
Method Article
Vibratome секционирования мыши Retina Подготовка фоторецептора культур
В этой статье
Резюме
Нейронные сетчатки мыши в возрасте 8 дней на вершине 4% желатина блока. После выделения фоторецепторов слоя (200 мкм) по vibratome, фоторецепторы высевают после механической и ферментативной диссоциации культуры. Фоторецептор слой может быть использован для молекулярных, биохимических анализов или трансплантации.
Аннотация
Сетчатка часть центральной нервной системы, которые организовал архитектуру, с нейронами в слоях от фоторецепторов, как палочки и колбочки в контакте с сетчатки пигментного эпителия в самой дальней части на сетчатке, учитывая направление света, и ганглиозных клеток в наиболее проксимальных расстояния. Эта архитектура позволяет изолировать фоторецепторов слоя vibratome срезов. Рассекали нейронной сетчатка мыши в возрасте 8 дней является плоским встроен в 4% желатина сверху ломтиком 20% фоторецептора слой желатина вниз. Использование vibratome и обоюдоострым лезвием бритвы, толстый внутренний сетчатки 100 мкм срезы. Этот раздел содержит ганглиозных клеток и внутренний слой со значительно биполярных клеток. Посредником часть 15 мкм отбрасывается до 200 мкм от внешней сетчатки, содержащей фоторецепторов извлекают. Желатин удаляют нагреванием при температуре 37 ° С. Кусочки наружного слоя являются incubaTed в 500 мкл раствора Рингера с 2 единицами активированного папаина течение 20 мин при 37 ° С. Реакцию останавливали добавлением 500 мкл 10% фетальной телячьей сыворотки (FCS), в Дульбекко в модификации Дульбекко (DMEM), затем 25 единиц ДНКазы I добавляют перед центрифугированием при комнатной температуре, промывают несколько раз, чтобы удалить сыворотку, и клетки ресуспендировали в 500 мкл DMEM и высевают в 1 × 10 5 клеток / см 2. Клетки выращивали до 5 дней в пробирке и их жизнеспособность набрал использованием живой / мертвый анализа. Чистоту культуры сначала определяется с помощью микроскопического наблюдения в ходе эксперимента. Чистота затем подтверждено путем посева и фиксации клеток на гистологическом слайд и анализа с использованием кроличьих поликлональных анти-SAG, фоторецептора маркер и мышиные моноклональные анти-RHO, стержень фоторецепторов специфическим маркером. С другой стороны, фоторецептор слой (97% стержней) могут быть использованы для анализа экспрессии гена или белка, и для трансплантации.
Введение
Сетчатка является неотъемлемой частью центральной нервной системы, имеющей архитектуру консервативный среди позвоночных. Нейроны нервной сетчатки организованы в слои, причем наиболее удаленная для падающего света, слой фоторецептора в тесном контакте с пигментного эпителия сетчатки (RPE) в задней части глаза. Палочек и колбочек photorecptors являются светочувствительные клетки, которые полагаются на Opsin чувствительных молекул для захвата фотонов. Эти молекулы заключены на дисковых мембран клеточной структуры, расположенных на внешней сегмента фоторецептора, который указывает в направлении ПЭС 1. Эта структура, которая наиболее часто влияет очень рано в случаях фоторецепторов вырождения, обновляется со скоростью 10% каждый день. Так называемый внутренний слой содержит большинство других нейронов, которые вычисляют сигнал, полученный от фоторецепторов, биполярного, амакринных и горизонтальных клеток, а также ганглиозных клеток. Эти последние с их аксоновобразуют пучок, который зрительный нерв. Это наслоение так консервативными, что биологи использовали термин перемещенных амакринные клетки, когда клетки находятся за пределами внутреннего плексиформной слоя 2. Слои нейронов распределяются внутри арматуры радиальная Мюллер глиальных клеток. Биполярные клетки связывают фоторецепторов ганглиозных клеток. Они расположены между внешним слоем плексиформной и внутренним слоем плексиформной. Ганглиозных клеток образуют внутренний слой плексиформную в связи с биполярными клетками. В amacrin клетки называются ассоциации клеток, расположенных во внутреннем плексиформной слоя между биполярных клеток и ганглиозных клеток. Внешняя сетчатый слой содержит горизонтальные клетки. Это уникальное расположение слоев нейронов центральной нервной системы позволяет выделить слой фоторецептора от внутреннего слоя клеток путем нарезки плоского монтажа сетчатки с использованием vibratome.
