
Method Article
機能向上レポーター・システムによる特異的マイコバクテリア・誤訳率の測定
要約
本稿では、機能的なレポーターシステムを用いて、モデル・ smegmatisの特定の翻訳誤差と誤訳を測定する2つの相補的な方法を提示します。この方法を使用して、高スループットの設定で、低スループットまたは相対エラー率で正確なエラー率を測定することができます。
要約
タンパク質への遺伝子の翻訳は誤りを起こしやすい。モデル系における並進誤差の平均速度はコドンあたり 1/10000 であると推定されるが、実際の誤差率は、研究されている種、環境、およびコドンによって大きく異なる。我々は以前に、マイコバクテリアが aminoacylated グルタミンおよびアスパラギン tRNAs の生成のための2段階の経路を使用することを示しており、これは具体的には誤訳率の変調に起因する比較的高いエラー率に関連していることを示す経路の必須成分は、amidotransferase GatCAB である。以前に使用されていたレニラルシフェリン-Firefly の二重ルシフェラーゼシステムを修正し、グルタミン酸での特定の誤訳率を測定するために、マイコバクテリアにおける誤訳率を測定しました。アスパラギンコドンに対するコドンおよびアスパラギン酸。このレポーターシステムは、特定のエラー率の正確な推定に適していましたが、過剰な操作手順の感度と要件の欠如は、高スループットのアプリケーションには適していませんでした。そこで、Nluc ルシフェラーゼと緑色蛍光タンパク質 (GFP) を用いて、中・高スループットの設定により適した2つ目の機能向上レポーターシステムを開発しました。このシステムを用いて、マイコバクテリアの誤訳を減少させることができる小分子としてカスガマイシンを同定した。ここで説明するレポーターは、特定のタイプのマイコバクテリア誤訳を測定するために使用されていますが、多くのモデルシステムで他のタイプの誤訳を測定するように変更することができます。
概要
分子生物学における情報の流れは、遺伝子情報を機能タンパク質に変換することを必要とします。すべての生物学的システムと同様に、遺伝子翻訳も測定可能な誤差を伴います。翻訳における誤差の割合の推定は、典型的には、コドン当たり約 1/10000 として引用される (リバス de Pouplana et al.1によって見直される)。しかし、エラー率は 10-5未満から 0.05/コドン1、2、3、4、5に大きく変化します。3桁以上の誤差率の広い範囲は、翻訳経路の複数のステップからエラーが発生する可能性があるという事実によるものです: aminoacylation6のストキャスティクス、変異、またはストレス誘発エラーから、7,8,9,図10は、アスパラギニル-およびグルタミニルペプチド-tRNAs5の生理的 misacylation、またはリボソーム復号エラー2、3、11である。程度までは 0.01/コドンを超える高いエラー率を示し、翻訳エラーが生理的機能1,12を実行し、誤訳が文脈特異的13である可能性があることを示唆しました。
我々と他の人々は、特に環境ストレス1、5、12、14、15、16 時に、遺伝子翻訳における自然発生的なエラーが適応可能であることを示している 、17、18。マイコバクテリアにおいて、aminoacylation 経路19、20の2段階の間接的グルタミン/アスパラギン tRNA によって生成された誤差は、第1のライン antituberculosis 抗生物質リファンピシンに対して著しく増加した耐性をもたらす5.そのため、小分子でマイコバクテリアの誤訳が減少すると、リファンピシンによる殺傷を増強可能性があると推測しました。我々は、マイコバクテリアの誤訳を減少させることができる化合物として天然に存在するアミノグリコシドカスガマイシンをスクリーニングして同定し、インビトロおよびインビボでの抗酸菌の増強リファンピシン媒介性の死滅を抑制し、世界で最も致命的な病原菌である結核22の世界的制御を脅かすリファンピシン耐性21の出現。
