
Method Article
事項をカットする場所: 郭清と眼のランドマークからマウス網膜の空間的なオリエンテーションの解析ガイド
要約
このプロトコルは、正確かつ確実に東洋の解剖学的領域でマウス単離網膜に深い眼のランドマーク、s オプシン免疫組織化学、Retistruct、およびカスタム コードを使用するため包括的な郭清と分析のガイドを提供します。
要約
正確かつ確実にマウス単離網膜の空間的なオリエンテーションを識別する視覚神経科学、密度の解析や網膜のセルタイプのサイズ グラデーションなど、多くの研究の重要な方向性のチューニングの方向神経節細胞といくつかの網膜疾患における地形変性パターンの検討。ただし、多くの異なる眼郭清の方法を文献で報告し、マウス網膜における網膜の方向をラベルに使用するがあります。方向のような調査で使用される方法はしばしば見落とされて、報告しないどのように網膜の方向を決定を引き起こす可能性が不一致文献および混乱の研究の間でデータを比較する際。角膜のやけどなど表面的な眼のランドマークが使われませんが最近腹直筋、脈絡裂のオプシン グラデーションなど深いランドマークより信頼性が低いことが示されています。ここでは、正確に分析し、マウス単離網膜の空間的なオリエンテーションを文書化する深い眼のランドマークの使用のため包括的なガイドを提供します。また 2 つのオプシンの抗体の有効性を比較して s オプシン免疫組織化学のためのプロトコルが含まれています。S オプシン グラデーションによると網膜の方向は、Retistruct ソフトウェアによる網膜再建とカスタム コードの回転を必要とするため、私たちはこれらのプログラムの両方を使用するために必要な重要なステップを発表しました。全体的に、このプロトコルの目標は、最も実験的プロトコルに適応できる正確な網膜向きのメソッドの信頼性と再現性のあるセットを提供することです。この作品の包括的な目標は、将来研究のため網膜方向メソッドを標準化することです。
概要
網膜神経科学の重要なと時々 見落された側面は適切な方向と全体マウント網膜の分析記録の部屋に電気生理学や組織学的のスライドの網膜の方向かどうか。これは、哺乳類の視覚系の調査のための最も広く使用されているモデルは、現在マウスの網膜を含む調査のため特に重要です。マウス網膜空間的に均一ではないが、メラノプシン神経節細胞、過渡 OFF α 神経節細胞、錐体オプシン1,2 などの機能的に異なる網膜細胞型の密度とサイズ グラデーションの最近の発見を明らかに ,3,4,5。その結果、網膜の方向性を決めるための手法がありますセル型オプシン分布2,3,6を含む実験結果方向が方向性のチューニング神経節細胞7,8,9, と網膜変性症10、11,12,13,14 の地形パターン.実際には、報告しないどのように網膜の方向を報告を引き起こす可能性が不一致文献および混乱の研究の間でデータを比較する際。したがって、研究者がこのような研究の結果を正確に解釈できるように網膜の方向を識別する方法を報告することが重要です。
網膜の方向の眼球摘出1,3,12,15,16,17 前に背側、腹側、鼻または一時的な角膜を得点によって識別される一般的 ,18,19外眼筋6,7など目の深い解剖学的ランドマークを染色したり、によって脈絡膜裂20,21, またはまたはs オプシン グラデーション2,3。腹直筋は上直筋、下直筋、内側直筋、外側直筋、いずれかの添付ファイルを二等分深い緩和カットすることによって背側、腹側、鼻と一時的な網膜をそれぞれ識別するために使用できます。ただし、ほとんどの実験では、1 つの直筋を用いたことで網膜22の方向付けのため十分です。目の開発の残骸である脈絡裂は、目の後ろにかすかな水平線として表示できます。この線の両端は、鼻腔またはグローブ23時間極のいずれかで終了します。最後に、s オプシンの発現は非対称マウスの腹側網膜に分散し、s オプシン抗体は免疫組織化学実験1で腹側の網膜を明らかにする使用ことができます。
Stabioらによる最近の仕事22角膜火傷等表面的な眼のランドマークが解剖学的スペース、ヒューマン エラーと角膜のやけどを作る時間と内側を使用する場合に可変性の原因で網膜の方向付けのため信頼性の低いメソッドであることを示した基準点として canthi。対照的に、上直筋、脈絡裂のオプシン グラデーションなどの深いランドマークは網膜22の方向付けのためのより信頼性の高い、正確なランドマークに示されています。しかし、これらの解剖学的ランドマークの同定文献で詳細に説明されていないユニークな解剖の手順が必要です。したがって、このプロトコルの目標は、上直筋、脈絡裂および s オプシン グラデーションを使用してマウス網膜の空間的なオリエンテーションを正確に識別する方法に関する包括的なチュートリアルを提供することです。さらに、我々 は s オプシン免疫組織化学のためのプロトコルと同様に、2 つのオプシンの抗体の有効性の比較を含まれています。
正確な網膜の向きに依存する研究に追加課題の 1 つは、記録室、料理、またはスライドで wholemount 網膜を平らにするために必要な大規模な緩和カットです。これはフラット二次元構造物としてイメージが作成されたとき、何が当然立体構造の解析のための課題を導入できます。それから収集したデータを分析する前に、その立体構造にフラット wholemount 網膜に戻ります Retistruct24と呼ばれるプログラムを使用できます。したがって、このプロトコルのセクションは Retistruct ソフトウェアを使用して、s オプシン immunostained マウス網膜の再構築に必要な手順を強調する専用です。我々 はまた正確に回転とオリエントのマウス網膜のオプシンとステンド グラスを開発した当社のカスタム MATLAB スクリプトを使用するためのプロトコルのセクションを含まれています。
