
Method Article
Otimizando o Crescimento de Cristais de Endotiapsina para Experimentos de Cristalografia Seriada
Neste Artigo
Resumo
O objetivo deste artigo é dar ao espectador uma compreensão sólida de como transformar seu protocolo de difusão de vapor de pequeno volume, para o cultivo de grandes cristais de proteína única, em um método de microcristalização em lote de grande volume para cristalografia seriada.
Resumo
Aqui, um protocolo é apresentado para facilitar a criação de grandes volumes (> 100 μL) de polpas microcristalinas adequadas para experimentos seriados de cristalografia em síncrotrons e XFELs. O método é baseado na compreensão do diagrama de fases do cristal de proteínas, e como esse conhecimento pode ser utilizado. O método é dividido em três etapas: (1) otimização da morfologia cristalina, (2) transição para batelada e (3) dimensionamento. O estágio 1 envolve encontrar cristais simples bem difratantes, esperançosamente, mas não necessariamente, apresentando-se em uma morfologia semelhante a um cubo. No Estágio 2, a condição do Estágio 1 é otimizada pelo tempo de crescimento do cristal. Esta estratégia pode transformar cristais crescidos por difusão de vapor em batelada. Uma vez que o crescimento do cristal pode ocorrer dentro de aproximadamente 24 h, um morfograma da proteína e da mistura precipitante pode ser plotado e usado como base para uma estratégia de escalonamento (Estágio 3). Quando os cristais podem ser cultivados em lote, a escala pode ser tentada e o tamanho e a concentração do cristal são otimizados à medida que o volume é aumentado. A endotiapepsina tem sido utilizada como proteína de demonstração para este protocolo. Algumas das decisões apresentadas são específicas da endotiapepsina. No entanto, espera-se que a forma como foram aplicados inspire uma forma de pensar sobre este procedimento que outros possam adaptar aos seus próprios projetos.
Introdução
A cristalografia de macromoléculas à temperatura ambiente (TR) é novamente popular na comunidade de biologia estrutural. O desenvolvimento de fontes de luz XFEL (X-ray Free Electron Laser) estimulou o desenvolvimento de abordagens de entrega de amostras de RT 1,2,3,4, e esses métodos têm sido aplicados a síncrotrons 5,6,7,8. Não só os métodos de RT abrem a possibilidade de estrangulamentos experimentais bomba-sonda 9,10,11,12, mas também há evidências crescentes de que eles promovem estados conformacionais alternativos dentro das proteínas 13,14,15,16,17.
No entanto, a principal razão pela qual os criométodos ganharam força sobre as abordagens de RT no final da década de 1990 foi a desaceleração dos danos da radiação por temperaturas cristalinas abaixo de zero18. O criométodo19 começou a permitir a coleta de um conjunto completo de dados a partir de um único cristal de proteína. Métodos modernos de RT em XFELs e síncrotrons resolveram o problema dos danos por radiação monocristalina através do desenvolvimento de estratégias rápidas de liberação de cristais (> 100 Hz) 1,2,3,4. Esses métodos permitem a coleta de um conjunto de dados completo a partir de milhares de cristais expostos individualmente. Essas abordagens de entrega de RT requerem, portanto, a produção de grandes quantidades de soluções contendo microcristais homogêneos (> 100 μL de < cristais de 50 μm). No entanto, como os criométodos tendem a exigir apenas cristais individuais, os métodos para criar tais polpas microcristalinas não são atualmente onipresentes nos laboratórios de cristalografia de proteínas.
Existem exemplos na literatura de como realizar partes do procedimento de otimização da microcristalização para amostras seriadas de cristalografia. Aqui, deve-se fazer uma distinção entre proteínas de membrana e proteínas solúveis. Protocolos para otimizar o crescimento de cristais de proteína de micromembrana cultivados em monooleína (ou algum outro lipídio), para fase cúbica lipídica (LCP), têm sido bem descritos20,21,22. No entanto, métodos para a microcristalização de proteínas solúveis, incluindo proteínas de membrana cultivadas em condições não-LCP, são geralmente escassos. Estudos anteriores focaram em partes específicas do processo, como a triagem de microcristais 23,24, o aumento da nucleação 24 e a escala usando difusão em interface livre 25, mas não um método completo.
No entanto, recentemente foi descritoum método 26 que tenta oferecer um protocolo completo. Como muitos aspectos da cristalografia de proteínas, ela não é nova. Muitas das ideias propostas já foram descritas por Rayment (2002)27. O método visa mostrar aos cristalógrafos como realizar a conversão de um único cristal cultivado usando difusão de vapor, para uma metodologia em batelada para cultivar milhares de microcristais. O método enfoca a difusão de vapor como um ponto de partida comum, já que 95% de todas as deposições do Protein Data Bank (PDB) provêm de cristais crescidos em placas de difusão de vapor26. A difusão de vapor não é, no entanto, o método ideal para microcristalização26, por isso é descrita uma metodologia para converter a difusão de vapor em cristalização em batelada. Uma vez que os cristais podem ser cultivados em lote, as rotas de escala para volumes maiores tornam-se mais viáveis. Dado os caprichos da cristalização de proteínas, os autores enfatizam que este método não é à prova de falhas. No entanto, o protocolo deve, pelo menos, fornecer uma visão sobre o "espaço de cristalização" de uma proteína.
Este método baseia-se no diagrama de fase de cristalização de proteínas e em como a compreensão desse diagrama pode atuar como um guia durante a otimização da microcristalização. Um diagrama de fase de proteína é comumente representado como um gráfico x/y com concentrações precipitantes e proteicas nos eixos x e y, respectivamente (Figura 1A). A partir do ponto de água pura (canto inferior esquerdo - Figura 1A), a concentração de proteína e precipitante aumenta até atingir a linha de solubilidade. A linha de solubilidade marca o ponto de supersaturação (linha roxa - Figura 1A). Quando uma proteína é supersaturada, a solução torna-se termodinamicamente instável e começará a separar-se em duas fases: "rica em proteínas" e uma solução saturada estável. Essa separação pode ocorrer em qualquer lugar além da linha de solubilidade e sua cinética depende das propriedades da proteína e dos componentes da solução.
Quando as concentrações de proteína e precipitante são muito grandes, a proteína se decompõe de forma instável fora da solução e resulta em precipitado amorfo (região rósea - Figura 1A). No entanto, a separação ordenada de fases pode ocorrer na região de nucleação [ver Garcia-Ruiz (2003)28 para descrição detalhada] e os nucleantes cristalinos têm propensão a se formar (região verde - Figura 1A). A nucleação e o crescimento removem a proteína da solução e movem a gota para a região metaestável, onde o crescimento pode continuar até que a linha de solubilidade seja atingida [ver McPherson e Kuznetsov (2014)29 para discussão detalhada]. O diagrama é, para a grande maioria das condições de cristalização, uma simplificação grosseira30. Apesar disso, no entanto, o diagrama ainda é de grande utilidade para os microcristalógrafos, pois o mapeamento do diagrama permite determinar a linha de solubilidade e a cinética de nucleação.
Em termos de criação de microcristais, os dois fatores durante a cristalização que precisam ser otimizados são o número de cristais (Xn) e sua dimensão média e mais longa (Xs). X n será proporcional ao número de eventos de nucleação (n ) (Eq. 1).
 Eq. 1
Eq. 1
X s é proporcional à concentração de proteína livre acima da linha de solubilidade (Ps) dividida por Xn (Eq. 2).
 Eq. 2
Eq. 2
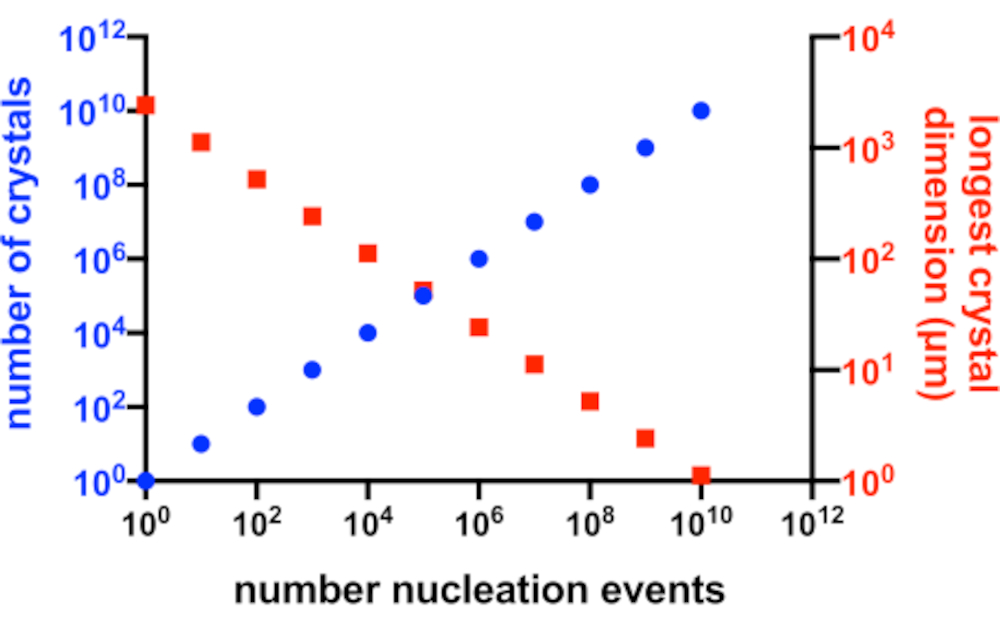
Em uma situação perfeita, cada evento de nucleação produziria um cristal possível e cada um desses cristais, teria igual acesso à proteína disponível em solução. A Figura 2 é uma representação gráfica de um cenário ideal da relação entre Xn e Xs. Na prática, o principal controle que um cristalógrafo tem sobre Xn e Xs é influenciando a quantidade de nucleação ou pela adição de cristais de sementes. O microcristalógrafo deve julgar como aumentar Xn de tal forma que uma concentração de cristal adequada e tamanho de cristal possam ser criados.
A maioria das técnicas de cristalização requer um "período de transição" (Figura 1B). Por exemplo, em um experimento de difusão de vapor, ao misturar a proteína e as soluções precipitantes, as concentrações de cada uma mudam à medida que a gota se equilibra com a solução do poço. Espera-se que essas mudanças façam a transição gradual da queda para a zona de nucleação, onde a propensão para cristalização aumentará. À medida que os cristais começam a se nuclear e crescem, a quantidade de proteína em solução começará a cair, diminuindo a probabilidade de nucleação adicional. A quantidade final de nucleação será específica da proteína e da condição, e também dependente da profundidade de penetração na zona de nucleação. Dada a penetração limitada da zona de nucleação de métodos que requerem uma etapa de transição, o nível de nucleação será limitado à taxa de nucleação no limite da região de nucleação metaestável.
Devido à importância de ser capaz de aumentar o nível de nucleação para um microcristalógrafo, é importante passar para uma metodologia de cristalização em batelada. O lote pode aproveitar melhor toda a região de nucleação (Figura 1C). Em métodos em batelada, a ideia é misturar a proteína e o precipitante de forma que uma solução supersaturada seja criada sem a necessidade de mudanças nas concentrações dos componentes. A nucleação deve ser possível imediatamente após a mistura. Os métodos em lote permitem, portanto, que toda a zona de nucleação seja teoricamente atingida. Qualquer aumento na cinética de nucleação além do limite de nucleação metaestável pode então ser utilizado.
Se o nível basal de nucleação de cristais não for suficiente para gerar um Xn grande, métodos de micro-semeadura podem ser usados. Na micro-semeadura, cristais pré-cultivados são quebrados para criar uma lama de fragmentos cristalinos que podem atuar como arcabouço para o crescimento de cristais frescos31,32. A microsemeadura tem sido amplamente utilizada na preparação seriada de amostras cristalográficas como forma de aumentar Xn sem a necessidade de aumentar a nucleação de cristais (Figura 1C).