Первоначально этот метод был использован для выделения фоторецепторы для Transplantation в глазах RD1 мышь, модели человеческого пигментный ретинит (RP) 3. RD1 мыши несет рецессивный мутацию в гене, который кодирует Pde6b для стержня-специфической фосфодиэстеразы бета-субъединицы. Рецессивные мутации этого результата генов в РП в людях 4. После стержень фоторецепторов выродились, пациент теряет ночного видения, и удивительно колбочек, которые не экспрессируют мутантный ген, вырождаются в качестве второго шага. Потому что конусы необходимы для цветового зрения и остроты зрения, пациенты постепенно становятся слепыми и эффективным средством для лечения болезни еще не разработаны. Прививкой фоторецептора слой из мыши дикого типа конус дегенерацию принимающей мыши задерживается 3,5. Стержни потерянных в стержне конуса дегенеративных модели не могут быть заменены трансплантации, поскольку синаптической связи между стержнями и биполярных клеток могут быть получены только при определенной стадии гetinal развития, отмечен началом NRL выражения 6. Слой фоторецепторов вводится хирургии в суб-сетчатки пространству стержня менее RD1 сетчатки, между ПЭС и внешней сетчатки, соответствующей только 3% из оставшихся фоторецепторов, конусов. Через две недели после операции, 40% из конусов из животного с трансплантированной нормальной слой фоторецептора выжили по сравнению с животного с трансплантированной нормальной слоем внутренних клеток сетчатки или с ложной операцией животного. Рельеф выживание конуса, распространяется из-за всей поверхности мутантного сетчатки, расположенной в положении пересаженной ткани указывает на то, что защитный эффект обусловлен диффундирующего молекулы 7.
Далее, мы использовали систему со-культуры, а также кондиционированной культуральной носитель для подтверждения того, что защитный эффект основан на секрецию белка стержнями 8,9. Мы предполагаем, что этот белок будет выражениеessed постоянно и, в частности стержнями и что их смерти в течение первого этапа заболевания вызовет вторичное конуса дегенерации с потерей защитного сигнала от стержней в не-клеточной автономном режиме 10. Из-за важности конуса опосредованной центрального зрения у приматов это предполагаемый белок будет весьма актуальным терапевтическим средством для РП. Сохранение конусы в РП бы теоретически допустить в общей сложности 1,5 млн пациентов во всем мире, чтобы стать слепым 11. Мы использовали высокое содержание подход скрининга и модель культуры конуса, обогащенные определить кДНК, кодирующей Род полученных Конус жизнеспособности фактор (RdCVF) от сетчатки библиотеки кДНК 12. RdCVF является сращивания продукт гена, который NXNL1, интересно гомологичный ген, кодирующий тиоредоксина белков, участвующих в окислительно-восстановительного гомеостаза 13. Второй сращивания продукт гена, RdCVFL является фермент, который защищает свою цель, тау-белок от окислительного DAmage 14. Администрация RdCVF предотвращает вторичное вырождение конусов и потери их зрительной функции в рецессивным, и доминирующее модель RP 12,15. Это свидетельствует о двух важных аспектов этой инновационной стратегии лечения 16. Во-первых, он может быть применен в большинстве случаев RP в гена-независимым способом. Во-вторых, в отличие от конкурирующих фактора CNTF, RdCVF выживание связано с содержанием зрительной функции 17. Отсутствие функциональной эффекта может объяснить причину отсутствия клинического эффекта от введения CNTF РП пациентов 18. RdCVF, скорее всего, одним из важных сигнала выживания между палочками и колбочками, так как конус спасение в пробирке подавляется RdCVF immunodepletion 12. Кроме того, нарушение гена конуса жизнеспособности стержня, полученных приводит к дисфункции фоторецептора и восприимчивость к окислительному стрессу 19.
Использованиефоторецепторов слой находится в начале координат идентификации RdCVF и нового сигнала окислительно-восстановительного участие в нейродегенеративных заболеваний 20. Эта рукопись описывает протокол, используемый для выделения и культивирования клеток из фоторецепторов слоя характеризуют деятельность RdCVF. Фоторецепторы можно поддерживать в культуре в течение 5 до 7 дней 21. Этот метод также может быть использован для изучения экспрессии специфических генов фоторецепторов.
протокол
ПРИМЕЧАНИЕ: процедура была одобрена комитетом по этике Дарвина из Университета Пьера и Марии Кюри (СЕ5 / 2009/048)
1. Подготовка раствора желатина, инструменты, Vibratome, медиакультуры и культуры плиты
- Подготовьте 20% раствора желатина по крайней мере, за один день до эксперимента.
- Под крышкой, добавить 500 мкл гентамицина [10 мг / мл] в бутылки емкостью 500 мл, содержащий СО 2 независимое среду (CO 2) -i.