翻訳誤差を研究するには、誤訳の測定方法を採用する必要があります。誤訳の測定のために開発された複数の方法があり、それぞれに長所と短所があります。簡単に言えば、精密質量分析法に基づく方法はいくつかの利点を有するが、その最も重要なものは、複数のタイプの翻訳誤差を検出するための新しいアルゴリズムを用いて、誤訳の比較的偏りのない測定が可能であることである18を行いました。しかしながら、質量分析法は、脱アミド化誤訳事象の測定にはあまり適しておらず、正確にはエラーが起こりやすい間接 tRNA aminoacylation 経路に起因するマイコバクテリアに発生する誤訳の種類である。これは、質量分析器23のための試料の処理において発生する高周波おける脱アミド化のためであり、極めて高いバックグラウンドシグナルをもたらす。したがって、この経路におけるエラーの検出については、遺伝的機能の増加のレポーターは、明確な利点を提供します。具体的には、適切な機能の利益のレポーターは、非常に低いバックグラウンドレートを持つことができ、非常に低いエラー率11の測定を可能にします。
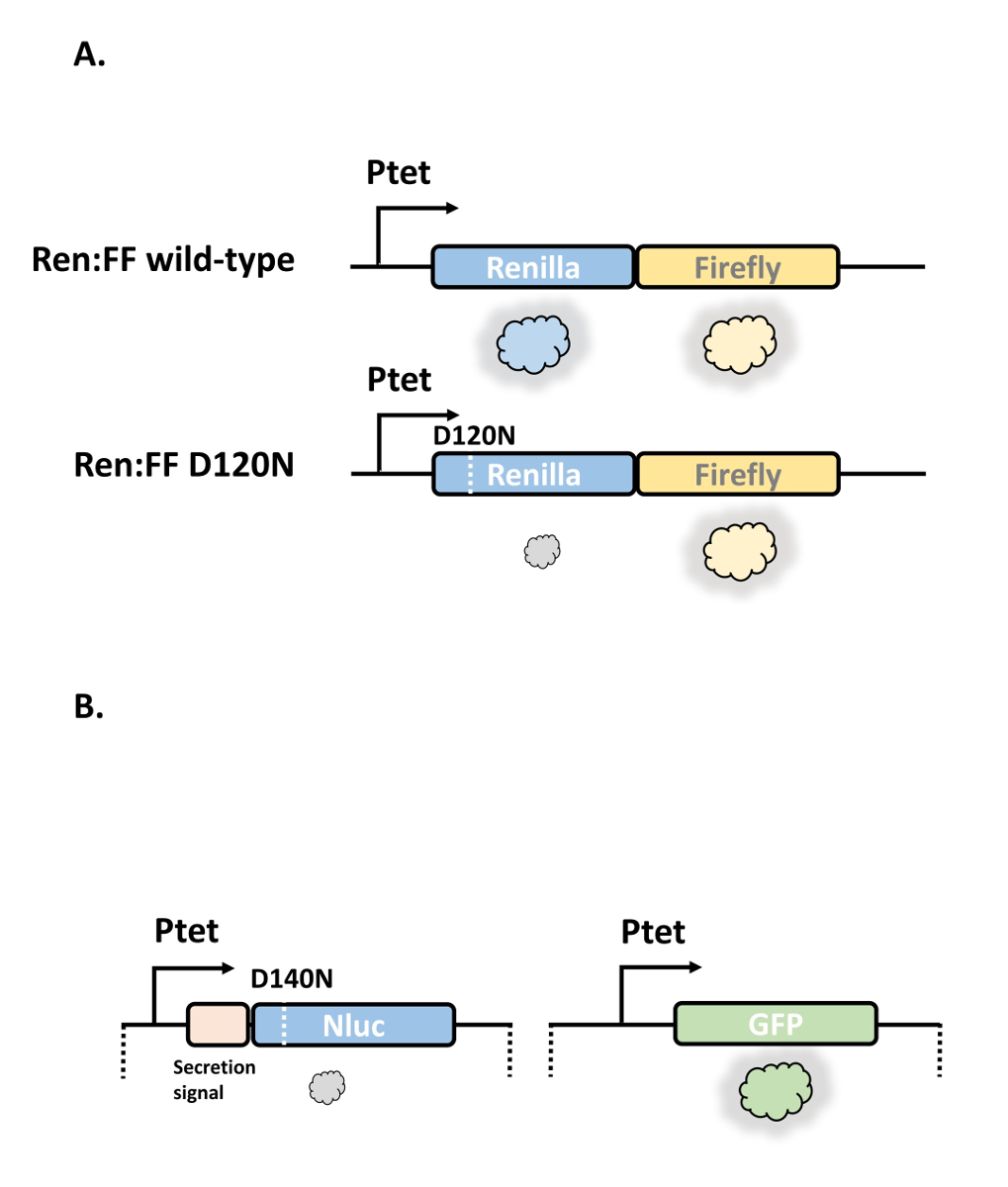
病原性結核菌による実験を行うには特殊な設備と特別な予防措置が必要であるため、我々は nonpathogenic smegmatis でほとんどの実験を行い、以前にそのことを示した。 この2種の間の結果は、概ね5,21と比較可能である。間接 tRNA aminoacylation 経路により生成されたマイコバクテリアにおける誤訳率を測定するために、大腸菌でのリボソームの復号エラーを測定するために開発済みのレニラルシフェリン-Firefly の二重ルシフェラーゼシステムを改良しました。11マイコバクテリアでの使用のため。我々は3つの特異的修飾を行った: 元のレポーターはマイコバクテリアで効率的に発現せず、したがって、配列はコドン最適化され、そしてその C 末端はホタルルシフェラーゼの3つのアミノ酸、セリン-リジン−ロイシンいくつかのシステム24において人身売買シグナルとして注釈付けされ、-アラニン-バリン25のイソロイシンを修飾した。元のレポーターは、ホタルルシフェラーゼに重大なリジン残基を変異させました。代わりに、我々は、レニラルシフェリンルシフェラーゼ中の批判的に保存されたアスパラギン酸 (D120) またはグルタミン酸 (E144) 残基をそれぞれ25 個(図 1) にアスパラギンとグルタミンに変異した。レポーターは、エピソームテトラサイクリン誘導性プラスミド pUV-tetOR (材料の表を参照) にサブクローニングしました。機能を持つレポーターは、非機能的11,26をレンダリングする酵素/蛍光タンパク質において、極めて保存された機能残基を変異させます。タンパク質の機能的変異体を合成する並進 (または理論的には転写) エラーは、翻訳されたタンパク質のサブセットにおいて測定可能な酵素活性をもたらすであろう。タンパク質の存在量の変化を補正するために、変異レポーターは、ベンチマークとして機能し、有し11の正確な定量を可能にする機能性タンパク質と共に作用します。レニラルシフェリン-ホタルの二重-ルシフェラーゼレポーターは、特定のマイコバクテリア誤訳率5,25 (プロトコルの図 2とセクション 1) の正確な測定を可能にしましたが、我々はすぐに誤訳率を変化させる分子の中・高スループットスクリーニングには適していないことがわかった。これは主に2つの理由、すなわち、a)レニラルシフェリンルシフェラーゼの効力の相対的な欠如が誤訳率を測定するのに最低1ml のマイコバクテリア培養/サンプルが必要であることを意味し、b) 前の細胞の溶解の要件酵素活性測定を必要とする過剰な手動処理: マイコバクテリア細胞は、溶解に比較的耐性のある厚いおよび多層の細胞壁およびエンベロープを有する。したがって、我々は、小容量 (例えば、96プレートシステム) で使用することができ、測定のための細胞溶解を必要としなかった新しい機能の関数型レポーターシステムを開発することを検討しました。我々は、非常に強力な Nluc ルシフェラーゼを用い、アスパラギンに突然変異したときに、2ログの機能の損失をもたらした臨界アスパラギン酸残基を同定した (図 1)。さらに、Nluc のサイズが小さいため、抗酸菌27の主要な分泌された抗原である抗原85A から N 末端分泌シグナルタグを持つことが可能になり、Nluc は培養上清中に分泌し、その要件を回避することができる。細胞溶解ベンチマークタンパク質である GFP は、変異 Nluc と同じプロモーターから表されたが、統合ベクター (材料の表を参照) から、無傷の細胞21で測定することができた (図 3)。これらの利点にもかかわらず、Nluc/GFP レポーター (プロトコルのセクション 2) には不利な点もあります: D140N 変異による Nluc 活性 (100-倍) の比較的控えめな減少は、極めて低い誤訳率の測定を許可しないでしょう、レポーターが翻訳誤差の正確な測定よりもスクリーニングツールとしてより適していること。さらに、Nluc には重要なグルタミン酸残基がありません。したがって、アスパラギン-アスパラギン酸のみの誤差率を測定することができます。.本研究で説明されている一般的な原則は、彼らのモデルシステムにおける他の特定の並進誤差率の正確かつ/または容易なの測定に適切なようにレポーターを使用するか、または変更するために調査者がこれらのレポーターを利用することを可能にすべきである選択。
