
Method Article
マウス胚中足骨の文化:軟骨内骨化の生理学的モデル
要約
We present a protocol to dissect and culture embryonic day 15 (E15) murine metatarsal bones. This highly physiological ex vivo model of endochondral ossification provides conditions closer to the in vivo situation than cells in monolayer or 3D culture and is a vital tool for investigating bone growth and development.
要約
The fundamental process of endochondral ossification is under tight regulation in the healthy individual so as to prevent disturbed development and/or longitudinal bone growth. As such, it is imperative that we further our understanding of the underpinning molecular mechanisms involved in such disorders so as to provide advances towards human and animal patient benefit. The mouse metatarsal organ explant culture is a highly physiological ex vivo model for studying endochondral ossification and bone growth as the growth rate of the bones in culture mimic that observed in vivo. Uniquely, the metatarsal organ culture allows the examination of chondrocytes in different phases of chondrogenesis and maintains cell-cell and cell-matrix interactions, therefore providing conditions closer to the in vivo situation than cells in monolayer or 3D culture. This protocol describes in detail the intricate dissection of embryonic metatarsals from the hind limb of E15 murine embryos and the subsequent analyses that can be performed in order to examine endochondral ossification and longitudinal bone growth.
概要
スケルトンは、イオン恒常性に、歩行及ぶ機能の範囲を持っている非常に、複雑な動的で複雑な器官です。骨および軟骨からなる、骨格が、それは、構造的、生化学的および機械的完全性1です維持するために動作する機能的に異なる細胞集団で構成されています。スケルトンの主な機能の1つは、発展と成長におけるその役割です。これらのプロセスは、タイトな内分泌および遺伝的制御を介して調節されている複数の細胞集団のオーケストレーションを必要とします。