プロトコル
ここで説明したすべてのメソッドは、機関動物ケアおよび使用委員会 (IACUC) アクロンの大学によって承認されています。
1. 網膜の方向を識別するのに上直筋ランドマークを使用
注: 上直筋、背側網膜 (表 1) のランドマークです。実験は、背側網膜の印を必要としない、手順 1 と手順 2 に進みます。
- マウスの安楽死のため承認された機関動物ケアおよび使用委員会プロトコルに従ってください。
- 世界中の一般的な方向を識別するためにマーク直後に安楽死 (図 1A) 角膜強膜国境に近い鼻と時間の canthi の間で直接背側の角膜の火傷をします。火傷 10 秒間焼灼ペンを加熱し、1 秒未満の背側の角膜にペンの先端に触れることによってマークを確認します。
注: 保持のため角膜を焼灼ペン長すぎるパンク グローブになります。
注: いくつかの焼灼ペンは、光を発するか中、材料表に掲げる焼灼ペンは発光しないとき加熱、バクテリオロドプシンの実験のための安全なオプションとなっています。 - 眼球摘出、軽くソケットから目を押して、グリップの下からグローブにカーブタイプ鉗子を使用します。世界を削除する視神経を切断しないでください。代わりに、移動しながら同時に優しく左から右へ世界中がソケットから外れるまでにソケットから世界を持ち上げてゆっくりと。
注: この動きは世界中がソケットから完全に削除される最後に世界に接続されたままに腹直筋になります。視神経もよく世界中に接続されたままになります。 - 郭清の媒体を含んでいるペトリ皿で接続されている腹直筋とグローブを転送します。目は左目、右目を追跡することを確認します。
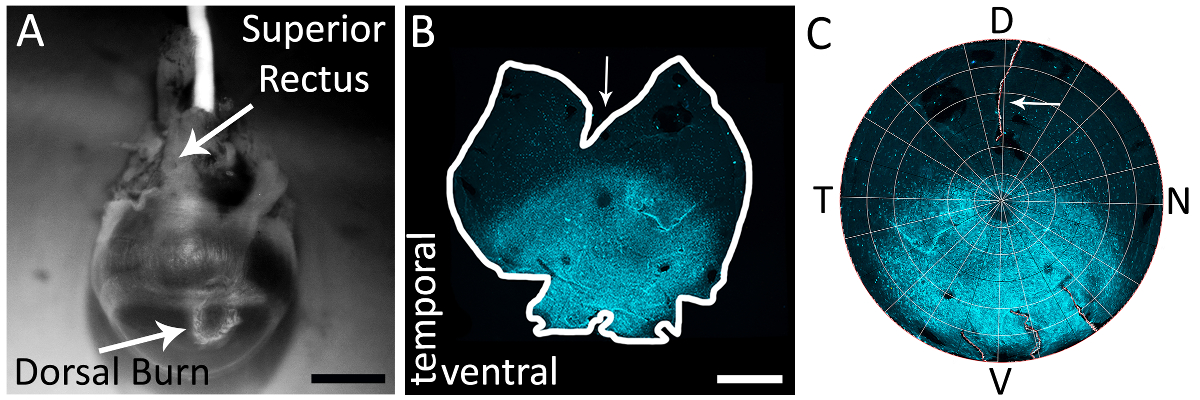
注: 切開は、彼らの実験的プロトコルと合わせて適切な解剖媒体を使用してください。 - 上直筋とを認識し、郭清範囲の下で視覚的に特定背側の角膜のやけどそれは関連付けられる (図 1A)。
- 解剖はさみを使ったり、20 G (0.9 mm × 25 mm) (材料の表を参照)、穿刺の針マークを燃やすで角膜。深い上筋を両断する視神経に向かって世界中にカットを緩和をします。このカットを分離し再構築した網膜は、図 1 bと1 Cに表示されます。
- 網膜の一部が公開されるまで手順 1.6 で穿刺した穴をそっと涙を鉗子 (材料の表を参照) の 2 つのセットを使用して網膜を分離を開始します。
メモ: 重要ですさらに涙を緩和のカットを引き起こすことができますこれが優しく、あまり強く引き裂くとして行われます。 - 強膜が完全に削除されるまで、離れて強膜から網膜をいじめるに鉗子を使用します。網膜が完全に分離するまでは、鉗子で虹彩、レンズ、硝子体、および任意の残りの構造を削除します。
注: プロトコルはここで一時停止可能性があります。組織は、s オプシン免疫組織化学的に修正するつもりですが場合、は、3.5 の手順に進みます。
2. 網膜の方向を識別するのに脈絡裂ランドマークを使用
注: 脈絡裂上の背中、目の強膜に存在は、(図 2 bおよび2 C鼻のポールに側頭極から実行表 1)。
- マウスの安楽死のための承認された機関動物ケアおよび使用委員会プロトコルに従ってください。
- 世界中の一般的な方向を識別するためにマーク直後に安楽死 (図 2A) 角膜強膜国境に近い鼻と時間の canthi の間で直接背側の角膜の火傷をします。火傷 10 秒間焼灼ペンを加熱し、1 秒未満の背側の角膜にペンの先端に触れることによってマークを確認します。
注: 保持のため角膜を焼灼ペン長すぎるパンク グローブになります。 - 目を摘出し、郭清媒体を含んでいるペトリ皿でグローブを転送します。目は左目、右目を追跡することを確認します。
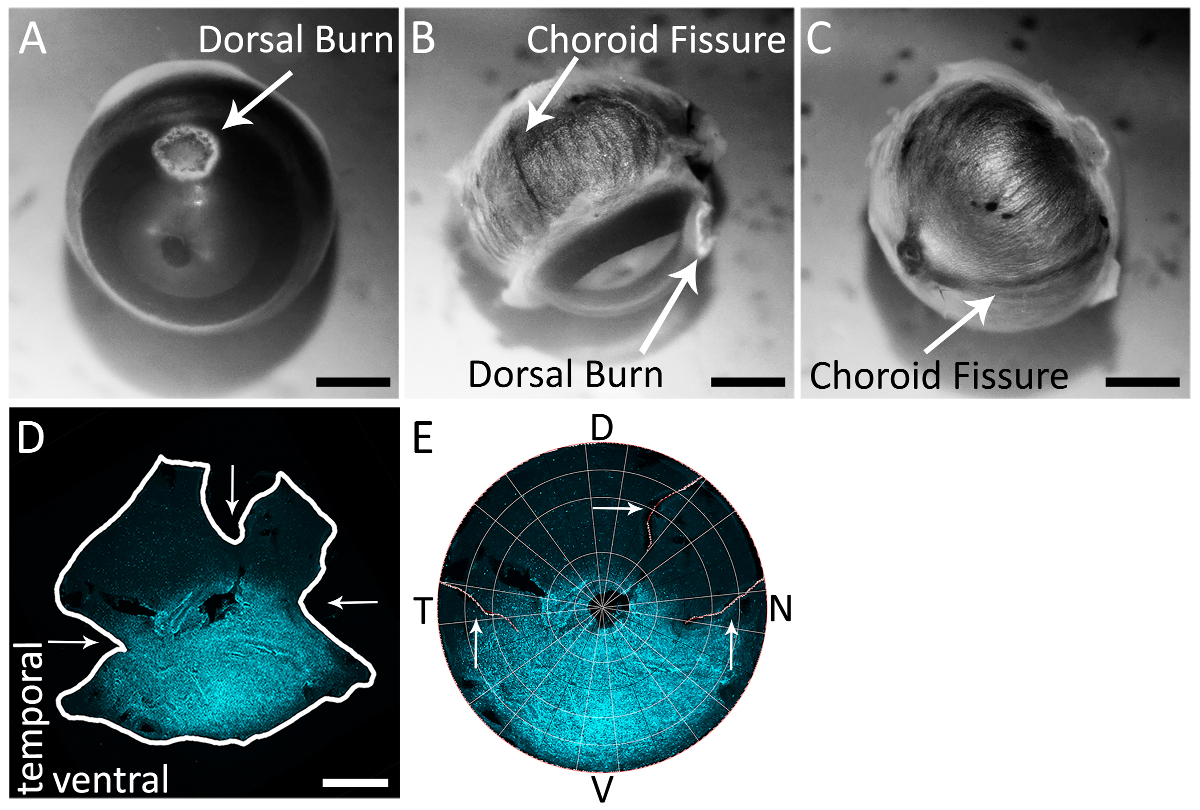
注: 切開は、彼らの実験的プロトコルと合わせて適切な解剖媒体を使用してください。 - 視覚的に検索して (図 2B, 2 C) 目の背面に脈絡裂を特定します。
注: 脈絡裂、赤外線光20下アイカップ内部表示されますも。 - 目がマウスでまだされたかどうか、背側の書き込みは優れた極に位置して、ペトリ皿のグローブに合わせます。
注意: 背の存在書き込みでグローブの鼻および時間的な面の識別は、かどうかそれは右または左の目がある限り記載されている: の場合右目、鼻の脈絡裂にやけどと、ロケーションの右側になりますl 脈絡裂に焼跡の左側になります。左目の場合は、一時的な脈絡裂に焼跡の右側になります、鼻の脈絡裂に焼跡の左側になります。
解剖はさみを使ったり、20 G (0.9 x 25 mm) は背の熱傷がある世界の中で (材料の表を参照) を 1 つ穿刺を針します。 - 作る浅い緩和視神経に対する背側の角膜の火傷があります。このカットは分離 (図 2D) 後の背側網膜の同定を可能にする、脈絡裂に垂直になります。
- 視神経への次の 2 つの深い緩和カット: 解剖のブレードを並べて 1 つは、目の後ろに一時的な脈絡裂線をはさみし、郭清のブレードを並べることによって 1 つは鼻の脈絡膜とをはさみ目の後ろの割れ目ライン。これらのカットは、図 2Dと2 eで分離・再生網膜に表示されます。