A transição da difusão de vapor para a batelada pode ser visualizada em um diagrama de fases como movendo o ponto de partida experimental das regiões não supersaturadas ou metaestáveis para a zona de nucleação. Isso pode ser feito aumentando-se as concentrações de proteínas e/ou precipitantes e/ou a relação dos dois dentro da gota (Figura 1D), observando-se em quais condições os cristais aparecem rapidamente (< 24 h)26. O equilíbrio completo da gota de difusão do vapor pode levar dias ou semanas33. Portanto, ao procurar condições que mostrem cristais aparecendo rapidamente, condições de lote podem ser encontradas sem ter que passar para formatos alternativos de triagem de cristalização, como micro-lote34,35,36,37.
Uma vez que a zona de nucleação foi encontrada, uma condição de lote foi encontrada e um morfograma - aqui, um diagrama de fase aproximado - pode ser criado. O morfograma é de grande utilidade quando se pensa em utilizar um protocolo de batelada semeada ou batelada reta. Plotando o Xn em função da proteína e da concentração precipitante, pode-se avaliar a cinética de nucleação26. Se X n permanecer baixo em toda a região de nucleação, o lote semeado pode ser necessário para tornar Xn grande o suficiente para limitar o crescimento de cristais. Essa avaliação é o primeiro passo no processo de escalonamento para volumes maiores (> 100 μL).
Este método foi projetado de tal forma que pudesse ser conduzido na maioria dos laboratórios de cristalização usando equipamento padrão de cristalização por difusão de vapor. Muitos estudos também têm sido realizados descrevendo técnicas para facilitar muitas partes desse processo, caso o equipamento esteja disponível. Estes incluem, mas não estão limitados a, espalhamento dinâmico de luz (DLS)25,27, imagem não-linear 20,24,25, difração de pó 20,24,27 e microscopia eletrônica 26 [ver Cheng et al. (2020)40 para uma bela revisão].
O objetivo deste trabalho é fornecer uma demonstração visual do método de transição da cristalização por difusão de vapor de pequeno volume (< 500 nL) para cristalização em batelada de grande volume (> de 100 μL). A endotiapepsina de Cryphonectria parasitica tem sido usada como um sistema de exemplo para demonstrar essa tradução. O tipo de experimento e o método de entrega da amostra para os quais os microcristais são necessários influenciarão a saída idealde X s 26. Para experimentos de mistura que requerem uma resolução de tempo de milissegundos41 ou bicos virtuaisdinâmicos a gás42, um X s final de < 5 μm pode ser desejável. Neste caso, o objetivo era produzir cristais de proteína que difratassem a aproximadamente 1,5 Å, para um experimento de bomba-sonda ativada por fótons, e usando uma abordagem de entrega de alvo fixo.
Para ilustrar as necessidades amostrais de um experimento de cristalografia seriado usando endotiapsina, a Tabela 1 mostra os parâmetros experimentais de um experimento hipotético. As informações da amostra foram baseadas no protocolo descrito a seguir. Dadas algumas estimativas conservadoras sobre as taxas de acerto e os requisitos de coleta de dados, 50 mg é a estimativa do consumo total da amostra para todo o experimento.
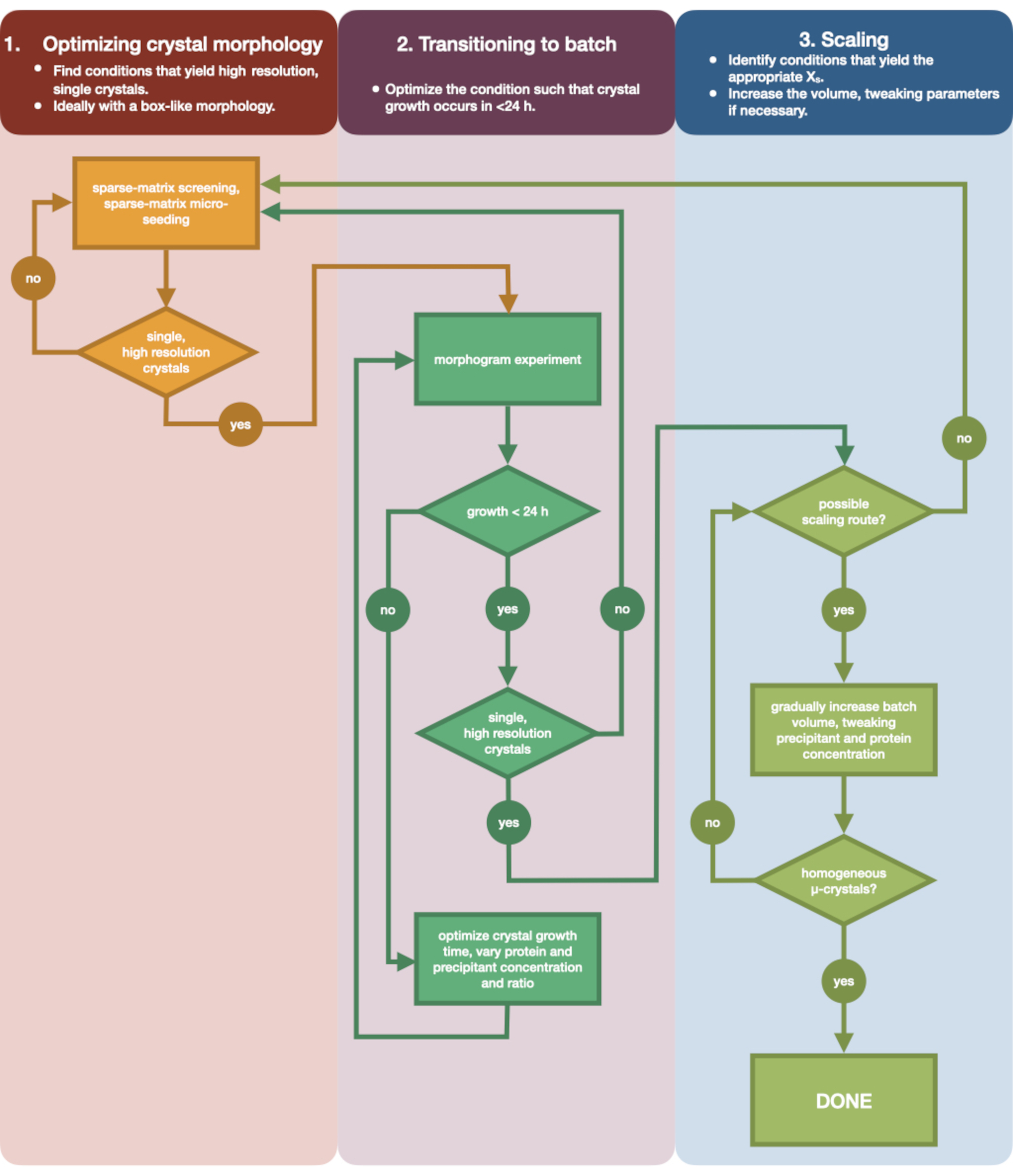
A Figura 3 mostra um fluxograma do processo completo de otimização, desde a cristalização inicial por difusão de vapor de pequeno volume até o lote em grande escala. Para a maioria dos projetos de cristalografia seriada, este protocolo começará na Etapa 2: 'transição para lote', uma vez que a proteína alvo já terá sido cristalizada. No entanto, o Passo 1 foi incluído para completude e para lembrar os leitores de sua importância. Encontrar uma condição que dê origem a um cristal bem difratante, único e grande é o melhor ponto de partida para a otimização de microcristais. Na Etapa 2, essa condição pode então ser otimizada da difusão de vapor para batelada, e um morfograma das regiões de nucleação e metaestável pode ser plotado. Feito isso, o dimensionamento da condição de lote para volumes maiores pode ser executado na Etapa 3. Ao final do fluxograma, um cristalógrafo terá criado um protocolo de microcristalização repetível, de grande volume (> de 100 μL), de microcristalização, em lote para endotiapsina. Este método pode então ser aplicado à sua proteína particular de interesse.
Protocolo
NOTA: Todos os experimentos de cristalização de gotas sentadas de 96 poços foram configurados usando placas de 2 ou 3 gotas. Um robô de manuseio de líquidos e um imageador/hotel de cristalização foram usados para facilitar a preparação e o monitoramento de todas as telas de 96 poços. Todas as concentrações de reagentes para experimentos de cristalização são dadas em suas concentrações iniciais antes da mistura.
1. Otimizando a morfologia dos cristais
NOTA: Passos 1.1.1. e 1.1.6. descrever como as condições de cristalização da endotiapepsina foram encontradas e como essas condições foram otimizadas para encontrar uma única condição que produzisse cristais únicos e bem difratantes.
- Otimização de matriz esparsa
- Preparar solução fresca de endotiapepsina.
NOTA: A endotiapepsina, quando adquirida como Superan 600, deve ser transferida para fora de sua solução de armazenamento e concentrada.- Preparar 3 L de acetato 0,1 M na pH 4,6 a 4 °C.
- Corte 20 cm da tubulação de diálise e lave brevemente no tampão. Sele uma extremidade da tubulação usando um clipe, coloque 50 mL da solução de endotiapepsina na tubulação e, em seguida, sele a outra extremidade.
- Deixar a solução dialisar durante, pelo menos, 4 h (ou durante a noite) a 4 °C em 1 L do tampão acetato de Na. Devido aos componentes do tampão de armazenamento, a solução na bolsa de diálise passará a ser de aproximadamente 100 mL.
- Transfira a bolsa de diálise contendo a endotiapsina para um litro fresco de 4 °C, acetato de 0,1 M Na pH 4,6. Repita este passo mais uma vez, de modo a que o tampão original tenha sido diluído 2000x contra o acetato de Na.
- A endotiapepsina agora estará em aproximadamente 10 mg/mL. Concentrar a 100 mg/mL usando um concentrador centrífugo de 10 kDa e uma centrífuga.
- Resfriar rapidamente a solução de endotiapsina em nitrogênio líquido em alíquotas de 50 μL e armazenar a -80 °C.
- Prepare uma tela PACT Premier de matriz esparsa de 96 poços.
- Usando um robô de manuseio de líquidos, dispense 100 nL de 70 mg/mL de endotiapepsina e 100 nL de solução de poço em um único subpoço por poço. Misture a proteína e a solução de poço 3 vezes após a adição do tampão de cristalização.
- Sele a placa e deixe durante 28 dias a 20 °C tirar imagens todos os dias durante a primeira semana e, posteriormente, todas as semanas durante 4 semanas.
- Análise matricial esparsa
- Identificar acertos que produzem cristais únicos de endotiapsina. A partir da tela PACT, as condições que continham MgCl2 cresceram como singletons em vez de aglomerados de agulhas.
- Otimização de matriz esparsa
- A partir do MgCl2 contendo as condições identificadas na Etapa 1.1.3.1, crie uma tela de 96 poços combinando e variando aleatoriamente os diferentes componentes do poço.
- Usando um robô de manuseio de líquidos, dispense 100 nL de 70 mg/mL de endotiapepsina e 100 nL de solução de poço em um único subpoço por poço. Misture a proteína e a solução de poço 3 vezes após a adição do tampão de cristalização.
- Sele a placa e deixe durante 28 dias a 20 °C tirar imagens todos os dias durante a primeira semana e, posteriormente, todas as semanas durante 4 semanas.
- Análise de otimização
- Usando um software de planilha adequado, classifique as condições de cristalização que dão origem aos cristais com base na qualidade do cristal e no nível de precipitação, nenhum cristal (0) a ideal (5) e baixo (0) a alto (5), respectivamente. Com relação à qualidade dos cristais, os critérios gerais são os monocristais com morfologia em forma de caixa.
- Realizar uma análise de correlação de Pearson entre os teores da condição de cristalização e a quantidade de cristais e o nível de precipitação.
- Plote esses dados como um mapa de calor. Procure componentes e condições que foram correlacionados com os desfechos preferidos.
- Análise de difração.
- Confirmar que os cristais cultivados a partir das condições identificadas na Etapa 1.1.5 são adequados para cristalografia seriada realizando um experimento de difração de raios X.