平らな骨、例えば肩甲骨、頭蓋及び胸骨を除いて、哺乳動物の骨格が軟骨内骨化と呼ばれるプロセスを介して形成されています。規制の両方の分子的および空間的にこのプロセスは、中胚葉2から主に由来間葉系細胞の縮合から始まります。転写因子SOX9の影響下で、細胞を目で縮合の電子内部は発現していると、そのようなII型コラーゲンおよびアグリカンなどの軟骨特定の細胞外マトリックス(ECM)成分を分泌する、軟骨細胞に分化します。結露のマージンの細胞を、Iコラーゲンと将来の骨3の輪郭を描く、軟骨膜を形成急行タイプ。その後、軟骨膜の骨幹は、転写因子、Runx2のとオステリックス4の発現が監督、骨芽細胞に分化します。これは、骨芽細胞の優位性をもたらし、この層は、骨の中間部周りの鉱化骨の襟を形成骨膜という名前に変更されます。軟骨足場の骨膜および浸潤の血管の浸透は、それに軟骨細胞の残骸や軟骨基質5の多くを再吸収haematopoetically由来の骨吸収細胞、破骨細胞を、持参発生します。形成された空間は、一次骨化を生じさせる骨幹によって充填されています次第に骨幹の中核を占め、骨膜と合流センター。他の細胞は骨髄を生じさせます。出産の周囲には、血管および骨幹細胞は、二次骨化中心を形成するために開発骨幹のいずれかの側(骨端)で軟骨が豊富なマトリックスを貫通しています。骨端成長板- -すべての長骨6の線形成長を調整する責任を負う長骨の両端に骨端と骨幹の間に位置する軟骨のバンドです。
成長板内の軟骨細胞は、最初に増殖し、その後、転写および成長因子の連続式でコーディネート形態学的に異なるゾーンを経て進行します。この一連のイベントの集大成がこの軟骨前駆体の中心で軟骨細胞の細胞周期と肥大からの出口です。肥大軟骨細胞を強く速達X型コラーゲンおよびマトリックスメタロプロテアーゼ-13(MMP13)と縦成長の方向のかなりの量の拡大は、哺乳動物7,8における骨の成長への最大の貢献を提供します。また、肥大軟骨細胞はマトリックス小胞の放出を介して、その周囲のECMの石灰化、膜構造は、それらのホスファターゼの包含(組織非特異的アルカリホスファターゼ(TNAP)とPHOSPHO1)を介して、非晶質リン酸カルシウムの製造に特化境界カルシウムタンパク質チャネリング(アネキシン)9-11。この石灰化軟骨は、破骨細胞の動員およびマトリックス再吸収の結果、基礎となる骨髄から血管に侵略されています。これは、彼らが急速に鉱化ある類骨の層を敷設石灰化軟骨の残党の表面に骨芽細胞の遊走とアライメントが続いています。一次海綿の網状骨は、後に二次海綿質12の層板骨に改造されています。中骨端核融合の結果軟骨内骨化13の停止。
乱れ発生および/または長手方向の骨の成長14を防止するために軟骨内骨化は、多くの内分泌および傍分泌ホルモンおよび成長因子による厳しい規制の下にあります。これらのプロセスを支える分子・細胞メカニズムのより良い理解は、人間の利益に向けて臨床の進歩の追求で、したがって必須です。異なる年齢、場所や種の成長板との間の類似点と相違点へのこれまでの研究では、大幅に成長板のダイナミクスを15,16の周囲の生理学的メカニズムの理解を改善しています。その環境からの軟骨細胞の除去は、COL2A1 17などの主要なマーカーの発現の消失を伴う、脱分化につながることができますようにしかし、単離された軟骨細胞の研究は、困難です。
マウスの中足骨、臓器の移植片培養、パイオランダの教授エリザベートバーガーらがoneered、 生体内 18 に見られる文化の模倣で骨の成長率として軟骨内骨化し、骨の成長を研究するための非常に生理のex vivoモデルです。ユニーク、中足骨器官培養は、軟骨形成の異なる段階で軟骨細胞の検査を可能にするため、文化19-21における細胞よりもin vivoの状況に近い条件を提供する、細胞-細胞および細胞-マトリックス相互作用を維持します。また、このモデルは、一次軟骨細胞培養は不可能である線形骨成長の直接的検査を可能にします。また、そのため、成長板のローカル効果の具体的な分析を可能に全身および局所因子の分離を可能にします。
中足骨の文化は、多数のパラメータの調査を可能にします。全長のとの鉱化地域の毎日の測定値基礎は、標準的な位相差顕微鏡及び画像解析ソフトウェアを用いて行うことができます。 in situハイブリダイゼーションおよび免疫組織染色で組織学は、容易に両方の細胞および分子実体の空間分解能、従来の細胞培養方法に比べて大きな利点を可能にする、中足骨切片上で実施してもよいです。免疫組織化学的手法は、軟骨細胞22を増殖することにより5-ブロモ-2'-デオキシウリジン(BrdU)の取り込みの検出および定量を含んでいます。また、中足骨組織のさらなる処理は、mRNA発現(RT-qPCRの)、タンパク質および細胞内シグナル伝達の解析(ウエスタンブロット)、または脂質組成(質量分析)の下流の分析を可能にするために行われてもよいです。これまでのほとんどの研究は、出生後の動物から培養した胚性中足骨ではなく、中足骨を使用します。両方のモデルが有益であることができる一方で、胚の骨の研究は、いくつかの利点があります:彼らは速く成長し、exploすることができます鉱化プロセス23-25の開始および進行を研究するited。
