
Method Article
Bリンパ球活性化を研究するための牽引力顕微鏡
要約
ここでは、B細胞に対する牽引力顕微鏡実験を行うために用いられるプロトコルを提示する。柔らかいポリアクリルアミドゲルの調製とその機能化、顕微鏡でのデータ取得、データ分析の概要について述べています。
要約
トラクションフォース顕微鏡(TFM)は、基材上の細胞によって生成される力の測定を可能にします。この技術は、弾性基板上の細胞引っ張りによって生成された実験的に観察された変位場からの動力力測定を推測する。ここで、B細胞受容体の抗原関与によって活性化された場合のB細胞によって発揮される力場の空間的および時間的構造を調べるTFMを適合させた。ゲル剛性、ビーズ密度、タンパク質の機能化は、細胞表面受容体に対して特異的に反応し、相互作用する比較的小さな細胞(〜6μm)の研究に最適化する必要があります。
概要
B細胞は免疫系の抗体産生細胞である。適応免疫応答を活性化するために、それらは、まずB細胞受容体(BCR)1と呼ばれる特異的受容体を介して天然型(すなわち、非処理された)で抗原を獲得する。このプロセスは、リンパ節B細胞ゾーンで起こる。一部の抗原がリンパ液を介してB細胞に到達できる場合でも、ほとんどの抗原、特に高分子量(リンパ管管の限界サイズである>70 kDa)は、実際に抗原提示細胞(APC)の表面にネイティブの形で提示され、典型的にはサブカプセル状の角座内膜または濾胞性デンドリック細胞、典型的には、眼下のマクロファージまたは濾胞性デンドリック細胞、レクチンまたはFc(Fc)を介して。この細胞との接触は、BCRがAPC関連抗原に力を発揮する免疫シナプスの形成につながる。BCRへの抗原の結合はBCRシグナル伝達を開始し、力発生メカニズムを活性化し得る。これらの力はBCRシグナル伝達を増幅するために重要である可能性がありますが、B細胞が抗原を抽出して内部化するためにも不可欠です。
最近の研究では、BCRは確かにメカノイ感受性2であることを示しています。例えば、硬質基質は、BCRシグナル伝達を強化した3を引き出す。また、免疫シナプスで発生する力は、単一のBBCを引っ張って抗原への親和性をプローブし、それによって親和性の判別を確実にする。したがって、抗原提示に対するB細胞の機械的応答を調べ、この応答を関与する受容体の種類(IgG/IgM)5、接着分子(インテグリンリガンド)、または薬理学的および遺伝子組み換え細胞(すなわち、BCRシグナル伝達または細胞骨格の下流のタンパク質のサイレンシング)の観点から解剖することは興味深い。
生理的剛性の基質に対する細胞の応答を観察する簡単な方法と、同時に、基質に及ぼす研究力は、トラクションフォース顕微鏡(TFM)である。TFMは、弾性基板上の細胞引っ張りによって生成される変位場を観察する。もともとゲルの変形は、位相対視7によってエラストマー自体のシワを介して観察されたが、蛍光マイクロビーズを受託マーカーとして挿入することで、より良い分解能を可能にし、それ以来標準8となっている。この方法は、ゲルに埋め込まれた接着細胞、組織、さらにはオルガノイドによって及ぼす牽引力を調べるのに使用されています。TFM のいくつかのバリエーションは、超解像顕微鏡(すなわち、STED10 またはSRRF11)との組み合わせ、TIRF顕微鏡12を可能にするゲルの屈折率の改変、ナノプリントパターン13によるビーズの置換、および平坦面14の代わりにナノピラーを用いた9を含む開発されている。これらのバリエーションの完全なレビューについては、コリン・ヨークら15を参照してください。
ここで提示されるプロトコルは、抗原被覆基質上でB細胞が加えた力を測定する手順を説明する。これらの力は、それらをクラスター化し、その後、抗原提示基質からそれらを抽出するためにリガンド(抗原)に適用されます。我々は、生理学的抗原提示基質の剛性、サイズ及びB細胞の関連コーティングを模倣するために標準TFMプロトコルを適応させました。このプロトコルは、同時に複数の細胞の研究を可能にし、蛍光顕微鏡技術および化学処理と組み合わせて使用することができる。しかしながら、単一分子力測定をプローブすることを目的としていないが、光学ピンセット16、分子張力プローブ17、18、生体膜力プローブ19、及び原子間力顕微鏡20がより適した技術である。他の単一細胞力測定法(例えば、マイクロピペット21またはマイクロプレート22)と比較して、TFMは、シナプスで加えられる力の完全なマップの再構成を、〜300nmの分解能で可能にする。これは、表面に及ぶ力の時空間的パターンを同定し、ゲルが共焦点イメージングと互換性があるように、特定のタンパク質(例えば、細胞骨格およびシグナル伝達タンパク質)のリクルートとそれらを相関させるために有用である。
3D TFMは可能ですが、使用した剛性やセットアップとは互換性がありません。3Dにおける変形は、突起力顕微鏡(細胞がめっきされる変形性膜をスキャンするAFM)および弾性共振器干渉応力顕微鏡(ERISM、光のハイライト及び数ナノメートルの精度で基板の変形を共鳴させるキャビティとして作用するゲル)などの他のより複雑な設定によって達成可能である25。これらの技術は非常に有望であるが、それらはまだB細胞で採用されていない。ナノピラー14など、他のタイプのTFMは、より再現性の高い基質を有するために使用することができる。しかし、この幾何学は細胞が柱を介在するので柔らかい細胞には適合せず、解析を複雑にします。このアプローチは、実際にT細胞で柱26の周りに構造を構築する細胞の能力を観察するために使用されています。