プロトコル
1.レニラルシフェリン-ホタル二重ルシフェラーゼレポーター
注:このメソッドを視覚的に表現するには、図 2を参照してください。
- 7H9 培地の 2 mL の-80 ° c のストックからの接種性マイコバクテリアレポーター株。野生型二重ルシフェラーゼ、および変異レニラルシフェリン(レポーター) 株は、誤訳率の計算を可能にするために使用される必要があります (ステップ1.7 を参照)。37° c で 1 ~ 2 日間、OD600が静止相 (od > 3) に達するまで振盪します。
- 0.1-0.5 の周りにアリコートし、OD600nmに希釈します。典型的な実験では、3つの独立した生物学的複製培養を使用する。Anhydrotetracycline (ATC) は、最終濃度 50 ng/mL までのレポーター発現 (テトラサイクリン誘導性プロモーターによって制御される) のテトラサイクリンアナログインデューサーです。
- 誤訳率21にカスガマイシンの効果を測定するために、培養物に異なる用量のカスガマイシン (示された用量については図 4を参照) を同時に加える (試験した用量では、カスガマイシンは抗菌活性を有さない)。各レポーターに対して、少なくとも1つの非誘導制御を含めることが重要であることに注意してください。培養-37 ° c で 4-6 h の培養を振盪と共に誘導する。
- 細菌培養液を 2 mL チューブに移し、室温で5分間 3220 x gで遠心分離し、バクテリアを下にペレットにします。上清を破棄します。
- 二重蒸留水で希釈された (1:1) 1x パッシブ溶解バッファー40μ l を添加することにより、細菌を破壊する。再懸濁された細菌の溶解物を白色の96ウェルプレートに移し、試料ごとに1つよく、室温で20分間振る。
注意:(30 分以上) 溶解バッファー内の細菌を過剰にインキュベートしないでください。 - 各ウェルに80μ l のホタル基材を加え、15秒間振り、積分時間として 1000 ms でルミノメーターして発光を測定した。自動インジェクターまたはマルチチャンネルピペットを使用して、ピペット操作によるエラーを回避します。
- レニラルシフェリン基板の80μ l を各ウェルに添加し、15秒間振盪し、積分時間として 1000 ms でルミノメーターすることにより発光を測定した。
- 背景の発光を減算-野生型m. smegmatis (すなわち、レポーターを含まない) または uninduced レポーター溶解物のいずれかを使用して測定され、測定値から。訂正された値を使用して、次の式11,25を使用して各条件の誤訳率を計算し、DN が変異レポーターひずみの活動を参照します。

2. Nluc/GFP レポーター
注:このメソッドを視覚的に表現するには、図 3を参照してください。
- 7H9 培地 2 mL の-80 ° c のストックから細菌レポーター株を接種する。37° c で 1 ~ 2 日間、OD600が静止相 (od > 3) に達するまで振盪します。
- サブカルチャーを 50 mL の7H9 培地に、そして、OD600が後期固定相 (> 4) に到達するまで増殖する。
- 96ウェルプレートに細菌を等分する前に、50 ng/mL の最終濃度で ATC を追加し、よく混ぜる。バルク培養の誘導により、すべてのウェルに同量のインデューサーが含まれ、レポーターの誘導が同期されます。各井戸の容積の100μ l が付いている明確な、丸底の 96-井戸の版に細菌をアリコート。
- 誤訳率に影響を与える小分子をスクリーニングするには、指定された濃度で化合物を加えて、井戸を選択します。このプロトコルの目的のために、カスガマイシン (図 5で示される用量で) を実例として使用してください。ウェルを選択するためにカスガマイシンの異なる投与量を追加します (各実験グループは、少なくとも2つの生物学的反復を含む必要があります)。
- 37° c のサンプルを 16-20 h でシェイクして誘導します。