このプロトコルは、詳細に胚15日目(E15)は、マウス胚の後肢から胚中足骨の複雑な解剖を説明します。また、解剖学的に別個の中足骨の成長と鉱化作用が示されています。
プロトコル
動物を対象とする手順は、英国の動物(科学的手順)に準拠法1986年に行われた、動物をホームオフィスに従って維持した科学的な目的のために、ブレッド付属または使用される動物の住宅とケアのための行動規範を発表しました。
注:野生型C57BL / 6Jマウスを使用して行った実験は、十分に確立され、以下に記載されています。彼らの成長と石灰化率は、本明細書に詳述したものに異なる場合がありますが、異なるマウス系統からの胚中足骨は、同様に、in vitroで培養することができます。
1.解剖条件と楽器の調製
- メディアや胚性原基の無菌性を確保するために、層流フード内のすべてのメディアの準備、組織の解剖と文化の作業を実行します。
- 文化や解剖メディアは使用前に37℃、5%CO 2インキュベーター内で少なくとも1時間平衡化することができます。
- 解剖を殺菌オートクレーブ処理によって超Vannasマイクロハサミと一緒にはさみ、デュモン#5とデュモン#4ピンセット(8センチメートルストレート、3ミリメートルブレード)。オートクレーブ処理後、70%エタノール中で使用する前に切開装置を浸します。
解剖と培養培地の調製
- 解剖媒体を準備します
- リン酸緩衝生理食塩水(PBS)(1:13)でα最小必須培地(ヌクレオシドなし、αMEM)で希釈し、フィルターを0.22ミクロン、直径33mmシリンジを介して滅菌前に2mg / m Lにウシ血清アルブミン(BSA)に再懸濁フィルタ。
注:解剖媒体を作製することができ、フィルター滅菌し、無期限に-20℃でアリコートで保存しました。
- リン酸緩衝生理食塩水(PBS)(1:13)でα最小必須培地(ヌクレオシドなし、αMEM)で希釈し、フィルターを0.22ミクロン、直径33mmシリンジを介して滅菌前に2mg / m Lにウシ血清アルブミン(BSA)に再懸濁フィルタ。
- 培養液を準備します
- (24ウェル培養プレートの各ウェル中の)培地300μlの中で培養胚の中足骨。 BSA w / vの0.2%を添加することにより培養培地を準備し; 5μg/ mLのL-ASCOrbicアシッドホスフェート; 1 mMのβグリセロリン酸(βGP;オプション - 結果の項を参照してください)。 0.05 mg / mlでのゲンタマイシンおよびαMEMに1.25μgのアムホテリシンBおよびフィルタは0.22μmで、33ミリメートルの直径のシリンジフィルターを通して滅菌します。
3.マウス胚中足骨の解剖と文化
- 英国のホームオフィスガイドラインに従って頸椎脱臼により、15日齢の胚を保有する、妊娠マウスをカリング
- 動物仰臥位を置き、70%エタノールを噴霧することにより、皮膚を消毒します。解剖ハサミを使用すると、皮膚と腹腔を露出させるために腹膜の両方を貫通し、正中線に大きな切開を行います。
- 体腔の背部に動物の2子宮角を見つけます。まず解剖ハサミで慎重に切開によって子宮間膜から各子宮を解放した後、最終的にはそれらのベースで自由子宮角を切断することにより子宮角を削除します。 Plac解剖媒体への電子の子宮角。
- 子宮角に沿って移植部位との間で切断することによって、各胚を分離し、鉗子を使用して、個々の嚢から胚を削除します。英国のホームオフィスガイドラインに従って断頭により胚をカル
- 70%エタノールを噴霧して、皮膚の消毒した後、近位大腿骨の周りに切開することにより胚からの後肢を除去するためにVannasマイクロはさみを使用しています。削除される可能性としてこれは後肢の限り保証します。中足骨の解剖の前に解剖媒体に沈めペトリ皿に後肢を置きます。
- 解剖顕微鏡下で、#4ピンセットを用いて、安定した後肢を保持しながら、#5ピンセットを使用してピンチとそれを引っ張ることによって、胚の足の周囲の皮膚を除去し始めます。これは、最も効果的に脛骨のレベルのまわりで皮膚をつかみ、指骨に向かって引っ張ることによって行われます。カットするには、この段階でのはさみの使用非常に薄い皮膚は必要ありません。
- 慎重#4ピンセットで胚足のtarsalsを保持し、#5ピンセットを使用して、廃棄tarsals近くにピンチオフすることにより、第1及び第五中足骨を取り除きます。
- 同じ位置に保持足で、残りの3つの指骨と中足骨の間の結合組織を破壊するために#5ピンセットを使用します。中足骨から指骨を除去するために、指骨と中足骨の間の関節で#5ピンセットの先端を挿入します。