注: または、一時的な脈絡裂で深いカットを行うことができます、不要なカット背側の角膜のやけどを作る鼻の脈絡裂で浅いカットを行うことができます。これにより少ない緩和カットと網膜の正確な方向。 - 網膜の一部が公開されるまで手順 2.7 および 2.8 で穿刺した穴をそっと涙を鉗子 (材料の表を参照) の 2 つのセットを使用して網膜を分離を開始します。
メモ: 重要ですさらに涙を緩和の cut(s) を引き起こすことができますこれが優しく、あまり強く引き裂くとして行われます。 - 強膜が完全に削除されるまで、離れて強膜から網膜をいじめるに鉗子を使用します。網膜が完全に分離するまでは、鉗子で虹彩、レンズ、硝子体、および任意の残りの構造を削除します。
注: プロトコルはここで一時停止可能性があります。組織は、s オプシン免疫組織化学的に修正するつもりですが場合、は、3.5 の手順に進みます。
3. マウス網膜における S オプシン グラデーションのラベリング
メモ: s オプシン光式、腹側網膜に1網膜の腹側の半分の優れたマーカーとなって分散非対称的。このメソッドは、固定の役に立つだけですと immunostained 組織 (表 1)。次の手順は、上記の方法のいずれかを使用して解剖されている網膜に適用できます。
- マウスの安楽死のための承認された機関動物ケアおよび使用委員会プロトコルに従ってください。
- 安楽死、直後後目を摘出し、郭清培地でシャーレの世界を配置します。目は左目と右目は網膜の解剖後に網膜の方向を識別するために追跡することを確認します。
注: 切開は、彼らの実験的プロトコルと合わせて適切な解剖媒体を使用してください。 -
網膜の一部が公開されるまで、優しく角膜に穴を引き裂く鉗子 (材料表) の 2 つのセットを使用して網膜を分離を開始します。
注意: 重要です涙に網膜を引き起こすことができますこれが優しく、あまり強く引き裂くとして行われます。 - 強膜が完全に削除されるまで、離れて強膜から網膜をいじめるに鉗子を使用します。網膜が完全に分離するまでは、鉗子で虹彩、レンズ、硝子体、および任意の残りの構造を削除します。
注: プロトコルはここで一時停止可能性があります。実験する前のヴィヴォの網膜を使用して場合は、次の手順を実行する前に実験を行います。 - 解剖はさみを使って、網膜の 4 つの緩和削減して、ようにそれは平ら。鉗子で膜上に網膜の各コーナーを軽く押すをニトロセルロース膜 (材料表) の網膜神経節細胞側をマウントします。
メモ: 緩和カットの場所は、網膜の方向 s オプシンのグラデーションを使用するときに任意指定できます。 - 鉗子を使用すると、転送マウントされた網膜 24 ウェル プレート (材料表) の最初の井戸の中には固定用 4% パラホルムアルデヒド (材料表) の 1 つの mL でいっぱい。常温 (材料表) の軌道シェーカーで 24 ウェル プレートを置き、40 分正確に網膜を修正します。
注: 次の洗浄と培養の手順のすべては軌道シェーカーで 24 ウェル プレートで完了する必要があります。 - よく満ちている 1 mL の 0.1 M PBS 第 2 に転送して室温で 15 分間の網膜を洗います。2 回連続 0.1 M PBS いっぱい 3 番目と 4 番目の井戸に網膜を転送することによって、この手順を繰り返します。
- ソリューション (1.7% トリトン X-100 および 5.2% ロバ血清 0.1 M PBS で;材料表参照) ブロックの 5 も含む 1 mL にマウントされた網膜を転送し、4 ° C で一晩インキュベート
- ウサギ抗のオプシンの一次抗体を追加 (材料の表を参照) 1: 500 の濃度の解決の妨害に、4 ° C で 3 日間インキュベート
- 六つの井戸を室温で 10 分ごとに 1 mL の 0.1 M PBS でいっぱいで順番に配置することによって 6 回網膜から余分な一次抗体を洗います。
- 新鮮なブロッキング液 (1.7% トリトン X-100 および 5.2% ロバ通常血清 0.1 M PBS) で井戸に網膜を配置し、ロバ抗家兎 Alexa 594 二次抗体を追加 (材料の表を参照してください)。4 ° C で一晩二次抗体と網膜を孵化させなさい
- 1 ml 室温で 10 分間新鮮な 0.1 M PBS の六つの井戸に順番に配置することによって 6 回網膜から余分な二次抗体を洗浄します。
- 鉗子を使用して、0.1 M PBS を含むシャーレにマウントされた網膜を転送します。網膜が添付されていないまで軽く網膜と膜間の鉗子の先端を挿入することによって硝酸セルロースの膜から網膜をリリースします。
- 網膜がガラスに付着するまで優しくで鉗子の肝いりでガラス顕微鏡スライドの網膜をマウントし、ペトリ皿からスライドを削除します。
- Aquamount のスライドに網膜をカバーし、#1.5 coverslip でそれをカバーします。スライド トレイにスライドを配置 (材料の表を参照) し、室温で 1 時間放置します。
- スライド トレイに格納しても冷蔵庫のスライドを返す (材料の表を参照) で使用しない場合は 4 ° C。スライドは 24 時間 coverslipped した後、乾燥を防ぐためにスライドの側面をシールするのにマニキュアを使用します。
注: プロトコルはここで一時停止することができます。
4 網膜の方向を識別するために S オプシンと網膜 Immunostained を再構築.