そのシンプルさにもかかわらず、ポリアクリルアミドゲルを使用したTFMは、多くの細胞の同時観察を可能にし、ベンチと蛍光顕微鏡を装備したラボで簡単かつ安価に実装することができます(共焦点/スピニングディスクをお勧めします)。
APCの生理的剛性を模倣するために、我々は、〜500Pa27 の剛性を有するポリアクリルアミドゲルを使用し、活性化抗原でゲルを機能化した。このプロトコルでは、ポリアクリルアミドゲルの表面を鶏卵リソザイム(HEL)で機能させた。これにより、抗原結合部位の関与を通じてBCRの刺激によって発生する力の測定が可能となる。MD4マウス由来のこの抗原とHEL特異的B細胞を用いると、抗原結紮28に応答して比較的均一な力発生が保証される。しかし、他の分子(B6マウスの抗IgMなど)はゲルに移植することができますが、これらの場合に発生する力はより不均一で強くなくなる可能性があります。B細胞は小さな細胞(直径は6μm)であるため、ビーズの数は最大だが追跡可能に最適化されています。基質に対して~kPaの力を発揮する大きな細胞の場合、比較的まばらなビーズを使用して、または単純粒子画像のvelocimetry(PIV)を行って変形場を再構築することで、満足のいく結果を得ることができます。しかし、~50Paほどの小さな応力を及ぼすBリンパ球などの小さな細胞では、変形場を再構築する際に所望の精度を達成するために、単一粒子追跡(粒子追跡用の軌跡、PTV)の使用が必要である。ビーズを個別に確実にトラッキングするには、対物レンズの倍率を少なくとも60倍、その数値開口を1.3程度にする必要があります。したがって、ゲルは比較的薄く(<50 μm)でなければならず、ビーズは目的の作業距離を超えているため、見えません。
主なプロトコルは、ゲル調製、ゲルの機能化およびイメージングの3つのセクションで構成されています。さらに2つのセクションは任意であり、蛍光細胞の抗原抽出の定量およびイメージングに専念している。
プロトコル
1. ゲル調製
- ゲル支持体のシラナイゼーション
- 2分間のUVランプ(残留オゾンへの暴露を避けるためにUVランプへの暴露前に30 s待つ)で、カバースリップまたはガラス底ペトリ皿(ゲルサポートとして使用されます)をアクティブにします。
- 200 μL のアミノプロピルトリメトキシシラン (APTMS) を使用して、カバースリップ/ガラス底皿を 5 分間シラナイズします。これにより、ゲルの共有結合に対する支持体を準備する。
- カバースリップ/ガラス底皿を超純水で十分に洗います。
- 真空吸引を使用してカバースリップ/ガラス底皿を乾燥させます。
- ゲルを平らにするために使用される18mmカバースリップの準備
- カバースリップを準備するには、まずセラミックカバースリップホルダーに入れます。その後、カバースリップホルダーを小さなビーカー(50 mL)に入れ、カバーリップの上にシリコン化試薬(4°Cで保存され、再利用可能)を注ぎ、それらを完全に覆うことを確認します。
- ビーカーをアルミホイルで覆い、室温で3分間インキュベートします。待っている間に、超純水で大きなビーカー(500 mL)を満たします。シリコン化試薬でインキュベーションを3分経過した後、カバースリップホルダーをカバースリップで水のビーカーに移します。
- カバーリップを超純水で徹底的に洗い流し、よく乾燥させ、紙の拭き取りを続けます。最良の結果を得るには、次のセクションに進んでください。
- ゲル重合
- 0.5 kPaのゲルの場合、40%アクリルアミドの75 μLを30 μLの2%ビサクリアミド(架橋剤)と895 μLのリン酸緩衝生理食塩(PBS)と混合します。このプレミックスは、4°Cで1ヶ月まで保存することができます。
- 0.5 kPaゲルプレミックスの167 μLに、1%(1.67 μL)のビーズ、ボルテックス、超音波処理をバスソニケーター(50-100 Wのパワーと周波数40 kHzの標準ベンチ超音波洗浄機)に5分間追加します。アルミニウムホイルを使用して光から保護されたミックスを保ちます。
注: プレミックスは、イニシエータ(TEMED)が追加されるまで重合しません。 - 重合を触媒するには、1%(1.67 μL)を10%w/v過硫酸アンモニウム(APS)に加えます。
- 重合を開始するには、0.1%(0.2 μL)N、N、N、N′、N'-テトラメチルエチレンジアミン(TEMED)を加えます。ピペットと混ぜます。APSとTEMEDを添加すると、ゲルは急速に重合するので、ゲル鋳造に迅速に進みます。
- ゲル鋳造
- 各カバースリップ/ガラス底皿にゲルミックスのピペット9 μL(中央に落とし、図1A)
- シラナイズ/疎水性カバースリップを配置し、ゲルを平らにします(図1B)。鉗子を使用して、カバースリップを押して、漏れ出し始めるまでカバースリップの全領域(図1C)にゲルが広がるようにします。
- カバースリップ/ガラス底皿を大きなペトリ皿に反転させ、ベンチでタップして、ゲル表面に向かうビーズを強制的に押し付けます(図1D)。
- アルミニウム箔で覆い、湿度の高いチャンバーで室温で重合するために1時間放置します(すなわち、蒸発を防ぐために皿の上に濡れた組織を置きます)。
- 1時間後、カバースリップの放出を容易にするためにサンプルにPBSを加える。慎重に、針を使用してカバースリップを取り外します(異なるシランを塗布すると、ゲルからカバースリップを簡単に剥がす必要があります、図1E)。
- ゲルをPBSに残します。
注意:ゲルは4°Cで5〜7日間PBSに保存できるようになりましたが、48時間以内に使用することをお勧めします。
2. ゲルの機能化