- 最後に、tarsalsと中足骨の間の結合組織を破壊するために#5ピンセットを使用します。再びtarsalsからtarsalsと中足骨、穏やか無料中足骨の間の関節空間に#5ピンセットの先端を配置します。
- 静かに新鮮な解剖培地中の#4ピンセットと場所で今すぐ無償中足骨を拾います。
注:慎重に解剖を各胚から6中足骨を提供する必要があります。 - 必要なすべての中足骨が解剖された場合には、carefu培養培地の300μLを含む予め温めた24ウェルプレートに個別にLLY場所の中足骨。最大14日間、5%CO 2インキュベーター内で37℃で培養中足骨。
注:これは彼らの鉱化能力26に影響を与えるとして、文化の少なくとも5日目まではE15培養のメディアを変更しないでください。この理由は不明であるが、線形成長およびマトリックスの石灰化は、中足骨から放出された成長因子による刺激に依存し得ます。
4.分析と中足骨の文化の操作
- 解剖(0日目)の日にデジタル取り付けられたカメラと画像解析ソフトウェアを用いて、顕微鏡を使用して最初の長手方向の測定を行います。それは文化の数日後に表示されたときに、中央鉱化帯の長さを測定します。
- 必要に応じて定期的に、その点mでの培養の最後の日まで(通常は毎第2日)、さらに測定を行いますetatarsal骨が必要な転帰(導入に説明する)に依存して処理されます。
- 文化22,24の0日目から増殖因子、ホルモンや薬理学的薬剤を、例えば 、様々な外因性因子と中足骨器官培養のサプリメント媒体。この種のすべての研究では、コントロールの未処理中足骨が必要とされています。
注:それは制御骨が対側脚から同等の原基であることをお勧めしますが、(下記図2参照)で培養E15、2位、3 位と4 番目の中足骨の任意の線形成長の違いや鉱化作用の可能性がされていません。
結果
この方法の目的は、縦骨の成長とECMの石灰化の検査のために胚の中足骨や文化、それらを単離することでした。光学顕微鏡により可視化されるように、本明細書に記載の我々のプロトコルを使用して、胚の中足骨が成功し、解剖しました。 図1に示すように、実施中足骨の測定値は、そのECMの長手方向の長さに成長し、石灰化する能力を示した( 図2および 3)。マトリックスの鉱化作用は、第1の骨原基の半ば骨幹で指摘され、簡単に光学顕微鏡で観察しました。カルセインの取り込みが新たに形成された鉱物の強化された可視化を提供することがありますが、無染色は必要ありません。
日付22,27,28に胚の中足骨を利用した研究は、解剖、2位、3 位と4 番目 (CENをプールされていますクトル3)中足骨は、 インビトロでの成長と鉱化作用に仮定すると匹敵する。ネズミ中足骨のインビボ発達は、しかし、3 番目と4 番目の数字は前に他のすべての中足骨と指骨29への鉱化作用の証拠を持っていることが明らかになりました。個別に単離し、培養E15、2位、3 位と4 番目の中足骨の成長と鉱化作用の可能性の評価は、培養7日目( 図2)の後に石灰化の外観や程度に有意差がないことが明らかになりました。ベースラインからの増加率には差は培養期間中に見つかりませんでした。鉱化作用の可能性には差が認められなかったので2 回目 3 回目と4 回目の中足骨をプールの標準プロトコルは、将来の研究のための有効な表示されます。
伝統的に、研究者らはメディアにβGPを追加しましたECMの石灰化を促進するために、培養胚の中足骨を使用。しかしながら、細胞培養の研究では、TNAP酵素がβGPを含有する骨形成培地に添加された場合、異所性ヒドロキシアパタイトミネラル沈着がまだもセル30の非存在下で起こることが示されています。中足骨の石灰化能力に様々な石灰化基質とアゴニストの効果を調べるために、我々は、サプリメントの範囲を含む培養培地中でE15の中足骨を培養し、画像や長さの測定を毎日記録しました。結果は、E15中足骨がまだ成長しβGPの不在下で彼らのECMを石灰化することを示しました。中足骨がβGP( 図3)の存在下で培養としてアスコルビン酸で培養E15中足骨が唯一のメディアは、長さと石灰化における同等の増加を示しました。
