
Method Article
大腸菌を用いた長期進化実験における毎日の移動、集団のアーカイブ、および適応度の測定
要約
このプロトコルでは、毎日の転送と定期的な凍結を実行して 大腸菌 長期進化実験(LTEE)を維持する方法と、進化した細菌の適応度の改善を測定するための競合アッセイの実施方法について説明します。これらの手順は、研究者が独自の微生物進化実験を開始するためのテンプレートとして役立ちます。
要約
長期進化実験(LTEE)は、35年以上にわたって77,000細菌世代にわたって単純な実験室環境に適応した 大腸菌 の12の集団を追跡しました。LTEEで使用されるセットアップと手順は、微生物の進化を研究するための信頼性が高く再現性のある方法を象徴しています。このプロトコルでは、まず、LTEE集団を新鮮な培地に移し、毎日培養する方法について説明します。次に、LTEE集団が汚染の兆候の可能性がないか定期的にチェックされ、アーカイブされて、後の研究のために恒久的な凍結された「化石記録」を提供する方法について説明します。これらの手順に含まれる複数の保護手段は、汚染を防ぎ、さまざまな問題が発生したときに検出し、実験の進行を著しく遅らせることなく中断から回復するように設計されています。LTEEで進化的変化の全体的なテンポと特徴を監視する1つの方法は、実験から集団と系統の競争力を測定することです。共培養競合アッセイの実施方法を説明し、結果から相対適応度を計算するためのスプレッドシートとRパッケージ(fitnessR)の両方を提供します。LTEEの過程で、一部の集団の行動は興味深い方法で変化し、全ゲノムシーケンスなどの新しいテクノロジーは、集団がどのように進化したかを調査するための追加の手段を提供しました。最後に、これらの変更に対応または利用するために、元のLTEE手順がどのように更新されたかについて説明します。このプロトコルは、進化と遺伝学、分子生物学、システム生物学、生態学の関係を研究するためのモデルシステムとしてLTEEを使用する研究者に役立ちます。より広義には、LTEEは、新しい微生物、環境、および質問で独自の進化実験を開始している人々のために、実証済みのテンプレートを提供します。
概要
1988年2月、リチャード・レンスキーは、カリフォルニア大学アーバイン校1で、定義されたグルコース制限増殖培地を含む12個のフラスコに大腸菌のクローン培養物を接種しました。翌日、彼は培養物の1%を各フラスコから新鮮な増殖培地を含む新しいフラスコのセットに移した。この1:100の希釈により、細菌集団は利用可能なグルコースを使い果たす前に100倍に拡大し、約62/3世代の細胞分裂に相当します。この手順は翌日繰り返され、それ以来毎日、数回の中断がありました。これらの毎日の転送は、実験が最初に1992年にミシガン州立大学に、次に2022年にテキサス大学オースティン校に移された後も続いています。その間ずっと、新しい突然変異がこれらの大腸菌集団に遺伝的変異を継続的に生み出し、自然淘汰は進化した細胞が祖先を凌駕する結果をもたらしました。
レンスキーは、進化のダイナミクスと再現性を調査するために、現在長期進化実験(LTEE)として知られているこの実験を設計しました。これらの質問に答えるために、彼は実験セットアップとそのプロトコルの設計にいくつかの重要な機能を含めました2。これらの特徴の1つは、モデル生物の慎重な選択でした。元の12の集団はすべて、直接の共通祖先である大腸菌B株REL606を共有する単一のコロニーから開始されました。この株が選ばれたのは、すでに実験室で一般的に使用されており、完全に無性生殖であり、プラスミドや無傷のプロファージを含まず3,4、その進化の研究をより簡単にするためです。実験を単純化した別の選択肢は、増殖後の各フラスコ内の細胞の密度を制限するために、増殖培地中の非常に低濃度のグルコースを使用することであった。低い細胞密度を使用することは、集団内の生態学的相互作用の進化の可能性を減らすことによって、集団適応度の変化の分析を容易にすることを目的としていました(例えば、交雑摂食による)5。
REL606は、araA遺伝子の点突然変異により、炭素およびエネルギー源(Ara−)としてʟ-アラビノースを使用することができません。LTEEを開始する前に、復元されたaraA配列を有する自然変異体(REL607)をREL6066から単離した。REL607は、ʟ-アラビノース(Ara+)上で増殖することができる。REL606はLTEE集団のうち6つを開始するために使用され、REL607は他の6つの集団を開始するために使用されました。アラビノースはLTEE中に使用される増殖培地には存在しないため、REL607はこれらの条件下でREL606と同じように動作します。しかし、テトラゾリウムアラビノース(TA)寒天培地に播種すると、Ara−細胞とAra+細胞はそれぞれ赤と白のコロニーを形成します。2つの祖先大腸菌株とその子孫を区別するこの方法は非常に有用である。LTEE集団間の相互汚染を検出するために使用できます。また、Ara-株または集団が互いに競合している場合のAra+株に対する適応度を測定するのにも役立ちます。適応度は、反対にマークされた競合他社の共培養を設定し、赤色と白色のコロニーの頻度(培養物の希釈液をTAプレート上に広げることによって得られる)が、競合他社が最初に混合されたときとLTEEと同じ条件下で1つ以上の成長サイクルの後の間でどのように変化するかを監視することによって測定されます。より適合した細胞タイプの表現は、各成長サイクル中に増加します。
LTEEのもう一つの重要な特徴は、進化する集団のサンプルが定期的にアーカイブされることです。グリセロールなどの凍結保護剤と混合すると、大腸菌細胞を凍結して後で復活させることができます7。LTEEプロトコルの一環として、75日ごと(約500世代に相当)に、新しいフラスコに移されなかった各集団の一部がグリセロールと混合され、複数のバイアルに分割され、冷凍庫に保存されます。この凍結された「化石記録」により、研究者はLTEEの最初の研究を行うことができ、さまざまな時点から進化した大腸菌集団を復活させ、祖先株と競合させて、適応度がどれだけ急速に増加しているかを追跡しました1。適応度の進化は、凍結された「化石記録」のより多くの「地層」が保存されるにつれて、定期的に再測定されてきました。これらの測定からの全体的な結論は、同じ環境で非常に多くの世代の進化の後でも、LTEEの適応度は今日まで改善し続けているということです8,9,10。
LTEEがこれほど長く続くことを可能にした理由は何ですか?元の質問と回答を可能にしたのと同じ機能の多くは、不運、人為的ミス、および世界の出来事による避けられない混乱に対する安全対策およびフェイルセーフとしても機能しています。毎日、培養物が新鮮な増殖培地に移されるとき、移管を行う研究者はAra−集団とAra+集団を交互に行います。次に、集団が凍結されたら、選択的および指示的寒天に播種して、「隣接する」集団が誤って相互汚染または混同されていないか(たとえば、白いコロニーが赤いコロニーのみを形成するはずの集団にある)、または外来微生物で汚染されていないか(たとえば、予期しないコロニーの形態や細胞密度)。人口が危険にさらされた場合、その祖先は冷凍庫から復活し、その場所に持ち越すことができます。したがって、Araマーカーと凍結アーカイブは、実験リソースと安全対策の両方として2つの目的を果たします。
その歴史は非常によく保存されており、簡単にアクセスできるため、LTEEサンプルは、実験の開始時には存在しなかった技術を使用して研究されています。例えば、LTEE集団における突然変異の動態を調べるために全ゲノムシーケンシングが使用され11,12,13,14,15、そしてトランスクリプトミクスおよびリボソームプロファイリングが遺伝子発現の変化を調べるために使用されている16,17。遺伝的ツールは、適応度およびさまざまな表現型への影響を理解するために、単一の突然変異またはいくつかの進化した突然変異の組み合わせによって異なる株を再構築するために使用されてきました18、19、20、21。凍結された「化石記録」のサンプルは簡単に補充されるため、実験履歴の一部または全部を他の研究所に出荷できます。LTEEサンプルは現在、南極大陸を除くすべての大陸に存在し、実験自体よりも若い研究者によって研究されています。その歴史的記録からのLTEEおよび進化した大腸菌サンプルおよび株の堅牢な方法は、他の質問および環境を調べる進化実験の出発点としても役立った22,23,24,25,26,27,28,29。

図1:LTEE手順の概要。 この図の拡大版を表示するには、ここをクリックしてください。
{kind=link}
ここでは、 大腸菌 長期進化実験で使用された3つのコアプロトコルを示します(図1)。(1)毎日の転送を実行する方法、(2)母集団サンプルとクローン分離株をアーカイブする方法、および(3)適応度の違いを測定するための共培養競合アッセイの実行および分析方法について説明します。これらのプロトコルがLTEEリソースの継続的な使用を促進し、新しい微生物進化実験の設計に役立つことを願っています。
プロトコル
1. LTEE人口の毎日の転送
注:12のLTEE集団は、前日のフラスコからの培養物の1%を新鮮な培地に接種することによって毎日移されます。このプロセスのステップを図 1 にまとめます。REL606株から開始された6つのAra−集団はA−1からA−6と指定され、REL607株から開始された6つのAra+集団はA+1からA+6と命名される。無菌技術と、集団を移送するためのスケジュールと順序を厳守することで、汚染やその他の混乱のリスクを最小限に抑えることができます。
- LTEE転送が行われる表面を、70%エタノールまたは10%漂白剤溶液で拭いて消毒します。ブンゼンバーナーに火をつけて局所的な上昇気流を作り、ガラス製品の炎上を可能にします。
注意: 汚染を防ぐために実験用手袋を着用してください。裸火の周りの安全のために、可燃性ではないニトリルのような材料で作られた手袋のみを使用することが重要です。 - オートクレーブで洗浄および滅菌された20 mLのホウケイ酸またはポリプロピレンビーカーで覆われた13個の50 mLホウケイ酸三角フラスコを準備します。フラスコに目に見える破片がないか確認し、完全にきれいでないものを交換します。
- 6つのフラスコに赤いマーカーを使用してA−1からA−6のラベルを付け、他の6つのフラスコに黒いマーカーを使用してA + 1からA + 6のラベルを付けます。最後に残ったフラスコに空白のラベルを付け、月/日の形式の日付と曜日を付けます。