- スルフォスクシニミジル6-(4'-アジド-2'-ニトロフェニルアミノ)ヘキサノエート(スルフォSANPAH)溶液を10 mM HEPESバッファーで0.5mg/mLで調製します。アルミ箔で覆われた4°Cで1週間まで保存できます。
- ゲルからPBSを吸引する。
- 室温でゲルにスルフォSANPAHの150 μLを加える(図1F)。
- ゲルを2分間UV処理に当て、スルフォSANPAHの部位を光活性化し、ゲル表面に貼り付けます。
- PBSで3回洗浄します(図1G)。
- ステップ 2.2 ~ 2.5 を繰り返します。
- 250 μL (100 μg/mL) を各ゲルに加え、アルミニウムホイルで覆ったまま、一晩4°Cの湿気の多いチャンバーで一晩インキュベートします(図1H)。
- HEL抗原を除去し、PBSで3回洗浄します。
注:HELは抗原として、そして接着分子として作用します。受容体に結合する他の分子(例えば、抗マウスIgM、ウシ血清アルブミン、オボアルブミン)またはインテグリンリガンド(例えば、LFA1に結合するICAM1)と混合することができる。必要に応じて、抗原抽出は、HELの蛍光版(タンパク質標識キットで分子を染色することによって得られる、ステップ4を参照)で観察することができる。バルクでの特定の濃度は、ガラス上と同じ表面濃度をゲルに得ない可能性があることに注意してください:直接比較が必要な場合は、二次染色で定量する必要があります。
3. 細胞のローディングとイメージング
- イメージングの前に、ゲルからPBSを取り出し、500 μLのB細胞培地(RPMI 1640、10%非補体胎児仔血清、1%ペニシリンストレプトマイシン、2%ピルビン酸ナトリウム、50uMメルカプトエタノールおよび1X非必須アミノ酸)を添加し、RTに平衡化させます。
- 細胞製剤
- 負の選択プロトコルに従って脾臓から原発B細胞を精製する(材料表を参照)。典型的な最終的なB細胞収量は1 x 107細胞の周りです。これをB細胞培地(RPMI-1640は10%の胎児の子牛血清、1%ペニシリン-ストレプトマイシン、0.1%メルカプトエタノールおよび2%ピルビン酸ナトリウムを補充する)に濃縮する。
- 必要に応じて、4°Cで最大6時間保存します。
- 画像取得前に37°Cで30分間セルを保持します。
- イメージング
- 熱および(おそらく)CO2 制御と共焦点顕微鏡を使用してください。
注意: 共焦点顕微鏡と回転ディスク顕微鏡のどちらを使用するかに関係なく、ピクセルサイズ<200 nmを使用して解析フェーズ(60x、NA 1.3など)でビーズを快適に追跡できる目的/ピンホールを使用することが重要です。蛍光顕微鏡も使用できますが、信号対ノイズ比が低く、個々のビーズの追跡が困難になる可能性があります。 - ビーズの2つの主要な層は、ゲルの底部と上部に表示されます。ゲル平面に焦点を当てます。
注:素敵なゲルは星空として現れ、ビーズはほぼ同じ平面上にほぼ均一に分布します。 - フレームレート5sで30分間の取得をプログラムします(これは実験のニーズに適応可能です、例えば、他の色を獲得し、zスタックを獲得するなど)
- ゲルから培地を吸引し、約200μLの培地をゲルに残します。ゲルを顕微鏡に置き、ビーズの表面層とゲル上の素敵な均一な領域を見つけます。
- 80 μLの細胞を加えます(焦点を維持するためにゲルに触れないようにしてください)。
- フォーカスが正しく、セルがエリア内で降下 (透過光の下) に表示されることを確認します。細胞がゲルに到達する前に取得を開始します。
- ゲル、振動、フォーカスドリフトとの偶発的な接触の場合は、フォーカスを調整します。
注:リラックスしたゲルの画像を収集することが重要であり、これはゲルに細胞が到着する前に撮影された任意の画像であることができます。
- 熱および(おそらく)CO2 制御と共焦点顕微鏡を使用してください。
4. 蛍光HEL抽出実験
- 蛍光色素(Alexa 555などのビーズとは異なる色)を結合して蛍光HELを調製し、 材料表を参照してください。
- ステップ2.7では、従来のHELを蛍光HELと交換する。
- 光の漂白を避けるために、低照度設定または低フレームレート(例えば、毎分2フレーム)で画像を取得します。
- HEL 抽出を定量化するには、次の式に従ってフレーム 0 の強度I(0) によって補正および正規化された各フレームI(t)のセル領域に積み込まれた強度を計算します。

注:フルオロフォアと共役した抗原は見えませんが(おそらくゲル表面でのフルオロフォアの焼入れによる)、ゲル上の存在は抗HELおよび蛍光二次抗体で確認できます。この蛍光体は、抗HELを塗布したカバースリップでゲルから剥がし、二次蛍光抗体(カバースリップ上)6で明らかにすることで、その蛍光性が実際に蛍光であることが確認できる。抽出された抗原のシグナルは非常に薄暗く、ビーズの漏出によってマスクされることもある。抗原抽出のみに関心がある場合は、ビーズを使わずにゲルを調製することをお勧めします(ステップ1.3.2および1.4.3を省略)。
5. 蛍光イメージング
- 野生型(例えば、ライフアクトGFPまたはミオシンII GFPマウスから)に対して行われた遺伝子組み換えマウスの脾臓からB細胞を精製することにより、蛍光B細胞を得る。
- 蛍光細胞のイメージングには、(可能であれば)水浸漬長距離40x-100x目的を有する回転ディスク顕微鏡を使用します。
- 露出時間とフレームレートを低く抑え、漂白を防ぐ。
注:Zの点広がり関数はゲルの存在によって非常に劣化するため、水浸しの目的を使用することをお勧めします。水浸漬目的を持つ生直立顕微鏡は、放出経路に(球状)細胞(および細胞核)が存在することによって誘導される強い球面収差に苦しんでいる。