- 共焦点顕微鏡とカメラの付いた epifluorescent 顕微鏡のオプシン グラデーションを可視化 (材料の表を参照) として網膜全体が表示されます(図 1 b、2 D、3 a、および1 つのイメージで、網膜をイメージ3 D). これは、低倍率でのセクションで網膜を画像と画像を一緒にステッチで行うことができます。
- 個人を特定できるので、網膜の名前を付けます。たとえば、名前に最初網膜再建"Retina1"。
- ダウンロードし、https://imagej.nih.gov/ij/download.html で ImageJ をインストールします。
- 再建されるが、フォルダーを空のままにする必要がありますそれぞれの網膜の個々 のフォルダーを作成します。たとえば、"Retina1"というフォルダーを作成します。この網膜を再構築に必要なすべてのファイルは、以降の手順でこのフォルダーにされます。
注: これらのフォルダーを含める必要がありますファイルのみ、Retistruct で分析することをファイルです。下記のもの以外のファイルは Retistruct ソフトウェアで開くことができませんの網膜になります。 - ファイルを選択する ImageJ で網膜の画像を開く →「Retina1」を選択すると開く。
- 画像に変更を加えず、タイトルのフォルダーに"image.png"として保存"網膜 1「ファイル → 名前を付けて保存 → png 形式を選択します。
注: ファイルを Retistruct ソフトウェア再構成のため網膜としてファイルを認識するために"image.png"という必要があります。 - 網膜の端を説明するには、セグメント化された [線] ツールを使用します。網膜の境界線の 2 つの隣接するスポットをクリックして、分割線] ツールは、本質的に「つなぐドット」アウトラインを作成する 2 つの隣接するスポット。繰り返して、全体の網膜に概説されています。網膜のアウトラインを分析を選択することによって"Retina1"のタイトルのフォルダーに"outline.roi"として保存 → ツール → ROI マネージャーは保存 → [t] → を追加します。
注: 網膜の端は、バック グラウンドに移行 s オプシン染色識別できます。 - 4.7 のステップの指示通り、視神経乳頭の境界線のアウトラインを作成するのに分割線] ツールを使用します。視神経乳頭のアウトラインを分析を選択することによって"Retina1"のタイトルのフォルダーに"od.roi"として保存 → ツール → ROI マネージャーは保存 → [t] → を追加します。
注: 視神経乳頭、網膜の真ん中に小さな穴として識別され、郭清の品質によって異なります。
注: すべてのファイル ("image.png"、"outline.roi"、"od.roi") Retistruct 復興のため必要今保存してくださいフォルダー"Retina1"。 - ダウンロード、インストール、および Retistruct プログラムを開くするには、Sterrattらの補足資料セクションにある Retistruct ユーザー ガイドで説明されている手順に従います24
- ウィンドウが登場して、Retistruct をクリックして上部に「オープン」のアイコン ウィンドウの左し、"Retina1"ディレクトリ フォルダーを選択します。
- スケール バーが存在しないことを示す画像ウィンドウが表示されます。「閉じる」をクリックし、網膜のイメージがボックスに表示されます。上にある「プロパティ」ボタンをクリックしてして網膜のアウトラインを視覚化する表示色 (図 5A) ウィンドウと変更の輪郭線の右。
- 重要: 網膜が右目または左 (図 5A) のパネルに左目からかどうかを指定します。
- 左側の「涙を追加」ボタンをクリックしてし涙またはカットの涙 (図 5A) の 3 つ頂点をクリックして網膜のある場所を指定します。これはカットの 3 つの頂点を接続する線を作成します。網膜にあるすべてのカットに対して繰り返します。
- 網膜のアウトラインの任意の点をクリックして、背側の網膜を指定します。大文字の文字"D"は、アウトライン (図 5B) にその時点で表示されます。
注意: 背側の網膜のオプシン グラデーションの反対側の網膜の暗い半分になります。ただし、Retistruct で背側網膜のマーキングは「背」のマーキングはこのステップで任意にすることができますので、網膜の背側半分を識別するための信頼性の高い方法ではありません。 - 上部に「網膜の再構築」ボタンをクリックして網膜左画面 (図 5B) の再構築。再建された網膜の極座標は、カットの概要 (図 5C) と同じ色で表示されますが表示されます。
- 再建された網膜およびすべてのそれに関連付けられているデータは"Retina1"フォルダー (図 5D) に保存されますので、画面の右上の「保存」ボタンをクリックしてします。
- 再建された網膜を保存するには、右側のパネル (図 5D) の"PDF"ボタンをクリックします。サイズ仕様を求めるボックスが表示されます。既定のサイズは次の手順のために許容します。このアクションは、"Retina1"フォルダー ディレクトリに"image.polar.pdf"として再建された網膜が保存されます。
- ペイント プログラム (またはその他のイメージの処理プログラム) で「image.polar.pdf」を開き、「ペイント」ツールを使用して (または類似の) 再建された網膜背景黒に変更します。"Retina1"フォルダー ディレクトリに"Retina1_reconstructed.tif"などの .tif ファイルとして再構築した網膜を保存します。
注: プロトコルはここで一時停止することができます。 - "Retina_Rotator.m"(補足資料を参照) と呼ばれる網膜を回転させるための MATLAB のコードをダウンロードしてください。独自のフォルダーでコード ファイルを配置フォルダーに他のファイルはないです。
- MATLAB、バージョン 2007 b を開くまたはそれ以降。MATLAB で開くコード ファイルをダブルクリックします。コマンド ウィンドウで"Retina_Rotator"を入力し、enter キーを押します。検索ウィンドウが表示されます。
注: コードは、.tif ファイルの特定です。回転するファイルが正しい形式でない場合、コードが網膜を正しく回転しないでしょう。4.17 と 4.18 の手順を参照してください。適切な形式で再建された網膜を保存します。 - 回転するファイルを開きます。たとえば、"Retina1_reconstructed.tif"を選択します。コード再建された網膜を分析し、自動的に元のファイルのあるフォルダーに"Retina1_reconstructed_rotated.tif"として回転した網膜を保存します。
- コードの網膜の分析が完了すると前に、と後の比較 (図 3 bと 3 C回転網膜の画像を表示ウィンドウが開きますも図 3E 、 3 f)。
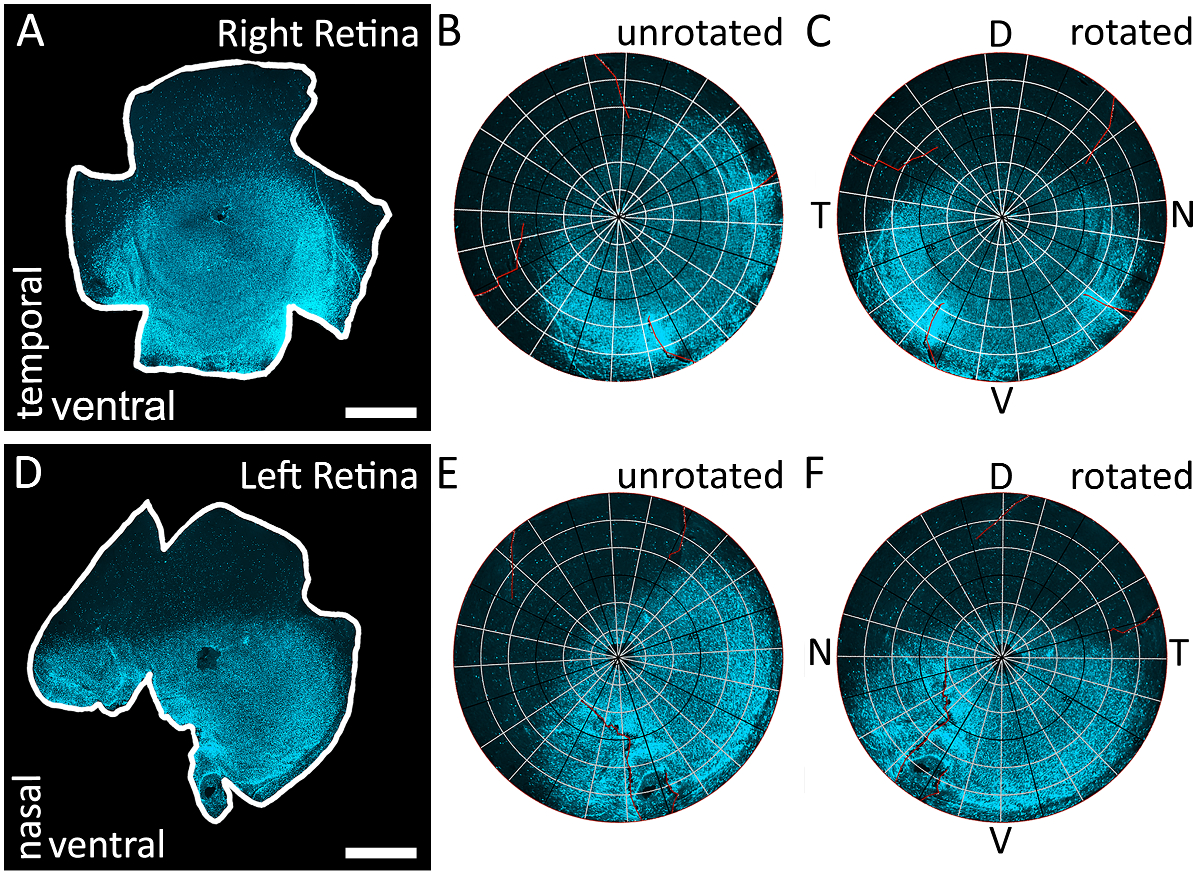