注:フィルムでプレートを密封する必要があります。また、すべてのエッジウェルは、試験井戸からの蒸発を制限するために、少なくとも200μ l の滅菌水を充填する必要があります。 - 各ウェルから80μ l をとり、マルチチャンネルピペットを使用して、サンプルを黒の96ウェルプレート (蛍光シグナル測定を最大化する) に移します。積分時間として20ミリ秒でルミノメーターして GFP 信号を測定します。
- GFP 信号を測定した後、10分間 3220 x gの上清の50μ l のプレートを、白色の底部96ウェルプレート (発光信号測定を最大化する) に遠心分離し、50μ l の Nluc 基板を各ウェルに添加し、それらをよく混合し、そして積分時間として 1000 ms とルミノメーターによって発光を測定してください。
- GFP 蛍光によって補正された Nluc 発光値を分割することによって Nluc/GFP 比を決定する: この尺度 (任意の単位で) は、アスパラギン誤訳に対するアスパラギン酸の相対的な尺度である。
結果
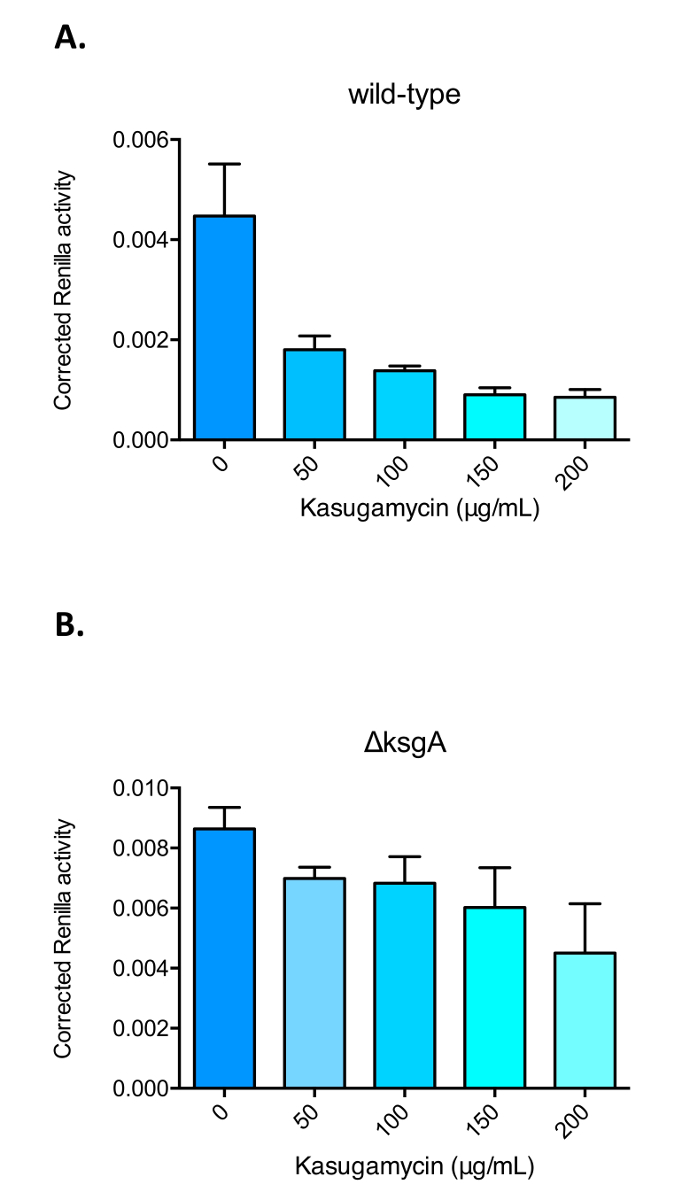
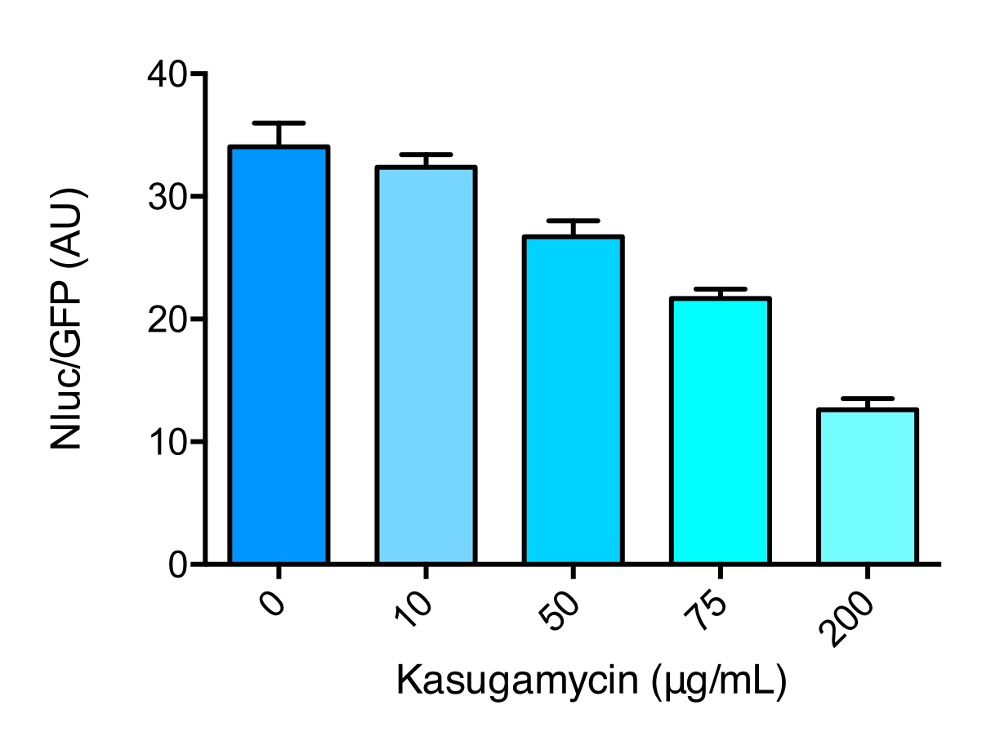
この作品で使用されている2つのレポーターシステムの概要を示す漫画を図 1に示します。図 2に、プロトコルのセクション1の概要と図 3のセクション2の概要を示します。カスガマイシンにおけるマイコバクテリア誤訳に対する影響を図 4に示したもので、レニラルシフェリン-ホタルレポーターシステムによって測定される。カスガマイシン作用の特異性を示すために、レポーターはまた、 ksgAが削除されたsmegmatisの株で表された (δ ksgA)。KsgA は rRNA ジメチルスルホキシドトランスフェラーゼで、 KsgAはカスガマイシンに対して比較的耐性があります。誤訳に対するカスガマイシン作用も、図 5に示すように NLUC/GFP レポーターを用いて測定した。

図 1: 漫画は、2つの機能の利得のレポーターシステムを説明します。(A)レニラルシフェリン-ホタルレポーターシステムは、融合タンパク質として発現する2つのルシフェラーゼ酵素と、テトラサイクリン誘導性プロモーターの制御下で構成されています。野生型二重酵素の発現は、レニラルシフェリンとホタルルシフェラーゼ (上) の両方の高い測定可能な活性をもたらす。重要なアスパラギン酸残基の変異は、D120、レニラルシフェリンにおいてはレニラルシフェリン酵素 (D120N) を不活性にレンダリングするが、ホタルルシフェラーゼは依然として活性である。アスパラギンのアスパラギンに対する誤訳 (この場合、生理的に misacylated した Asp-trna のAsn trna5,21) は、わずかな割合で翻訳されたレニラルシフェリンタンパク質の活性を獲得し、を測定することができる。野生型二重酵素と比較した変異レポーターのレニラルシフェリン/Firefly 活性の比較により、アスパラギンの計算をアスパラギン酸誤訳率にすることができます。(B) NLUC/GFP レポーターシステムは、同様の原理で動作します。変異 Nluc (D140N) 遺伝子は、抗原85A 由来の N 末端分泌シグナルを有し、主要な分泌型マイコバクテリアタンパク質である。Nlucとgfp遺伝子の両方が、同一のテトラサイクリン誘導性プロモーターから発現し、gfp を相対発現ベンチマークとして使用できるようにします。野生型 Nluc 制御株の欠如に注意してください: このレポーターは主にスクリーニングに使用され、主画面に対して相対誤訳率の測定で十分です。この図の大規模なバージョンを表示するには、ここをクリックしてください。
{kind=link}

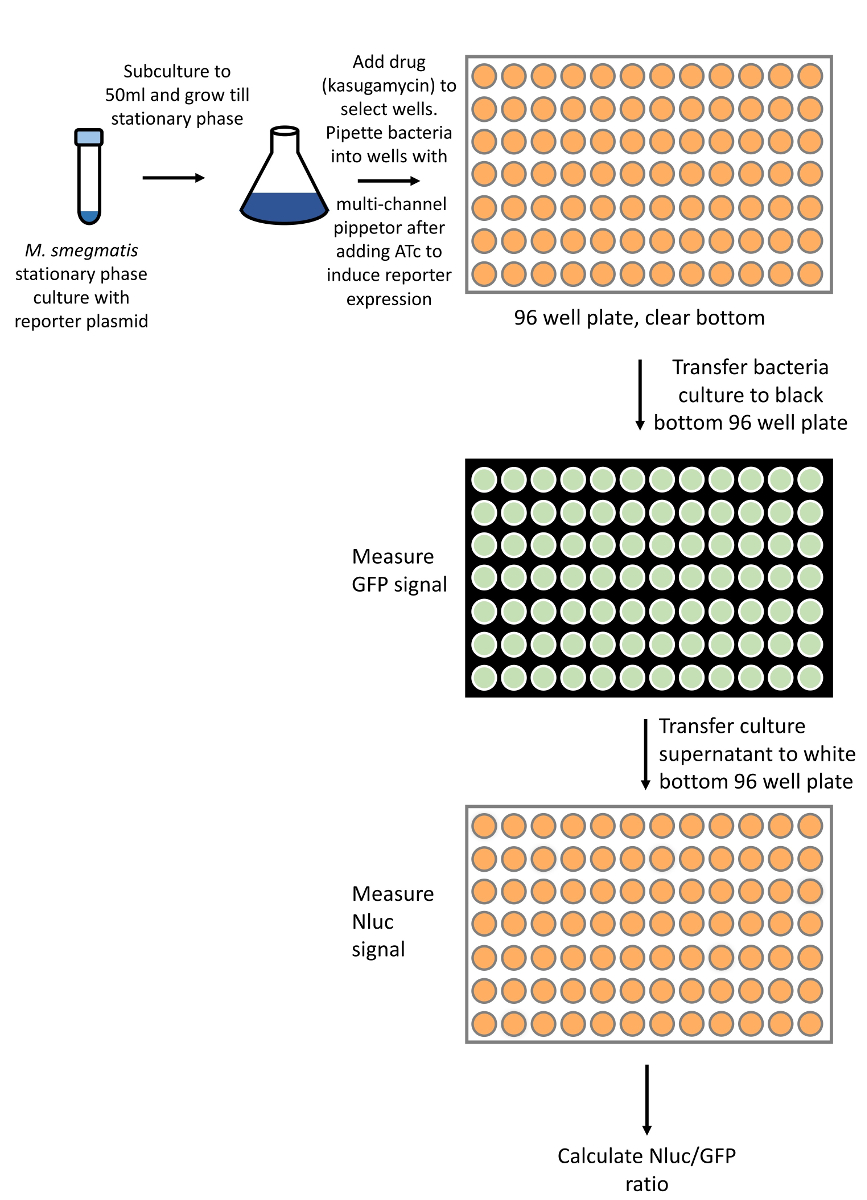
図 2:レニラルシフェリン-ホタルレポーターを用いた誤訳率の測定の概要 (プロトコルのセクション 1)。Smegmatisの新鮮な培養物を、二重-ルシフェラーゼレポーターを発現するプラスミドで形質転換し、増殖させる。レポーター発現が誘導され、治験化合物または条件がテストされます。4-6 h のレポーター発現に続いて、培養物をペレット化し、溶解させるとともに、溶解液を二重ルシフェラーゼ活性について試験します。この図の大規模なバージョンを表示するには、ここをクリックしてください。
{kind=link}