6. 分析
注:データ分析は、一般的に、ドリフトのためにスタック全体を修正し、各フレーム内のビーズを見つけ、参照フレームに対するそれらの動きを追跡し(細胞の不在時に撮影)、変位場を補間し、フーリエ変換29を使用して応力を得る問題を反転させることによって行われます。このため、オンラインリポジトリ30からダウンロード可能なImageJマクロとMATLABプログラムの組み合わせを使用することをお勧めします。
- イメージのスタックとして ImageJ でムービーを開く
- マクロ "Crop_and_save.ijm" を実行します。
- 「長方形」ツールで対象地域(ROI)を選択し、't'キーを使用してROIリストに追加します。
- セルをトリミングする場合は、不動ビーズの領域を 5 ~ 10 ピクセル以上含める必要があります。境界または他のセルに近すぎるセルを分析から除外します。終了したら「OK」をクリックします。
- マクロはセルのマスクを提案します: これが満足のいく場合は「OK」をクリックします。問題がなければ、[OKでない]をクリックし、選択ツール(例:"フリーハンド"または"Oval")で閉じた領域を手動で選択し、[続行]をクリックします。
- MATLAB を開き、"TFM_v1.m" を実行します。
- 必要なパラメータを入力する:特に画像のプロパティ(ピクセルサイズ、取得時間間隔)とゲルのプロパティ(ヤングモジュラスE、ポアソン比)を確認してください。
- 参照イメージは、デフォルトで最初に設定されます。必要に応じて別のフレームに設定するか、外部ファイルをロードするには「0」に設定します。
- 元のファイルと同じディレクトリにソフトウェアの出力を見つけます(説明については、User_notice.pdfファイルを参照)。これには、ビーズの予備トラック(「FILENAME.fig」)、時間の経過に伴う収縮エネルギーのプロット(「FILENAME_energy.fig」)、セル(エネルギー、面積、瞬間など FILENAME_finaltable)に統合されたいくつかの量のテーブル(エネルギー、面積、瞬間など)、変位と力場を含む構造、ビーズの映画、変位場、ストレスとエネルギー(任意のaviリーダーで開くことができる)が含まれます。
注: 入力パラメータでは、「窓サイズ」は変位が補間される窓であり、従って応力と変位場の最終的な解像度です。これは、少数の(デフォルトでは4)ピクセルに設定されています。ビーズがない領域を補間して、人為的に解像度を上げるため、これを減らすことは推奨されません。
結果
細胞の大きさを考えると、相関技術(粒子画像のvelocimetryなど)を介してビーズの変位マップを抽出するアルゴリズムは、一般的にあまり正確ではない。しかし、必要な解像度の程度に応じて、無料のFiji / ImageJプラグイン31、32を使用して質的な結果を簡単に得ることができます。このアプローチは刺激と非刺激条件を比較するのに十分であるが、徹底的な分析のために我々は、ビーズを個別に追跡し、個々のビーズの変位33の補間として所定の時点で変位フィールドマップを提供するオンラインリポジトリ30からダウンロード可能なソフトウェアを使用することを推奨する。この時点で、いくつかの定量化が可能です。たとえば、(変位がゲル表面に対する応力接線によってのみ発生すると仮定することによって)、ソフトウェアは、その特定の変位マップを引き起こす各点での応力も提供します。これは「反転問題」の一種です:ある時点での変位は、他のすべてのポイントに適用されるすべての力の合計に依存します。「反転アルゴリズム」は、基板の物理的パラメータ(その剛性(ヤング率)とポアソン比を考慮に入れます。直接アルゴリズムは、通常、非常に正確ですが、計算にコストがかかります。フーリエ変換に基づくアルゴリズムは、私たちと同様に、本質的にフーリエ空間でのデコンボリューションを実行し、より効率的ですが、いくつかのエラーが発生する傾向があります(主に補間ステップによる)。これらのアルゴリズムは、一般的に、小さな局所(そして潜在的にアーティファクタル)変位が応力場の計算にあまりにも関連することを防ぐパラメータの調整を必要とします(Tikhonov正規化パラメータ8,29;ダイアログウィンドウの「正規化」変数。ここでは通常、5 x 10-19に設定します。より高度な解釈と分析(例えば、時空間相関、局所的な動き、蛍光チャネルとの相関)のために、我々は、この分野の専門家との協力を推奨します。計算方法のレビューについては、 シュワルツら 9を参照してください。
前述したように、正しいビーズ画像は「星空」のように見え、明るいスポットの均一でランダムな分布(図2A)。ビーズの数が少なすぎる場合(図2B)、または画像が焦点が合っていない場合(図2C)の場合、データと分析は信頼できません。B細胞がゲルの表面に落ち着くと、細胞の下のビーズは、ゲル上の細胞によって及ぶ牽引力のために動き始める。ビードが追跡できないフレームは破棄する必要があります。
チェックとして、「基準フレーム」を比較するビーズの動きを目で観察することができるが、典型的には、細胞の基板との最初の接触の前に1つ。概算結果は、図 3Aで行われているように、単一の粒子追跡(例えば、トラックメイト、フィジー 34)から得ることができます。解析では、参照画像内のビーズのセグメンテーション (「FILENAME.fig」)がコントロールとして提供されます。