- Carregar uma amostra dos cristais de endotiapepsina de cada uma das condições identificadas em suportes que permitam a coleta de dados a 100 ou 293 K e realizar um experimento de difração de raios X. Se trabalhar sob crioterapia, use 25% de etilenoglicol como crioprotetor.
- Processe esses dados por meio de um pacote de software adequado. Os cristais de endotiapsina devem difundir-se para além de 1,5 Å. Verifique se há geminação, pois cristais geminados podem complicar significativamente o processamento de dados cristalográficos seriais.
- Se os cristais forem singletons e difratarem para 1,5 Å prossiga para o Passo 2. Caso contrário, volte para a Etapa 1.1.2 e tente telas de matriz mais esparsas para identificar condições promissoras. Após as análises realizadas nas etapas 1.1.5. e 1.1.6., uma condição de cristalização de 25% (p/v) PEG 6.000, Tris-HCl 0,1 M pH 7,0 e 0,15 M MgCl2 deve ter sido encontrada como ideal aproximadamente.
- Preparar solução fresca de endotiapepsina.
2. Transição para lote
- Experimento de morfograma
- Crie um estoque de sementes de micro-cristal.
NOTA: É uma boa prática ao fazer sementes-estoques, para fazer as sementes a partir de cristais cultivados especificamente para a tarefa. Isso ajuda muito na reprodutibilidade. Outras ideias apresentadas nos Passos 2.1.1.1 a 2.1.1.11 são sempre usar os cristais cultivados a partir de um número padrão de poços - aqui 5 - e alicitar os estoques uma vez que eles são feitos para anular os ciclos de congelamento-descongelamento.- Preparar uma placa de cristalização de 96 poços com poços contendo o tampão de cristalização: 25% (p/v) PEG 6.000, 0,1 M Tris-HCl pH 7,0 e 0,15 M MgCl2.
- Usando um robô de manuseio de líquidos, dispense 200 nL de endotiapepsina descongelada 70 mg/mL e 200 nL de solução de poço em um único subpoço por poço. Misture a proteína e a solução de poço 3 vezes após a adição do tampão de cristalização.
- Sele a placa e deixe por 24 h.
- Encher um tubo de centrífuga de 1,5 mL com 250 μL de tampão de cristalização e 10-15 esferas de vidro de 1 mm. Deixe o tubo da centrífuga no gelo para esfriar por 5-10 min.
- Selecione 5 poços com cristais, abra os poços com um bisturi e, usando uma ponta de pipeta, esmague os cristais nos poços.
- Aspirar 1 μL de tampão do tubo de centrífuga gelado e usar para homogeneizar a lama de cristal triturada. Uma vez homogêneo, aspirar toda a lama e coletar no tubo de centrífuga resfriado.
- Repita a etapa 2.1.2.6 para cada um dos 5 subpoços.
- Vórtice o tubo de centrífuga contendo o tampão, as lamas agrupadas e as contas a 1000 rpm por 30 s.
- Retorne o tubo da centrífuga ao gelo por 30 s.
- Repita as etapas 2.1.2.8 e 2.1.2.9 mais duas vezes.
- O estoque de sementes já está pronto e pode ser aliquotado em lotes de 10 μL e armazenado a -20 °C.
- Realizar experimento de morfograma.
- Prepare uma tela de grade de 2 gotas e 96 poços. Variar a concentração de PEG 6.000 de 5 a 40% (p/v) ao longo das colunas da placa, mantendo o tampão e o sal a 0,1 M Tris-HCl pH 7,0 e 0,15 M MgCl2, respectivamente.
- Preparar uma diluição sequencial de endotiapepsina em acetato de 0,1 M Na pH 4,6 de 100 a 12,5 mg/mL ao longo de 8 etapas. Uma concentração diferente de endotiapepsina será utilizada para cada fileira da placa.
- Usando um robô de manuseio de líquidos, distribua 150 nL de endotiapepsina em ambos os subpoços 1 e 2. No sub-poço 1, dispensar 150 nL da solução do poço. No subpoço 2, multiaspirar 50 nL de caldo de semente descongelado e 100 nL de solução de poço e, em seguida, dispensar ambos na solução proteica. Misturar as soluções 3 vezes após a adição do tampão de cristalização.
- Sele a placa e deixe a 20 °C tirando imagens a cada 0, 3, 6, 12, 18, 24 h, depois todos os dias durante a primeira semana e todas as semanas para as quatro seguintes. Se a imagem automática não for possível, não se preocupe com a imagem horária no dia 1.
- Crie um estoque de sementes de micro-cristal.
- Análise do morfograma
- Observando as imagens obtidas após 24 h, estime o número de cristais presentes em cada poço e registre essas estimativas na planilha "geradora de morfogramas" fornecida. Estas estimativas não têm de ser precisas; Contar individualmente milhares de microcristais, se presentes, não é prático ou necessário. Principalmente tentar garantir que as estimativas sejam consistentes ao longo de toda a chapa.
NOTA: A regra das 24 horas foi baseada nas observações feitas em Beale et al (2019)26. As condições de cristalização por difusão de vapor podem levar dias ou semanas para se equilibrarem. Os cristais que aparecem rapidamente são mais propensos a ter crescido através de um processo em lote do que pelo equilíbrio gradual dos componentes da gota. O critério de 24 h é, portanto, um tanto arbitrário e um tempo de corte exato entre um experimento de difusão em lote e vapor dependerá da mistura específica da condição [ver Beale et al. (2019) 26 para mais detalhes]. - Inserir as concentrações iniciais de endotiapepsina e PEG 6.000 nas caixas indicadas.
- A planilha plotará automaticamente os resultados no formato tradicional de diagrama de fase com precipitante e concentração de proteína nos eixos x e y , respectivamente. Condições de poços que só dão origem a cristais em suas gotas semeadas indicam a região metaestável do diagrama (azul transparente), enquanto condições que possuem cristais tanto nas gotas semeadas quanto nas não semeadas indicam a zona de nucleação (verde sólido).
NOTA: Idealmente, a maioria da zona de nucleação deve estar presente no diagrama (ou seja, existem alguns poços claros na parte inferior do diagrama e algum precipitado deve ser visível em altas concentrações de proteínas e precipitantes). Se este não for o caso, talvez, repita o experimento, mas aumente a concentração de proteína e/ou precipitante (se possível). - Se o cristal tiver aparecido em menos de 24 h, avance para o Passo 2.3.1. Caso contrário, prossiga para a Etapa 2.4 e continue otimizando para o lote.
- Observando as imagens obtidas após 24 h, estime o número de cristais presentes em cada poço e registre essas estimativas na planilha "geradora de morfogramas" fornecida. Estas estimativas não têm de ser precisas; Contar individualmente milhares de microcristais, se presentes, não é prático ou necessário. Principalmente tentar garantir que as estimativas sejam consistentes ao longo de toda a chapa.
- Análise de cristais
- Como dito no final da Etapa 1, antes de passar para a próxima etapa, certifique-se de que esses cristais tenham a morfologia e a qualidade de difração desejadas. No que diz respeito à morfologia, os cristais são observavelmente desgeminados e formando-se como singletons em vez de estruturas semelhantes a bolas de agulha ou leques? Com relação à difração, coletar dados de difração dos cristais, se possível. Se esses cristais não difratarem, é improvável que os cristais crescidos em um volume maior difratem.
- Coloque uma amostra dos cristais de endotiapsina do experimento de morfograma em suportes que permitam a coleta de dados em 100 ou 293 K e realize um experimento de difração de raios X. Se trabalhar sob crioterapia, use 25% de etilenoglicol como crioprotetor.
- Processe esses dados por meio de um pacote de software adequado. Os cristais de endotiapsina devem difundir-se para além de 1,5 Å. Através da amostra de cristais, observe o tamanho da célula, o número total de observações e a mosaicidade; Estas medidas darão uma indicação quanto à homogeneidade dos cristais difratantes.
- Se a morfologia cristalina e a qualidade da difração forem suficientes, prossiga para a Etapa 3.
- Otimize o tempo de crescimento do cristal.
NOTA: A análise do morfograma (Etapa 2.2) terá dado uma indicação do ponto de partida da cristalização (ou seja, a região do diagrama de fases onde a gota está localizada quando as soluções precipitante e proteica foram misturadas). A queda está na região metaestável ou abaixo da linha de solubilidade? A cristalização em batelada começa na zona de nucleação (Figura 1C). O objetivo desta etapa é mover esse ponto de partida abaixo da linha de solubilidade ou da região metaestável, para a zona de nucleação (Figura 1D). Se as sementes caírem do Passo 2.2. produziram cristais rapidamente, isso é uma indicação de que a mistura de gotas já está na região metaestável, se não, então é provável que a gota não esteja supersaturada.- Otimizando o tempo de crescimento dos cristais.
- Usando a mesma tela da Etapa 2.1.3, prepare um experimento de cristalização por difusão de vapor de 96 poços em uma placa de 3 gotas.
- Aumentar a concentração de proteína inicial de endotiapepsina no eixo y (ou seja, concentrar ainda mais a proteína, talvez 120 mg/mL para endotiapepsina).
- Efectuar uma diluição em série, como no passo 2.1.3.2, de modo a que cada linha da placa contenha uma concentração de proteínas sequencialmente mais baixa.
- Use diferentes proporções de queda em cada uma das três gotas na placa: 1:1, 1:2 e 2:1, proteína:precipitante.
- Veja ou imagine a placa no primeiro dia às 0, 3, 6, 12, 18, 24 h e depois todos os dias durante a primeira semana e todas as semanas para as quatro seguintes. Se a imagem automática não for possível, não se preocupe com a imagem horária no dia 1.
- Identifique as gotas que produzem os cristais que aparecem mais rapidamente e faz destes os pontos de partida de otimizações repetidas até que o crescimento dos cristais ocorra dentro de 24 h.
- Quando uma condição de cristal que aparece rapidamente tiver sido identificada, retorne à Etapa 2.1 para reposicionar o morfograma como um prelúdio para começar a escalar.
- Otimizando o tempo de crescimento dos cristais.
3. Dimensionamento
- Classifique as rotas de escala. Nesta fase, não é necessário decidir sobre uma única rota de escala, apenas identificar e classificar as opções para que elas possam ser exploradas por sua vez. À medida que o volume da mistura em lote é aumentado durante o procedimento de escala, ocorrerão mudanças na taxa de nucleação e na faixa de tamanhos de cristais. No entanto, estes podem ser superados por ajustes cuidadosos das concentrações dos componentes à medida que o volume escalonado é aumentado.
NOTA: As etapas 3.1.1 e 3.1.2 descrevem como discernir, a partir do morfograma, se um protocolo de lote ou lote semeado é mais apropriado.- Protocolo de lote direto
- O Xn na zona de nucleação é proporcional à concentração de proteína e/ou precipitante? Ou seja, Xn aumenta em função da concentração de precipitantes e/ou proteínas? -Sim? Vá para a Etapa 3.1.1.2. Não? Vá para a Etapa 3.1.2.
- Localize as condições que produzem cristais do tamanho necessário e vá para a Etapa 3.2.
- Protocolo de lotes semeados
- O Xn é plano através da zona de nucleação? Ou seja, Xn não aumenta em função da concentração de precipitantes e/ou proteínas.
- Localize as condições de semeadura que produzem cristais do tamanho necessário e vá para a Etapa 3.2. Se todos os cristais forem muito grandes - vá para o Passo 3.1.2.3.
- Repetir o experimento do morfograma (Etapa 2.1), mas desta vez aumentar a concentração do estoque de sementes utilizado nos poços semeados. O estoque de sementes pode ser aumentado com o uso de mais cristais em sua criação. Por exemplo, em vez de 5 poços na Etapa 2.1.1.5, use 10 poços.
- Visualize ou visualize a placa ao longo das primeiras 0, 3, 6, 12, 18, 24 h.
- O Xn deveria ter aumentado e o Xs diminuído nas gotas semeadas. Repita esse ciclo se cristais menores forem necessários e, em seguida, siga um protocolo de lote semeado.