- В отдельном стакане, добавляют 20 мл CO2-I (с шагом 1.1.1) и нагревают при 95 ° С на нагревательном блоке при постоянном перемешивании в течение 15 мин. Заметим, изменение цвета на желтый, что свидетельствует о том, что раствор готов для дальнейшего использования.
- Взвешивание 4 г желатина и постепенно бедных к раствору полученного выше (этап 1.1.2) с увеличенной перемешивании и нагревании при 95 ° С в течение 45 мин, чтобы получить раствор желтого цвета. Дайте раствор желатина, чтобы охладить подкапот течение примерно 5 минут.
- Налейте 4 мл раствора желатина в 35 мм чашки для культивирования диаметром без введения пузырьков воздуха. Хранить блюдо при 21 ° С в течение 30 мин. Обложка посуду пластиковой пленкой, и превратить блюда с ног на голову. Возможно, хранить посуду на 4 ° C в течение 2 месяцев до начала использования.
- Подготовка 4% раствор желатина.
- Под капотом, добавить 25 мл CO 2 -i среды в стерильный пластиковый контейнер и тепла при 42 ° С на нагревательном блоке. Удалить среду из нагревательного блока и постепенно добавить 1 г желатина в горячую среду и перемешать. Быстро вернуть контейнер на нагревательный блок (42 ° C) и держать в тепле во время процесса.
- Настройку в vibratome аппарат:
- Удалить посуду, содержащие 20% раствор желатина хранили при 4 ° С. Вырезать желатин с помощью скальпеля. Переверните желатин кусочек и приклейте его на черном опорного диска в vibratome с помощью капли супер клей.
- Перерыв лезвие на две половины, и вставьте одну половину в vibratome проведения розетку. Вставьте черный поддержки диск на vibratome аппарата. Включите черную ручку с правой стороны. Закрепите держит сосуд на голове vibratome. Добавьте достаточное количество CO 2 -i среды в vibratome бака для покрытия желатина блок.
- Включите vibratome и сократить три 100 мкм ломтики желатина блока. Держите желатин блок для дальнейшего использования.
- Подготовка 40 мл культуральной среды:
- Подготовка модифицированный Дульбекко среде Игла (DMEM) с 10% эмбриональной телячьей сыворотки (FCS) под капотом. Подготовка 1 мг / мл поли-D-лизина с использованием фосфатный буфер солевой раствор (PBS), рН 7,4.
- В 96-луночный планшет, добавьте 2 мкг / см 2 поли-D-лизина, чтобы покрыть дно лунок. Инкубируют планшет в течение 45 мин при 37 ° С в течение 5% CO 2 инкубаторе. После инкубации удалить PBS и заменить 200 мкл/ А DMEM и место снова при 37 ° С в 5% СО 2 инкубатора перед использованием.
2. Вскрытие всей сетчатки
- Выяснять мыши:
- Жертвоприношение мышь в соответствии с Европейской директивой 2010/63: путем смещения шейных позвонков. Отрежьте голову изогнутые ножницы и поместите его в 100 мм Диаметр чашки Петри после дезинфицировать глаз с дезинфицирующим средством.
- Снимите глаз после отдельностоящий зрительный нерв секционирования очень осторожно, используя изогнутый сцепление. Затем поместите оба глаза в 35 мм чашки Петри, содержащую стерильный CO 2 -i среды при 21 ° С.
- Удаление сетчатки:
- Сделать отверстие на уровне глаз конечности с помощью 18 G иглы, чтобы иметь возможность ввести ножницы в глаз земного шара. Представьте прямые ножницы в отверстие и аккуратно вырезать склеры ниже радужной оболочки на всем периметре глазного шара, чтобы удалить цилиарного тела.
- Удалить роговицы и объектив и держать заднюю камеру глаза с сетчаткой, прилегающей к склеры. С двух прекрасных ручек, удалить стекловидное тело находится внутри задней камеры без повреждения сетчатки. Выполните четыре радиальных разрезов (сетчатка / склеры), чтобы разрешить уплощение сетчатки.
- Подъем глазной шар и очистить его как «оранжевых». Снимите сетчатку от склеры и сетчатки пигментного эпителия (РПЭ). Снимите сетчатку еще привязаны на уровне зрительного нерва очень аккуратно с мелкими изогнутые ножницы.
- Отделить оставшиеся стекловидного от периферии к центру сетчатки с двух тонких захватов. Убедитесь, что все стекловидное тело полностью удаляется, чтобы позволить уплощение сетчатки.
- Отрежьте оконечность пластиковой пипетки и передачи сетчатки в 35 мм Диаметр чашки Петри, содержащей CO 2 -i среду с пластиковой пипетки.