我々が提案するソフトウェアを用いて、変位(図3B)および応力場(各画素における局所応力のベクトル及び変位場からの反転によって得られた各時間点、 図3C)を得ることができる。セルの面積に集積された変位および力場のスカラー積は、セルが基板上で発揮する総作業を提供する(図4A)。この計算には、プロトコルのステップ6.2で導入されたセルのマスクが必要です。
2つの生物学的条件(HEL対非活性化基質BSA、または野生型対ノックアウト)を比較する場合、平均曲線(図4B)または、さらに総合的に、エネルギーが高原に到達する最後の時間(20分)の平均値を計算することが有用である(図4C)。力の空間的情報が関連する場合、各条件の単一の時点を比較することが可能です(図4D)。より詳細な分析については、クマリら6 を参照してください。
蛍光抗原抽出時間の経過の一例は 図5A:シナプスで示された抗原がゲルから離れている場合の蛍光シグナルの進行性の出現を示す。15細胞を超える信頼区間(平均の標準誤差)を持つ平均抽出曲線を 図5Bに示します。

図1:ゲルの調製とその機能化を示す概略図。手順はプロトコルで説明されています。この図の大きなバージョンを表示するには、ここをクリックしてください。
{kind=link}

図2:異なる品質のビーズ画像の3つの例。 (B) ビーズの数が少なすぎる画像と、フォーカスが不十分な画像の例。この図の大きなバージョンを表示するには、ここをクリックしてください。
{kind=link}

図3:力場を抽出する画像の処理(A)ビーズの画像の例(白色の細胞の輪郭、透過画像から抽出)、時間t= 5分(赤いオーバーレイ)および変位(矢印)での時間t= 0分(スケールバー5μm)に対するビーズ追跡。(B)補間変位場(ベクトルの震えと大きさの地図として表され、矢印は変位[nm]に比例し、右側のカラーバーを参照)。底: マグニチュードのより滑らかなイメージ(バイキュービック関数による補間によって得られる)。(C)パネルBの変位場からの応力場(ベクトル震えと大きさマップとして表され、矢印はせん断応力[Pa]に比例し、右側のカラーバーを参照)、底: マグニチュードのより滑らかなイメージ(バイキュービック関数による補間によって得られる)。この図の大きなバージョンを表示するには、ここをクリックしてください。
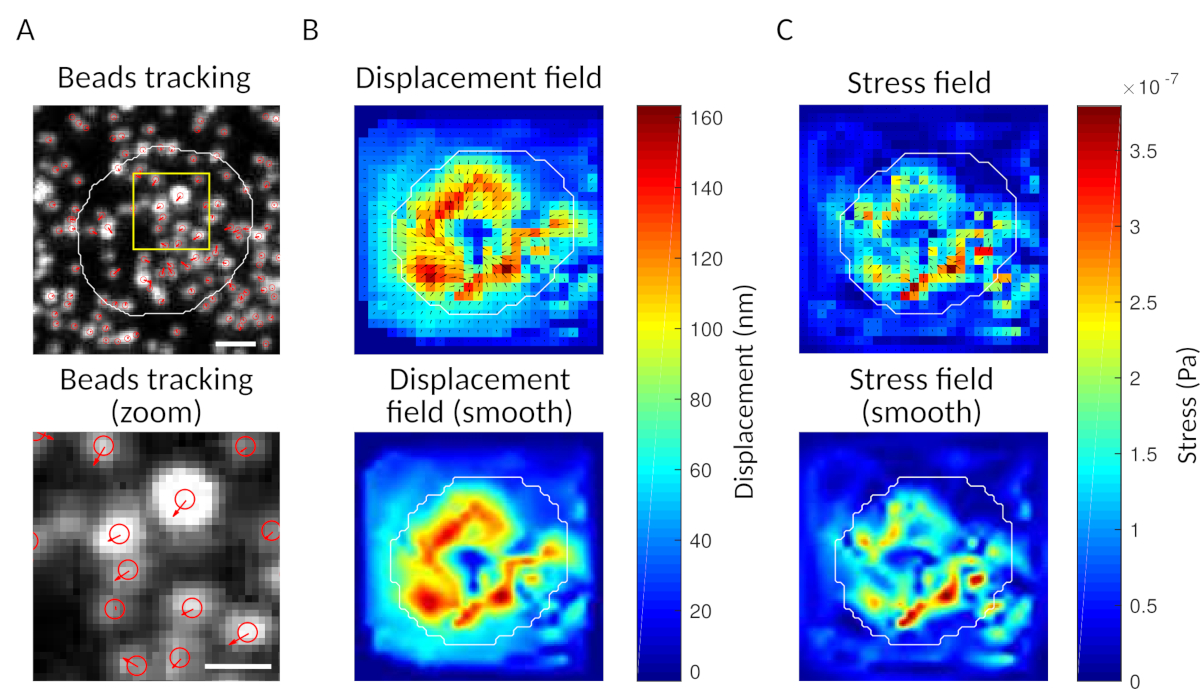{kind=link}

図4:力および変位場から抽出できる情報の例(A)単一細胞に対する時間内のエネルギーの進化の例:高原相(灰色で強調表示)は約10分後に現れる(B)平均エネルギー曲線の比較と(C)は、HEL(活性化)コーティングされたゲルおよび35細胞の相対的な高原レベルの比較(非活性化)コーティングされたゲル(±の異片間皮質)に対して有意性を示した。(D) HELおよびコントロールBSA状態のためのストレスのタイムラプスカラーマップ;大きさと震えのプロットの両方が表示されます。これらの画像は、クマリら 6. から適応されています。この図の大きなバージョンを表示するには、ここをクリックしてください。
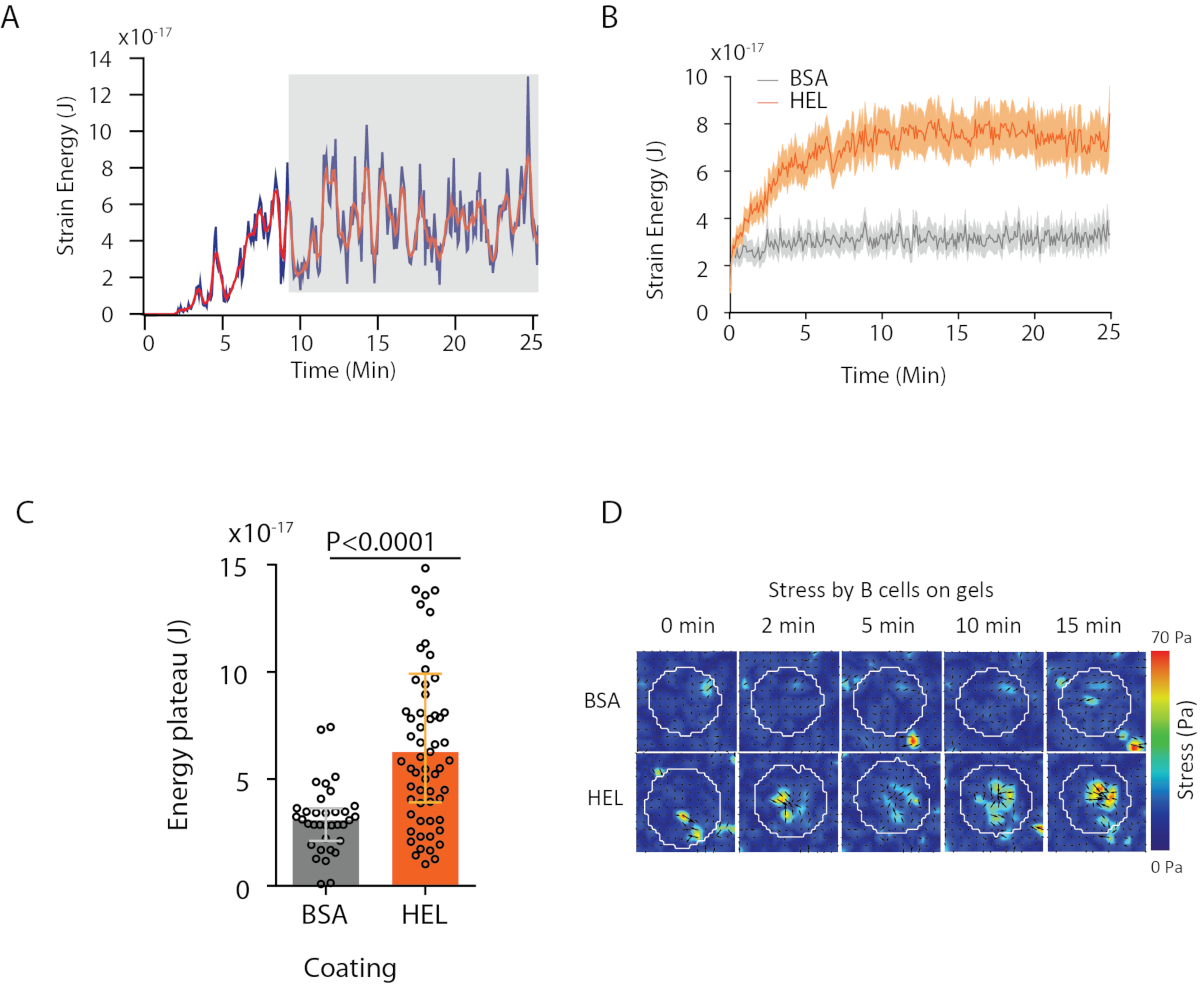{kind=link}

図5:蛍光抗原を用いた実験例(A)蛍光HELの抽出の経過時間(以下:最大のパーセンテージ、スケールバー=3μm)。(B) 時間経過に合った抗原の集合(平均±SEM、n = 15)。これらの画像は、クマリら 6. から適応されています。この図の大きなバージョンを表示するには、ここをクリックしてください。
{kind=link}
ディスカッション
ここで説明するTFM法は、B細胞の活性機械的能力の系統的研究を可能にする。B細胞の文脈では、これは抗原を抽出し、内在化する能力に関連している。他のTFM法と比較して、ここで提示されるプロトコルは単純で再現性が高い:剛性は、ガラス微小球のインデントによって測定され、Hertzモデルを使用して、400〜600 Pa.Bセル35 だけでなくTセル36にもうまく使用されている。ナノピラー(Tリンパ球37にも使用される)と比較して、平らな均質な表面を提供し、したがってゲルの相互作用が主に表面に接するように制約されるため、結果は解釈しやすくなる。
我々が述べたプロトコルは、抗原提示基質上のB細胞によって及ぶ力の時空間的ダイナミクスへのアクセスを与える。空間レベルでは、これは力の局在化の情報を提供し、蛍光顕微鏡と組み合わせて、実験者が特定の分子(すなわち、細胞骨格またはBCRシグナル伝達カスケードの成分)の存在と局所的な力を相関させることができます。時間レベルでは、量(総エネルギーや総応力など)を統合して、時間ポイントごとに1つの値を提供し、ノイズを低減することができます。これにより、時間の牽引力(成長および高原)の進化と脈動パターンの存在を観察することができます。
分析のための重要な実験的側面は以下のように説明される。(i)細胞密度:正しい分析を行うために、細胞は十分に分離されるべきである。セルの周囲に独自のサイズの空の領域がある場合、セルは分析可能であると考えています。(ii)透過画像:分析でマスクとして使用する実験中に細胞の少なくとも透過画像を収集することが推奨される。(iii) 画像内のビーズの数: シナプス中のビーズの数が30~200(すなわち、1~8~8ビーズ/μm²)の画像のみを分析することを推奨します。密度が低い場合、適切なマップの変位再構築はできません。高いビーズ密度は、単一の粒子追跡を信頼性の低いものにします。(iv) ビーズの数は実験中に一定である必要があります。ただし、イメージング条件の変動が小さいために変動が発生する可能性があります(特に、ビーズが互いに近すぎる場合)。フォーカスドリフトが発生した場合は修正する必要があり、問題のあるフレームは破棄する必要があります。(v) ゲルの品質: ひび割れが多すぎるゲル、ビーズの分布のばらつき、厚すぎるゲルは廃棄する必要があります。(vi) 細胞の種類によっては、繰り返し露光した後、遅い時点の細胞(>300フレーム)が光毒性の影響を受ける可能性があります。データと比較する「ベースライン」として、セルを欠いたマスク上でプログラムを実行することをお勧めします。これは、実験条件だけによるノイズレベルの大きさを提供します。