- Protocolo de lote direto
- Dimensionamento gradual
- Escalonamento em placas de 96 poços. A partir do morfograma da endotiapepsina, um método em batelada reta usando a condição de cristalização 0,1 M Tris-HCl pH 7,0, 0,15 M MgCl2 e 30% (p/v) PEG 6.000, foi inicialmente selecionado para raspagem. 100 mg/mL de endotiapepsina misturada ao tampão de cristalização na proporção de 1:1.
- Preparar 2-3 poços em uma placa de 2 poços de 96 poços com 100 μL de Tris-HCl 0,1 M pH 7,0, 0,15 M MgCl2 e 30% (p/v) PEG 6.000.
- Usando 100 mg/mL de solução de endotiapepsina recém-descongelada, dispensar 0,5 μL de proteína e 0,5 μL de precipitante por poço, selar e armazenar a 20°C.
- Visualize ou visualize a placa ao longo das primeiras 0, 3, 6, 12, 18, 24 h. Observe quaisquer alterações no intervalo de Xs e Xn.
- Se tiverem ocorrido alterações, repita os passos 3.2.1.1 a 3.2.1.2, mas aumente ou diminua a concentração deproteínas , precipitantes e/ou sementes para restaurar quaisquer alterações ao intervalo de X s e Xn.
- Quando o intervalo de Xs e Xn for aceitável, avance para o Passo 3.2.2.
- Dimensionamento em placas suspensas de 24 poços
- Prepare um único poço de uma placa suspensa de 24 poços untando as bordas do poço com graxa a vácuo.
- Preparar 0,5 mL de Tris-HCl 0,1 M pH 7,0, 0,15 M MgCl2 e 30% (p/v) PEG 6.000 e encher bem o untado.
- Usando solução de endotiapepsina recém-descongelada, pipete 1 μL de proteína na superfície de uma lamínula de vidro. Pipetar 1 μL de tampão de cristalização sobre a gota de proteína e misturar usando a pipeta.
- Visualize ou visualize a placa ao longo das primeiras 0, 3, 6, 12, 24 h. Observe quaisquer alterações no intervalo de Xs e Xn.
- Se tiverem ocorrido alterações, repita os passos 3.2.2.1 a 3.2.2.4, mas aumente ou diminua a concentração deproteínas , precipitantes e/ou sementes para restaurar quaisquer alterações na gama de X s e Xn.
- Quando/se o intervalo de Xs e Xn for aceitável, avance para o Passo 3.2.2.7.
- Repetir os passos 3.2.2.1 a 3.2.2.5, aumentando gradualmente o volume total do experimento para 10 μL.
- Uma vez com um volume igual ou superior a 10 μL, proceder aos tubos de centrifugação no passo 3.2.3.
- Dimensionamento em tubos de centrífuga
NOTA: O refinamento da condição do lote de endotiapepsina ocorreu principalmente no ponto de volumes de 200 μL (ver Resultados, Escala). O processo iniciou-se com uma condição de cristalização de 0,1 M Tris-HCl pH 7,0, 0,15 M MgCl2 e 30% (p/v) PEG 6.000. No entanto, a concentração de PEG acabou mudando para 40% (p/v). As sementes também foram necessárias para controlar o Xn, e para evitar que os cristais crescessem muito grandes, o crescimento dos cristais teve que ser extinguido. As etapas 3.2.3.1 a 3.2.3.7 detalham o processo de otimização da condição. Passo 3.2.4. Descreva o protocolo do lote final.- Preparar 1 mL de tampão de cristalização: 0,1 M Tris-HCl pH 7,0, 0,15 M MgCl2 e 30% (p/v) PEG 6.000.
- Usando endotiapepsina recém-descongelada 100 mg/mL, adicione 25 μL de proteína a um tubo de centrífuga de 1,5 mL.
- Misture bem o tampão de cristalização com a solução proteica numa proporção de 1:1 com uma ponta de pipeta. Colocar o tubo num revólver/rotador com agitação elevada a 20 °C.
- Tome alíquotas regulares (5, 10, 30, 60 min, 2, 5, 10, 24 h) de 2,5 μL e visualize em um hemocitômetro. Registre o intervalo Xn eX s .
- Se tiverem ocorrido alterações, repita as etapas 3.2.3.1. até 3.2.3.4. mas aumentar ou diminuir a concentração deproteínas , precipitantes e/ou sementes para restaurar quaisquer alterações na faixa de X s e Xn
- Quando o intervalo de Xs e Xn for aceitável, avance para o passo 3.2.3.7.
- Repetir os passos 3.2.2.1 a 3.2.2.5, aumentando gradualmente o volume total do experimento para 200 μL ou mais, conforme necessário.
- Protocolo final de lotes semeados
- Preparar caldo de sementes.
- Preparar 2 mL de tampão de cristalização: 0,1 M Tris-HCl pH 7,0, 0,15 M MgCl2 e 40% (p/v) PEG 6.000.
- Usando endotiapepsina recém-descongelada, adicione 100 μL de proteína a um tubo de centrífuga de 1,5 mL.
- Misture bem o tampão de cristalização com a solução proteica numa proporção de 1:1 com uma ponta de pipeta. Colocar o tubo num revólver/rotador com agitação elevada a 20 °C durante 24 h para permitir o crescimento de cristais de 50 μm.
- Adicione 10-15 contas de vidro de 1 mm à pasta de cristal de 50 μm.
- Vórtice o tubo centrífugo contendo a lama e as contas a 1000 rpm por 30 s.
- Retorne o tubo da centrífuga ao gelo por 30 s.
- Repita as etapas 3.2.4.1.5 e 3.2.4.1.6 mais 10 vezes.
- Isso agora é 200 μL de um estoque de sementes de 1x. Diluir o estoque de sementes 10x com a adição de 1,8 mL de tampão de cristalização. Aliquot o estoque de sementes de 10x em lotes de 50 μL e armazenar a -20 °C.
- Protocolo de lotes semeados.
- Preparar tampão de cristalização: 0,1 M Tris-HCl pH 7,0, 0,15 M MgCl2 e 40% (p/v) PEG 6.000.
- Em um tubo de centrífuga, misturar 100 μL de tampão de cristalização com 50 μL de caldo recém-descongelado de 10x.
- Usando endotiapepsina recém-descongelada, adicione 150 μL de proteína a um tubo de centrífuga de 1,5 mL.
- Misture bem a mistura tampão de cristalização/semente com a solução de endotiapsina com uma ponta de pipeta e coloque o tubo em um revólver/rotador com alta agitação a 20 °C.
- Monitore a cristalização tomando alíquotas regulares de 2,5 μL e visualize os cristais em um hemocitômetro. Registre o intervalo Xn eX s .
- Após aproximadamente 80 min, quando os cristais atingirem um Xs de 15 μm, extinguir a reação pela adição de 150 μL de acetato de 0,05 M Na pH 4,6, 0,05 M de Tris-HCl pH 7,0, 0,075 M MgCl2 e 20% (p/v) de PEG 6.000 (solução composta de tampão endotiaapepsina e tampão de cristalização, misturado 1:1).
- Conservar os cristais a 20 °C.
- O protocolo produziu uma faixa de tamanho de cristal e um número aceitáveis para o experimento pretendido? Sim - FEITO - Não - retorne ao Passo 3.1. e tente uma opção de dimensionamento alternativa. Por exemplo, uma relação proteína:precipitante diferente pode ser possível ou adicionar sementes se isso não foi feito anteriormente. Quando todos eles estiverem esgotados, pode ser necessário encontrar uma nova condição na Etapa 1.
- Preparar caldo de sementes.
- Escalonamento em placas de 96 poços. A partir do morfograma da endotiapepsina, um método em batelada reta usando a condição de cristalização 0,1 M Tris-HCl pH 7,0, 0,15 M MgCl2 e 30% (p/v) PEG 6.000, foi inicialmente selecionado para raspagem. 100 mg/mL de endotiapepsina misturada ao tampão de cristalização na proporção de 1:1.
Resultados
Otimizando a morfologia cristalina
O passo 1, otimizando a morfologia cristalina, foi incluído para lembrar o leitor de sua importância. Pode ser possível criar microcristais perfeitos a partir de bolas de agulha de baixa difração; no entanto, os autores sugerem que é melhor otimizar os dois separadamente. Primeiro, encontre condições que dão origem a um único cristal através da difusão de vapor e , em seguida, converta essas condições em lote, em vez de tentar combinar as duas etapas juntas. A descoberta de condições altamente nucleantes, nesta fase, não é necessária; morfologia e qualidade de difração são os principais objetivos.
Antes de iniciar a microcristalização da endotiapepsina, foi realizada uma análise das condições de cristalização da estrutura depositada a partir do PDB. Condições de cristalização e protocolos aproximados puderam ser obtidos para 47 das 48 deposições de enthothiapepsina. Estes foram amplamente todos baseados na primeira cristalização de endotiapepsina conduzida por Moews e Bunn (1970)46. Dadas as semelhanças dessas condições e sua origem "clássica", uma tela de matriz esparsa de difusão de vapor de 96 poços foi realizada para explorar uma variedade maior de condições de cristalização. A endotiapepsina foi concentrada a 70 mg/mL e uma tela de matriz esparsa PACT47 foi realizada em uma placa de 96 poços a 20 °C misturando 100 nL de proteína com 100 nL de solução de poço. Todas as condições deste experimento após 36 h deram origem a cristais. No entanto, uma análise da morfologia cristalina indicou que algumas condições podem ser melhores para a otimização da microcristalização.
A Figura 4A mostra uma queda da tela PACT que foi amplamente representativa daquelas observadas na maioria das placas. À primeira vista, pode ser tentador pensar que esses cristais podem valer a pena otimizar ainda mais para a microcristalização. Os cristais são grandes e parece haver nucleação significativa. No entanto, a morfologia cristalina geral não é ideal. Em primeiro lugar, os cristais não são observáveis singletons, pois parece que vários cristais estão crescendo a partir de pontos de nucleação únicos. Em segundo lugar, o tamanho do cristal é altamente assimétrico, com o crescimento ocorrendo principalmente em um único eixo. Tais cristais são teoricamente mais propensos a se alinhar preferencialmente quando entregues ao feixe de raios X. Ambas as características apresentam problemas durante a coleta e processamento de dados cristalográficos seriados.
A Figura 4B, entretanto, mostra cristais de endotiapsina crescidos na presença de MgCl2. Esta morfologia foi consistente em todas as condições que continham MgCl 2 e, portanto, sugeriu que sua morfologia era devida ao MgCl2. As condições de MgCl2 produziram cristais únicos, mais semelhantes a caixas, que representaram um melhor alvo para os experimentos seriais finais.
Havia quatro condições dentro da tela PACT que continham MgCl2. Para melhor compreender a influência de todos os diferentes componentes dessas condições na cristalização da endotiapepsina, uma otimização aleatória foi realizada. Foi criada uma tela contendo uma combinação aleatória dos tampões e precipitantes em uma faixa de concentrações e pHs. A concentração de MgCl2 também foi variada e, em seguida, as gotas resultantes foram arbitrariamente graduadas de 0-5 (sendo 0 sem cristais ou precipitação) em termos de sua qualidade visual de cristais e nível de precipitação.
A Figura 5A mostra um mapa de calor dos resultados de uma análise de correlação de Pearson entre o nível de precipitação e a qualidade do cristal, e as variáveis de tela (exemplos das gotas deste experimento são mostrados na Figura 5B, C e D). Os resultados indicaram que o pH da solução foi altamente correlacionado com o nível de precipitação, com tamponamentos alcalinos resultando em maior precipitação. A concentração de MgCl2 correlacionou-se levemente com o nível de precipitação, assim como o pH e a concentração de precipitante com a qualidade dos cristais.