- 滅菌済みの10 mL血清学的ピペットを使用して、13本のフラスコのそれぞれに9.9 mLのDM25培地を満たします。蓋として機能するビーカーを取り外した後、ビーカーを交換する前に、各フラスコの口に火をつけます。各フラスコを満たす間にピペットの先端を炎上させる。
注意: DM25の作成手順はオンラインで入手できます30.プラスチック製の血清学的ピペットを使用する場合は、プラスチックが溶けないように、先端を炎上させるか、炎の中での時間を制限してください。 - 前日のLTEEフラスコを振とうインキュベーターから取り出します。
- 各フラスコを光にかざして調べ、濁度と色を評価し、フラスコの完全性を確認し、異物の存在を探します。
注:肉眼では、すべてのAra-およびAra+培養物は、培地中のクエン酸塩の成長により、他の培養物よりも~10倍濁るA-3を除いて、ブランクと比較してわずかに濁っているように見えます。多くの外部の微生物汚染物質もクエン酸塩で増殖する可能性があるため、A-3以外の集団の濁度の増加は汚染を示している可能性があります。転送前の LTEE カルチャのイメージについては、「代表的な結果」セクションを参照してください。 - オプション:ブランク1 mLと各培養液1 mLを1 cmのプラスチックキュベットにピペッティングし、装置をブランキングした後、分光光度計を使用して600 nm(OD600)で光学濃度を読み取ることにより、各LTEE培養液に予想される濁度があることを確認します。
注:この追加の手順は、LTEEを初めて使用する研究者や、濁度を目で判断することに自信がない研究者や、疑わしい異常を文書化して調査する場合に役立ちます。OD600値が期待どおりである場合に増殖し続ける細胞集団を汚染するリスクを最小限に抑えるために、新しいフラスコへのその日の通常の転送(次の手順)を完了した後にのみ、前日のフラスコからOD600を測定するためのサンプルを採取します。LTEE カルチャの一般的な OD600 値については、「 代表的な結果 」セクションを参照してください。 - 滅菌フィルターチップ付きのP200マイクロピペッターを使用して、各LTEEフラスコから新鮮なDM25を含む対応するフラスコに100 μLの培養液を移します。A−1から始めて、A+1を転送します。その後、-母集団と+母集団を交互に続けます。どの培養物が移されたかを追跡するには、フラスコからピペッティングした後、フラスコを左に移動します。
注:Ara−とAra+ の集団間の移動と交替の厳密な順序は、相互汚染と取り違えの防止と検出 に役立ちます。厳格な無菌技術を守ってください:各転写に新しいピペットチップを使用し、キャップを外した直後とリキャップする前にフラスコの口に火をつけ、各転写の間に70%エタノールで湿らせた糸くずの出ない紙ワイプでマイクロピペッターのバレルとエジェクターを拭き取ります。微量でも培養物を殺す可能性があるため、マイクロピペッターの消毒には漂白剤を使用しないでください。 - 新しく接種したフラスコを37°Cで24±1時間インキュベートし、直径1インチで120 rpmの軌道振とうを行います。
- 前日からの培養物を4°Cで保存する。 これらのバックアップ カルチャを 2 日間保持します。この時点で3日前に4°Cで保存された古い培養物を破棄します。
注:前の2日間の培養物は、必要に応じて、問題や事故が発生した場合、または転送前に前日の培養物の汚染が発見された場合(たとえば、奇妙な着色や予期しない粒子状物質)に実験を再開するための2つの完全なバックアップセットを提供します。 - 転送ログノートブックに、転送を行った研究者の時間、日付、転送番号、名前またはイニシャル、培養に問題がないかどうか、およびその他の関連情報を入力します。(1)前日のブランクが汚染されている、(2)フラスコまたはその蓋にひびが入ったり壊れたりしている、(3)フラスコに異物が含まれている、(4)移送中にフラスコがひっくり返ったり落下したりした場合、または(5)これらのフラスコからの継続に疑問を投げかけるその他のイベントまたは観察がある場合、手順1.12-1.14に進みます。
- 前日のLTEE文化に問題、事故、または汚染の疑いがある場合は、それらから転送しないでください。代わりに、12個の培養物のセット全体を4°Cで保存して、後で検査してさらに特性評価します。
- 前日から移し替えたバックアップ培養液を入れたフラスコを取り出し、4°Cで保存した。 それらをベンチトップに置いて室温に温めます。各フラスコを静かに旋回させて、細胞を再懸濁します。
- バックアップフラスコから新鮮な培地を含む新しいフラスコのセットに移し、手順1.6-1.11の説明に従って通常どおり実験を続行します。バックアップ カルチャが使用されたことを転送ログにメモし、前日と同じ転送番号を記録します。
注:1つの母集団のフラスコにのみ問題が指摘されている場合でも、すべての集団で経過した世代数が同相に保たれるように、バックアップフラスコから12の集団すべてを転送します。4°Cで保管されたバックアップフラスコに汚染が認められた場合は、母集団サンプルの手順3.1〜3.2で説明されている手順を使用して、影響を受けるLTEE集団を凍結ストックから再開する必要があります。LTEE の転送番号は、復活後の DM25 で最初の培養が成長するまで増分しないでください。
2. LTEE 母集団のアーカイブ
注:LTEE集団のサンプルは、75回の転送ごとに凍結されます。個体群は、100倍の移入希釈後、毎日~6 2/3世代増加するため、この期間は~500世代に相当します。アーカイブ中、LTEE集団は、汚染をチェックするためにさまざまな種類の寒天培地にメッキされます。任意選択で、これらのプレートから代表的なクローンを選び出し、この時点でアーカイブすることができる。これらの手順を 図 1 にまとめます。
- 凍結予定の前日または数日前に、最小グルコース(MG)、最小アラビノース(MA)、テトラゾリウムアラビノース(TA)の3種類の寒天プレートを準備します。寒天の種類ごとに12枚のプレートに加えて、いくつかのエキストラを作ります。また、少なくとも250 mLの0.85%(w / v)滅菌生理食塩水と50 mLの80%(v / v)滅菌グリセロールを準備します。
注:すべてのメディアとソリューションのレシピはオンラインで入手できます30.LTEE が通常のアーカイブ スケジュールの 500 の倍数である世代に達する前日は、最後のフリーズダウンから 74日目 に、問題が検出または疑われるときに 4 °C のバックアップ フラスコからの転送によって追加された日数を加えた日です。 - オプション:LTEE集団からクローン分離株をアーカイブする場合は、追加の消耗品を準備します:各集団から3つのクローンを単離するには、72 MGプレート、80 mLの80%(v/v)グリセロール、および370 mLのDM1000が必要です。
- 計画された凍結の前日に毎日のLTEE転送のステップ1.2を実行するときは、12個のフラスコの追加セットを準備します。追加のフラスコの6つxA−1からxA−6に赤いマーカーを使用してラベルを付け、他の6つのxA + 1からxA + 6に黒いマーカーを使用してラベルを付けます。
注:「x」は、追加のフラスコセットがアーカイブに使用されることを示し、LTEEの毎日の転送を並行して継続するために使用される他のフラスコセットと区別します。 - 毎日のLTEE転送のステップ1.4を実行する場合は、アーカイブに使用する追加のフラスコに、25 mLの血清学的ピペットを使用して14.85 mLのDM25を満たします。
- 手順 1.5-1.11 の説明に従って、通常の LTEE 転送を完了します。次に、ステップ1.8の手順を繰り返しますが、今回は前日のLTEE培養液のそれぞれから150 μLを、アーカイブに使用する14.85 mLの新鮮なDM25の追加のフラスコに移します。
注意: この手順およびそれ以降のすべての手順では、これらのガイドラインに従って汚染や取り違えを避けてください。母集団A−1から始めて、次にA + 1を転送し、次に−母集団と+母集団を交互に続けます。集団を切り替えるときは、70%エタノールで湿らせた糸くずの出ないペーパーワイプでマイクロピペッターのバレルとエジェクターを拭き取ります。フラスコと試験管をトレイまたはラックからピペッティングした後、またはそれらとの間でピペッティングした後、フラスコと試験管を移動して、どの移送が完了したかを追跡します。 - ステップ1.9で説明したように、12本のフラスコのセットを37°Cで24±1時間、直径120 rpmの軌道振とうしながら、直径1インチのオービタル振とうしながらインキュベートします。
- 凍結当日にLTEE転送が実行される少なくとも1時間前に、LTEE集団をメッキするための物資を準備します。
- 12 個の MG、12 個の MA、および 12 個の TA 寒天プレートを選択します。それぞれを目視検査して、明らかな汚染がないことを確認します。
- 12のLTEE集団(A−1からA + 6)のそれぞれについて、各タイプのプレートの1つにラベルを付けます。
注意: プレートにラベルを付けるときは、シャーレの底の側面に書き込みます。これは、寒天の下からコロニーを調べたり写真を撮ったりするときにコロニーを覆い隠さないために重要です。これらは混同される可能性があるため、蓋に書かないでください。 - 寒天プレートを37°Cのインキュベーターに20分以上入れて温めてから、ステップ2.10で使用します。
- 9.9mLの生理食塩水を含む24本の試験管を準備します。それぞれ2本のチューブの12セットにそれらを配置します。
- プレートと同じ方法で12本の試験管の2セットのそれぞれにラベルを付け、LTEE母集団識別子の下に「1」または「2」を追加して、その母集団の希釈に使用する順序を指定します。
- 通常どおりLTEEの毎日の転送を継続するフラスコを使用して、手順1.1〜1.11を実行します。ステップ1.5の間に、アーカイブ用の余分な培養物を含む12個のフラスコも振とうインキュベーターから取り出します。
- 12個の追加のフラスコのそれぞれから培養液100μLをピペットでピペットし、そのLTEE集団について対の生理食塩水を第1試験管にアーカイブする。これらの100倍希釈液でチューブを完全にボルテックスします。次いで、それぞれ100μLを生理食塩水の対応する第2のチューブにピペットで入れる。最終的な10,000倍の培養希釈液を徹底的に渦流します。