注: このコードは、腹側 (明るい) 半分が下、背 (明度) 半分従って正確に定位 s オプシン グラデーション1によると網膜の上に再建された網膜を回転します。場合は網膜が右目から左目が記載されているか、鼻と一時的な棒の置き場所も向き (図 3) のこのメソッドから推定できます。
結果
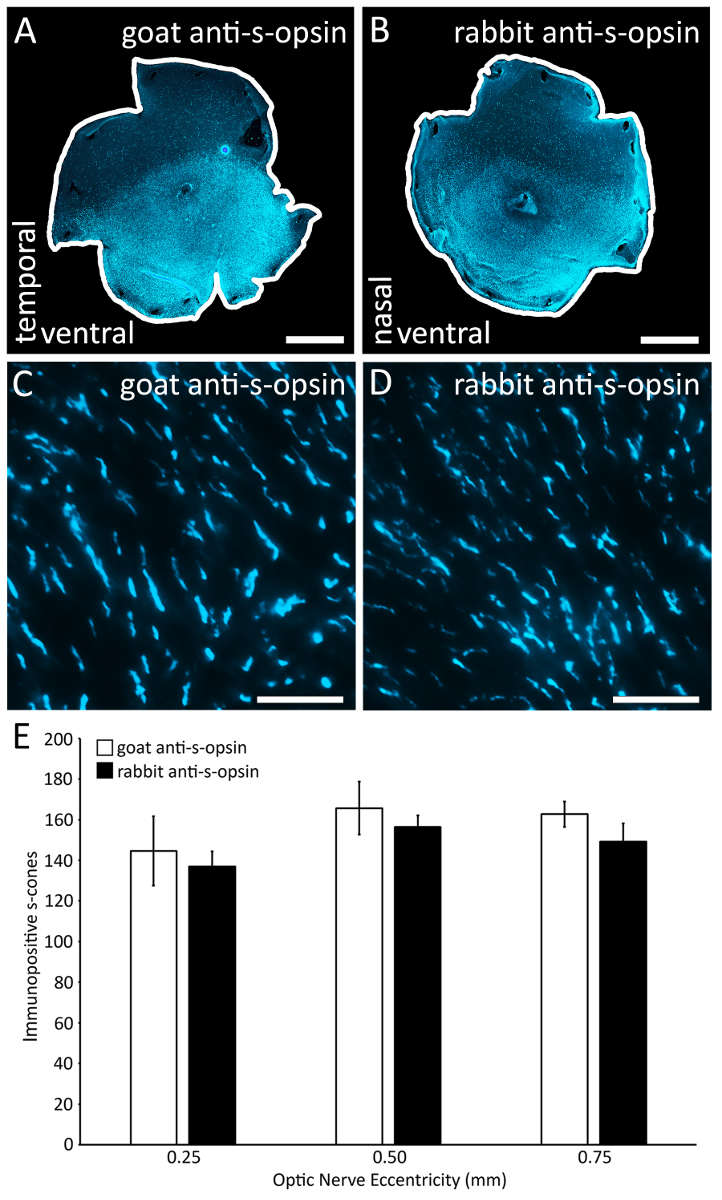
正確かつ確実に直筋を二等分単一緩和カットは、背側の網膜 (図 1) を識別します。脈絡裂正確かつ確実に鼻と一時的な網膜を深い緩和時間と鼻の脈絡裂 (図 2) に沿って識別します。この例では、緩和のカットもしました背側網膜の網膜 (図 2D、垂直の矢印) の背/腹軸を識別するために。将来 dissectors によってレプリケーションの目的のこれらのプロセスの手順を示します。S オプシン免疫組織化学の組み合わせ (図 3 a と3 D)、(3 b, 3 e) Retistruct ソフトウェアによる再建とカスタム MATLAB コード (3 C、3 f) と正確な回転が可能、鼻と時間極これは網膜が右または左目 (図 3) かどうかがわかっている場合と同様に、網膜の腹側と背側半分の id。S オプシン コーン (図 4A ~ D) をラベリング効果の 2 つの一般的に使用される s オプシン一次抗体も比較した: 両方ヤギ アンチのオプシンの一次抗体、ウサギ抗のオプシンの一次抗体に効果的にラベルを付ける同じマウス s オプシン コーン (図 4E)。
緩和カットされた s オプシン再建 immunostained 網膜上識別され、s オプシン勾配によって決定される方位の場所を比較しました。当社のカスタム MATLAB コード (補足資料参照)、網膜のオプシン染色の最高濃度のある腹、真配置、正確に回転しました (直) の 90 ° で 0 ° (鼻の真の鼻背脈絡裂) と時間 (時間的脈絡裂) の 180 ° 真します。角をカット緩和個々 の価値は、網膜を s オプシン グラデーションによると回転した後に ImageJ で角度ツールを使用して決定されました。各緩和カットタイプの平均角度を計算したし、ポーラー プロット (図 6) でプロットしました緩和カット種類ごとの平均値。平均すると、筋肉のカットは上直筋識別 96.3 ± 4.3 ° 背極 (n = 11) (図 6)。鼻の脈絡裂 6.7 ± 5.8 ° 鼻ポールと一時的な脈絡裂示さ 172.0 ± 4.4 ° 時間極 (n = 9;図 6)。

図 1: 右目の背側の網膜を正確に識別する上直筋を使用しています。(A) 焼灼ペン (白い矢印) で作られた角膜強膜の国境近くの背側角膜火傷の例。上直筋はこのビュー (白い矢印) に表示されますも。全体の例 (B) 除去のマウントされた網膜は、上直筋を二等分して背側網膜に作られたカット。矢印は、上直筋を二等分によって背側網膜に作られたカットを緩和深を示しています。網膜は一次抗体ヤギ抗-s-オプシンと汚れる (材料の表を参照) と二次抗体ロバの反やぎ Alexa 594 (材料の表を参照してください励起: 590 nm; 排出量: 620 nm) (水色)。網膜をテキサス赤フィルター付き epifluorescent 顕微鏡をイメージしました (595 nm)。(C) A 網膜 Retistruct で再建され、MATLAB のカスタム コードを使用して回転 (補足資料を参照してください)、上直筋効くと目に見えるカット (白い矢印)。D: 背, v: 腹側、t: 時間、n: 鼻。スケール バー = 1 mm.この図の拡大版を表示するのにはここをクリックしてください。
{kind=link}

図 2: 脈絡裂を使用して正確に右目の網膜の鼻と時間の極を識別する。(A) 焼灼ペンで作られた角膜強膜の国境近くの背側角膜火傷の例。(B) 脈絡膜裂強膜 (白い矢印) で目の後ろに表示されます。背側の角膜のやけどは時間的脈絡裂から約 90 ° に位置する、このビューに表示も。(C) 脈絡膜裂強膜、角膜強膜の枠線を走行視神経からの目の後ろに表示されます。(D) は網膜に染まったヤギ抗-s-オプシン (材料の表を参照してください) と二次抗体ロバの反やぎ Alexa 594 (材料の表を参照してください励起: 590 nm; 排出量: 620 nm) 脈絡裂カット (水平 (シアン)矢印) 切り (垂直方向)、背側の緩和します。網膜をテキサス赤フィルター付き epifluorescent 顕微鏡をイメージしました (595 nm)。(E) 網膜は Retistruct で再建され、背の緩和カットと鼻で、一時的な脈絡裂カット表示 (白い矢印) カスタム MATLAB コード (補足資料を参照してください) を使用して回転します。D: 背, v: 腹側、t: 時間、n: 鼻。スケール バー = 1 mm.この図の拡大版を表示するのにはここをクリックしてください。
{kind=link}

図 3: s オプシンのグラデーションを使用して網膜のすべての 4 本の棒を識別する。S オプシンのラベルに immunostained をされており、テキサス赤フィルター付き epifluorescent 顕微鏡を用いたイメージング右目から網膜の例 (A) 解剖 (595 nm)。地形向き、s オプシン グラデーションによって決まりますので、この網膜でカット、任意です。(B) Retistruct A で網膜の再建の結果。S オプシン グラデーションがない正しく並んでいること網膜がカスタム MATLAB コード (補足資料を参照してください) を実行されていないので注意してください。(C) カスタム コードで網膜を回転の結果。網膜は、s オプシン染色の最高濃度は下部にありますように回転し、腹側の網膜として識別されています。網膜は、右の目からは、一時的なポールは位置 90 ° 背極から反時計回りに、鼻ポール位置 90 ° 背極から時計回りに。(D) 網膜の例は左眼のオプシンのラベルに immunostained をされており、テキサス赤フィルターでイメージから解剖 (595 nm)。地形向き、s オプシン グラデーションによって決まりますので、この網膜でカット、任意です。(E) Retistruct と D で網膜をデジタル再構成の結果。S オプシン グラデーションがない正しく並んでいること網膜がカスタム コードによって回転されていないので注意してください。(F) カスタム コードと D で網膜を回転の結果。網膜は、s オプシン染色の最高濃度は下部にありますように回転し、腹側の網膜として識別されています。網膜は、左の眼からは、鼻のポールの位置 90 ° 背極から反時計回りと一時的な棒が位置 90 ° 背極から時計回りに。D: 背, v: 腹側、t: 時間、n: 鼻。スケール バー = 1 mm.この図の拡大版を表示するのにはここをクリックしてください。
{kind=link}