図 3: 誤訳率 Nluc-GFP レポーター (プロトコルのセクション 2) の測定の概要。Smegmatisの新鮮な培養物を、NLUC および GFP レポーターで形質転換し、試験されるすべてのプレートについて十分な量に成長させる (〜 10 mL/96 ウェルプレートを想定)。細菌培養物を各ウェルにピペットで入れる直前に、バルク培養に ATC を追加してレポーターの発現を誘導する。一晩それらを振って、井戸をインキュベートします。相対誤訳率を測定するには、培養物をブラックプレートに移し (蛍光検出の感度を上げるため)、GFP 蛍光を測定し、その後、プレートをスピンして細菌をペレットにします。上清を Nluc の活性測定のために白いプレートに移す。この図の大規模なバージョンを表示するには、ここをクリックしてください。
{kind=link}

図 4: カスガマイシンの存在下において誤訳率を測定するレニラルシフェリン-ホタルレポーターを用いた代表的な結果。アスパラギン酸の誤訳率 (A) 野生型m. Smegmatisおよび (B) ksgAのために削除された株 (δ ksgA) カスガマイシン (Ksg) による処理の後、レニラルシフェリン-Firefly デュアルで測定したもの記者。培養物を、細胞溶解および測定の前に、指示された用量 (ミクログラム/ミリリットル) で6時間にわたってカスガマイシンで治療した。補正された Ren/FF 比 (y 軸) は、アスパラギン誤訳率に対するアスパラギン酸の指標である。KsgAの削除は、カスガマイシンによる変調に対してより耐性の高いベースライン誤訳率をもたらす。バーは平均値を示し、標準偏差は誤差となります。この図の大規模なバージョンを表示するには、ここをクリックしてください。
{kind=link}

図 5: カスガマイシンの存在下における誤訳率を測定した Nluc レポーターを用いた代表的な結果である。Nluc/GFP レポーターによって測定される野生型m. smegmatisにおけるアスパラギンに対するアスパラギン酸の相対誤訳率は、カスガマイシン (ksg) 一晩 (16 時間) での処理に続く。バーは平均値を示し、標準偏差は誤差となります。この図の大規模なバージョンを表示するには、ここをクリックしてください。
{kind=link}
ディスカッション
ここで説明するプロトコルは、多種多様な生物における誤訳率の測定に適合させることができる。プロトコルを他のシステムに適応させる際には、留意すべき考慮事項がいくつかあります。まず、測定の目的を考慮する必要があります。機能的なレポーターを用いて誤訳率を正確に測定するために必要なのは、(a) ロバスト読み出しアッセイ (例えば、酵素機能 [この場合、ルシフェラーゼ活性] の広い線形範囲) です。また、(b) レポータータンパク質の重要な残留物における機能突然変異の損失を調べて、予想される誤訳の速度を下回り、機能の非常に重要な損失をもたらします。例えば、予想される誤訳率が約 10-3/codon である場合、関数変異の損失は1000よりも大きくなければなりません。さもなければ、誤訳事象のより低い範囲は敏感に検出されません。最後に、機能的なレポーターを用いて誤訳率を正確に測定するには、(c) 堅牢なベンチマークレポーターが必要です (この場合、プロトコルのセクション1では firefly ルシフェラーゼ)。理想的には、ベンチマークレポーターは、変異酵素レポーターに融合し、それらが両方とも等モル比で表されることを保証する (すべての意図と目的のために) 必要があります。ベンチマーク (およびプライマリレポーター) の読み出しは、露出した環境への摂動に対して堅牢である必要があります。例えば、野生型 GFP の蛍光は pH28の変化による摂動の影響を強く受けており、マクロファージの貪食マイコバクテリアにおける誤訳率の測定基準としては不適当である。pH 729の下にある phagosomes。一方、小分子スクリーニングなどの用途では、一次スクリーンに対する最も重要な考慮事項は、測定の再現性 (精度とは対照的に精度)、手動処理の最小化、および小文化のボリューム。誤訳率の正確な測定はそれほど重要ではなく、二次アッセイとして行うことができます。最後に、このプロトコルに記載されたレポーターは、ルシフェラーゼ酵素の酵素的機能に基づいて、細菌集団の平均誤訳率を測定するだけであるが、単一細胞の不均一性についての情報を与えることはできないということに留意すべきである。誤訳率のばらつき。適応表現型における単一細胞変動性の重要性を考えると、30,31,32, 誤訳事象の測定のための蛍光レポーターが開発された12,33、マイコバクテリア5における誤訳の測定を含む。