- 10,000倍の培養希釈液を含む各チューブから80 μLをピペットで、その集団の標識されたTA、MG、およびMAプレートにピペットで送ります。好ましいものとして、滅菌散布棒または滅菌散布ビーズのいずれかを使用して、寒天表面全体に液体を均一に広げます。12種類の母集団すべてが3種類の培地すべてにメッキされるまで繰り返します。
- 必要に応じて、寒天に液体が見えなくなるまでプレートを乾燥させます。プレートを逆さまにして(寒天側を上にして)37°Cに設定した重力対流インキュベーターに入れます。
注意: プレートを逆さまにインキュベートすると、寒天が乾燥するのを防ぎ、結露が寒天表面に滴り落ちるのを防ぎます。インキュベーション中に寒天表面上の液体中の細胞が移動すると、コロニーが汚れ、コロニー数が不正確になる可能性があります。 - 3 mLの滅菌80%(v/v)グリセロールを、アーカイブ用に指定された12個の予備フラスコのそれぞれに加えます。渦巻き、穏やかにボルテックスして完全に混ぜます。
- 各フラスコから、サンプルの一意の識別子、サンプルが属するLTEE集団、凍結された世代、混合(母集団)サンプルであること、および日付でラベル付けされた滅菌クライオバイアルに混合物を分配します。6 mLを1つの大きなバイアルに、1.25 mLを6つの小さなバイアルのそれぞれにピペットで入れます。
注:大きなバイアルは作業ストックです。1つの小さなバイアルは、作業在庫が使い果たされたり汚染されたりした場合のバックアップです。他の5つの小さなバイアルは、他のラボに送ることができるコピーです。 - 充填したバイアルを-80°Cで凍結します。
- 24時間および48時間のインキュベーション後のTA、MG、およびMAプレート上のコロニーの成長と形態を調べて文書化します。
注:REL606およびREL607の祖先と、76,000世代で播種された12のLTEE集団のそれぞれによって形成されたコロニーの画像と説明については、「 代表的な結果 」セクションを参照してください。 - オプション: クローン分離株をアーカイブする場合は、以下のステップを実行します。
- MGプレートからLTEE集団ごとに3つのクローン分離株(コロニー)を選び、それぞれを新しいMGプレートに別々にストリークし、これらのプレートを37°Cで16〜24時間インキュベートします。
注:異なる形態のコロニーが存在する場合、LTEEの標準的な方法は、最初に最も一般的なタイプを選択し、次に少数派タイプからさらにコロニーを選択することにより、最大の多様性をサンプリングすることです。細胞を広げる前にシャーレの底の下側にドットをマークし、成長後に各マークに最も近い分離コロニーを選択することにより、ランダムサンプリング戦略を使用することもできます。 - 翌日、新しいMGプレート上の各プレートから代表的なコロニーをストリークし、これらのプレートを37°Cで16〜24時間インキュベートします。
- 翌日、各MGプレートから1つの単離されたコロニーを10mLの新鮮なDM1000を含むフラスコに接種する。また、追加のフラスコに10 mLのDM1000を充填し、培地汚染をテストするための未接種ブランクとして機能します。
- フラスコを37°Cで16〜24時間インキュベートし、直径1インチで120 rpmの軌道振とうを行います。
- インキュベーション後、2 mLの滅菌80%(v/v)グリセロールを各フラスコに加え、旋回させて混合します。
- 各フラスコから1.25 mLのアリコートを、各クローンの一意の識別子、LTEE集団、起源の世代、クローンサンプルであること、および日付でラベル付けされた小さな滅菌バイアルに分配します。
- 充填したバイアルを-80°Cで凍結します。
- MGプレートからLTEE集団ごとに3つのクローン分離株(コロニー)を選び、それぞれを新しいMGプレートに別々にストリークし、これらのプレートを37°Cで16〜24時間インキュベートします。
3.競争力のあるフィットネスアッセイ
注:LTEEでは、生殖適応度は、異なる細菌が毎日の移植と同じ条件下で1つ以上の24時間の培養サイクルにわたって達成する倍増の相対数の観点から定量化されます。具体的には、ある競合他社と別の競合他社の相対的な適応度は、共文化で直接競争するときの実現倍増率の比率です。ペアの各競合他社は、LTEEの凍結された「化石記録」の一部として以前にアーカイブされた完全な集団またはクローン分離株である可能性があります。あるいは、一方または両方の競合相手は、それらの効果をテストするために特定の突然変異を追加または削除するように遺伝子組み換えされたクローンであってもよい。2つの競合相手は、この遺伝子マーカーがこのアッセイ中にそれらを区別するために使用されるため、反対のAra+/Ara- 状態を持っている必要があります。競合アッセイの全体的なワークフローを 図2に示します。共培養フェーズの期間を1日から3日(またはそれ以上)に延長して、ほぼ均等に一致する競合他社間の差をテストする際の適応度推定の精度を向上させることができます。このプロトコルの他の重要な考慮事項と可能な変更については、 Disussion を参照してください。

図2:競合アッセイのフローチャート。 1日コンペティションアッセイの完全な手順を示します。3日間の手順は、1日目と2日目の代替経路を、1日の競技の1日目と同じ方法で3日目にメッキするまで続きます。 この図の拡大版を表示するには、ここをクリックしてください。
{kind=link}
- 備品を準備する
- 使用する競合のLTEE株および/または集団の数、および競合のペアごとに実行される反復競合アッセイの数を決定します。次の手順で説明するように、必要な消耗品を準備します。
注:すべてのメディアとソリューションのレシピはオンラインで入手できます30.競技実験のすべての日に必要なフラスコと試験管は、事前に、または使用する日に必要に応じて充填することができます。フラスコと試験管が事前に満たされている場合は、蒸発を最小限に抑えるために、暗所で室温で保管してください。TAプレートは、培養希釈液を広げることができるように、注がれた後に十分に乾燥できるように、使用する少なくとも2日前に準備する必要があります。ピペッティングミス、汚染されたプレート、またはその他の小さな事故が発生した場合に実験を続行できるように、常にいくつかの追加のフラスコ、試験管、およびTAプレートを準備してください。 - 復活の日(-2日目)には、20 mLビーカーで覆われた滅菌50 mL三角フラスコに、競合他社として使用する大腸菌の 菌 株または集団ごとに9.9 mLのDM1000またはリソジェニーブロス(LB)のいずれかを入れます。もう1つのフラスコに9.9 mLの同じ培地を満たし、未接種のブランクとして機能します。
- プレコンディショニング日(-1日目)には、1本の試験管に競技者ごとに9.9 mLの0.85%(w / v)滅菌生理食塩水を、2つのフラスコに2つの競合他社間の反復アッセイごとに9.9 mLのDM25を充填し、もう1つのフラスコに9.9 mLのDM25をブランク用に充填します。
- 競技開始日(0日目)には、1つのフラスコに9.9 mLのDM25を満たし、1つの試験管に9.9 mLの0.85%(w / v)滅菌生理食塩水を満たし、競技アッセイ複製ごとに1つのTAプレートを準備します。もう1つのフラスコに9.9mLのDM25を入れてブランクとして使用します。
- 代替案:最初の大会を過ぎた数日間の競技の日ごとに、競技ごとに9.9 mLのDM25を1つのフラスコに充填し、ブランク用に9.9 mLのDM25をもう1つのフラスコに充填します。
- 競技の最終日(例: 1日目または3日目)には、2本の試験管に9.9 mLの0.85%(w / v)滅菌生理食塩水を満たし、競技の複製ごとに1つのTAプレートを準備します。
- 使用する競合のLTEE株および/または集団の数、および競合のペアごとに実行される反復競合アッセイの数を決定します。次の手順で説明するように、必要な消耗品を準備します。
- -2日目:DM1000またはLBで競技者を個別に復活させる
- 競合他社ごとに、DM1000またはLBのいずれかの9.9 mLで満たされたフラスコにラベルを付けます。 同じ培地のバッチから9.9 mLで満たされた追加のフラスコにラベルを付けて、汚染をテストするための未接種ブランクとして機能します。
注:凍結ストックはLBまたはDM1000で復活し、凍結保存された細胞のより均一で予測可能な回収を実現します。凍結保護剤として使用されるグリセロールは 大腸菌によって代謝される可能性があり、サンプルがDM25で復活した場合、予想よりも高い細胞密度につながります。LBおよびDM1000は、この合併症が無視できるほど高い細胞密度への増殖をサポートします。 - 競合株の凍結ストックを含むクライオバイアルを-80°Cの冷凍庫から取り出します。バイアルを使用している間、バイアルをアイスバケットで冷やしてください。
- 各凍結ストックが解凍した後、それを徹底的にボルテックスして大腸 菌 細胞を再懸濁します。クローンを復活させる場合は、新鮮な培地を含むフラスコに12 μLの凍結ストックを接種します。集団を復活させる場合は、フラスコに120μLの凍結ストックを接種します。
注:120 μL容量の凍結ストックが集団に使用されるため、復活する細胞の数は、LTEE集団の1%が新しいフラスコに移されたときの毎日のボトルネックとほぼ同じになります。凍結ストックを複数回解凍およびボルテックスすると、細胞にストレスがかかり、時間の経過とともにストックの生存率が低下する可能性があります。特定のLTEE集団またはクローンを競技会で複数回使用する場合は、ストックの複数のコピーを再成長させて凍結し、1つのコピーが何度も解凍および再凍結されないようにすることをお勧めします。 - リバイバルフラスコとブランクを37°Cで一晩(16〜24時間)インキュベートし、直径1インチで120 rpmの軌道振とうします。
- 競合他社ごとに、DM1000またはLBのいずれかの9.9 mLで満たされたフラスコにラベルを付けます。 同じ培地のバッチから9.9 mLで満たされた追加のフラスコにラベルを付けて、汚染をテストするための未接種ブランクとして機能します。
- -1日目:DM25で競技者を個別に事前調整する
- 競合他社ごとに、9.9 mLの生理食塩水で満たされた試験管にラベルを付けます。2組の競合他社間の反復競合アッセイごとに、9.9 mLのDM25で満たされた2つの50 mLフラスコに、それぞれ複製番号と競合他社の1人の名前を記載したラベルを付けます。ブランクとして機能するように、9.9 mLのDM25で満たされた1つの追加のフラスコにラベルを付けます。