我々はまた、レンチウイルスを培養metataに外因性DNAをトランスフェクトするために使用され得ることを示していますrsals。ここでは、正常に中足骨あたり2×10 6緑色蛍光タンパク質(GFP)ウイルス粒子を含む培養(解剖のすなわち日)の0日目に中足骨をトランスフェクトしました。 500:ウイルス形質導入を可能にするために、ポリブレンは同時に1で培養物に添加しました。培養物を4とインキュベートする前に、発生するE15中足骨石灰化に必要な「ノーメディア変更を、7日間、5分間、6-ジアミジノ-2-フェニルインドール(DAPI)染色を出て、直接共焦点顕微鏡を用いて可視化しました( 図4)。コンセプト実験の私達の証明は明らかに効果的な伝達を示しています。しかし、彼らのトランスフェクションし、効果的な遺伝子操作を評価するために、中足骨の全体を通してセクションを検討するのが賢明だろう。

図1: 標準的な方法は、Tを測定するために使用彼の長さと胚中足骨の鉱化ゾーン。(A)の培養0日目E15の中足骨の合計の長さの測定(解剖の日)は、中足骨の中心を通って撮影しました。 (B)培養液中で7日後の中足骨の総長手方向の長さの測定。中足骨のわずかな曲率に注意してください。 (C)文化の中で7日後の鉱化帯の長さの測定。 この図の拡大版をご覧になるにはこちらをクリックしてください。
{kind=link}

図2: 成長と解剖学的に別個のE15中足骨の石灰化能力第二のイメージングと計測、ワットE15マウスの後肢から3番目と4番目の中足骨。実行されます。 (A)培養期間を通じて第二、第三及び第四中足骨の連続画像。文化の日ごとに0日目からの長さは(B)の割合が増加。培養期間にわたって第二、第三及び第四中足骨の(C)ミネラルの長さは合計中足骨の長さの割合として表しました。データは、平均±平均の標準誤差(SEM)として表現された(n = 6)。 (D)文化の各日におけるC57BL / 6J中足骨の長さ。中のデータは平均±標準偏差として表された(n = 6)されています。ブラックバー= 1mmである。 この図の拡大版をご覧になるにはこちらをクリックしてください。
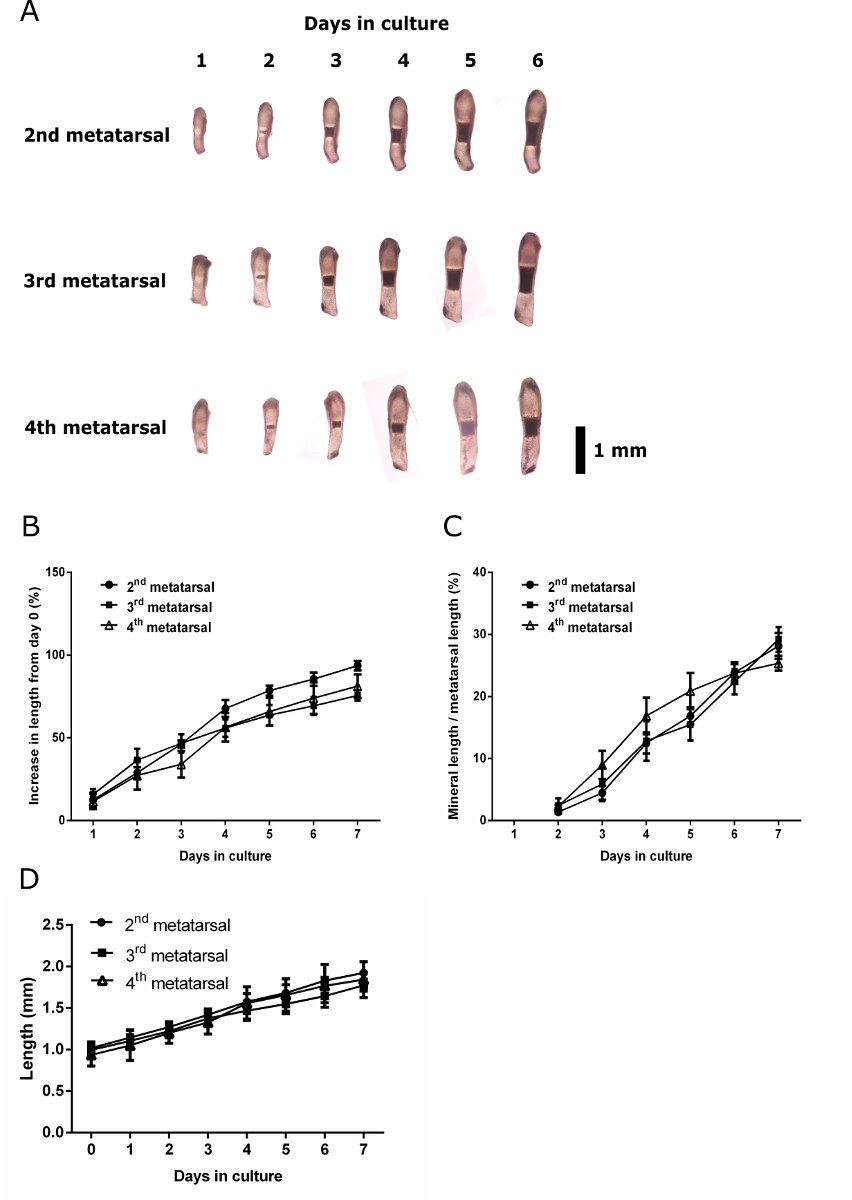{kind=link}

図3: 野生型の成長と鉱化容量様々な骨形成メディアでE15中足骨培養。(A)のみアスコルビン酸、1mMのβGP、3 mMのホスホコリ、1.5 mMの塩化カルシウムやメディアを含む3 mMの塩化カルシウムで培養中足骨の連続画像。中足骨の0日目からの長さは(B)の割合の増加は、様々なサプリメントで培養しました。様々な培地で培養中足骨の(C)ミネラルの長さは合計中足骨の長さのパーセンテージとして表しました。データは、として平均±平均の標準誤差(SEM)で表される(N = 6)。ブラックバー= 1mmである。 この図の拡大版をご覧になるにはこちらをクリックしてください。
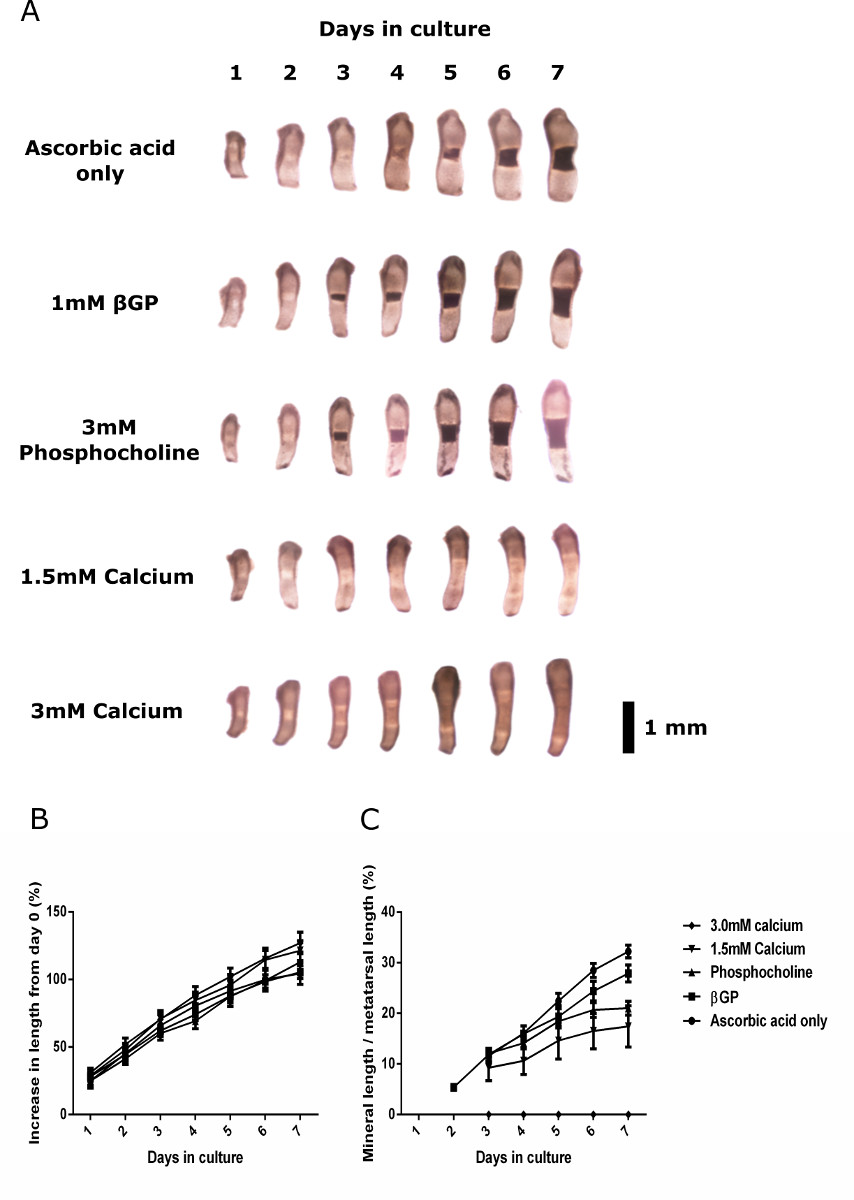{kind=link}

図4:GFP ウイルスPとE15中足骨のトランスフェクション物品(A)E15中足骨がコンセプトの証明のためのGFPのウイルス粒子でトランスフェクトしました。 (B)骨は、DNAの可視化のためのDAPIで染色しました。 (C)二重の画像が中足骨の全体にわたってGFPウイルスの成功したトランスフェクションを示します。白いバー= 0.5ミリメートル。 この図の拡大版をご覧になるにはこちらをクリックしてください。
{kind=link}
ディスカッション
マウス胚中足骨の文化は、軟骨内成長と鉱化作用の高度生理学的モデルを提供します。初期の研究では、E15マウス中足骨は骨格の成長および分化の正常なパターンを受けることが確認されました。軟骨(軟骨細胞)と骨(骨芽細胞と破骨細胞)細胞およびそれらのそれぞれのコラーゲン性マトリックスは、in vivoで観察されたものと区別がつきません。しかし、このモデルのいくつかの認識限界があります。破骨細胞分化の速度は、骨の成長(しかし分化軟骨ではない)であるとして損なわれる。骨の成長は、in vivoで観察されたものよりも約50%遅く、局所因子( 例えば 、IGF-1、BMPの、 など )31の生産に影響を与える全身要因の不備に起因する可能性があります。また、十分に骨がその機械的環境に敏感であることが認識され、これは、Cに応答培養中足骨、の場合であります機械的負荷32,33でhanges。このプロトコルで説明したようにそのため、無負荷の状況では、鉱化作用、ミネラル吸収が損なわれる可能性があります。それにもかかわらず、中足骨のモデルは、直接インビトロ培養系で 2Dと3Dを介して可能でない線状の骨の成長を測定する能力を提供しています。
我々は、E15マウス中足骨の解剖と文化に関わるプロトコルの包括的な説明を提供し、実際に、初めて、実験のための第2、 第3および第 4中足骨をプールの有効性を確認しています。