古典的な接着での牽引力を測定するために使用されるゲルは、焦点接着(アクチンの流れとシグナル伝達分子の採用)で起こるプロセスの調査を可能にする—力が38、39に加えられるポイント。しかし、シナプスの力は、焦点接着を介して適用されません。B細胞免疫シナプスにおける力発生の時空間パターンは、最近までこの方法を用いて定量的に調べられていない。TFMを用いて、我々は、最近の研究6で提示されたB細胞免疫シナプスにおける力パターニングを初めて観察し、リンパ球の研究における励ましの視点を開いた。
特に、この方法は、細胞がゲルに到達する前に撮影した画像を力計算の基準画像として採用している。通常のTFMプロトコルは、トリプシンで細胞を取り外した後、実験の最後に参照画像を撮ることを示唆しています。これにより、実験者はセルに含まれる領域を検索できます。これはここでも可能ですが、トリプシンは抗原コーティングされたゲルからB細胞を取り外す際にはかなり非効率的であり、剥離を長く待つ必要があり、ゲル修飾および動きのリスク(データセット全体を利用不能にする)が高くなります。
ここで提示される方法は柔軟であり、他のタンパク質をゲル表面に移植することができるので、免疫シナプスにおける他のシグナルの効果を研究するために適用することができる(例えば、インテグリンリガンドおよび免疫グロブリンが試験された)、さらには蛍光抗原(セクション4参照)。さらに、細胞は、薬物治療および局所摂動のために実験者にアクセス可能なままである。最後に、この方法は、固定細胞のイメージングにも対応しています。これらの観察のために、カバースリップにゲルを作り、細胞を汚し、スライドにカバースリップを接着し、取り付けメディアと別のカバースリップを追加することをお勧めします。その後、ゲルを通して画像の劣化を避けるために、上のゲルで観察が行われます。
考えられる落とし穴は、重合およびコーティングにおけるゲルの変動である。重合の問題は主にイニシエータ/触媒の品質によるものです。また、ゲルは、特に組み立て直後に使用しない場合に膨張する可能性があります。この問題はゲルの機械的特性に劇的な影響を与えないようですが、ビーズ層を目的に到達不能にして、ゲルを効果的に役に立たないようにすることができます。この問題が発生した場合は、条件ごとに余分なゲルを用意することをお勧めします。コーティングには一定のばらつきがあり、希釈したばかりのSulfo SANPAHを持つことは非常に重要です。
結論として、我々は、BCRリガンドによって活性化されたときの免疫シナプスでB細胞が及ぼす力を測定するための簡単で、安価で再現可能な方法を説明した。適切な受容体リガンドを用いて他のリガンドや他の種類のリンパ球(記憶B細胞、T細胞など)に対する反応を研究するように適合させることができる。
開示事項
著者らは開示するものは何もない。
謝辞
著者らは、M.ボルガー・マンロの批判的な読書に感謝し、ニコンイメージングCenter@CNRS-InstitutCurieとPICT-IBiSA、パリ、パリ、画像取得とキュリー動物施設のサポートのためにフランスバイオイメージング国家研究インフラのメンバーを認める。PP は CNRS によってサポートされていました。AKとJPは、パリ・デカルト博士課程のフェローシップとエコール・ドクターレFIRE(プログラム・ベッテンコート)によって支援されました。このプロジェクトは、PP(ANR-10-JCJC-1504-Immuphy)とAMLD(ANR-PoLyBex-12-BSV3-0014-001、ERC-ストラパセミ-GA 243103)への助成金によって資金提供されました。
資料
| Name | Company | Catalog Number | Comments |
| 3-aminopropyltrimethoxysilane (APTMS) | Sigma-Aldrich | 281778 | Store aliquoted, protected from humidity |
| 40% Acrylamide Solution | Biorad | 1610140 | |
| Alexa555 microscale protein labeling kit | Molecular Probes | A30007 | |
| Ammonium Persulfate (APS) | Sigma-Aldrich | A3678 | |
| B cell Isolation Kit, Mouse | Miltenyi Biotec | 130-090-862 | |
| B-mercaptoethanol | Gibco | 31350-010 | |
| 2% Bis Solution | Biorad | 161-0142 | |
| Bovine Serum Albumin (BSA) | Euromedex | 04-100-812-C | |
| Coverslip 18mm | VWR | 631-1580 | |
| Fetal calf serum | PAA | A15-151 | Decomplemented (40min @56°C) |