- 復活した競合他社の文化を含むフラスコをインキュベーターから取り出します。濁度を目で調べて、成長したこと、明らかな汚染がないことを確認します。
- 各フラスコからその競技者の生理食塩水の試験管に100μLをピペットで入れます。
注:このステップは培養物を100倍に希釈しますが、これはLBおよびDM1000の方がLTEEで使用されるDM25環境よりもはるかに高い細胞密度であるために必要です( 代表的な結果を参照)。 - 希釈培養物から100 μLを新鮮なDM25を入れたフラスコにピペッティングする直前に、各希釈チューブを完全にボルテックスします。これらのプレコンディショニングフラスコのうち2つを、各反復アッセイに2つ、各競合他社に1つずつ接種します。
- プレコンディショニングフラスコとブランクを37°Cで24±1時間インキュベートし、直径1インチにわたって120 rpmの軌道振とうを行います。
- 0日目:最初のカウントのために競技者とプレートを混合して競争を開始します
- 競合アッセイの複製ごとに、9.9 mLのDM25で満たされた1つのフラスコと、9.9 mLの生理食塩水で満たされた1つの試験管にラベルを付けます。フラスコとチューブに、競合他社の各ペアと競合アッセイの複製数を一意に識別する方法でラベルを付けます。ブランクとして機能するように、9.9 mLのDM25で満たされた1つの追加のフラスコにラベルを付けます。
- プレコンディショニングフラスコをインキュベーターから取り出します。濁度を目で調べて、成長したこと、明らかな汚染がないことを確認します。
- 50 μLのAra− 競技用フラスコを、新鮮なDM25で満たされた最初の反復競技用フラスコに移します。すぐに、50 μLのAra+ 競合他社を同じ競技用フラスコに移し、穏やかに旋回させて混合します。
- 競合他社のすべてのペアのすべての反復について、ステップ3.4.3を繰り返します。
注:競合フラスコには、DM25で増殖した 大腸菌 培養物が全体で100倍に希釈されており、LTEEでは毎日の移動後に経験するのと同じ条件の細胞です。転送とミキシングの実行順序は重要です。両方の競合他社を各フラスコにすぐに次々に追加して、どちらも新鮮な培地で成長し始めないようにします。たとえば、すべての競技用フラスコにAra− 培養物を追加せずに、戻ってすべてのAra+ 株を追加します。 - 新たに接種した各競技用フラスコから100 μLのピペットを、その競合アッセイ用に標識された生理食塩水の試験管に、これらの各チューブに合わせた事前調整されたDM25培養物の全体の10,000倍希釈が含まれるように複製します。
- 競技用フラスコとブランクを振とうインキュベーターに入れます。競技用フラスコを37°Cで24±1時間インキュベートし、直径1インチで120 rpmの軌道振とうを行います。
- 同日、競技用フラスコをインキュベーターに入れた直後に、ステップ3.4.5の各試験管を完全にボルテックスし、ステップ2.10で説明したように、これらの10,000倍希釈液80 μLをTAプレート上に広げました。各プレートの底面の側面に、混合された菌株のペア、反復数、および「Day 0」をラベル付けして、各競合他社の初期表現を決定するために使用されることを示します。
- TAプレートを重力対流インキュベーター内で逆さまにインキュベートし、Ara−とAra+の両方の競合物のコロニーが見えて区別できるようになるまで、37°Cでインキュベートします。一般に、これは16〜24時間以内に発生しますが、一部の進化した株ではさらに時間がかかる場合があります。各プレート上のAra−(赤)およびAra+(白)コロニーの数を数え、結果を記録します。
注:TAプレート上のAra- コロニーとAra+ コロニーの色の違いは、プレートを4°Cで保存しても時間の経過とともに明確でなくなるため、インキュベーターから取り出したらできるだけ早くカウントする必要があります。Ara− およびAra+ 細胞によって形成されたコロニーの典型的な外観を示すTAプレートの画像が 代表的な結果に含まれています。このセクションには、一般的な「エッジケース」コロニーの画像(たとえば、異なるコロニータイプの重複または成長)があり、それらを数える方法について説明します。競合他社のいずれかによってTAプレート上に形成されたコロニーの増殖速度と形態が以前に特徴付けられていない場合は、競合他社がまだ互いに分離している0日目に、生理食塩水で10,000倍希釈した80 μLを広げます。次に、37°Cで16〜24時間以上インキュベートした後、これらのコントロールプレート上のコロニーを調べます。
- 代替案:1日目と2日目:3日間の競技を続ける
- 競合アッセイの複製ごとに、9.9 mLのDM25で満たされた1つのフラスコに標識を付けます。競合他社の各ペア、反復数、および競合アッセイの日を一意に識別する方法でフラスコにラベルを付けます。ブランクとして機能するように、9.9 mLのDM25で満たされた1つの追加のフラスコにラベルを付けます。
- 前日からの競技用フラスコをインキュベーターから取り出します。濁度を目視で調べて、予想される成長を確認し、汚染を検出します。
- 各競技用フラスコから100 μLを、競技当日の新鮮な培地の対応するフラスコに移します。
- 新しい競技用フラスコとブランクを振とうインキュベーターに入れます。直径1インチにわたって120 rpmの軌道振とうしながら、37°Cで24±1時間インキュベートします。
- 続行する前に、コンテストの3.5.1日目に手順3.5.1〜3.5.4を繰り返します。
- 1日目または3日目:競技を終え、最終カウントのためにプレートを立てる
- 競技用フラスコごとに、9.9 mLの生理食塩水で満たされた2本の試験管を用意します。競合他社の各ペア、反復数、およびそれらが1回目または2回目の希釈用であるかどうかを一意に識別する方法でラベルを付けます。
- 競技用フラスコをインキュベーターから取り出します。濁度を目で調べて、それらが成長し、明らかな汚染がないことを検出します。
- 各競技用フラスコから100μLをその複製のための生理食塩水の最初のチューブにピペットで入れる。得られたチューブには、DM25培養物の100倍希釈液が含まれています。
- ボルテックス各100倍希釈チューブを十分に混合し、その複製のために生理食塩水の2番目のチューブに100μLをピペットで入れた。得られたチューブには、DM25培養物の10,000倍希釈液が含まれています。
- 10,000倍希釈液を含む各試験管を完全にボルテックスし、ステップ2.10で説明したようにTAプレート上に80μLを広げます。各プレートの底面の側面に、混合された菌株のペア、反復数、および1日の競技の場合は「1日目」、3日間の競技の場合は「3日目」のラベルを付けて、各競技者の最終的な表現を決定するために使用されます。
- TAプレートを37°Cでインキュベートし、ステップ3.4.8の説明に従って、成長後のAra−およびAra+ コロニーをカウントします。
注:すべての転写およびめっきステップを通じて、各競合アッセイの反復数を追跡します。異なる反復アッセイ間でどの最終カウントと初期カウントが対応するかを混乱させると、同じ2つの競合他社がそれぞれに混在している場合でも、適合度の推定値が正しくなくなります。
- 計算とプロットの適合度
- Excelを使用して相対適合度を計算およびプロットする場合は、XLSスプレッドシート(補足ファイル1)をダウンロードします。Rを使用している場合は、fitnessRパッケージ31 をインストールし、カンマ区切り値(CSV)テンプレート(補足ファイル2)をダウンロードするか、ビネットの指示に従ってこのファイルの新しいコピーを生成します。
- ダウンロードしたファイルの指定された細胞またはカラムで実施された競合アッセイの「転写希釈」100を入力します。競技者が共存培養された毎日の成長サイクルの総数を「転送数」として入力します(たとえば、3日間の競技の場合は3)。
- 指定されたセルまたは列に、参照株を「競合者1」、試験株または母集団を「競合菌2」として、競合者の各ペアの名前を入力します。
- コンペティションアッセイの複製ごとに、それぞれの初期コロニー数と最終コロニー数をダウンロードしたファイルの指定された列に入力します。
- Excelスプレッドシートを使用している場合、この推定値の平均相対適合度値と95%信頼限界が表示されるようになりました。競合他社のさまざまな組み合わせの結果を別のシートにコピーし、結果を要約したグラフを作成します。R を使用してデータを分析する場合は、fitnessR パッケージのビネットの指示に従ってこれらの計算を実行し、計算値を含む CSV ファイルを出力して、結果をプロットします。
結果
LTEE培養物の出現と濁度
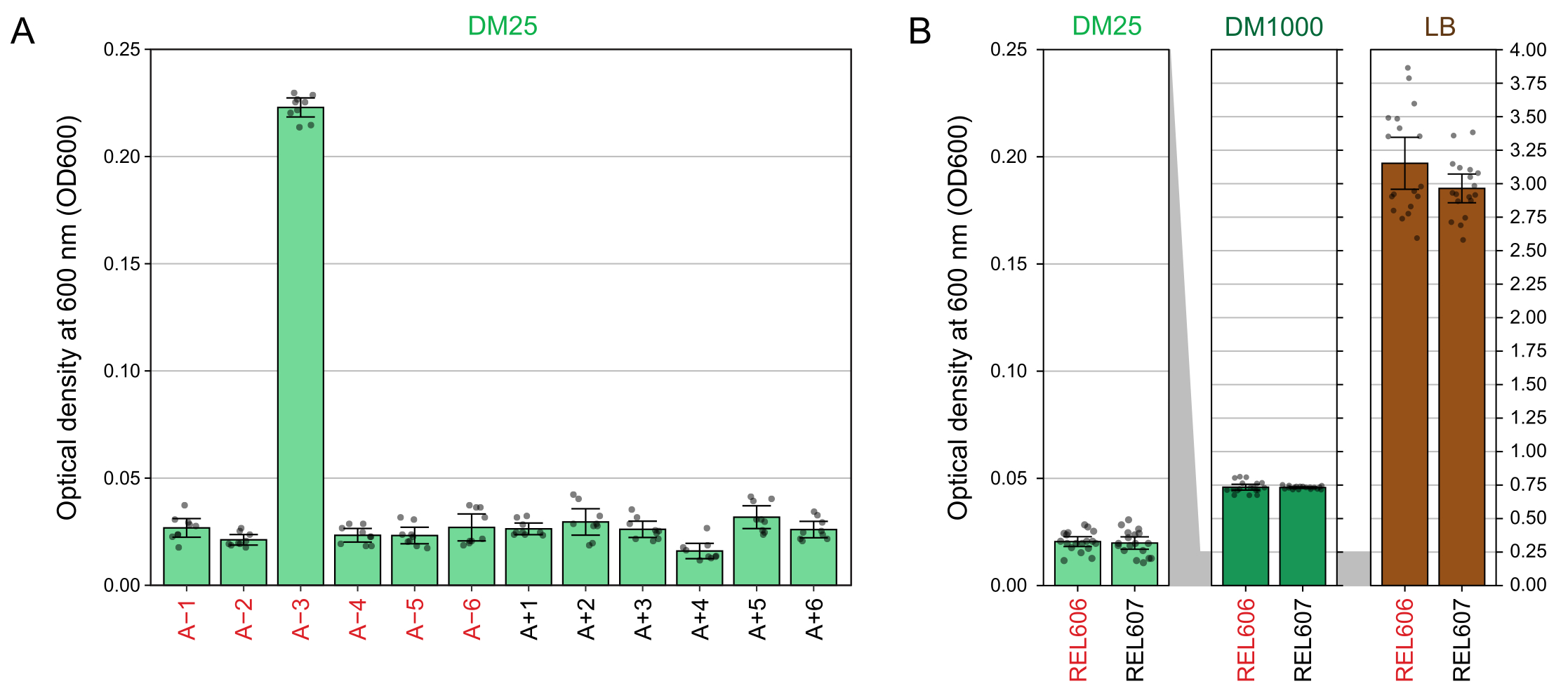
DM25のグルコース濃度が低いため、完全に増殖したLTEE集団の濁度は、12個のフラスコのうち11個でかろうじてしか見えません。LTEE培養物を目視で検査し、正常な増殖と汚染の兆候を確認する場合(ステップ1.6)、LTEE集団を含む各フラスコをブランクと並べて比較する必要があります(図3A)。例外は集団A−3であり、これはクエン酸塩を追加の炭素およびエネルギー源として使用するように進化したため、より高い細胞密度に達します32。REL606およびREL607祖先株のDM25培養物の濁度は、典型的な進化集団の濁度と同様です(図3B)。LTEE株および集団は、グルコースの濃度が高く、LBの濃度がはるかに高いため、DM1000ではより高い密度に成長します(図3B)。A−3 LTEE集団のDM25培養物の密度は、DM25およびDM1000におけるREL606の培養密度の中間である(図3C)。

図3:LTEE培養の外観 。 (A)実験が76,253 1/3世代に達した日のDM25の24時間の成長後の12のLTEE集団を含むフラスコは、ブランクの横に描かれています。(B)DM25、DM1000、およびLBで24時間増殖させたREL606およびREL607祖先の培養物を含むフラスコが、メディアブランクと一緒に描かれています。(C)DM25のA−3集団フラスコの濁度がDM25およびDM1000のREL606祖先とどのように比較されるかを示す同じフラスコの写真を並べてズームインしました。 この図の拡大版を表示するには、ここをクリックしてください。