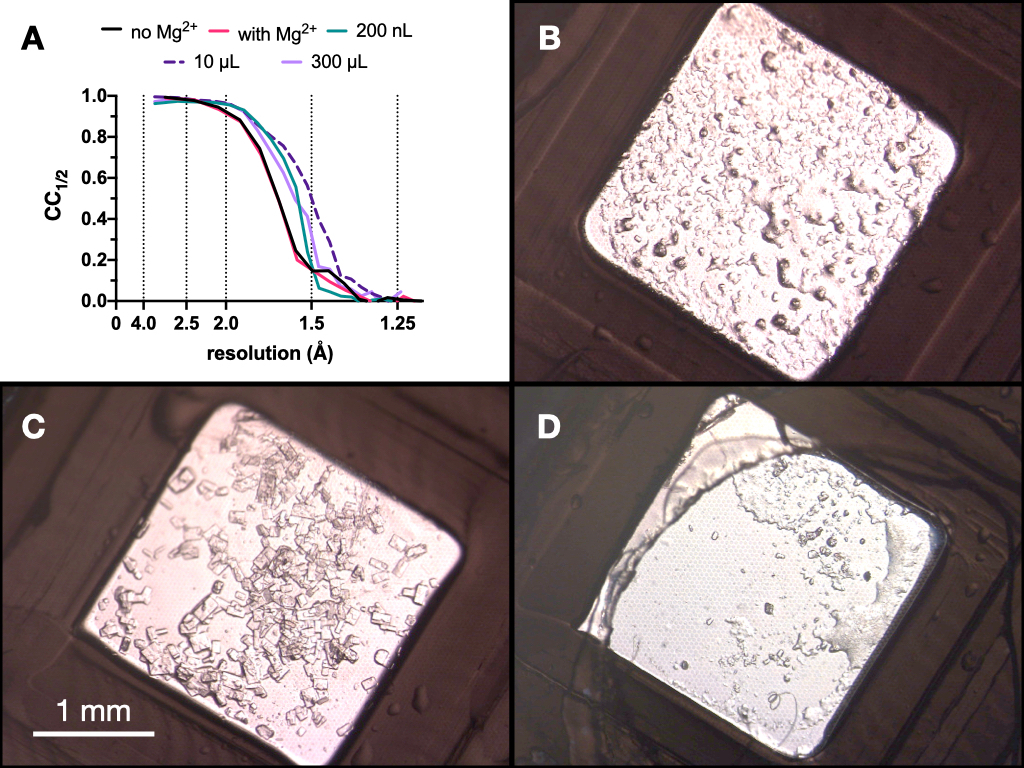
Com base nesses resultados, optou-se por levar os cristais cultivados em Tris-HCl 0,1 M pH 7,0, 0,15 M MgCl2, 20% (p/v) PEG 6.000 para a próxima etapa do protocolo - Transição para batelada. A morfologia dos cristais foi aceitável e uma análise da difração de raios X e das métricas de qualidade dos dados desses cristais sugeriu que não houve diferença significativa entre os cristais crescidos dentro e fora da presença de Mg2+ (Figura 9).
Transição para lote
Para muitas otimizações de microcristalização de cristalografia serial, a Etapa 2 será o ponto de partida. A proteína de interesse já terá sido cristalizada para criocristalografia e o protocolo de cristalização precisará agora de transformação para criar polpas de microcristais. Este protocolo utilizou apenas placas de difusão de vapor de 96 poços para realizar a transformação em batelada, uma vez que a difusão de vapor é o método de cristalização utilizado por 95% das entradas de PDB26. O protocolo evitou a mudança para o microlote34,35,37, uma vez que essa transição ainda poderia incorrer em uma otimização semelhante. Isso não quer dizer que esse protocolo só possa ser feito em placas de difusão de vapor. Todas as etapas apresentadas, também funcionariam em microbatelada se este fosse o método original de cristalização.
Para avaliar a cristalização da endotiapepsina na condição escolhida, foi criado um morfograma - ou diagrama de fase rugosa. O objetivo do experimento do morfograma é triplo. Em primeiro lugar, uma análise do morfograma é de grande utilidade ao avaliar rotas de escala na Etapa 3 - Escala. Em segundo lugar, o morfograma atua como uma ferramenta de otimização, ajudando a descobrir as condições de difusão de vapor que dão origem aos cristais via batelada [ou seja, cristais que aparecem rapidamente (< 24 h)]. Em terceiro lugar, se os cristais não apareceram rapidamente, uma análise das gotas semeadas pode dar ao cristalógrafo uma ideia da localização aproximada da condição atual no diagrama de fases. Por exemplo, se as condições semeadas dão cristais, mas as não semeadas não, é provável que essas condições estejam na região metaestável.
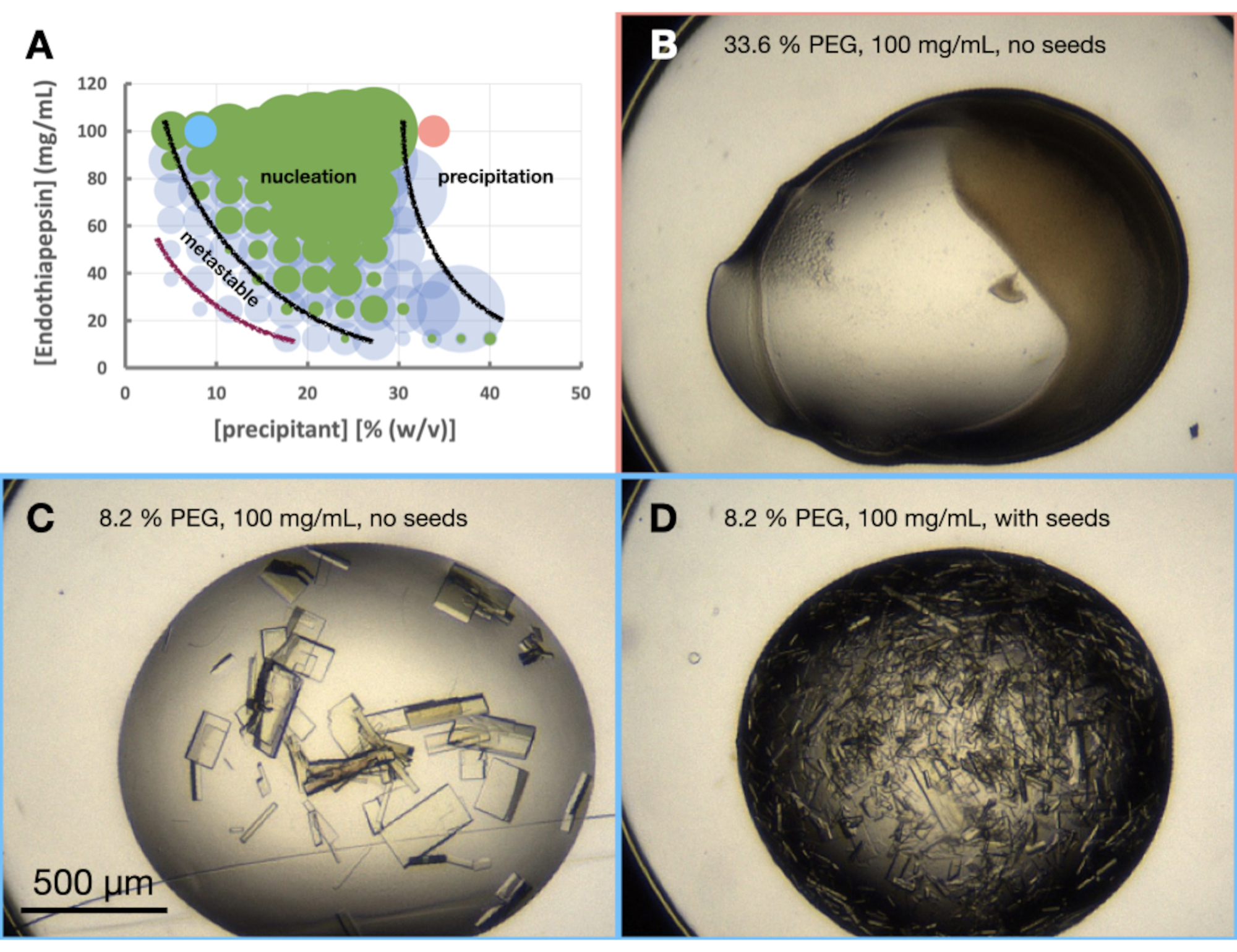
O experimento de morfograma da endotiapepsina foi realizado com base na condição Tris-HCl 0,1 M pH 7,0, 0,15 M MgCl2, 20% (p/v) PEG 6.000. As concentrações de proteína e PEG variaram de 100 a 12,5 mg/mL e 5 a 40% (p/v), respectivamente. As gotas foram analisadas e os resultados plotados por meio da planilha fornecida (Figura 6A).
Também já estava claro no estágio de otimização da morfologia cristalina que o crescimento de cristais de endotiapepsina nessa condição, e nessas concentrações de proteínas, resultaria em cristais crescidos em menos de 24 h. Isso indicou que a cristalização estava ocorrendo através de um lote em vez de um processo de difusão de vapor. Os cristais cultivados nessas condições foram, portanto, adequados para escalonamento para volumes maiores.
Se os cristais não tivessem sido visíveis nas gotas não semeadas após 24 h, então seria provável que a cristalização ainda dependesse de uma transição (Figura 1B) e, portanto, não de um lote. Neste caso, os resultados do experimento do morfograma ainda são de interesse. Eles dão uma indicação do provável ponto de partida para a cristalização no diagrama de fases e, portanto, como a otimização subsequente deve proceder. Olhe para as gotas semeadas. As sementes permitirão o crescimento de cristais na região metaestável, independentemente da nucleação. Por exemplo, se cristais aparecem dentro de 24 h nas gotas semeadas, mas não nas gotas não semeadas, isso indica que parte da região metaestável pode ser observada. Se nenhum cristal for observado nas gotas semeadas ou não semeadas, todos os poços permanecerão subsaturados.
Escala
Observando o morfograma (Figura 6A), algumas observações puderam ser feitas. A quantidade de nucleação parece ser afetada tanto pelas concentrações de proteína quanto pelas precipitantes. Havia também uma demarcação muito clara das gotas que levavam à precipitação proteica, com gotas contendo: nada, cristais ou precipitados (Figura 6B). A adição de sementes (Figura 6D) também aumentou muito Xn quando comparada às gotas sem sementes (Figura 6C). Levando todos esses resultados em conjunto, optou-se por escalar um protocolo de batelada e de lote semeado a 30% (p/v) de PEG 6.000 e 100 mg/mL de endotiapsina.
O escalonamento inicial do teste foi feito em placas suspensas de 24 poços. Os volumes de gota foram aumentados gradualmente para que qualquer mudança no comportamento de cristalização pudesse ser observada (Figura 7). Como pode ser visto, tanto nas gotas não semeadas quanto nas semeadas ocorreu o crescimento de cristais. Todas as gotas não semeadas cresceram uma variedade de tamanhos de cristais, mas predominantemente cristais grandes (100-200 μm - dimensão mais longa). As gotas semeadas, no entanto, produziram cristais menores (5 - 50 μm - maior dimensão). Esses testes iniciais sugeriram que as sementes seriam necessárias para diminuir Xs, mas também, que essa condição deveria ser adequada para volumes maiores.
Quando o volume foi aumentado em 200 μL, o volume de cristalização foi continuamente agitado durante o crescimento do cristal. A principal razão para essa agitação foi garantir que a solução de cristalização permanecesse homogênea e que os cristais em crescimento não se depositassem no fundo ou nas laterais dos tubos. O assentamento de cristais pode levar a uma população heterogênea de cristais com cristais muito grandes e pequenos. A agitação da solução de cristalização também pode promover nucleação44,45.