誤訳の測定のための要件を考慮したが、このプロトコルに記載されているような遺伝的利益のレポーターは、1つのタイプの誤訳 (すなわち、1つのアミノ酸置換) しか測定できないことに留意すべきである) レポーターあたりのコドンである。したがって、異なる誤訳事象の測定には、複数のレポーターが必要となる。例えば、リジルオキシダーゼ tRNA による近接性コドンのリボソーム復号エラーによる誤訳の測定を1つの位置で行うためには、少なくとも16人のレポーターが2、3、11を必要とする。高精度質量分析とバイオインフォマティクスにより、多くの異なるタイプの誤訳イベントを同時に18の潜在的な識別が可能になります。ただし、これらの方法にも注意が伴います。一般に、これらの関数は、機能向上のレポーターよりも精度が低くなります。さらに、先に述べたように、それらは特定のタイプの誤訳、すなわちおける脱アミド化23で conflated ことができるものの検出にはあまり適していません。最後に、質量分析法は、中またはハイスループットスクリーニングのための読み出しとして不適当である。
他のシステムにおける誤訳の測定のためのこれらのレポーターおよびプロトコルの適合のために、さらなる考慮は、所望のモデルシステムにおけるレポーターのロバストな表現を含む。我々は当初、Farabaugh や同僚によって開発された二重ルシフェラーゼシステムを改変なしで、我々のマイコバクテリアシステムで使用しようと試みたが、レポーター発現の欠如のために成功しなかった。広範なトラブルシューティングでは、コドンの最適化と、レポーターのマイナーシーケンスの変更の両方が、ロバストな表現25 (および上記を参照) を可能にすることが必要であることが確認されました。他のシステムで使用するには、レポーターの同様の適応が必要であると考えられます。
最後に、測定される誤訳事象の種類について検討すべきである。例えば、停止コドン読破 (ナンセンス抑制) を測定する必要がある場合、停止コドンの機能変異の消失は、原発性レポータータンパク質34の重要でない領域内に導入される必要がある。さもなければ、重要な残留物 (ひいてはレポーター機能) を回復させる特定の無意味な抑制イベントだけが、真の誤訳率のかなりの過小評価を引き起こす可能性があるアッセイによって測定されます。多数の生物1、12、13、35において翻訳エラーが適応的な役割を果たす可能性があるという証拠が増えている。しかし、誤訳事象の測定は、依然として少数のモデル種に限定されています。誤訳を測定するための敏感な方法の適応は、生理学と病理学における翻訳ミスの役割に対する科学者の理解をさらに高める可能性を秘めている。
開示事項
作者は何も開示することはありません。
謝辞
この作品は、ビル・アンド・メリンダ・ゲイツ財団 (OPP1109789)、中国国立自然科学財団 (31570129)、清華大学医学部から B.J. B.J. への助成金によって支えられ、ウェルカムの信頼を受けています。調査員 (207487/z/17/Z)。
資料
| Name | Company | Catalog Number | Comments |
| Middlebrook 7H9 | BD Difco | 271310 | |
| Anhydrotetracycline | Cayman Chemical | 10009542 | |
| Kasugamycin | sigma | K4013 | |
| Dual-luciferase reporter assay system | promega | E1960 | |
| Nano-Glo luciferase assay | promega | N1120 | |
| Fluoroskan Ascent FL luminometer | Thermo | / | |
| Assay Plate, 96 Well White, Flat Bottom High Binding, No Lid | Costar | 3922 | |
| Assay Plate, 96 Well Black, Flat Bottom High Biding, No Lid | Costar | 3925 | |
| 96 Well Cell Culture Cluster, Flat Bottom with Lid | Costar | 3599 | |
| Non-commercial reagents (plasmids) | |||
| pUV-TetOR-RenFF | NA | NA | episomal shuttle plasmid that allows tetracycline-inducible expression of the wild-type dual luciferase reporter |