{kind=link}
DM25(ステップ1.7)で増殖させた培養物の600 nm(OD600)での光学密度の分光光度計の読み取り値は、LTEE集団(図4A)とその祖先(図4B)の両方について、これらの目視観察と一致します。これらの測定値は、汚染やミスが疑われる場合の成長を定量的に比較して文書化するために使用できます。76,000〜76,500世代のLTEE集団の測定では、クエン酸塩で成長するように進化した集団であるA−3のOD600は平均0.223であることがわかりました(0.218-0.227、95%信頼区間)。他の11集団のOD600は平均0.0252であった(0.0239-0.0265、95%信頼区間)。11人の正常集団の間でOD600測定値にわずかではあるが有意なばらつきがあった(F10,88 = 5.1035、 p = 7.5×10−6)。LTEE集団は、約5〜6時間のインキュベーション後に定常期に達します。それらが午前中に移された場合、成長は同じ日の午後半ばから遅くまでに見られます。微生物の多くの種はクエン酸塩で好気的に成長することができます。したがって、A-3以外の集団における濁度の増加は、外部汚染の兆候である可能性があります。

図4:LTEE培養の濁度 。 (A)実験の76,000〜76,500世代の間の3つの異なる日の24時間成長サイクル後の12のLTEE集団の600nm(OD600)での光学密度。3つの異なる日のそれぞれにおける3つの1 mLアリコートのOD600値がポイントとしてプロットされます。同じ日からのブランクの3つの異なるアリコートの平均OD600値をこれらの値から差し引いた。塗りつぶされたバーは平均を示します。エラーバーは95%信頼限界です。(B)DM25、DM1000、およびLBにおけるREL606およびREL607祖先の培養物のOD600。各条件および菌株について、2つの異なる培養の3つの異なる日のそれぞれにおける3つの1-mLアリコートのOD600値がポイントとしてプロットされます。同じ日からのブランクの3つの異なるアリコートの平均OD600値をこれらの値から差し引いた。塗りつぶされたバーは平均を示し、エラーバーは95%信頼限界です。パネル間の灰色の影付きの領域は、DM25パネルとDM1000およびLBパネルの間でOD600軸がどのように再スケーリングされるかを示しています。 この図の拡大版を表示するには、ここをクリックしてください。
{kind=link}
LTEEコロニーの成長と形態
異なる培地にプレーティングして集団の汚染をチェックすると(ステップ2.15)、REL606とREL607の祖先とすべての進化した集団は、最小グルコース(MG)寒天プレート上に半透明でやや不規則なエッジを持つ白いコロニーを形成します(図5A)。MG寒天の組成は、グルコースの濃度が高いことを除いて、毎日のLTEE転送に使用されるDM25の組成と同じであるため、進化したLTEE集団は、祖先よりもMG上に大きなコロニーを形成することがよくあります。DM25の細胞密度が高いため、A−3集団は、他の集団と同じ体積で播種すると数倍多くのコロニーを持ち、コロニーのサイズが制限される可能性があります。最も一般的なタイプの汚染微生物は、MG上に真っ白で不透明で完全に円形のコロニーを形成します。
最小アラビノース(MA)寒天培地では、REL607の祖先とAra+ 集団は通常、すべてわずかに半透明の白いコロニーを形成します。この典型的な成長パターンは、アラビノースで成長の欠陥を進化させ、MAでコロニーを形成しなくなったA+6を除いて、76,000世代にわたってAra+ 集団で持続しています(図5B)。DM25でのLTEE転移中にアラビノースで増殖を維持する選択はないため、実験が続くにつれて、他のAra+ 集団も最終的にMA寒天プレート上でコロニーの形成を停止する可能性があります。A−3を除いて、Ara− 集団はMA寒天上にコロニーを形成しないが、綿密な検査により、寒天中の微量栄養素のために微小コロニーが明らかになる可能性がある。A−3集団は、これらの細胞がこの培地にも存在するクエン酸塩上で増殖することができるため、MA上に多数の小さなコロニーを形成する。MA上の汚染物質コロニーはまれです。

図5:汚染を検出するためのLTEE集団のめっき。 実験が76,026 2/3世代に達した日のREL606およびREL607祖先および12のLTEE集団の希釈液を、(A)MG、(B)MA、および(C)TA寒天プレートに播種し、24時間および48時間後に写真撮影した。すべての培養物に対して同じ希釈が行われたが、LTEE集団のプロトコルに記載されている量の半分の量が祖先のために播種され、それらのより高い細胞密度をいくらか説明した。 この図の拡大版を表示するには、ここをクリックしてください。
{kind=link}
テトラゾリウムアラビノース(TA)寒天培地では、REL606祖先とすべてのAra-集団が赤色のコロニーを形成すると予想され、REL607祖先とすべてのAra+集団は一般に白色のコロニーを形成するはずです(淡いピンク色または桃色の色合いを含む)(図5C)。LTEEの祖先は堅牢なコロニーを形成し、16〜24時間以内にTA寒天上でAra−およびAra+として容易に識別できます。もともと、この違いは、Ara−集団とAra+集団の間の相互汚染を検出するために使用できました。しかし、TA寒天培地は、毎日の移入に使用される化学的に定義されたDM25培地よりも複雑な栄養組成を有しており、LTEEの大腸菌がこれらの条件下で頑強に増殖する能力を維持するための進化的圧力はありませんでした。その結果、一部の進化したLTEE集団は現在、TAプレート上での成長が悪く、コロニーを形成するのに48時間かかるか、まったく確実に成長していません。進化したLTEE集団によってTA上に形成されたコロニーの色と形態も、祖先と比較して変化し、互いに分岐しています。いくつかの異常なコロニーの存在は、必ずしも汚染の兆候ではありません。アラビノースの利用に影響を与える機能喪失変異の可能性が高いため、特にAra+からAra-にLTEE株のAraマーカー状態を切り替える自然突然変異が、araA活性を回復する復帰変異よりも発生する可能性があります。Araマーカーの状態を切り替える突然変異は、超突然変異(A−1、A−2、A−3、A−4、A+3、およびA+6)を進化させた集団でより一般的です13。TA寒天培地では、他の種の汚染微生物は、LTEE株や集団によって形成されたものとは異なり、明確な白い境界で囲まれた赤い中心を持つ小さな完全に円形のコロニーを形成することがよくあります(常にではありません)。
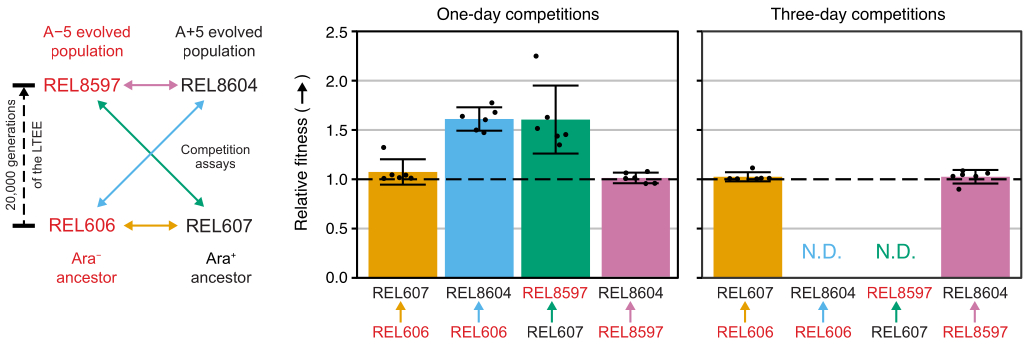
共文化コンペティション結果
2つのLTEE祖先(それぞれREL606とREL607)のすべてのAra−とAra+のペアと、20,000世代でアーカイブされたA−5とA + 5の集団サンプル(それぞれREL8597とREL8604)の間の競争は、異なるAraマーカー状態を持つコロニーをTA寒天で区別してカウントする方法を示しています(ステップ3.4.8と3.6.6)(図6).コロニーを、DM1000での復活から始まった1日および3日間のアッセイの前後に、競合他社の各ペアについて6つの複製フラスコについてカウントした(表1)。進化したLTEE集団の培養物はDM25の祖先株の培養物よりも低い細胞密度に達するため、同じ希釈率と体積で播種されたコロニーの総数は、競合物が混合されたコロニーの総数によって異なります。この違いは、実験の最初の数千世代の間にすべてのLTEE集団で発生した細胞サイズの増加の進化の結果です8,33。

図6:TA寒天プレートにプレーティングした競合アッセイ。 競合アッセイからのTA寒天プレートの例。REL606とREL607は、それぞれLTEEのAra− とAra+ の祖先です。REL8597とREL8604は、LTEEの凍結された「化石記録」から、それぞれ20,000世代のA−5とA + 5の集団です。株の各ペア間の1回の反復アッセイに対応するTAプレートが、競技の0日目、1日目、および3日目について示されています。プレートは、37°Cで24時間の成長後に撮影されました。 REL606およびREL8597の競合細胞の細胞はAra− であり、赤いコロニーを形成する。REL607およびREL8604の競合の細胞はAra+ であり、白いコロニーを形成します。 この図の拡大版を表示するには、ここをクリックしてください。
{kind=link}
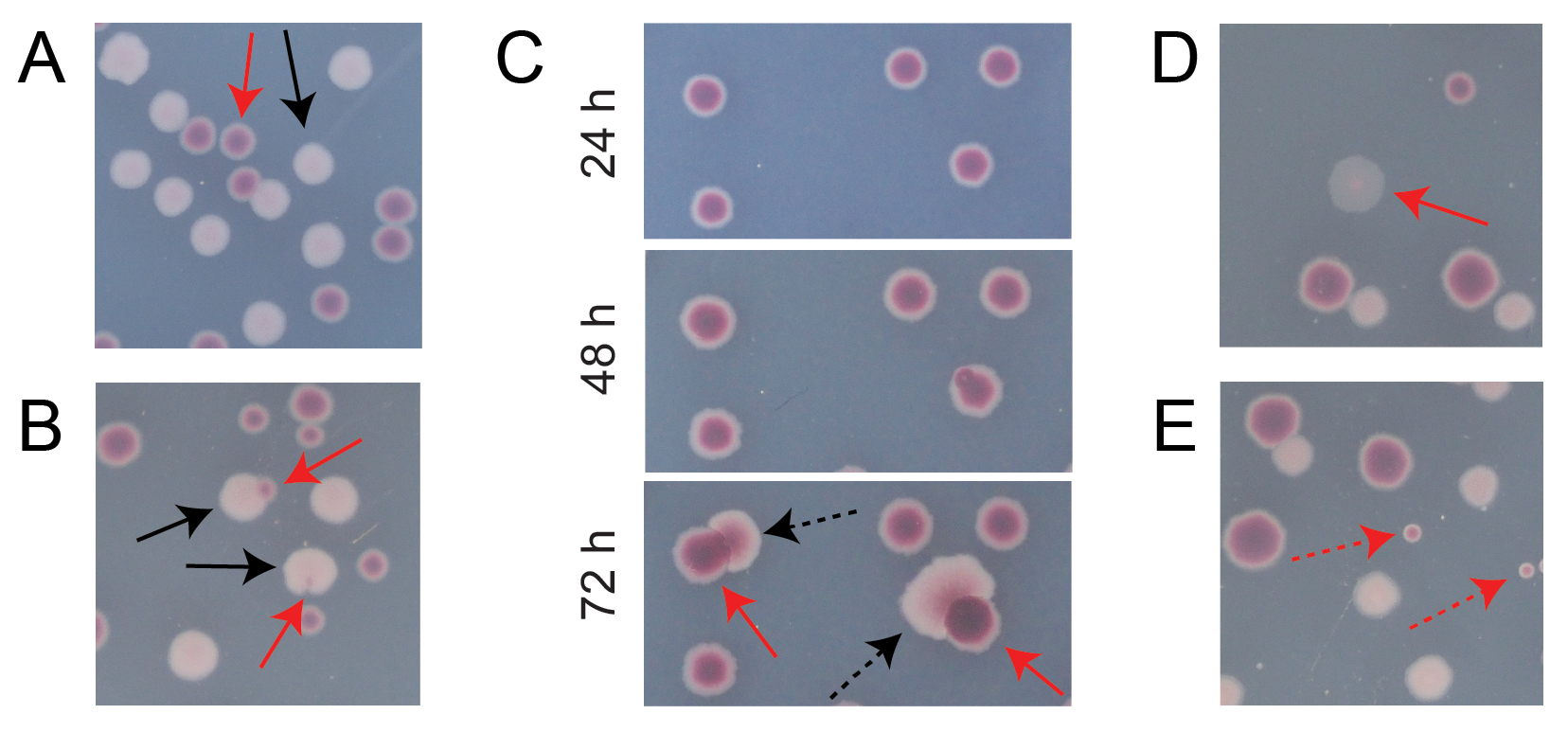
典型的な競合TAプレート上のほとんどのコロニーは、異なるタイプの最初は円形のコロニーが一緒に成長した数を数えやすい方法で、十分に分離または重なり合っています(図7A)。しかしながら、2つの色の混合である異型のコロニーまたは成長を数える方法が明らかでない状況が生じるかもしれない。まず、白いAra+コロニーと赤いAra-コロニーが重なると、Ara+コロニーが繁茂してAra-コロニーを包む傾向があります。この状況では、大きなAra+コロニーの小さな赤いパッチまたは半透明の「ギャップ」をAra-コロニーとして数える必要があります(図7B)。第二に、自発的なAra+変異体がAra−コロニーで時折発生します。これらの変異体は通常、追加の栄養素としてアラビノースにアクセスするとより速く成長するため、赤いコロニーの内部からより迅速に広がる白いセクター(乳頭)として現れます(図7C)。これらの白人セクタのコロニーは、1つのAra-コロニーとしてカウントされ、Ara+コロニーはカウントされません。この状況は、プレートを48時間以上インキュベートするとより一般的になります。第三に、時には半透明のピンクがかったコロニーが観察される(図7D)。これらはAra-競合他社によって形成されます。最後に、寒天の調製中、または培養希釈液を表面に広げるときに、内部がわずかに異なる赤の色合いを持つ少数の円形コロニーがTAプレート上に成長することがあります(図7E)。これらの汚染物質コロニーはカウントされるべきではありません。TAプレートのいずれかに多くの異型コロニーがあるために競合培養の汚染が疑われる場合は、その複製を除外する必要があります。