Infelizmente, o PEG 6.000 não semeado a 30% (p/v) não produziu cristais, de modo que a concentração de PEG foi aumentada para 35% (p/v). Esse aumento melhorou acentuadamente a cristalização, com uma faixa final de Xn e Xs de 3,6 ± 1,2 x 106 cristais·mL-1 e 42 ± 4,1 μm, respectivamente (Figura 8A e B - preto). Apesar de uma melhoria significativa e uma concentração de cristais aceitável, os cristais finais eram muito grandes para o experimento planejado, então novas otimizações foram realizadas. Para reduzir o tamanho dos cristais finais foram exploradas duas vias (Figura 1E): diminuir a concentração de proteína para tentar limitar o crescimento final dos cristais (Figura 8A e B - rosa quente) e aumentar a concentração de PEG para tentar aumentar a nucleação (Figura 8A e B - verde).
A redução da concentração de proteínas, infelizmente, também reduziu drasticamente o Xn, que acabou produzindo cristais ainda maiores. O aumento da concentração de PEG para 40% produziu uma faixa final de Xn eX s de 3,1 ± 0,7 x 106 cristais·mL-1 e 39 ± 2,3 μm, respectivamente. Estes não foram significativamente diferentes dos 35%, mas como o tamanho final do cristal foi reduzido, esta condição foi continuada com as otimizações adicionais.
Para aumentar o Xn, foram adicionadas sementes. Isso aumentou drasticamente o Xn (1,1 ± 1,8 x 108 cristais·mL-1) e levou a um Xs menor (4,2 ± 4,0 μm) (Figura 8A e B - roxo tracejado). Esses cristais, embora muito adequados para alguns experimentos de cristalografia seriada, foram considerados muito pequenos, de modo que a concentração das sementes adicionadas foi alterada.
Essa sintonia do estoque de sementes adicionado, no entanto, mostrou-se difícil de repetir de forma confiável; portanto, tentou-se a extinção. Após a adição de um estoque de sementes, o tamanho do cristal foi monitorado e, uma vez atingido um tamanho de cristal adequado (aproximadamente 10 - 20 μm), a cristalização em batelada foi extinta (Figura 8C e D). A têmpera foi proposta, no que diz respeito à microcristalização, em Kupitz et al (2014)25. Embora talvez não seja um método ideal, uma vez que a solução proteica acabará sendo desperdiçada26, a técnica foi muito útil nessa situação, pois o crescimento de cristais foi difícil de controlar. A ideia por trás da têmpera é retornar rapidamente a mistura de cristalização a um ponto logo acima da linha de solubilidade (Figura 1F). Uma vez que a solução tenha retornado à linha de solubilidade, a solução retornou a uma solução saturada estável e não ocorrerá mais crescimento de cristais.
Tentar extinguir uma reação de cristalização não é isento de riscos. Se for adicionada uma solução de têmpera demasiado grande, a proteína em solução pode ser diluída de tal forma que a linha de solubilidade é ultrapassada. Neste caso, a solução ficará subsaturada e os cristais começarão a se dissolver. Para evitar isso, é possível estimar a quantidade de solução de têmpera necessária com base nos resultados do morfograma. No ponto de têmpera, tome a concentração da solução de proteína. Comparando a concentração de proteína na linha de solubilidade e a concentração de proteína em solução, pode-se estimar a diluição necessária.
A versão temperada do experimento de PEG 6.000 10 x sementes diluídas a 40% (p/v) forneceu uma concentração final de cristais e uma faixa de tamanho de 2,6 ± 3,1 x 106 cristais·mL-1 e 15 ± 3,9 μm, respectivamente.
Durante todo o processo, coletas de dados de raios-X dos cristais de endotiapsina foram coletadas na linha de luz Swiss Light Source PXII usando um foco de 10 x 30 μm, uma energia de 12,4 keV atenuada em 80%, e sob condições criogênicas. Os dados foram processados por meio de mostradores e a Figura 9 mostra a comparação do CC1/2. Nenhuma mudança dramática no CC1/2 foi observada ao longo da otimização.

Figura 1: Uma visão geral da cristalização de transição e de lote e métodos de dimensionamento mapeados em um diagrama de fase. Não. As zonas e limites do diagrama arquetípico de fases de cristalização de proteínas. As concentrações de precipitante e proteína são plotadas nos eixos x e y , respectivamente, com o ponto de água pura na origem. A linha roxa indica o limite de supersaturação da proteína, e as zonas metaestáveis, de nucleação e de precipitação são mostradas em azul, verde e rosa, respectivamente. Não. Um exemplo dos limites de penetração da zona de nucleação de um método de cristalização de "fase de transição", como a difusão de vapor. Neste experimento teórico, a queda precipitante e as concentrações de proteínas começam logo abaixo da linha de solubilidade - ainda não supersaturada. Enquanto a queda se equilibra, as concentrações do componente da gota aumentam de tal forma que a gota se torna supersaturada e continua a se mover - ou fazer a transição - para a zona de nucleação. Após a nucleação do cristal, a concentração de proteína em solução começa a cair. A concentração continua a cair à medida que os cristais crescem até finalmente parar na linha de solubilidade. A linha pontilhada azul marca um limite teórico da transição para a zona de nucleação. Assim que a nucleação começa, a concentração de proteína diminui, impedindo uma maior penetração. Não. Exemplo de trajetórias de cristalização em batelada e em batelada semeada. Em batelada, a mistura da proteína e do precipitante deve criar uma solução supersaturada dentro da zona de nucleação para que o crescimento de cristais possa ocorrer. Em lotes semeados, não é estritamente necessário estar na zona de nucleação devido à adição de micro-sementes, para que locais na região metaestável também possam ser explorados. Não. Uma otimização hipotética do experimento de cristalização mostrado em B da difusão de vapor para batelada. O ponto de partida original da difusão do vapor transitou, através do vetor de otimização resultante, para a nova posição de partida; dentro da zona de nucleação. O vetor resultante é o produto de duas otimizações: um aumento nas concentrações de proteína e precipitante. Não. Otimizações de exemplo ao dimensionar as condições do lote para adaptar o Xn e o Xs finais. Não. Extinção do experimento de cristalização pela adição de tampão de cristalização. É essencial que a têmpera não retire a concentração de proteína da região metaestável e, portanto, abaixo do ponto de supersaturação proteica. Caso contrário, os cristais começarão a se dissolver novamente em solução. Não. e C . foram adaptados de Beale et al (2019)26 com a permissão dos autores. Clique aqui para ver uma versão maior desta figura.
{kind=link}

Figura 2: Aumento de Xn e diminuiçãode X s. A relação idealizada entre o número de cristais produzidos a partir de um experimento de cristalização e sua dimensão média mais longa. Para a criação deste gráfico, foi utilizada a cristalização de uma proteína modelo hipotética de 10 kDa. A proteína cristalizou-se na concentração de 10 mg/mL e produziu cristais P2 1 21 21 com dimensões de 49x50x51 Å. Cada evento de nucleação foi assumido como produzindo um cristal. O crescimento dos cristais foi assumido como homogêneo em todas as faces. Clique aqui para ver uma versão maior desta figura.
{kind=link}

Figura 3: Fluxograma mostrando os passos para otimizar um cristal cultivado em um experimento de difusão de vapor de pequeno volume (<500 nL) em um experimento de grande volume (> de 100 μL). A otimização de cristais é dividida em três etapas: (1) Otimização da morfologia cristalina. (2) Transição para lote. (3) Dimensionamento. Na Etapa 1 é importante identificar cristais adequados para microcristalização. Algumas proteínas estão presentes apenas em uma morfologia de cristal, independentemente da condição de cristalização. No entanto, vale a pena procurar condições que dêem origem a cristais únicos, semelhantes a cubos, ou o mais próximo possível destes. Cristais únicos, semelhantes a cubos, hipoteticamente e anedoticamente, geralmente darão origem a melhores resultados de experimentos de cristalografia seriada. Uma vez selecionada a morfologia cristalina e confirmada a difração, é necessário mover o experimento de cristalização da difusão de vapor para o lote (Etapa 2). Aqui, os cristais devem ser otimizados pelo seu tempo de nucleação. O objetivo é encontrar condições que produzam cristais que aparecem rapidamente (> 24 h), pois essas condições provavelmente atingirão a zona de nucleação imediatamente e, portanto, são em lote. Uma vez que uma condição na zona de nucleação foi encontrada, um morfograma pode ser criado. O morfograma permite que a maior parte da zona de nucleação seja mapeada e potenciais rotas de escala identificadas para o Estágio 3. O volume de uma condição de lote identificada pode então ser gradual ou rapidamente dimensionado em tamanho para produzir um volume final de >100 μL. Clique aqui para ver uma versão maior desta figura.
{kind=link}

Figura 4: Análise das condições de cristalização da endotiapepsina a partir de uma tela de matriz esparsa PACT. A. e B. são fotos após 24 h dos poços A4 e C10, respectivamente, da tela PACT . Os componentes do tampão de cristalização são destacados na figura. O tampão SPG é ácido succínico, dihidrogenofosfato de sódio e glicina misturados em uma proporção molar de 2:7:7. Clique aqui para ver uma versão maior desta figura.
{kind=link}

Figura 5: Análise da otimização da cristalização da endotiapepsina a partir das condições PACT MgCl2 . Não. Um mapa de calor dos resultados de uma análise de correlação de Pearson entre o pH tampão, a concentração de MgCl2 e a concentração de precipitantes e o nível de precipitação e a qualidade do cristal. O nível de precipitação e a qualidade dos cristais foram avaliados arbitrariamente em uma escala de 0-5 (sendo 0 sem cristais ou precipitação) após 24 h. B. C. e D . mostram exemplos da cristalização e precipitação em três gotas diferentes. A condição de cristalização e as avaliações do nível de precipitação e da qualidade dos cristais também são mostradas. Clique aqui para ver uma versão maior desta figura.
{kind=link}

Figura 6: Morfograma de endotiapepsina quando cristalizado em Tris-HCl 0,1 M pH 7,0, 0,15 M MgCl2 e PEG 6.000. Não. Um morfograma criado a partir da planilha "phase-diagram-generator" fornecida. O número relativo de cristais em cada gota é denotado pelo tamanho dos círculos, e os resultados da gota 1 (proteína e precipitante) e da gota 2 (proteína, precipitante e sementes) são destacados em verde e azul, respectivamente. Os valores das concentrações de proteína e precipitante, nos eixos x e y, respectivamente, denotam valores pré-misturados de cada volume, em vez de volumes finais. Com base nos resultados, linhas pretas e uma linha roxa foram desenhadas para mostrar os limites da zona de nucleação e da zona metaestável, respectivamente. B.C. e D. mostram alguns exemplos de resultados do experimento. Os pontos vermelhos e azuis marcados em A. indicam as localizações de B. e C. e D., respectivamente. Clique aqui para ver uma versão maior desta figura.
{kind=link}

Figura 7: Ensaios iniciais de raspagem da endotiapepsina em placas suspensas de 24 poços. As mesmas concentrações de proteína e precipitante foram utilizadas para todas as trilhas: 100 mg/mL de endotiapsina em acetato de 0,1 M Na pH 4,6 e 0,1 M Tris-HCl pH 7,0, 0,15 M MgCl2 e 30% (p/v) PEG 6.000, respectivamente. Todas as imagens exibidas foram tiradas após 24 h e os volumes finais de gota são rotulados em cada imagem. O painel esquerdo (A, D e G) é uma mistura 1:1 de proteína e precipitante, o painel do meio (B, E e H) é uma mistura 1:2:3 de sementes, precipitante e proteína e o painel direito (C, F e I) são imagens ampliadas do painel do meio. Clique aqui para ver uma versão maior desta figura.
{kind=link}

Figura 8: Análise da microcristalização da endotiapepsina em volumes de 200-300 μL. A. e C. mostram como Xn mudou ao longo do tempo de experimento. Não. e D. mostram comoX s (dimensão mais longa) mudou ao longo do tempo. Os resultados dos experimentos foram separados para maior clareza. A linha pontilhada vermelha em C. e D. mostra o ponto em que a têmpera foi realizada. Clique aqui para ver uma versão maior desta figura.