| pUV-TetOR-Ren-D120N-FF | dual-luciferase reporter with mutated Renilla | ||
| pUV-TetOR-Ag85ASec-Nluc-D140N | NA | NA | episomal shuttle plasmid with a tetracyclin-inducible secretable version of mutated Nluc luciferase |
| pMC1S-GFP | NA | NA | Mycobacterial integrated (L5 site) vector for tetracycline-inducible expression of GFP |
参考文献
- Ribas de Pouplana, L., Santos, M. A., Zhu, J. H., Farabaugh, P. J., Javid, B. Protein mistranslation: friend or foe. Trends in Biochemical Sciences. 39 (8), 355-362 (2014).
- Leng, T., Pan, M., Xu, X., Javid, B. Translational misreading in Mycobacterium smegmatis increases in stationary phase. Tuberculosis (Edinburgh). 95 (6), 678-681 (2015).
- Manickam, N., Nag, N., Abbasi, A., Patel, K., Farabaugh, P. J. Studies of translational misreading in vivo show that the ribosome very efficiently discriminates against most potential errors. RNA. 20 (1), 9-15 (2014).
- Netzer, N., et al. Innate immune and chemically triggered oxidative stress modifies translational fidelity. Nature. 462 (7272), 522-526 (2009).
- Su, H. W., et al. The essential mycobacterial amidotransferase GatCAB is a modulator of specific translational fidelity. Nature Microbiology. 1 (11), 16147 (2016).
- Li, L., et al. Naturally occurring aminoacyl-tRNA synthetases editing-domain mutations that cause mistranslation in Mycoplasma parasites. Proceedings of the National Academy of Sciences of the United States of America. 108 (23), 9378-9383 (2011).
- Li, L., et al. Leucyl-tRNA synthetase editing domain functions as a molecular rheostat to control codon ambiguity in Mycoplasma pathogens. Proceedings of the National Academy of Sciences of the United States of America. 110 (10), 3817-3822 (2013).
- Ling, J., Soll, D. Severe oxidative stress induces protein mistranslation through impairment of an aminoacyl-tRNA synthetase editing site. Proceedings of the National Academy of Sciences of the United States of America. 107 (9), 4028-4033 (2010).