| Fluorodishes FD35 | World Precision Instruments, Inc | FD35100 | |
| Fluosphere: carboxylate-modified, 0.2um, dark red | Molecular Probes | F8807 | |
| Hen Egg Lysozyme | Sigma-Aldrich | L6876 | Stocked in aliquote 100mg/ml |
| MEM Non-Essential Amino Acids Solution (100X) | Thermofisher/Gibco | 11140035 | |
| N,N,N',N'-tetrametiletilendiammine (TEMED) | Euromedex | 50406-B | |
| PBS (Phosfate Buffer Saline) | Gibco | 10010-015 | |
| Penicillin–streptomycin | Gibco | 15140-010 | |
| RMPI 1640 – Glutamax I | Thermofisher | 61870-010 | |
| Sigmacote | Sigma-Aldrich | SL2 | |
| Sodium pyruvate | Gibco | 11360-039 | |
| sulfosuccinimidyl 6-(4'-azido-2'-nitrophenylamino)hexanoate (Sulfo-SANPAH) | Thermo Scientific | 22589 |
参考文献
- Yuseff, M. -. I., Pierobon, P., Reversat, A., Lennon-Duménil, A. -. M. How B cells capture, process and present antigens: a crucial role for cell polarity. Nature Reviews. Immunology. 13 (7), 475-486 (2013).
- Spillane, K. M., Tolar, P. B cell antigen extraction is regulated by physical properties of antigen-presenting cells. The Journal of Cell Biology. 216 (1), 217-230 (2017).
- Shaheen, S., Wan, Z., et al. Substrate stiffness governs the initiation of B cell activation by the concerted signaling of PKCβ and focal adhesion kinase. eLife. 6, (2017).
- Natkanski, E., et al. B cells use mechanical energy to discriminate antigen affinities. Science. 340 (6140), 1587-1590 (2013).
- Wan, Z., Chen, X., et al. The activation of IgM- or isotype-switched IgG- and IgE-BCR exhibits distinct mechanical force sensitivity and threshold. eLife. 4, (2015).
- Kumari, A., Pineau, J., et al. Actomyosin-driven force patterning controls endocytosis at the immune synapse. Nature Communications. 10 (1), 2870 (2019).
- Dembo, M., Wang, Y. L. Stresses at the cell-to-substrate interface during locomotion of fibroblasts. Biophysical Journal. 76 (4), 2307-2316 (1999).
- Sabass, B., Gardel, M. L., Waterman, C. M., Schwarz, U. S. High resolution traction force microscopy based on experimental and computational advances. Biophysical Journal. 94 (1), 207-220 (2008).
- Schwarz, U. S., Soiné, J. R. D. Traction force microscopy on soft elastic substrates: A guide to recent computational advances. Biochimica et Biophysica Acta. 1853 (11), 3095-3104 (2015).
- Colin-York, H., Shrestha, D., et al. Super-Resolved Traction Force Microscopy (STFM). Nano Letters. 16 (4), 2633-2638 (2016).