{kind=link}

Figura 9: Resultados de CC1/2 e imagens de cristais obtidas em cada etapa do processo de microcristalização para avaliar a qualidade da difração. Não. CC1/2 plotado contra resolução a partir de dados coletados de cristais cultivados: com e sem Mg - parte da otimização do Estágio 1, em um volume de 200 nL, um volume de 10 μL e o volume final de 300 μL. BC e D. mostram os cristais a partir do volume de 200 nL, 10 μL e 300 μL, respectivamente. Clique aqui para ver uma versão maior desta figura.
{kind=link}
| Informações sobre proteínas | |
| Proteína | Endotiapepsina |
| Peso Molecular (kDa) | 33.8 |
| Grupo espacial | P12 11 |
| a, b, c (Å) | 45.2, 73.3, 52.7 |
| α, β, ɣ (°) | 90.0, 109.2, 90.0 |
| Parâmetros de destino fixo | |
| Volume carregado por chip (μL) | 150 |
| Aperatures por cavaco | 25,600 |
| Concentração de cristais necessária (cristais/mL) | 500,000 |
| Informações de exemplo | |
| Massa proteica utilizada para produzir 200 μL de amostra (mg) | 10 |
| Maior dimensão do cristal (μm) | 15 |
| Concentração de cristais (cristais/mL) | 2,500,000 |
| Variáveis experimentais | |
| Número de pontos de tempo necessários | 5 |
| Número de imagens necessárias por ponto de tempo | 50,000 |
| Taxa de acerto (padrões integrados/imagens coletadas) | 0.3 |
| Metas fixas necessárias por ponto de tempo (arredondadas para cima) | 7 |
| Requisitos de amostra | |
| Volume de amostra necessário por ponto de tempo (μL) | 1,050 |
| Volume total da amostra necessário para o experimento (mL) | 5.25 |
| Massa total de proteína necessária (mg) | 52.5 |
Tabela 1: Um exemplo dos requisitos de amostra para um experimento hipotético de bomba-sonda óptica realizado usando alvos fixos. A proteína utilizada neste experimento teórico foi a endotiapsina. Os parâmetros de alvo fixo foram baseados em experimentos relatados em Ebrahim et al (2019)48 e Davy et al (2019)49. As informações da amostra foram provenientes do protocolo relatado neste artigo em vídeo e as variáveis experimentais foram estimativas conservadoras baseadas na experiência vivida. As seguintes necessidades amostrais foram posteriormente calculadas, tendo em conta os pressupostos anteriores.
Discussão
O método apresentado mostra como otimizar a cristalização da endotiapepsina de grandes cristais (≥ 100 μm de maior dimensão), cultivados em telas de matriz esparsa de 96 poços, para microcristais, cultivados em tubos de centrífuga (volume de 300 μL) via batelada. A ideia por trás do protocolo é que as etapas tomadas para otimizar a endotiapepsina também possam ser usadas para outras proteínas. Em última análise, respondendo ao problema de criar grandes volumes (>100 μL) de micro-cristais (10-20 μm) para experimentos de cristalografia seriada em XFELs e síncrotrons.
O protocolo divide a tarefa de microcristalização de grandes volumes em três etapas: (1) Otimização da morfologia cristalina, (2) Transição para lote e (3) Dimensionamento. Na Etapa 1, a gama de formas cristalinas que uma proteína pode criar deve ser explorada em placas de difusão de vapor. Condições que dão origem a cristais únicos, semelhantes a caixas, que difratizam para a resolução requerida devem ser o objetivo. Na Etapa 2, as condições selecionadas podem então ser transformadas da difusão de vapor em batelada. Aqui, o critério de otimização é o tempo de crescimento dos cristais e encontrar condições que dão origem aos cristais de proteína dentro de 24 h. Um morfograma também pode ser plotado dando ao experimentador uma ideia da localização da linha de solubilidade e limites da zona de nucleação. Este morfograma é de grande utilidade na Etapa 3, Escala. O morfograma dará uma indicação de se a nucleação sozinha pode aumentar Xn e reduzir Xs. À medida que o volume do experimento é aumentado, Xn e Xs podem ser continuamente avaliados como os principais critérios de sucesso de escala.
No caso da endotiapsina, o passo 1 desenterrou o que potencialmente era uma morfologia cristalina até então desconhecida para a endotiapsina. Essa morfologia tinha o mesmo grupo espacial que os relatados anteriormente, mas, importante para a cristalografia serial, uma forma mais parecida com uma caixa. Monocristais também pareciam crescer a partir de pontos de nucleação únicos, ao contrário dos leques criados a partir de outras condições (Figura 4). Para a condição selecionada, o Passo 2 já estava parcialmente satisfeito, pois o crescimento dos cristais ocorreu em < de 24 h. O morfograma indicou que um protocolo de batelada reta ou semeado pode ser bem-sucedido ao escalar na Etapa 3. A escala inicial, em lote reto, criou uma condição que produziu cristais com uma faixade Xn e X s de 3,6 ± 1,2 x 106 cristais·mL-1 e 42 ± 4,1 μm, respectivamente. Esses cristais, embora aceitáveis para alguns experimentos seriados de cristalografia, foram considerados muito grandes. Assim, otimizações adicionais foram realizadas. O protocolo final produziu cristais com uma faixa de concentração e tamanho de 3,1 x 106 cristais·mL-1 e 15 ± 3,9 μm, respectivamente. Isso foi mais do que ideal para os experimentos planejados.
O método se concentra na transformação de cristais de proteína "solúveis" cultivados em placas de difusão de vapor em batelada. A razão para esse foco é que a grande maioria dos cristais de proteínas solúveis é cultivada via difusão de vapor26. No entanto, os conceitos apresentados também podem ser aplicados a cristais de proteína solúvel cultivados usando outros métodos, como micro-batelada. Os conceitos também podem ser aplicáveis a cristais de proteína de membrana cultivados em LCP; pois este também é um processo de cristalização em batelada.
Um aspecto chave do protocolo é o processo de transformação das condições dos cristais crescidos em placas de difusão de vapor para que possam ser cultivados em batelada. Para essa transformação, o método utiliza o critério proposto por Beale e colaboradores (2019)26. Cristais crescidos através de um processo em batelada, mesmo em placas de difusão de vapor, se formarão rapidamente (< 24 h). Este critério é uma aproximação baseada na velocidade de equilíbrio da gota de difusão de vapor, e é mais verdadeiro para condições precipitantes baseadas em PEG. No entanto, as condições de cristalização conterão uma grande variedade de compostos que influenciarão o tempo de equilíbrio. O equilíbrio das condições de cristalização à base de sal, por exemplo, cloreto de amônio altamente concentrado, pode acontecer em 1-2 dias. Portanto, o critério de 24 h pode não ser verdadeiro para condições à base de sal. Condições à base de sal também podem apresentar diagramas de fases mais complexos26,30 que podem não estar de acordo com o arquétipo apresentado neste protocolo. Pode ser necessária uma redução do critério de tempo para condições à base de sal para 12 ou 6 h se a escala em volumes maiores se revelar impossível.
Outra limitação desse método é sua aparente complexidade. O protocolo seguido para otimizar a microcristalização da endotiapepsina realmente mudou relativamente pouco a condição original da tela de matriz esparsa. O primeiro acerto observado na tela PACT foi de 0,1 HEPES pH 7,0, 0,2 M MgCl2 e 20% (p/v) PEG 6.000. O tampão de cristalização escalonado final foi 0,1 Tris-HCl pH 7,0, 0,15 M MgCl2 e 40% (p/v) PEG 6.000. Também é muito possível que a mudança no tampão de HEPES para Tris-HCl, e a concentração de MgCl2 , tenham contribuído pouco para o sucesso do processo. Deixando o aumento na concentração de PEG 6.000 a única otimização, e que poderia ter sido alcançada de forma bem simples.
No entanto, esta avaliação é também demasiado simplista. Ele não apenas desconta os problemas encontrados durante o escalonamento (ou seja , o uso de sementes e têmpera), mas também o fato de que só porque essa proteína se mostrou simples, não há garantia de que a próxima também se provará ser. As etapas aconselhadas no protocolo, foram concebidas porque otimizar a escala de volumes de cristalização de proteínas pode ser muito caro para proteínas. Ao longo dos sete ensaios de escalonamento de endotiapsina que são mostrados, 100 mg de proteína foram consumidos. É certo que algumas dessas etapas foram realizadas para mostrar suas consequências à luz desse protocolo. Mesmo assim, 100 mg de uma proteína, mais potencialmente mais 50 mg de proteína consumida durante um experimento (Tabela 1), podem ser um investimento significativo em tempo ou dinheiro.
Felizmente, não está claro se essa massa de amostra necessária é onipresente em todas as proteínas. A endotiapepsina era altamente solúvel e, portanto, necessitava de uma grande concentração de proteínas para atingir a supersaturação. Em outros (atualmente em fase de otimização), a supersaturação pode ser alcançada em 10 ou até 5 mg/mL. Tais variáveis são específicas de proteínas e precisam ser abraçadas quando aparecem.
Outras limitações do método incluem sua dependência de equipamentos complexos, como robôs de manuseio de líquidos para a criação de telas e placas, e imageadores para criar imagens automáticas de placas quando necessário. Rotinas alternativas têm sido oferecidas para limitar a necessidade de alguns desses equipamentos, mas o protocolo será mais demorado de ser seguido sem eles. O protocolo também sugere testar a difração de cristais otimizados. Para cristalógrafos sem acesso regular a um síncrotron, esses testes podem ser desafiadores. Controles em cada etapa podem não ser necessários, mas esses testes são altamente recomendados uma vez que um acerto tenha sido identificado, e pré e pós-escala. Cristais não difratantes em um XFEL não são, infelizmente, uma ocorrência incomum. Diante disso, é melhor errar do lado da cautela em relação às suposições sobre a difração de cristais.
Em última análise, este protocolo e os resultados apresentados aqui oferecerão um guia, ideias e um exemplo para aqueles que lutam com a produção de amostras para experimentos de cristalografia seriada. Espera-se que, à medida que a cristalografia seriada for mais desenvolvida, as demandas de amostra da técnica sejam reduzidas de tal forma que a necessidade de protocolos como este seja reduzida. No entanto, mesmo neste evento, as estratégias aqui apresentadas ainda serão úteis para aqueles que desejam explorar o espaço de cristalização de sua proteína.
Divulgações
Os autores não têm conflitos de interesse a declarar.
Agradecimentos
Este projeto recebeu financiamento do programa de investigação e inovação Horizonte 2020 da União Europeia ao abrigo do acordo de subvenção Marie Skłodowska-Curie n.º 701647. Muito obrigado pela assistência e apoio dos cientistas da linha de luz da linha de luz suíça X10SA-PXII.