図7:TA寒天培地上のAra−およびAra+コロニーをカウントする際に遭遇するエッジケース。各パネルでは、カウントされるべきいくつかのAra−およびAra+コロニーが、それぞれ赤と黒の実線の矢印でマークされています。カウントすべきでないコロニーは、表示されるタイプに対応する破線の矢印で示されます。全ての写真は、パネルCを除いて24時間のインキュベーション後に撮影した。 (A)正常なAra−およびAra+コロニーの例。(B)Ara+コロニーがAra−コロニーの近くに繁茂している例で、白いコロニーの外側に透明な隙間としてかろうじて見えるものを含む。これらの各ケースを、各タイプに1つずつ、2つのコロニーとして数えます。(C)Ara+変異株を生むAra−コロニーの例。各ケースを単一のAra−コロニーとしてのみ数えます。発生する白いセクター(乳頭)は、コロニー内で発生するAra+変異体によるものです。コロニーの同じフィールドは、24時間、48時間、および72時間の成長に続いて示されています。(D)半透明のピンク色のコロニーの例。それをアラ−として数えます。(e)大腸菌ではない微生物による外部汚染により形成されたコロニーの例。これらは赤ですが、小さく、完全に円形で、明確な白い境界があります。この図の拡大版を表示するには、ここをクリックしてください。
{kind=link}
Excelスプレッドシート(補足ファイル1)を使用するか、CSVテンプレートに入力されたコロニー数(補足ファイル2)に対してRのfitnessRパッケージ関数を実行して、これらの競技会のコロニー数を分析すると、2つの祖先はアッセイの精度の範囲内で適応度の点で区別がつかず、20,000世代のA-5とA + 5の両方の集団が祖先よりも有意に適合していることがわかります。 そして、どちらの進化した集団も他の集団よりも有意に適合していないこと(ウェルチの t検定、 p > 0.05)(図8)。相対適応度推定の精度は、密接に一致したペアの1つ(REL606対REL607)の1日競技と比較して、3日間の競技で向上します。これらの測定の精度は、必要に応じて、より多くの成長サイクルでより長い競争を実施することにより、さらに向上させることができます。ただし、数日間の競技の結果は、一方の競技者が他方の競技者に比べて非常に豊富になり、不適合なタイプのコロニーがほとんどまたはまったくないため、2つの株の比率を正確に決定できなくなると有益ではありません。これは、進化した20,000世代の集団に対する祖先の3日間の競争(REL606対REL8604およびREL607対REL8597)の場合です(図6 および 表1)。
表1:競合フィットネスアッセイからのコロニー数。 6回の反復による1日および3日間の競合アッセイを、2つのAra− および2つのAra+ 競合体のすべてのペアワイズ組み合わせについて実施した。REL606とREL607は、それぞれLTEEのAra− とAra+ の祖先です。REL8597とREL8604は、LTEEの凍結された「化石記録」から、それぞれ20,000世代のA−5とA + 5の集団です。 この表をダウンロードするには、ここをクリックしてください。

図8:競合アッセイを使用して測定された相対適応度。 LTEEの祖先と20,000世代のA−5およびA+5 LTEE集団との間の1日間および3日間の競合アッセイの結果。左の図は、4つのペアワイズ競技を色分けされた両方向矢印で示しています。2つのAra− (赤いラベル)と2つのAra+( 黒いラベル)の競合他社の各組み合わせを6倍の複製でテストしました。 表1 のコロニー数をfitnessRパッケージ31を用いてRで分析し、結果をggplot2パッケージ(バージョン3.4.0)34を用いてプロットした。フィットネスは、ラベルの矢印が矢印の元の競技者に対して向かっている競技者として表示されます(たとえば、REL8604とREL606)。各競合アッセイ反復のコロニーカウントから推定された相対適応度値(点)、競合相手のペアの平均相対適応度値(図と同じ色分けで塗りつぶされたバー)、および95%信頼区間(エラーバー)が示されています。3日目のプレートには祖先のコロニーがゼロまたは非常に少なかったため、祖先と進化した集団との間の3日間の競争の相対適応度値(ND)を決定できませんでした( 表1を参照)。 この図の拡大版を表示するには、ここをクリックしてください。
{kind=link}
補足ファイル 1.相対的な適応度を計算するためのExcelスプレッドシートファイル。このファイルをダウンロードするには、ここをクリックしてください。
補足ファイル 2.fitnessR パッケージを使用して R の相対適合度を計算するためのカンマ区切り値入力ファイルテンプレート。このファイルをダウンロードするには、ここをクリックしてください。
ディスカッション
LTEEの長期的なレジリエンスとその手法
大腸菌長期進化実験(LTEE)は、現在4年目を迎えています。どのような期間の微生物進化実験でも、再現性のある環境を維持し、汚染を回避し、サンプルをアーカイブし、フィットネス度を正確に測定することが重要です。LTEEは、均質な環境を作り出すよく振とうフラスコの使用や、低い細胞密度をサポートする化学的に定義された増殖培地の使用など、これらの目的を達成するためのいくつかの実績のある戦略を示しています。さらに、LTEEは、進化環境において容易にスクリーニングされ、選択的に中立である表現型(コロニーの色)を与える遺伝子マーカーが異なる祖先株を採用しています。この実験計画機能は、内部および外部の汚染を識別する手段を提供し、フィットネスの測定を容易にします。ただし、1988年以降LTEEで使用されているすべての手順と保護手段が同等に堅牢であることが証明されているわけではありません。LTEEが始まったときに信頼できたいくつかの方法は、大腸菌集団が進化するにつれて効果が低下しました。幸いなことに、これらの問題のある方法は、実験の開始以来開発された技術を使用して拡張または置き換えることができます。
汚染の検出
汚染の検出はLTEEにとって重要です。汚染には、LTEE集団間(相互汚染)と環境からの微生物(外部汚染)の2種類があります。ほとんどの場合、無菌技術を慎重に使用し、培地調製および毎日の移送中に細心の注意を払うことで、両方のタイプの汚染を防ぐことができますが、それらは起こります。実験の早い段階で、TA寒天培地へのめっきは、Ara−集団とAra+集団の間で常に交互に起こるため、交差汚染の事例を検出するために使用できた。特定のバクテリオファージに対するこれらの大腸菌の感度と耐性のフィンガープリントは、LTEE集団を、それらを汚染する可能性のある一般的に使用される大腸菌実験室株と区別できる設計上の特徴でもあることを意図していました4。しかし、これらの遺伝子マーカーは、実験が進むにつれて信頼できなくなっています(たとえば、一部の集団はTA寒天培地上にコロニーを形成しなくなりました)10,35。幸いなことに、実験中に別々の進化の歴史を経験したため、集団は遺伝的に分岐し、交差汚染の検出に使用できる新しい遺伝子マーカーを作成しました。たとえば、各集団は、pykF遺伝子とnadR遺伝子の突然変異の独自の組み合わせを進化させました14、36、37。私たちは時々、PCR増幅とサンガー配列決定を行い、異常な形態または色のコロニーが交差汚染によるものであるかどうかをテストします。全ゲノムおよび全集団のシーケンシングのコストが下がり続けるにつれて、LTEE集団の定期的なシーケンシングがまもなく可能になり、それによって汚染の兆候を監視する新しい機会がもたらされる可能性があります。
競技フィットネスの測定
LTEEが元の方法を超えて成長した別のケースは、進化した大腸菌の適応度が実験環境で増加し、ここで説明するプロトコルを使用して祖先と比較した今日の集団の適応度を直接測定できなくなったことです。進化した集団は祖先を凌駕し、1日の競争の後に数えられる祖先コロニーがほとんどまたはまったく残っていません。この大きな適応度差に対処するための1つのアプローチは、菌株の不均等な開始比を使用し、適合度の低い競合物(たとえば、90 μLの祖先と10 μLの進化した競合物)に向かって混合される初期体積に重みを付けることです。第2のアプローチは、LTEE祖先よりも高い適応度を有する進化したAra−クローンを同定し、MA寒天培地上での選択によってその自然発生的なAra+復帰変異体を単離し、次いで競合アッセイを用いてその復帰株がその親と同じ適応度を有することを確認することである6,38。この新しいAra−/Ara+ペアは、REL606/REL607の代わりに一般的な競合株のセットとして使用できます。理想的には、共通の競合物質として選択された進化したAra−クローン(およびそのAra+復帰体)は、実験で関心のあるすべての株に対して中間の適応度を持ちます。LTEEの最初の50,000世代にわたって、これら2つのアプローチ(不均等な開始比または共通の競合他社を使用)は、典型的なアプローチ39と比較して有意に異なる適応度測定値を生成しませんでした。
競争プロトコルに対するこれらの変更は、常に真実であるとは限らない特定の単純化された仮定を行います。1つは、フィットネス測定は推移的であるということです。つまり、2つの母集団と共通の競合株を別々に競合させると、2つの母集団の相対的な適合度を推測できます。この関係は、大部分がLTEE40について当てはまることが見出されているが、他の実験41については当てはまらない。この不一致の理由の1つは、負の周波数依存適応度効果の進化である可能性があります。この状況は、LTEEの集団A−2からの2つの異なる分岐系統から分離された系統が互いに競合するときに発生します19,42。それぞれがまれな場合、相互給餌のために利点があり、それはそれらの共存を安定させる。異なる変異セットを持つ系統の長期的な共存を示すシーケンシングデータは、同様の相互作用が他のLTEE集団でも発生した可能性があることを示唆していますが14,43、それらが適応度の推定値を著しく変更するほど強力であるかどうかは明らかではありません。最後に、LTEE32の集団A−3におけるクエン酸塩の好気的成長の進化は、これらの細胞の適応度が、クエン酸塩を使用できない細胞と競合するときに「プライベート」リソースの使用を組み込むことを意味し、これらの結果の解釈を複雑にします。これらの例外にもかかわらず、低グルコース濃度とよく振られた環境の使用は、LTEE株と集団の間の適応度比較を行うことを間違いなく簡素化しました。