E17 / E18から中足骨は、一般的に(培養14日以上IE)長期間のex vivoで増殖するそれらの異常な電位による骨成長のメカニズムを調べるために使用されます。それらは、胚の縦骨の成長の成長因子の役割を調査するために、十分に確立されたモデルです。さらに、出生後の中足骨の解剖培養は、一般的に、出生後の骨の成長と胎児の骨成長が異なっ21調節されることが理解されているように、出生後の骨の成長を取り囲むメカニズムを描写するために使用されます。培養生後中足骨で観察された骨の成長は長期試験で自分の可能性を制限し、この出生後の段階で、骨の成長の詳細全身影響を強調し、しかし、胚の基礎25,34で観察されたものよりも著しく小さいです。実際、マウスから3日目の生後中足骨は、一般的に測定可能な成長34を得るために使用することができ、最新のステージです。
ここでは、E15での胚の中足骨の単離のために私たちのプロトコルを記述します。 E15中足骨のヒドロキシアパタイト鉱物が存在しないことは、それらが縦骨の成長を取り囲むメカニズムも開始しないだけを調べるために無類のモデルを提供することを意味します骨格の石灰化。末端肥大軟骨細胞ゾーンの鉱化マトリックスは、その後石灰化している骨の特定のマトリックス(類骨)を敷設する骨芽細胞に侵入するための足場を提供し、そのようなこの最初の鉱化作用として成功し、機能的な骨形成35のために不可欠です。実際にE15の中足骨を利用する最近の報告数は、石灰化プロセス28,36,37の調査のための彼らのユニークな能力を確認しました。また、研究者らは、骨の成長/石灰化の設定外のモデルとして胚中足骨の可能性を強調表示し、血管新生38のモデルとして、E15の中足骨の使用が記載されています。
私たちが説明するプロトコルは、層流フード、解剖を行うための解剖顕微鏡、および切除した原基の培養のためのCO 2インキュベーターを含むのみ標準的な実験装置を必要とします。また、非常に基本的なCULαMEM、BSA、抗生物質、抗真菌剤、およびL-アスコルビン酸(ヒドロキシプロリンおよびヒドロキシリジンの合成における補因子、コラーゲン39の製造のための2つの必須アミノ酸)を含むチャー培地は、動物血清のような未定義の添加剤の使用を回避します。伝統的に、研究者らは、培養胚の中足骨に使用されるメディアにβGPを追加したが、我々の結果で明らかにされるように、追加のリン酸源の使用は、この培養系で成功石灰化のために必要とされていません。これは、ECMの石灰化を支えるメカニズムを理解することの追求に重要な知見です。
中足骨は以前、長骨の発達40の調節にトランスフォーミング成長因子βの役割を探求するのSmad2のドミナントネガティブ型を含むアデノウイルスに感染しています。ここでは、またトンを示す、GFPウイルス粒子と野生型E15中足骨のトランスフェクションが成功を明らかにする帽子レンチウイルス技術は、容易に足骨培養における遺伝子発現を操作するために採用することができます。同様に、中足骨器官培養系は、より良好な骨成長過程における特定の遺伝子の役割を理解するために遺伝的に改変されたマウスからの骨の成長を比較するには優れたモデルです。我々の以前の研究では、軟骨内骨成長におけるサイトカインシグナル-2(SOCS2)の抑制の役割を決定するために、中足骨器官培養系を利用しています。 SOCS2欠損マウスから中足骨を使用して、我々は、成長ホルモンは、成長ホルモン治療34に応答しない野生型の中足骨とは異なり、成長因子(IGF-1)のようなインスリンのその長手成長独立してシミュレートすることが可能であることを示しています、 41。これは、中足骨の培養物は、異なる遺伝子および/または軟骨内骨成長の外因性因子の影響を調べるために操作可能な範囲を強調する。
要約すると、我々はdetaiを持っています操作および分析のさまざまな方法を使って調べることができる胚の中足骨の抽出成功と文化のための方法を導きました。このex vivoでのモデルは、従って、単層又は三次元培養物中の細胞よりも多くの生理学的モデルを提供すること、ならびに軟骨形成の異なる段階における軟骨細胞が含まれて、細胞-細胞及び細胞-マトリックス相互作用を維持します。中足骨器官培養は、したがって、軟骨内骨化を担当する分子機構を調べるためのユニークなモデルであり、骨の生理学と病態生理の両方の我々の理解を促進するために必須です
開示事項
The authors have nothing to disclose.