Materiais
| Name | Company | Catalog Number | Comments |
| Swissci 96-well 2-Drop plates | Molecular Dimensions | MD11-002 | 96-well 2-drop crystallisation plate |
| Swissci 96-well 3-Drop plates | Molecular Dimensions | MD11-003 | 96-well 3-drop crystallisation plate |
| mosquito LCP liquid handling robot | sptlabtech | mosquito LCP | Crystallisation robot |
| ClearVue Sheets | Molecular Dimensions | MD6-015 | 96-well crystallization plate seals |
| Safe-Tube 1.5 mL | Eppendorf | 30120086 | 1.5 mL centrifuge tubes |
| Scaple | Swan and Morton | No. 3 scalple and No. 3 handle | Scalple for cutting open plate seals |
| MS 3 Vortex | IKA | 3319000 | Vortex for mixing solution and making seed stocks |
| 24-well XRL Plate | Molecular Dimensions | MD3-11 | 24-well hanging-drop plates |
| Tube revolver/rotator | Thermo Fischer Scientific | 88881001 | Tube revolver for mixing solution during scaling |
| Eppendorf Research plus pipettes | Eppendorf | Range of manual pipettes, 0.5-10, 1-20, 10-100, 100-1000 µL | |
| Eppendorf pipette tips | Eppendorf | Range of tip sizes for manual pipettes | |
| Suparen 600 | Prochem AG | Suparen 600 | Endothiapepsin solution |
| Sodium Acetate | Sigma-Aldrich | 241245-1KG | Sodium Acetate |
| Tris | Merck | 8382T014 | Tris |
| Magnesium Chloride | Sigma-Aldrich | M2670-1kg | Magnesium Chloride |
| PEG 6,000 | Sigma-Aldrich | 81255-1kg | PEG 6,000 |
| Ethelyene glycol | Sigma-Aldrich | 324558-1L | Ethelyene glycol for cyro-protecting the crystals |
| PACT Premier HT screen | Molecular Dimensions | MD1-36 | PACT Premier 96-well crystal screen |
| DOW CORNING high vacuum grease | Molecular Dimensions | MD6-02 | Grease for sealing 24-well plates |
| Hirschmann 22 x 22 mm glaser cover slides | Hirschmann | 8000104 | Cover slides for sealing 24-well sitting drop plates |
| Crystal pins | PSI | Manufactured inhouse | Thin-film supports for micro-crystals. |
| 1-1.3 mm SiLibeads Type S | Faust | 6239547 | Glass beads for making mico-seed stocks |
| Macbook Pro | Apple | Macbook Pro | Computer for performing data analysis |
| CCP4 software suite | CCP4 | Diffraction pattern data processing software | |
| Excel | Microsoft | Microsoft Office | Plotting tool for phase diagram |
| Hausser Scientific Bright-Line counting chamber | Thermo Fischer Scientific | 02-671-51B | Tool to calculate crystal concentration |
| PACT Premier | Molecular Dimensions | MD1-29-ECO | Sparse-matrix crystallization screen |
| Rock Imager | Formulatrix | Rock Imager | Temperature controlled crystal plate storage and imager |
| Rock MakerWeb | Formulatrix | Rock MakerWeb | Crystal plate creation and image storage stoftware |
| Formulator | Formulatrix | Formulator | 96-well crystal screen creation liquid handling robot |
| Leica MZ16 Microscope | Leica | Leica MZ16 | Light microscope |
| LAS V4.6 | Leica | LAS V4.6 | Software for Leica microscopes |
| Spectra/Por 3.5 kDa dialysis tubing | Spectrumlabs | Spectra/Por 3 Dialysis Membrane | 3.5 kDa dialysis membrane |
| Dialysis tubing closures | Spectrumlabs | Spectra/Por 3 Duniversal Closures | Clips to seal the dialysis tubing ends |
| Amicon 10 kDa centrifugal concentrator | Merck-Millipore | Amicon Ultra-15 10 kDa centrifugal concentrator | 10 kDa centrifugal filter |
| 5810 R swing bucket centrifuge | Eppendorf | 5810 R Centrifuge | Swing bucket centrifuge |
Referências
- DePonte, D. P., et al. Gas dynamic virtual nozzle for generation of microscopic droplet streams. Journal of Physics D: Applied Physics. 41 (19), 195505 (2008).
- Hunter, M. S., et al. Fixed-target protein serial microcrystallography with an x-ray free electron laser. Scientific Reports. 4 (1), 6026 (2014).
- Weierstall, U., et al. Lipidic cubic phase injector facilitates membrane protein serial femtosecond crystallography. Nature Communications. 5 (1), 1-6 (2014).
- Roessler, C. G. G., et al. Acoustic Injectors for Drop-On-Demand Serial Femtosecond Crystallography. Structure. 24 (4), 631-640 (2016).
- Sherrell, D. A., et al. A modular and compact portable mini-endstation for high-precision, high-speed fixed target serial crystallography at FEL and synchrotron sources. Journal of Synchrotron Radiation. 22 (6), 1372-1378 (2015).
- Roedig, P., et al. A micro-patterned silicon chip as sample holder for macromolecular crystallography experiments with minimal background scattering. Scientific Reports. 5 (1), 1-11 (2015).
- Botha, S., et al. Room-temperature serial crystallography at synchrotron X-ray sources using slowly flowing free-standing high-viscosity microstreams. Acta Crystallographica Section D Biological Crystallography. 71 (2), 387-397 (2015).
- Weinert, T., et al. Serial millisecond crystallography for routine room-temperature structure determination at synchrotrons. Nature Communications. 8 (1), 542 (2017).
- Tenboer, J., et al. Time-resolved serial crystallography captures high-resolution intermediates of photoactive yellow protein. Science. 346 (6214), 1242-1246 (2014).
- Nango, E., et al. A three-dimensionalmovie of structural changes in bacteriorhodopsin. Science. 354 (6319), 1552-1557 (2016).
- Suga, M., et al. Light-induced structural changes and the site of O=O bond formation in PSII caught by XFEL. Nature. 543 (7643), 131-135 (2017).
- Mehrabi, P., et al. Liquid application method for time-resolved analyses by serial synchrotron crystallography. Nature Methods. 16 (10), 979-982 (2019).
- Halle, B. Biomolecular cryocrystallography: structural changes during flash-cooling. Proceedings of the National Academy of Sciences of the United States of America. 101 (14), 4793-4798 (2004).
- Fraser, J. S., et al. Hidden alternative structures of proline isomerase essential for catalysis. Nature. 462 (7273), 669-673 (2009).
- Fenwick, R. B., van den Bedem, H., Fraser, J. S., Wright, P. E. Integrated description of protein dynamics from room-temperature X-ray crystallography and NMR. Proceedings of the National Academy of Sciences of the United States of America. 111 (4), 445-454 (2014).
- Keedy, D. A., et al. Mapping the conformational landscape of a dynamic enzyme by multitemperature and XFEL crystallography. Elife. 4, (2015).
- Thomaston, J. L., et al. XFEL structures of the influenza M2 proton channel: Room temperature water networks and insights into proton conduction. Proceedings of the National Academy of Sciences of the United States of America. 114 (51), 13357-13362 (2017).
- Haas, D. J., Rossmann, M. G. Crystallographic studies on lactate dehydrogenase at -75 °C. Acta Crystallographica Section B Structural Crystallography and Crystal Chemistry. 26 (7), 998-1004 (1970).
- Hope, H. Cryocrystallography of biological macromolecules: a generally applicable method. Acta Crystallographica Section B Structural Science. 44 (1), 22-26 (1988).
- Wu, W., et al. Batch crystallization of rhodopsin for structural dynamics using an X-ray free-electron laser. Acta Crystallographica Section:F Structural Biology Communications. 71 (7), 856-860 (2015).
- Ishchenko, A., Cherezov, V., Liu, W. Preparation and delivery of protein microcrystals in lipidic cubic phase for serial femtosecond crystallography. Journal of Visualized Experiments. (115), e54463 (2016).
- Andersson, R., et al. Well-based crystallization of lipidic cubic phase microcrystals for serial X-ray crystallography experiments. Acta Crystallographica Section D: Structural Biology. 75 (10), 937-946 (2019).
- Luft, J. R., et al. The detection and subsequent volume optimization of biological nanocrystals. Structural Dynamics. 2 (4), 041710 (2015).
- Lee, D. B., et al. Supersaturation-controlled microcrystallization and visualization analysis for serial femtosecond crystallography. Scientific Reports. 8 (1), 1-10 (2018).
- Kupitz, C., et al. Microcrystallization techniques for serial femtosecond crystallography using photosystem II from Thermosynechococcus elongatus as a model system. Philosophical transactions of the Royal Society of London. Series B, Biological Sciences. 369 (1647), 20130316 (2014).
- Beale, J. H., et al. Successful sample preparation for serial crystallography experiments. Journal of Applied Crystallography. 52, 1385-1396 (2019).
- Rayment, I. Small-scale batch crystallization of proteins revisited: An underutilized way to grow large protein crystals. Structure. 10 (2), 147-151 (2002).
- García-Ruiz, J. M. Nucleation of protein crystals. Journal of Structural Biology. 142 (1), 22-31 (2003).
- McPherson, A., Kuznetsov, Y. G. Mechanisms, kinetics, impurities and defects: Consequences in macromolecular crystallization. Acta Crystallographica Section F:Structural Biology Communications. 70 (4), 384-403 (2014).
- Rupp, B. Origin and use of crystallization phase diagrams. Acta crystallographica. Section F, Structural biology communications. 71, 247-260 (2015).
- Luft, J. R., DeTitta, G. T. A method to produce microseed stock for use in the crystallization of biological macromolecules. Acta Crystallographica Section D Biological Crystallography. 55 (5), 988-993 (1999).
- Ireton, G. C., Stoddard, B. L. Microseed matrix screening to improve crystals of yeast cytosine deaminase. Acta crystallographica. Section D, Biological crystallography. 60 (3), 601-605 (2004).
- Forsythe, E. L., Maxwell, D. L., Pusey, M. Vapor diffusion, nucleation rates and the reservoir to crystallization volume ratio. Acta Crystallographica Section D Biological Crystallography. 58 (10), 1601-1605 (2002).
- Chayen, N. E., Shaw Stewart, P. D., Maeder, D. L., Blow, D. M. IUCr An automated system for micro-batch protein crystallization and screening. Journal of Applied Crystallography. 23 (4), 297-302 (1990).
- Chayen, N. E., Shaw Stewart, P. D., Blow, D. M. Microbatch crystallization under oil - a new technique allowing many small-volume crystallization trials. Journal of Crystal Growth. 122 (1-4), 176-180 (1992).
- Chayen, N. E. Comparative studies of protein crystallization by vapour-diffusion and microbatch techniques. Acta Crystallographica Section D: Biological Crystallography. 54 (1), 8-15 (1998).
- D'Arcy, A., Mac Sweeney, A., Stihle, M., Haber, A. The advantages of using a modified microbatch method for rapid screening of protein crystallization conditions. Acta Crystallographica - Section D Biological Crystallography. 59 (2), 396-399 (2003).
- Darmanin, C., et al. Protein crystal screening and characterization for serial femtosecond nanocrystallography. Scientific Reports. 6, 25345 (2016).
- Gati, C., et al. Atomic structure of granulin determined from native nanocrystalline granulovirus using an X-ray free-electron laser. Proceedings of the National Academy of Sciences of the United States of America. 114 (9), 2247-2252 (2017).
- Cheng, R. K. Y. Towards an optimal sample delivery method for serial crystallography at XFEL. Crystals. 10 (3), 215 (2020).
- Schmidt, M. Mix and Inject: Reaction Initiation by Diffusion for Time-Resolved Macromolecular Crystallography. Advances in Condensed Matter Physics. 2013, 1-10 (2013).
- Oberthuer, D., et al. Double-flow focused liquid injector for efficient serial femtosecond crystallography. Scientific Reports. 7 (1), 44628 (2017).
- McPherson, A., Gavira, J. A. Introduction to protein crystallization. Acta crystallographica. Section F, Structural biology communications. 70, 2-20 (2014).
- Yaoi, M., et al. Effect of stirring method on protein crystallization. Japanese Journal of Applied Physics, Part 2: Letters. 43 (10), 1318 (2004).
- Castro, F., Ferreira, A., Teixeira, J. J., Rocha, F. Influence of Mixing Intensity on Lysozyme Crystallization in a Meso Oscillatory Flow Reactor. Crystal Growth & Design. 18 (10), 5940-5946 (2018).
- Moews, P. C., Bunn, C. W. An X-ray crystallographic study of the rennin-like enzyme of Endothia parasitica. Journal of Molecular Biology. 54 (2), 395-397 (1970).
- Newman, J., et al. Towards rationalization of crystallization screening for small- to medium-sized academic laboratories: the PACT/JCSG+ strategy. Acta crystallographica. Section D, Biological. 61, 1426-1431 (2005).
- Ebrahim, A., et al. Resolving polymorphs and radiation-driven effects in microcrystals using fixed-target serial synchrotron crystallography. Acta Crystallographica Section D Structural Biology. 75, 151-159 (2019).
- Davy, B., et al. Reducing sample consumption for serial crystallography using acoustic drop ejection. Journal of Synchrotron Radiation. 26 (5), (2019).
Reimpressões e Permissões
Solicitar permissão para reutilizar o texto ou figuras deste artigo JoVE
Solicitar PermissãoThis article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. Todos os direitos reservados