後の世代では、LTEE集団の一部はTA寒天培地上にコロニーを形成しなくなり、改変されたプロトコルを使用しても競争実験を行うことが困難または不可能になります10。コロニーの成長を必要としない代替法は、アンプリコン44内の2つの代替対立遺伝子を含むリードの割合をカウントするために次世代シーケンシングを使用するFREQ-seqなど、2つの競合の相対的な表現を決定するために潜在的に使用することができる。この方法または同様の方法は、祖先配列に対して、Ara対立遺伝子または pykF や nadRなどの新しく進化した突然変異で使用できる可能性があります。他のタイプの中性マーカーを導入する遺伝子組み換えを行うことも、相対的な適応度を測定するために使用することができる。例えば、蛍光タンパク質遺伝子は、フローサイトメトリー45を用いて競合をカウントできるように、LTEE分枝実験において細胞の染色体に挿入されている。同じ競合用フラスコ内で2つ以上の菌株を混合する可能性を開く別のアプローチは、PCR増幅および配列決定が可能なバーコードを異なる競合他社のゲノムに挿入することです。このアプローチは、進化実験46における系統追跡に使用されている。フローサイトメトリーとバーコードシーケンシングはどちらも、2つの株とコロニーカウントのはるかに極端な比率を正確に測定できるため(寒天プレートでカウントできる<500に対して、10,000>細胞/ゲノムをクエリできるため)、これらの方法を使用すると、一般的な競合他社と比較して測定できる適応度の違いに関してダイナミックレンジを拡大することも約束されます。
長期微生物進化実験のための代替設計
そのすべての長所のために、LTEEは完璧ではありません。その設計の特定の側面は、労働集約的であり、人為的ミスの影響を受けやすくなっています。たとえば、実験を続けるには、研究者が毎日研究室に来て、三角フラスコの間にピペットを挟む必要があります。また、少数の競合他社でも適度な複製でテストされている場合、滅菌ガラス製品、培地、インキュベータースペース、コロニーカウントの要件が急速にエスカレートするため、競合実験は困難なロジスティック上のハードルをもたらす可能性があります。96ウェルマイクロプレートで動作するピペッティングロボットなどのラボラトリーオートメーションシステムや、ケモスタットやタービドスタットなどの連続培養システムを利用しない理由についてよく聞かれます。答えは簡単です:LTEEは、ある意味で、それ自身の長い歴史の囚人です。50 mL三角フラスコで特定の速度で振とうする10 mLの培養物から逸脱することは、実験を根本的に変えるリスクがあるためです。これらの集団が何十年にもわたって適応してきた環境の微妙な側面(例えば、曝気の量)は、マイクロプレートまたは連続培養システムで変化します。各移動における集団のボトルネックも異なり(たとえば、マイクロプレートでは小さい)、進化のダイナミクスが変化する可能性があります。要するに、ここで説明した方法から逸脱すると、LTEEは別の実験になるか、少なくとも進化の軌道を乱す不連続性を導入するリスクがあります。
新しい進化実験を設計する研究者は、潜在的な利点と欠点を認識しながら、微生物集団を増殖させるこれらの他の方法を検討する必要があります。ピペッティングロボットを使用してマイクロウェルプレート内の集団を移送することは、いくつかの点でロジスティック的に単純であり、この方法で増殖できる複製集団の数が多いため、非常に強力であることがわかります47、48、49。しかし、現在のほとんどのセットアップでの自動移送は、完全に無菌条件下では行われないため、外部汚染の可能性が高まります。汚染を防ぐために、増殖培地はしばしば抗生物質で補給され、それは進化に影響を与える環境の特徴になります。マイクロウェルプレートでの移送も、相互汚染イベントが発生しやすくなります。最後に、マイクロウェルプレートの環境は、特に振盪されていない場合、壁の成長、凝集、および1つのウェルに複数のニッチを作成することによって進化を複雑にする可能性のあるその他の現象を選択する傾向があります。リッチメディアまたは高濃度の栄養素を使用して、小さな井戸で個体数を大きく保つと、これらの複雑さが悪化する可能性があります。このような相互作用が発生すると、フィットネスの測定と解釈がはるかに困難になる可能性があります。
微生物進化のための連続培養システムには、新鮮な培地を絶えずポンプで送り込み、培養液をポンプで排出するケモスタットと、自動センシングとポンピングによって培養物を定期的に希釈して細胞を一定の増殖状態に維持するタービドスタットがあります。これらのシステムは、微生物を常に栄養素50を持つ環境に保つことで、微生物が成長と飢餓の間で移行することを回避するため、微生物の生理学と進化をモデル化する場合に非常に役立ちます。光学密度、O2消費量、pH、および培養環境と成長の他の側面をリアルタイムで測定するセンサーを追加することもできます。しかし、現在の連続培養システムでは、カスタムセットアップを構築するために高価な機器の購入または専門知識が必要です51,52,53,54。また、細胞が培養室に付着して希釈を逃れる壁成長は、定期的に滅菌しない限り、連続培養システムの進化のダイナミクスを悩ませます。これらの制約のために、これまでのほとんどのケモスタットおよびタービドスタット進化実験は、連続移動進化実験と比較して、限られた期間であったり、独立して進化する集団が比較的少なかったりしました。
結論
ここでLTEEについて示す方法は、そのユニークな歴史的記録を研究し、これらの 大腸菌 集団の無制限の進化を継続するために重要です。また、ラボラトリーオートメーションを利用したり、LTEEから意図的に省略された自然環境に見られる複雑さのさまざまな要素を追加したりする可能性のある新しい進化実験を検討している他の人に出発点を提供します。1988年以来、実験的進化は分野として繁栄してきました。この間、世界中の研究所の研究者は、進化を研究し、創造的な実験計画を導入し、新しい技術を使用して結果を監視することで革新するためのこのアプローチの計り知れない柔軟性を実証しました。LTEEの手法はエンドポイントではありませんが、今後もこの分野の基盤となることを期待しています。
開示事項
利益相反は宣言されていません。
謝辞
リチャード・レンスキーと 、大腸菌 を使った長期進化実験の研究と維持に貢献した多くの研究者、特にNeerja Hajelaに感謝します。LTEE は現在、米国国立科学財団 (DEB-1951307) によってサポートされています。
資料
| Name | Company | Catalog Number | Comments |
| 2,3,5-Triphenyltetrazolium chloride (TTC) | Sigma-Aldrich | T8877 | |
| 20 mL Glass Beaker | Sigma-Aldrich | CLS100020 | |
| 50 mL Erlenmeyer Flasks | Sigma-Aldrich | CLS498050 | |
| Agar | Sigma-Aldrich | A1296 | |
| Ammonium Sulfate | Sigma-Aldrich | AX1385 | |
| Antifoam | Sigma-Aldrich | A5757 | |
| Arabinose | Sigma-Aldrich | A3256 | |
| Freezer Box (2") | VWR | 82007-142 | |
| Freezer Box (3") | VWR | 82007-144 | |
| Freezer Box Cell Divider (49-place) | VWR | 82007-150 | |
| Freezer Box Cell Divider (81-place) | VWR | 82007-154 | |
| Freezer Vials (1/2-Dram) | VWR | 66009-816 | |
| Freezer Vials (2-Dram) | VWR | 66010-560 | |
| Glucose | Sigma-Aldrich | G8270 | |
| Glycerol | Fisher Scientific | G33 | |
| Magnesium Sulfate | Sigma-Aldrich | M7506 | |
| Metal Tray | Winco | SPJP-202 | |
| Petri Dish | Fisher Scientific | FB0875712 | |
| Potassium Phosphate Dibasic Trihydrate | Sigma-Aldrich | P5504 | |
| Potassium Phosphate Monobasic | Sigma-Aldrich | P5379 | |
| Sodium Chloride | Sigma-Aldrich | M7506 | |
| Sodium Citrate Tribasic Dihydrate | Sigma-Aldrich | C7254 | |
| Test Tube Cap (18mm) | VWR | 10200-142 | |
| Test Tube Rack (18mm, steel) | Adamas-Beta | N/A | Test Tube Racks Stainless Steel Grid Arrangement 72 Holes (17-19 mm) |
| Test Tubes (18 x 150 mm) | VWR | 47729-583 | |
| Thiamine, Hydrochloride | Millipore | 5871 | |
| Tryptone | Gibco | 211705 | |
| Yeast Extract | Gibco | 212750 |
参考文献
- Lenski, R. E., Rose, M. R., Simpson, S. C., Tadler, S. C. Long-term experimental evolution in Escherichia coli. I. Adaptation and divergence during 2,000 generations. The American Naturalist. 138 (6), 1315-1341 (1991).
- Fox, J. W., Lenski, R. E. From here to eternity-the theory and practice of a really long experiment. PLoS Biology. 13 (6), e1002185(2015).