謝辞
We are grateful to the Biotechnology and Biological Sciences Research Council (BBSRC) for funding DH (BB/J01446X/1), CF & VEM (Institute Strategic Programme Grant Funding BB/J004316/1). We also acknowledge Arthritis Research UK for funding to KAS (20413). We thank Miss Elaine Seawright for her assistance with the experiments detailed, Dr. Neil Mackenzie for his assistance with the lentiviral techniques, and Mr Darren Smith and Mr Alex Robertson (Roslin Institute BRF) for their assistance with the animal studies.
資料
| Name | Company | Catalog Number | Comments |
| Dissection scissors | World Precision Instruments | 14393 | Autoclave sterilise before use |
| Dumont #5 tweezers | World Precision Instruments | 500342 | Autoclave sterilise before use |
| Dumont #4 tweezers | World Precision Instruments | 500339 | Autoclave sterilise before use |
| SuperFine Vannas scissors | World Precision Instruments | 501778 | |
| Absolute ethanol | Fisher Scientific | E/0650DF/17 | Dilute to 70% v/v in dH2O |
| α-MEM without nucleosides | Thermo Fischer Scientific | 22561021 | |
| Bovine serum albumin | Sigma Aldrich | A3059 | |
| Syringe filters (0.22 μm) 33 mm diameter | Merck Millipore | SLGP033RS | |
| Phosphocholine chloride calcium salt tetrahydrate | Sigma Aldrich | P0378 | Add directly to the media prior to experimentation |
| L-ascorbic acid-2-phosphate | Sigma Aldrich | A8960 | Make up stock soution at 5mg/ml and aliquot at -20 °C |
| Calcium chloride dihydrate | Sigma Aldrich | C5080 | Make up stock soution at 150mM and aliquot at -20 °C |
| β-glycerophosphate disodium salt hydrate | Sigma Aldrich | G9422 | Make up stock soution at 1mM and aliquot at -20 °C |
| Gentamicin | Thermo Fischer Scientific | 15710049 | |
| Amphotericin B | Thermo Fischer Scientific | 15290018 | |
| Nunc cell-culture treated 24-well plates | Thermo Fischer Scientific | 142475 | |
| Polybrene | Sigma Aldrich | H9268 | |
| DAPI | Thermo Fischer Scientific | D3571 |
参考文献
- Farquharson, C., Staines, K. The skeleton: no bones about it. J. Endocrinol. 211, 107-108 (2011).
- Berendsen, A. D., Olsen, B. R. Bone development. Bone. 80, 14-18 (2015).
- Kronenberg, H. M. Developmental regulation of the growth plate. Nature. 423, 332-336 (2003).
- Komori, T. Regulation of bone development and extracellular matrix protein genes by RUNX2. Cell Tissue Res. 339, 189-195 (2010).
- de Crombrugghe, B., Lefebvre, W. Regulatory mechanisms in the pathways of cartilage and bone formation. Curr. Opin. Cell Biol. 13, 721-727 (2001).
- Hall, B. K. Bones and Cartilage: Developmental and Evolutionary Skeletal Biology. , Elsevier Academic Press. (2005).
- Farquharson, C., Jefferies, D. Chondrocytes and longitudinal bone growth: The development of tibial dyschondroplasia. Poult. Sci. 79, 994-1004 (2000).
- Cooper, K. L., et al. Multiple phases of chondrocyte enlargement underlie differences in skeletal proportions. Nature. 495, 375-378 (2013).
- Wuthier, R. E., Lipscomb, G. F. Matrix vesicles: structure, composition, formation and function in calcification. Front. Biosci. 16, 2812(2011).
- Roberts, S., Narisawa, S., Harmey, D., Millan, J. L., Farquharson, C. Functional involvement of PHOSPHO1 in matrix vesicle-mediated skeletal. J. Bone Miner. Res. 22, 617-627 (2007).
- Cui, L., Houston, D. A., Farquharson, C., MacRae, V. E. Characterisation of matrix vesicles in skeletal and soft tissue mineralisation. Bone. 87, 147-158 (2016).
- Brighton, C. T. Structure and Function of Growth Plate. Clin. Orthop. , 22-32 (1978).
- Shim, K. S. Pubertal growth and epiphyseal fusion. Annals of pediatric endocrinology & metabolism. 20, 8-12 (2015).
- Staines, K. A., Pollard, A. S., McGonnell, I. M., Farquharson, C., Pitsillides, A. A. Cartilage to bone transitions in health and disease. J. Endocrinol. 219, R1-R12 (2013).
- Hunziker, E. B., Schenk, R. K., Cruzorive, L. M. Quantitation of Chondrocyte Performance in Growth-Plate Cartilage During Longitudinal Bone-Growth. Journal of Bone and Joint Surgery-American. 69A, 162-173 (1987).
- Breur, G. J., Vanenkevort, B. A., Farnum, C. E., Wilsman, N. J. Linear Relationship Between The Volume Of Hypertrophic Chondrocytes And The Rate Of Longitudinal Bone-Growth In Growth Plates. J. Orthop. Res. 9, 348-359 (1991).
- Ma, B., et al. Gene expression profiling of dedifferentiated human articular chondrocytes in monolayer culture. Osteoarthritis Cartilage. 21, 599-603 (2013).