- Stubb, A., Laine, R. F., Guzmán, C., Henriques, R., Jacquemet, G., Ivaska, J. Fluctuation-Based Super-Resolution Traction Force Microscopy. BioRxiv. , (2019).
- Gutierrez, E., Tkachenko, E., et al. High refractive index silicone gels for simultaneous total internal reflection fluorescence and traction force microscopy of adherent cells. Plos One. 6 (9), 23807 (2011).
- Bergert, M., Lendenmann, T., et al. Confocal reference free traction force microscopy. Nature Communications. 7, 12814 (2016).
- Schoen, I., Hu, W., Klotzsch, E., Vogel, V. Probing cellular traction forces by micropillar arrays: contribution of substrate warping to pillar deflection. Nano Letters. 10 (5), 1823-1830 (2010).
- Colin-York, H., Fritzsche, M. The future of traction force microscopy. Current Opinion in Biomedical Engineering. 5, 1-5 (2018).
- Feng, Y., et al. Mechanosensing drives acuity of αβ T-cell recognition. Proceedings of the National Academy of Sciences of the United States of America. 114 (39), 8204-8213 (2017).
- Spillane, K. M., Tolar, P. DNA-Based Probes for Measuring Mechanical Forces in Cell-Cell Contacts: Application to B Cell Antigen Extraction from Immune Synapses. Methods in Molecular Biology. 1707, 69-80 (2018).
- Stabley, D. R., Jurchenko, C., Marshall, S. S., Salaita, K. S. Visualizing mechanical tension across membrane receptors with a fluorescent sensor. Nature Methods. 9 (1), 64-67 (2011).
- Merkel, R., Nassoy, P., Leung, A., Ritchie, K., Evans, E. Energy landscapes of receptor-ligand bonds explored with dynamic force spectroscopy. Nature. 397 (6714), 50-53 (1999).
- Hinterdorfer, P., Dufrêne, Y. F. Detection and localization of single molecular recognition events using atomic force microscopy. Nature Methods. 3 (5), 347-355 (2006).
- Sawicka, A., Babataheri, A., et al. Micropipette force probe to quantify single-cell force generation: application to T-cell activation. Molecular Biology of the Cell. 28 (23), 3229-3239 (2017).
- Desprat, N., Guiroy, A., Asnacios, A. Microplates-based rheometer for a single living cell. Review of Scientific Instruments. 77 (5), 055111 (2006).
- Labernadie, A., Bouissou, A., et al. Protrusion force microscopy reveals oscillatory force generation and mechanosensing activity of human macrophage podosomes. Nature Communications. 5, 5343 (2014).
- Bouissou, A., Proag, A., et al. Protrusion force microscopy: A method to quantify forces developed by cell protrusions. Journal of Visualized Experiments. (136), 57636 (2018).
- Kronenberg, N. M., Liehm, P., et al. Long-term imaging of cellular forces with high precision by elastic resonator interference stress microscopy. Nature Cell Biology. 19 (7), 864-872 (2017).
- Basu, R., Whitlock, B. M., et al. Cytotoxic T cells use mechanical force to potentiate target cell killing. Cell. 165 (1), 100-110 (2016).
- Bufi, N., Saitakis, M., et al. Human Primary Immune Cells Exhibit Distinct Mechanical Properties that Are Modified by Inflammation. Biophysical Journal. 108 (9), 2181-2190 (2015).
- Goodnow, C. C., Crosbie, J., et al. Altered immunoglobulin expression and functional silencing of self-reactive B lymphocytes in transgenic mice. Nature. 334 (6184), 676-682 (1988).
- Butler, J. P., Tolić-Nørrelykke, I. M., Fabry, B., Fredberg, J. J. Traction fields, moments, and strain energy that cells exert on their surroundings. American Journal of Physiology. Cell Physiology. 282 (3), 595-605 (2002).
- . MBPPlab/TFM_v1: Software for Time dependent Traction Force Microscopy Available from: https://github.com/MBPPlab/TFM_v1 (2019)
- Tseng, Q., Duchemin-Pelletier, E., et al. Spatial organization of the extracellular matrix regulates cell-cell junction positioning. Proceedings of the National Academy of Sciences of the United States of America. 109 (5), 1506-1511 (2012).
- . ImageJ plugins by Qingzong TSENG Available from: https://sites.google.com/site/qingzongtseng/ (2019)
- Plotnikov, S. V., Sabass, B., Schwarz, U. S., Waterman, C. M. High-resolution traction force microscopy. Methods in Cell Biology. 123, 367-394 (2014).
- Schindelin, J., Arganda-Carreras, I., et al. Fiji: an open-source platform for biological-image analysis. Nature Methods. 9 (7), 676-682 (2012).
- Wang, J., Lin, F., et al. Profiling the origin, dynamics, and function of traction force in B cell activation. Science Signaling. 11 (542), (2018).
- Hui, K. L., Balagopalan, L., Samelson, L. E., Upadhyaya, A. Cytoskeletal forces during signaling activation in Jurkat T-cells. Molecular Biology of the Cell. 26 (4), 685-695 (2015).
- Bashour, K. T., Gondarenko, A., et al. CD28 and CD3 have complementary roles in T-cell traction forces. Proceedings of the National Academy of Sciences of the United States of America. 111 (6), 2241-2246 (2014).
- Gardel, M. L., Sabass, B., Ji, L., Danuser, G., Schwarz, U. S., Waterman, C. M. Traction stress in focal adhesions correlates biphasically with actin retrograde flow speed. The Journal of Cell Biology. 183 (6), 999-1005 (2008).
- Stricker, J., Sabass, B., Schwarz, U. S., Gardel, M. L. Optimization of traction force microscopy for micron-sized focal adhesions. Journal of Physics. Condensed Matter. 22 (19), 194104 (2010).
転載および許可
このJoVE論文のテキスト又は図を再利用するための許可を申請します
許可を申請さらに記事を探す
This article has been published
Video Coming Soon
Copyright © 2023 MyJoVE Corporation. All rights reserved