図 4: s オプシン コーンをラベルに 2 つのプライマリ s オプシン抗体の比較。(A) A 網膜ヤギ抗のオプシンの一次抗体で染色 (材料の表を参照してください)。 (B) 同じマウスの他の網膜に染まったウサギ抗のオプシン一次抗体 (材料の表を参照してください)。(C) A ヤギ抗のオプシンの一次抗体で染色網膜から代表 (0.1 × 0.1 mm2)Epifluorescent 顕微鏡を用いた 40 倍の倍率で撮影した画像。網膜から (D) の代表的な地域 (0.1 × 0.1 mm2) に染まったウサギ抗-s-オプシン (材料の表を参照)、一次抗体の代替。イメージは、40 倍の倍率で epifluorescent 顕微鏡で撮影されました。(E) 両抗体のラベル s コーンの外側のセグメントの数が同じヤギ抗-s-オプシンとウサギ抗-s-オプシン テスト網膜の染色陽性の錐体の数に差がないので離心率 (n = 2;アドホックの記事ボンフェローニ テストと分散分析p > 0.05)。スケール バー = 1 mm (A B);25 μ m (C ・ D)。この図の拡大版を表示するのにはここをクリックしてください。
{kind=link}

図 5: s オプシンと網膜 immunostained を再構築する Retistruct ソフトウェアを使用するための視覚的なガイドします。アウトライン表示で Retistruct で開かれた A (A) 網膜と、「涙」に追加。「涙」のポイントは、スーパーイン ポーズの白い矢印で示されます。この網膜にあるすべてのカットは、します解剖時に網膜の方向をマークする特定のランドマークが使用されていない任意の。重要なボタンの輪郭が赤に。すべての「涙」追加 (B) A 網膜と網膜の端に"D"が付いて背網膜。「網膜の再構築」ボタンが表示されます注意してください。重要なボタンの輪郭が赤に。(C) 網膜の再建のプロセス。再建された網膜の極座標は、シアン (青い矢印のカット位置を明確にする重ね合わせ) でカット、緩和を示す右に表示されます。(D) Retistruct を通じて網膜を実行する最終的な結果。元 wholemount 網膜そのまま左側と右側再建された網膜に表示されます。緩和のカットは、シアン (カット位置を明確にする重ね合わせ白い矢印) で表示されます。この図の拡大版を表示するのにはここをクリックしてください。
{kind=link}

図 6: 上直筋と脈絡膜裂を正確にマウス網膜の向きを使用できます。カットを緩和、上直筋から得られる角度の極座標または脈絡裂は Retistruct と再建されている網膜にカットできます。緩和カットされた s オプシン再建 immunostained 網膜に識別され、s オプシン グラデーションの場所の場所を調べた。カスタムの MATLAB のコードを使用すると、正確に網膜を回転して s オプシン染色の最高濃度が腹側に位置する、真背 (90 ° 上直筋)、真の鼻 (鼻脈絡裂の 0 °)、真の一時的な脈絡膜のための一時的な (180 °裂) 各網膜を求めた。カット アングルは、ImageJ と平均の角度で決定されたそれぞれの個別緩和の値は、緩和の型をカットごとに算出しました。上直筋カット識別 96.3 ± 4.3 ° 背極 (n = 11)。鼻の脈絡裂 6.7 ± 5.8 ° 鼻ポールと一時的な脈絡裂示さ 172.0 ± 4.5 ° 時間極 (n = 9)。この図の拡大版を表示するのにはここをクリックしてください。
{kind=link}
| 深いランドマーク | 角膜のやけどの場所 | 識別される網膜の極 | 実験的アプリケーション |
| 上直筋 | 背側 | 背側 | ライブまたは固定 |
| 鼻の脈絡裂 | 背側 | 鼻 | ライブまたは固定 |
| 一時的な脈絡裂 | 背側 | 一時的です | ライブまたは固定 |
| S オプシン グラデーション | どれも | 背、腹、鼻、時間 | 固定 |
表 1: 深いランドマーク、網膜の極彼らを識別するとかライブや固定組織のアプリケーションに使用できます。
ディスカッション
ない包括的な標準化されたプロトコルを確認し解剖学的スペースでマウス単離網膜の方向をラベリングされています。プロトコルの詳細ここで標準化することによって、この空白を埋めるしようと深い解剖学的ランドマークを使用する方法について詳しく説明するポイントを確実に参照として識別網膜の方向。このプロトコルの深い解剖学的ランドマークが角膜の火傷22など表面的なランドマークよりマウス網膜の方向付けのためのより正確で信頼性の高いメソッドを提供することが示されています。したがって、網膜の方向の角膜の火傷に頼ってきた研究にあったかもしれないより大きいエラー脈絡裂、腹直筋などのランドマークに頼ってきた研究よりも向き。この不一致は、結果を解釈し、正確な網膜の向きに依存する研究間で比較を行うこの標準化された議定書の意義と必要性を強調表示します。全体的にみて、ビジョン研究者に従うの一般的な方法を提供する標準化されたプロトコルがなくなり、網膜を識別するための標準メソッドを使用が発生するデータ集録の交絡変数の存在向きです。
紹介した方法は、簡単に再現性と実験的プロトコルの多くの種類に適用されます。実際には、このプロトコルの最大の利点の 1 つはその適応性です。データ集録 (テーブルを最適化する実験的パラメーターに最適なランドマークを選択ことができます脈絡裂、s オプシン式と直筋ランドマークすべて発見されている網膜方向22を確実に同定するため1). さらに、郭清の方法はさらに網膜の方向を明確にまとめることができます。網膜のすべての 4 本の棒を方向づけるために s オプシン免疫組織化学と脈絡裂カットを組み合わせることができますたとえば、: 脈絡裂カットによって鼻と一時的な半球を識別でき、s オプシン免疫組織化学を識別することができます腹側と背側の半球。まだ、このプロトコルの適応性は、生理学実験の時間に敏感な性質によって制限可能性があります。ランドマークを識別、角膜のやけど、緩和カットを実行にかかる時間は、体外実験で重要な組織死で起因できる、未満の最適なこれらの郭清の方法のいくつかあります。幸いなことに、深いランドマークを識別し、緩和、解剖に脈絡裂または上直筋解離法に精通するなるとカット迅速に郭清のルーチンの部分になるし、大幅追加しないでください。郭清の長さ。組織の生存率はもはや問題 (図 3 網膜向きのホックを投稿の s オプシン グラデーションを使用してお勧めただし、ここで説明されている手順非常に時間に敏感な実験の時間に追加できることを認識、).S オプシンは、すべての 4 本の棒を特定すること、網膜を方向づけるための効果的な方法のために網膜を染色: 背側と腹側の極に網膜を分割し、可能にするかどうかによって鼻と時間の同定が極 s オプシン染色網膜右または左の目 (図 3) からです。したがって、このプロトコルが実験的パラメーターを満たすことができる正確な網膜方向法の信頼性と再現性のあるセットを提供と考えています。