- Daegelen, P., Studier, F. W., Lenski, R. E., Cure, S., Kim, J. F. Tracing ancestors and relatives of Escherichia coli B, and the derivation of B strains REL606 and BL21(DE3). Journal of Molecular Biology. 394 (4), 634-643 (2009).
- Studier, F. W., Daegelen, P., Lenski, R. E., Maslov, S., Kim, J. F. Understanding the differences between genome sequences of Escherichia coli B strains REL606 and BL21(DE3) and comparison of the E. coli B and K-12 genomes. Journal of Molecular Biology. 394 (4), 653-680 (2009).
- Barrick, J. E., Lenski, R. E. Genome dynamics during experimental evolution. Nature Reviews Genetics. 14 (12), 827-839 (2013).
- Lenski, R. E. Experimental studies of pleiotropy and epistasis in Escherichia coli. II. Compensation for maladaptive pleiotropic effects associated with resistance to virus T4. Evolution. 42 (3), 425-432 (1988).
- Calcott, P. H., Gargett, A. M. Mutagenicity of freezing and thawing. FEMS Microbiology Letters. 10 (2), 151-155 (1981).
- Lenski, R. E., Travisano, M. Dynamics of adaptation and diversification: a 10,000-generation experiment with bacterial populations. Proceedings of the National Academy of Sciences of the United States of America. 91 (15), 6808-6814 (1994).
- Wiser, M. J., Ribeck, N., Lenski, R. E. Long-term dynamics of adaptation in asexual populations. Science. 342 (6164), New York, N.Y. 1364-1367 (2013).
- Lenski, R. E., et al. Sustained fitness gains and variability in fitness trajectories in the long-term evolution experiment with Escherichia coli. Proceedings of the Royal Society B: Biological Sciences. 282 (1821), 20152292(2015).
- Barrick, J. E., et al. Genome evolution and adaptation in a long-term experiment with Escherichia coli. Nature. 461 (7268), 1243-1247 (2009).
- Blount, Z. D., Barrick, J. E., Davidson, C. J., Lenski, R. E. Genomic analysis of a key innovation in an experimental Escherichia coli population. Nature. 489 (7417), 513-518 (2012).
- Tenaillon, O., et al. Tempo and mode of genome evolution in a 50,000-generation experiment. Nature. 536 (7615), 165-170 (2016).
- Good, B. H., McDonald, M. J., Barrick, J. E., Lenski, R. E., Desai, M. M. The dynamics of molecular evolution over 60,000 generations. Nature. 551 (7678), 45-50 (2017).
- Consuegra, J., et al. Insertion-sequence-mediated mutations both promote and constrain evolvability during a long-term experiment with bacteria. Nature Communications. 12 (1), 980-980 (2021).
- Cooper, T. F., Rozen, D. E., Lenski, R. E. Parallel changes in gene expression after 20,000 generations of evolution in Escherichia coli. Proceedings of the National Academy of Sciences of the United States of America. 100 (3), 1072-1077 (2003).
- Favate, J. S., Liang, S., Cope, A. L., Yadavalli, S. S., Shah, P. The landscape of transcriptional and translational changes over 22 years of bacterial adaptation. eLife. 11, e81979(2022).
- Khan, A. I., Dinh, D. M., Schneider, D., Lenski, R. E., Cooper, T. F. Negative epistasis between beneficial mutations in an evolving bacterial population. Science. 332 (6034), 1193-1196 (2011).
- Plucain, J., et al. Epistasis and allele specificity in the emergence of a stable polymorphism in Escherichia coli. Science. 343 (6177), 1366-1369 (2014).
- Quandt, E. M., Deatherage, D. E., Ellington, A. D., Georgiou, G., Barrick, J. E. Recursive genomewide recombination and sequencing reveals a key refinement step in the evolution of a metabolic innovation in Escherichia coli. Proceedings of the National Academy of Sciences of the United States of America. 111 (6), 2217-2222 (2014).
- Leon, D., D'Alton, S., Quandt, E. M., Barrick, J. E. Innovation in an E. coli evolution experiment is contingent on maintaining adaptive potential until competition subsides. PLoS Genetics. 14 (4), e1007348(2018).
- Bennett, A. F., Lenski, R. E., Mittler, J. E. Evolutionary adaptation to temperature. I. Fitness responses of Escherichia coli to changes in its thermal environment. Evolution. 46 (1), 16-30 (1992).
- Kibota, T. T., Lynch, M. Estimate of the genomic mutation rate deleterious to overall fitness in E. coli. Nature. 381 (6584), 694-696 (1996).
- Friesen, M. L., Saxer, G., Travisano, M., Doebeli, M. Experimental evidence for sympatric ecological diversification due to frequency-dependent competition in Escherichia coli. Evolution. 58 (2), 245-260 (2004).
- Cooper, T. F. Recombination speeds adaptation by reducing competition between beneficial mutations in populations of Escherichia coli. PLoS Biology. 5 (9), e225(2007).
- Cooper, T. F., Lenski, R. E. Experimental evolution with E. coli in diverse resource environments. I. Fluctuating environments promote divergence of replicate populations. BMC Evolutionary Biology. 10, 11(2010).
- Quan, S., et al. Adaptive evolution of the lactose utilization network in experimentally evolved populations of Escherichia coli. PLoS Genetics. 8 (1), e1002444(2012).
- Deatherage, D. E., Kepner, J. L., Bennett, A. F., Lenski, R. E., Barrick, J. E. Specificity of genome evolution in experimental populations of Escherichia coli evolved at different temperatures. Proceedings of the National Academy of Sciences of the United States of America. 114 (10), E1904-E1912 (2017).
- Izutsu, M., Lake, D. M., Matson, Z. W. D., Dodson, J. P., Lenski, R. E. Effects of periodic bottlenecks on the dynamics of adaptive evolution in microbial populations. BioRixv. , 4457(2021).
- Chavarria-Palma, J. E., Blount, Z. D., Barrick, J. E. LTEE Media Recipes. , (2022).
- Barrick, J. E., Lake, D. M. fitnessR: fitnessR-v1.0.0. barricklab. , (2023).
- Blount, Z. D., Borland, C. Z., Lenski, R. E. Historical contingency and the evolution of a key innovation in an experimental population of Escherichia coli. Proceedings of the National Academy of Sciences of the United States of America. 105 (23), 7899-7906 (2008).
- Grant, N. A., Magid, A. A., Franklin, J., Dufour, Y., Lenski, R. E. Changes in cell size and shape during 50,000 generations of experimental evolution with Escherichia coli. Journal of Bacteriology. 203 (10), 22(2021).
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis. , Springer-Verlag. New York. (2016).
- Meyer, J. R., et al. Parallel changes in host resistance to viral infection during 45,000 generations of relaxed selection. Evolution. 64 (10), 3024-3034 (2010).
- Woods, R., Schneider, D., Winkworth, C. L., Riley, M. A., Lenski, R. E. Tests of parallel molecular evolution in a long-term experiment with Escherichia coli. Proceedings of the National Academy of Sciences of the United States of America. 103 (24), 9107-9712 (2006).
- Barrick, J. E., Deatherage, D. E., D'Alton, S. LTEE-Ecoli: genomics resources for the Long-Term Evolution Experiment with Escherichia coli. , Available from: https://github.com/barricklab/LTEE-Ecoli (2022).
- Izutsu, M., Lenski, R. E. Experimental test of the contributions of initial variation and new mutations to adaptive evolution in a novel environment. Frontiers in Ecology and Evolution. 10, 958406(2022).
- Wiser, M. J., Lenski, R. E. A comparison of methods to measure fitness in Escherichia coli. PLoS One. 10 (5), 0126210(2015).
- de Visser, J. A. G. M., Lenski, R. E. Long-term experimental evolution in Escherichia coli. XI. Rejection of non-transitive interactions as cause of declining rate of adaptation. BMC Evolutionary Biology. 2 (1), 19(2002).
- Paquin, C. E., Adams, J. Relative fitness can decrease in evolving asexual populations of S. cerevisiae. Nature. 306 (5941), 368-371 (1983).
- Rozen, D. E., Lenski, R. E. Long-Term Experimental Evolution in Escherichia coli. VIII. Dynamics of a balanced polymorphism. The American Naturalist. 155 (1), 24-35 (2000).
- Quandt, E. M., Gollihar, J., Blount, Z. D., Ellington, A. D., Georgiou, G., Barrick, J. E. Fine-tuning citrate synthase flux potentiates and refines metabolic innovation in the Lenski evolution experiment. eLife. 4, e09696(2015).
- Chubiz, L. M., Lee, M. -C., Delaney, N. F., Marx, C. J. FREQ-Seq: a rapid, cost-effective, sequencing-based method to determine allele frequencies directly from mixed populations. PLoS One. 7 (10), e47959(2012).
- Gallet, R., Cooper, T. F., Elena, S. F., Lenormand, T. Measuring selection coefficients below 10-3: method, questions, and prospects. Genetics. 190 (1), 175-186 (2012).
- Levy, S. F., et al. Quantitative evolutionary dynamics using high-resolution lineage tracking. Nature. 519 (7542), 181-186 (2015).
- Lang, G. I., Botstein, D., Desai, M. M. Genetic variation and the fate of beneficial mutations in asexual populations. Genetics. 188 (3), 647-661 (2011).
- Frenkel, E. M., et al. Crowded growth leads to the spontaneous evolution of semistable coexistence in laboratory yeast populations. Proceedings of the National Academy of Sciences. 112 (36), 11306-11311 (2015).
- Jordt, H., et al. Coevolution of host-plasmid pairs facilitates the emergence of novel multidrug resistance. Nature Ecology and Evolution. 4 (6), 863-869 (2020).
- Gresham, D., Dunham, M. J. The enduring utility of continuous culturing in experimental evolution. Genomics. 104 (6), 399-405 (2014).
- Miller, A. W., Befort, C., Kerr, E. O., Dunham, M. J. Design and use of multiplexed chemostat arrays). Journal of Visualized Experiments. (72), e50262(2013).
- Toprak, E., et al. Building a morbidostat: an automated continuous-culture device for studying bacterial drug resistance under dynamically sustained drug inhibition. Nature Protocols. 8 (3), 555-567 (2013).
- Wong, B. G., Mancuso, C. P., Kiriakov, S., Bashor, C. J., Khalil, A. S. Precise, automated control of conditions for high-throughput growth of yeast and bacteria with eVOLVER. Nature Biotechnology. 36 (7), 614-623 (2018).
- Ekkers, D. M., Branco Dos Santos, F., Mallon, C. A., Bruggeman, F., Van Doorn, G. S. The omnistat: A flexible continuous-culture system for prolonged experimental evolution. Methods in Ecology and Evolution. 11 (8), 932-942 (2020).
転載および許可
このJoVE論文のテキスト又は図を再利用するための許可を申請します
許可を申請さらに記事を探す
This article has been published
Video Coming Soon
Copyright © 2023 MyJoVE Corporation. All rights reserved