- Thesingh, C. W., Burger, E. H. The role of mesenchyme in embryonic long bones as early deposition site for osteoclast progenitor cells. Dev. Biol. 95, 429-438 (1983).
- Scheven, B. A., Hamilton, N. J. Longitudinal bone growth in vitro: effects of insulin-like growth factor I and growth hormone. Acta endocrinologica. 124, 602-607 (1991).
- Coxam, V., Miller, M. A., Bowman, M. B., Miller, S. C. Ontogenesis of IGF regulation of longitudinal bone growth in rat metatarsal rudiments cultured in serum-free medium. Archives of physiology and biochemistry. 104, 173-179 (1996).
- Andrade, A. C., Chrysis, D., Audi, L., Nilsson, O. Methods to study cartilage and bone development. Endocr Dev. 21, 52-66 (2011).
- Mushtaq, T., Bijman, P., Ahmed, S. F., Farquharson, C. Insulin-like growth factor-I augments chondrocyte hypertrophy and reverses glucocorticoid-mediated growth retardation in fetal mice metatarsal cultures. Endocrinology. 145, 2478-2486 (2004).
- Owen, H. C., Miner, J. N., Ahmed, S. F., Farquharson, C. The growth plate sparing effects of the selective glucocorticoid receptor modulator, AL-438. Mol. Cell. Endocrinol. 264, 164-170 (2007).
- MacRae, V. E., Farquharson, C., Ahmed, S. F. The restricted potential for recovery of growth plate chondrogenesis and longitudinal bone growth following exposure to pro-inflammatory cytokines. J. Endocrinol. 189, 319-328 (2006).
- Chagin, A. S., Karimian, E., Sundstrom, K., Eriksson, E., Savendahl, L. Catch-up growth after dexamethasone withdrawal occurs in cultured postnatal rat metatarsal bones. J. Endocrinol. 204, 21-29 (2010).
- Haaijman, A., et al. Inhibition of terminal chondrocyte differentiation by bone morphogenetic protein 7 (OP-1) in vitro depends on the periarticular region but is independent of parathyroid hormone-related peptide. Bone. 25, 397-404 (1999).
- Haaijman, A., Dsouza, R. N., Bronckers, A., Goei, S. W., Burger, E. H. OP-1 (BMP-7) affects mRNA expression of type 1, II, X collagen, and matrix Gla protein in ossifying long bones in vitro. J. Bone Miner. Res. 12, 1815-1823 (1997).
- Staines, K. A., et al. MEPE is a novel regulator of growth plate cartilage mineralization. Bone. 51, 418-430 (2012).
- Kaufman, M. The atlas of mouse development. , 3, Academic Press. (1992).
- Boskey, A. L., Roy, R. Cell Culture Systems for Studies of Bone and Tooth Mineralization. Chem. Rev. 108, 4716-4733 (2008).
- Minkin, C., et al. Skeletal development and formation of osteoclast-like cells from in situ progenitors in fetal mouse metatarsals cultured in chemically defined. Bone Miner. 12, 141-155 (1991).
- Kleinnulend, J., Veldhuijzen, J. P., Burger, E. H. Increased calcification of growth plate cartilage as a result of compressive force in vitro. Arthritis Rheum. 29, 1002-1009 (1986).
- Kleinnulend, J., Veldhuijzen, J. P., Vanstrien, M. E., Dejong, M., Burger, E. H. Inhibition of osteoclastic bone-resorption by mechanical stimulation in vitro. Arthritis Rheum. 33, 66-72 (1990).
- Dobie, R., et al. Increased linear bone growth by GH in the absence of SOCS2 is independent of IGF-1. J. Cell. Physiol. 230, 2796-2806 (2015).
- Heilig, J., Paulsson, M., Zaucke, F. Insulin-like growth factor 1 receptor (IGF1R) signaling regulates osterix expression and cartilage matrix mineralization during endochondral ossification. Bone. 83, 48-57 (2016).
- Huesa, C., et al. The functional co-operativity of tissue-nonspecific alkaline phosphatase (TNAP) and PHOSPHO1 during initiation of skeletal mineralization. Biochemistry and Biophysics Reports. 4, 196-201 (2015).
- Staines, K. A., et al. Endochondral Growth Defect and Deployment of Transient Chondrocyte Behaviors Underlie Osteoarthritis Onset in a Natural Murine Model. Arthritis & Rheumatology. 68, 880-891 (2016).
- Song, W., et al. The fetal mouse metatarsal bone explant as a model of angiogenesis. Nature Protocols. 10, 1459-1473 (2015).
- Libby, P., Aikawa, M. Vitamin C, collagen, and cracks in the plaque. Circulation. 105, 1396-1398 (2002).
- Alvarez, J., Serra, R. Unique and redundant roles of Smad3 in TGF-beta-mediated regulation of long bone development in organ culture. Dev. Dyn. 230, 685-699 (2004).
- Pass, C., MacRae, V. E., Huesa, C., Ahmed, S. F., Farquharson, C. SOCS2 is the critical regulator of GH action in murine growth plate chondrogenesis. J. Bone Miner. Res. 27, 1055-1066 (2012).
転載および許可
このJoVE論文のテキスト又は図を再利用するための許可を申請します
許可を申請さらに記事を探す
This article has been published
Video Coming Soon
Copyright © 2023 MyJoVE Corporation. All rights reserved