として、任意の変更された網膜の郭清を伴う、郭清法の有効性は切開と隔離されている組織の品質の精度によって制限されます。解剖時に任意の組織が失われた場合、正確な再建のため、網膜が破損するとあまりにも Retistruct と MATLAB プログラムされません確実に再構築および網膜の方向を設定することができます。したがって、実験データ収集のために使用前に郭清法を実践することが重要です。解剖の種類は、ここで困難ではありませんを説明しながら、特定のランドマークと網膜の方向を識別する再現性の確保に練習されなければなりません。さらに、正しいランドマークと確信させるのにデータ収集を開始する前に解剖学的ランドマークを視覚的に識別する解剖実習が使用されていることが不可欠です。特定の凝固切開装置の精度を確認する方法の 1 つはいずれかの脈絡裂にカットまたは上直筋を切り取って固定マーカーは、したがって、dissectio の精度に依存しないので s オプシン グラデーションにカットの位置を比較(名) 潜在的な dissectors は、カットは図 1に示すように正確なランドマークと再建された網膜の例に再建された網膜を比較することもと図 2。基本的には、潜在的な解剖特定解離型のこのプロトコルで説明されている手順を実行する必要があります、かどうかは上直筋または脈絡膜裂法と結果の妥当性を確立する s オプシン グラデーションを比較、特定のバイナリ。網膜の不正確な方向に切開がランドマークの場所について確認の場合がありますので、デフォルトに影響を与えるデータの収集と解釈。
開示事項
著者が明らかに何もありません。
謝辞
ブルターニュは、その技術支援のジェシカ Onyak、リュー親切に私たちは彼 epifluorescent 顕微鏡を使用させるために感謝したいと思います。サポートの確認: NIH R15EY026255-01 とカール ・ Kirchgessner 財団。
資料
| Name | Company | Catalog Number | Comments |
| 0.1 M Phosphate Buffered Saline | Sigma-Aldrich | P5244 | |
| Axioplan2 Epifluorescent Microscope | Zeiss | N/A | |
| Clear Nailpolish | N/A | N/A | |
| Corning LSE Low Speed Orbital Shaker | Sigma-Aldrich | CLS6780FP | |
| Costar TC-Treated 24-well Plates | Sigma-Aldrich | CLS3524 | |
| Dissection Microscope | Olympus | SZ51 | |
| Donkey anti-Goat Alexa 594 | Life Technologies | A11058 | |
| Donkey anti-Rabbit Alexa 594 | Life Technologies | A21207 | |
| Donkey Normal Serum | Millipore | 566460 | Use at 5.2% (52 μL with 86 μL of 20% Triton X-100 and 863 μL of 0.1M PBS for 1 mL of blocking solution) |
| Fisherbrand Superfrost Plus Microscope Slides | Fisher Scientific | 12-550-15 | |
| Goat anti-s-opsin | Santa Cruz Biotechnologies | sc-14363 | Not commerically available as of 2017 |
| Graefe Curved Forceps | Fine Science Tools | 11052-10 | |
| ImageJ or FIJI | National Institute of Health | N/A | Freely available software |
| Low Temperature Cautery Ophthalmic Fine Tip Cauterizer | Bovie Medical Corporation | AA00 | |
| MATLAB | MathWorks | N/A | At least version 2007b or later |
| Micro Cover Glasses | VWR International | 48393-241 | |
| Micro Slide Trays | VWR International | 82020-913 | |
| Moira Ultra Fine Forceps | Fine Science Tools | 11370-40 | |
| Nitrocellulose membrane | Millipore | HAWP04700 | |
| Paraformaldehyde | Electron Microscopy Sciences | 15714-S | Use at 4% (25 μL and 875 μL of 0.1 M PBS for 1 mL of fixative) |
| PrecisionGlide Needle 20G (0.90 mm x 25 mm) | BD PrecisionGlide | 305175 | |
| Pyrex Glass Petri Dish | Sigma-Aldrich | CLS3160152 | |
| R | The R Project for Statistical Computing | N/A | Freely available software; version 3.4.3 or later |
| Rabbit anti-s-opsin | Millipore | ABN1660 | |
| Retiga R3 Microscope Camera | Qimaging | 01-RET-R3-R-CLR-14-C | |
| Retistruct | N/A | N/A | Freely available software compatiable with Windows 7 or Windows 10 |
| Shandon Aqua-Mount Slide Mounting Media | Fisher Scientific | 14-390-5 | |
| Triton X-100 | Sigma-Aldrich | T8787 | Use 1.7% (86 μL of 20% Triton-X with 52 μL of Donkey Normal Serum and 863 μL of 0.1 M PBS for 1 mL of blocking solution) |
| Vannas Spring Dissection Scissors | Fine Science Tools | 15000-03 | |
| 5MP USB Microscope Digital Camera | AmScope | MU500 | To be used with the Olympus Dissection Microscope |
参考文献
- Applebury, M. L., et al. The murine cone photoreceptor: A single cone type expresses both S and M opsins with retinal spatial patterning. Neuron. 27 (3), 513-523 (2000).