- Raina, M., et al. Reduced amino acid specificity of mammalian tyrosyl-tRNA synthetase is associated with elevated mistranslation of Tyr codons. Journal of Biological Chemistry. , (2014).
- Wu, J., Fan, Y., Ling, J. Mechanism of oxidant-induced mistranslation by threonyl-tRNA synthetase. Nucleic Acids Research. 42 (10), 6523-6531 (2014).
- Kramer, E. B., Farabaugh, P. J. The frequency of translational misreading errors in E. coli is largely determined by tRNA competition. RNA. 13 (1), 87-96 (2007).
- Evans, C. R., Fan, Y., Weiss, K., Ling, J. Errors during Gene Expression: Single-Cell Heterogeneity, Stress Resistance, and Microbe-Host Interactions. mBio. 9 (4), (2018).
- Mohler, K., Ibba, M. Translational fidelity and mistranslation in the cellular response to stress. Nature Microbiology. 2, 17117 (2017).
- Bullwinkle, T. J., et al. Oxidation of cellular amino acid pools leads to cytotoxic mistranslation of the genetic code. Elife. 3, (2014).
- Fan, Y., et al. Heterogeneity of Stop Codon Readthrough in Single Bacterial Cells and Implications for Population Fitness. Molecular Cell. 67 (5), 826-836 (2017).
- Fan, Y., et al. Protein mistranslation protects bacteria against oxidative stress. Nucleic Acids Research. 43 (3), 1740-1748 (2015).
- Schwartz, M. H., Pan, T. Temperature dependent mistranslation in a hyperthermophile adapts proteins to lower temperatures. Nucleic Acids Research. 44 (1), 294-303 (2016).
- Schwartz, M. H., Waldbauer, J. R., Zhang, L., Pan, T. Global tRNA misacylation induced by anaerobiosis and antibiotic exposure broadly increases stress resistance in Escherichia coli. Nucleic Acids Research. , (2016).
- Curnow, A. W., et al. Glu-tRNAGln amidotransferase: a novel heterotrimeric enzyme required for correct decoding of glutamine codons during translation. Proceedings of the National Academy of Sciences of the United States of America. 94 (22), 11819-11826 (1997).
- Rathnayake, U. M., Wood, W. N., Hendrickson, T. L. Indirect tRNA aminoacylation during accurate translation and phenotypic mistranslation. Current Opinion in Chemical Biology. 41, 114-122 (2017).
- Chaudhuri, S., et al. Kasugamycin potentiates rifampicin and limits emergence of resistance in Mycobacterium tuberculosis by specifically decreasing mycobacterial mistranslation. eLife. 7, (2018).
- Toosky, M., Javid, B. Novel diagnostics and therapeutics for drug-resistant tuberculosis. British Medical Bulletin. 110 (1), 129-140 (2014).
- Robinson, N. E., Robinson, A. B. Deamidation of human proteins. Proceedings of the National Academy of Sciences of the United States of America. 98 (22), 12409-12413 (2001).
- Miura, S., et al. Urate oxidase is imported into peroxisomes recognizing the C-terminal SKL motif of proteins. European Journal of Biochemistry. 223 (1), 141-146 (1994).
- Javid, B., et al. Mycobacterial mistranslation is necessary and sufficient for rifampicin phenotypic resistance. Proceedings of the National Academy of Sciences of the United States of America. 111 (3), 1132-1137 (2014).
- Wong, S. Y., et al. Functional role of methylation of G518 of the 16S rRNA 530 loop by GidB in Mycobacterium tuberculosis. Antimicrobial Agents and Chemotherapy. 57 (12), 6311-6318 (2013).
- Wiker, H. G., Harboe, M. The antigen 85 complex: a major secretion product of Mycobacterium tuberculosis. Microbiological Reviews. 56 (4), 648-661 (1992).
- Roberts, T. M., et al. Identification and Characterisation of a pH-stable GFP. Scientific Reports. 6, 28166 (2016).
- Li, H., Wu, M., Shi, Y., Javid, B. Over-Expression of the Mycobacterial Trehalose-Phosphate Phosphatase OtsB2 Results in a Defect in Macrophage Phagocytosis Associated with Increased Mycobacterial-Macrophage Adhesion. Frontiers in Microbiology. 7, 1754 (2016).
- Aldridge, B. B., et al. Asymmetry and aging of mycobacterial cells lead to variable growth and antibiotic susceptibility. Science. 335 (6064), 100-104 (2012).
- Rego, E. H., Audette, R. E., Rubin, E. J. Deletion of a mycobacterial divisome factor collapses single-cell phenotypic heterogeneity. Nature. 546 (7656), 153-157 (2017).
- Zhu, J. H., et al. Rifampicin can induce antibiotic tolerance in mycobacteria via paradoxical changes in rpoB transcription. Nature Communications. 9 (1), 4218 (2018).
- Lant, J. T., et al. Visualizing tRNA-dependent mistranslation in human cells. RNA Biology. 15 (4-5), 567-575 (2018).
- Grentzmann, G., Ingram, J. A., Kelly, P. J., Gesteland, R. F., Atkins, J. F. A dual-luciferase reporter system for studying recoding signals. RNA. 4 (4), 479-486 (1998).
- Melnikov, S. V., van den Elzen, A., Stevens, D. L., Thoreen, C. C., Soll, D. Loss of protein synthesis quality control in host-restricted organisms. Proceedings of the National Academy of Sciences of the United States of America. , (2018).
転載および許可
このJoVE論文のテキスト又は図を再利用するための許可を申請します
許可を申請さらに記事を探す
This article has been published
Video Coming Soon
Copyright © 2023 MyJoVE Corporation. All rights reserved