- Hughes, S., Watson, T. S., Foster, R. G., Peirson, S. N., Hankins, M. W. Nonuniform distribution and spectral tuning of photosensitive retinal ganglion cells of the mouse retina. Curr Biol. 23 (17), 1696-1701 (2013).
- Sondereker, K. B., Onyak, J. R., Islam, S. W., Ross, C. L., Renna, J. M. Melanopsin ganglion cell outer retinal dendrites: Morphologically distinct and asymmetrically distributed in the mouse retina. J Comp Neurol. 525 (17), 3653-3665 (2017).
- Bleckert, A., Schwartz, G. W., Turner, M. H., Rieke, F., Wong, R. O. L. Visual space is represented by nonmatching topographies of distinct mouse retinal ganglion cell types. Current Biology. 24 (3), 310-315 (2014).
- Warwick, R. A., Kaushansky, N., Sarid, N., Golan, A., Rivlin-Etzion, M. Inhomogeneous Encoding of the Visual Field in the Mouse Retina. Curr Biol. 28 (5), 655-665 (2018).
- Valiente-Soriano, F. J., et al. Distribution of melanopsin positive neurons in pigmented and albino mice: evidence for melanopsin interneurons in the mouse retina. Front Neuroanat. 8, 131 (2014).
- Sabbah, S., et al. A retinal code for motion along the gravitational and body axes. Nature. 546 (7659), 492-497 (2017).
- Vaney, D. I., Sivyer, B., Taylor, W. R. Direction selectivity in the retina: Symmetry and asymmetry in structure and function. Nat Rev Neurosci. 13 (3), 194-208 (2012).
- Huberman, A. D., et al. Genetic identification of an On-Off direction-selective retinal ganglion cell subtype reveals a layer-specific subcortical map of posterior motion. Neuron. 62 (3), 327-334 (2009).
- Ueki, Y., Ramirez, G., Salcedo, E., Stabio, M. E., Lefcort, F. Loss of Ikbkap causes slow, progressive retinal degeneration in a mouse model of familial dysautonomia. eNeuro. 3 (5), (2016).
- Maiorano, N. A., Hindges, R. Restricted perinatal retinal degeneration induces retina reshaping and correlated structural rearrangement of the retinotopic map. Nat Commun. 4, 1938 (2013).
- Hadj-Said, W., et al. Quantitative and topographical analysis of the losses of cone photoreceptors and retinal ganglion cells under taurine depletion. Invest Ophthalmol Vis Sci. 57 (11), 4692-4703 (2016).
- Tao, Y., et al. The temporal topography of the N-Methyl- N-nitrosourea induced photoreceptor degeneration in mouse retina. Sci Rep. 5, 18612 (2015).
- Risner, M. L., Pasini, S., Cooper, M. L., Lambert, W. S., Calkins, D. J. Axogenic mechanism enhances retinal ganglion cell excitability during early progression in glaucoma. Proc Natl Acad Sci U S A. , (2018).
- Estevez, M. E., et al. Form and function of the M4 cell, an intrinsically photosensitive retinal ganglion cell type contributing to geniculocortical vision. J Neurosci. 32 (39), 13608-13620 (2012).
- Kolesnikov, A. V., Kefalov, V. J. Transretinal ERG recordings from mouse retina: Rod and cone photoresponses. J Vis Exp. (61), (2012).
- Lin, B., Wang, S. W., Masland, R. H. Retinal ganglion cell type, size, and spacing can be specified independent of homotypic dendritic contacts. Neuron. 43 (4), 475-485 (2004).
- Ortin-Martinez, A., et al. Number and distribution of mouse retinal cone photoreceptors: differences between an albino (Swiss) and a pigmented (C57/BL6) strain. PLoS One. 9 (7), 102392 (2014).
- Zhang, H., et al. The degeneration and apoptosis patterns of cone photoreceptors in rd11 Mice. J Ophthalmol. 2017, 9721362 (2017).
- Wei, W., Elstrott, J., Feller, M. B. Two-photon targeted recording of GFP-expressing neurons for light responses and live-cell imaging in the mouse retina. Nat Protoc. 5 (7), 1347-1352 (2010).
- Wang, J., et al. Anatomy and spatial organization of Muller glia in mouse retina. J Comp Neurol. 525 (8), 1759-1777 (2017).
- Stabio, M. E., et al. A novel map of the mouse eye for orienting retinal topography in anatomical space. J Comp Neurol. 526 (11), (2018).
- Lamb, T. D., Collin, S. P., Pugh, E. N. Evolution of the vertebrate eye: Opsins, photoreceptors, retina and eye cup. Nat Rev Neurosci. 8 (12), 960-976 (2007).
- Sterratt, D. C., Lyngholm, D., Willshaw, D. J., Thompson, I. D. Standard anatomical and visual space for the mouse retina: Computational reconstruction and transformation of flattened retinae with the Retistruct package. PLoS Comput Biol. 9 (2), 1002921 (2013).
転載および許可
このJoVE論文のテキスト又は図を再利用するための許可を申請します
許可を申請さらに記事を探す
This article has been published
Video Coming Soon
Copyright © 2023 MyJoVE Corporation. All rights reserved