3. Уплотнение Flat-mounted Retina на Желатин блока
- Удалить 20 - 30 мл CO 2 -i среды от vibratome бака (желатин блок свободен), чтобы разрешить уплотнение плоской монтажа сетчатки.
- Передача сетчатки в стекле с помощью пластиковой пипетки и добавить каплю CO 2 -i среднего до передачи желатина ломтик с фоторецептора вниз на желатиновой среза.
- Приложить сетчатки в желатиновую блока, осторожно инъекционных нагретого 4% желатина на одной стороне блока между сетчаткой и желатина блока и одновременно исключении теплый желатин на другой стороне.
- Отберите 4% желатина с пипетки Пастера. Подождите 10 мин, чтобы позволить полную герметизацию. Добавить CO 2 -i среду, чтобы погрузить блок и лезвие полностью.
4. Структурирование Сетчатка
Примечание: Этот шаг является критическим для получения слоя фоторецепторных клеток без других клеток сетчатки (фиг.1).
- Начиная сверху на желатиновой блока, вырезать 100 мкм серийных срезов для достижения сетчатки. Затем вырезать 100 - 120 мкм секции внутреннего слоя. В зависимости от применения, этот слой может быть отброшены или хранили в жидком азоте. Убедитесь, что скорость vibratome очень медленно (в 1 или 2) в течение секционирования внутренний слой или слои фоторецепторов.
- Выполнение промежуточных 15 мкм секций сетчатки, соответствующей границе между внутренней и внешней сетчатки. Соблюдайте этот раздел под микроскопом. Отсутствие кровеносных сосудов указывает, что внешний сетчатки было достигнуто.
- Вырезать 200 мкм внешней сетчатки (фоторецепторов слой) с желатином и перенести его на 35 мм блюдо диаметром, заполненный 3 мл CO 2 -i среды. Держать его на льду, пока все фоторецептор слой секции всех сетчатки не получают.
5. разлетным клетки фоторецепторов
- Возьмите блюдо (ES), содержащий фоторецепторов слой (слои) и положить его (их) в термостате при 37 ° С в течение 10 мин.
- Использование щипцов аккуратно отделить фоторецепторов слой из желатина и передать его в новое блюдо, наполненный 3 мл раствора Рингера. Повторите этот шаг (5.2) для каждого препарата фоторецепторов слоя.
- Инкубируют 2 единиц папаина с 25 мкл раствора активатора (1,1 мМ ЭДТА; 5,5 мМ L-цистеина, 60 мкМ β-меркаптоэтанол) в 5 мл стерильной пробирке из полипропилена и инкубировать ее 30 мин при 37 ° С в течение 5% CO 2 инкубаторе , Активация папаина достигается при 37 ° С. Во время этой инкубации, вырезать фоторецепторов слой в 2 мм 2 штук (не слишком малых) и передать эти куски в 5 мл пробирки.
- Промыть сетчатки в два раза с 1,5 мл раствора Рингера с последующим гравитационным осаждением. Удалить все остатки раствора Рингера из трубки с образцом.
- Добавить 475 мкл раствора Рингера в пробирку, содержащую активированный папаин и перемешать. Добавить этот раствор в пробирку, содержащую сетчатку.
- Инкубируйте пробирку с сетчатки в течение 20 мин при 37 ° С в течение 5% CO 2 инкубаторе. Остановить реакцию, добавив 1 мл 10% FCS в DMEM. Добавить 25 ЕД дезоксирибонуклеазы I (ДНКазы I), чтобы расщепленной ДНК из смерти клеток. Тщательно гомогенизации суспензии клеток с использованием 1-мл пипетки. Спина в 50 мкг в течение 6 мин при комнатной температуре.
- Жидкость над осадком сливают, чтобы удалить следы сыворотки DMEM и добавить среду с добавками (см таблицу конкретной reagents_equipment) к клеточной гранул и ресуспендируют тщательно клеточной суспензии с 1 мл пипетки. Спина в 50 мкг в течение 6 мин при комнатной температуре. Повторите этот шаг (5,7) еще раз.
6 Культивирование клеток фоторецепторов
- Фоторецептор слой мыши на PN8 содержит примерно от 1 до 1500000 клеток с использованием этой методики. Семенной клетки фоторецепторов в 1х 10 5 клеток / см 2 в культуральной пластины с питательной среды и культуры клеток в течение 5 дней при 37 ° С в течение 5% CO 2 инкубаторе. На 5-й день, перейти к иммунохимии и Вестерн-блоттинга исследования следующие методы, предоставленных поставщиками антитела.
ПРИМЕЧАНИЕ: Шаги, чтобы остановить культуры и предварительные тесты описаны в работе 21.
Результаты
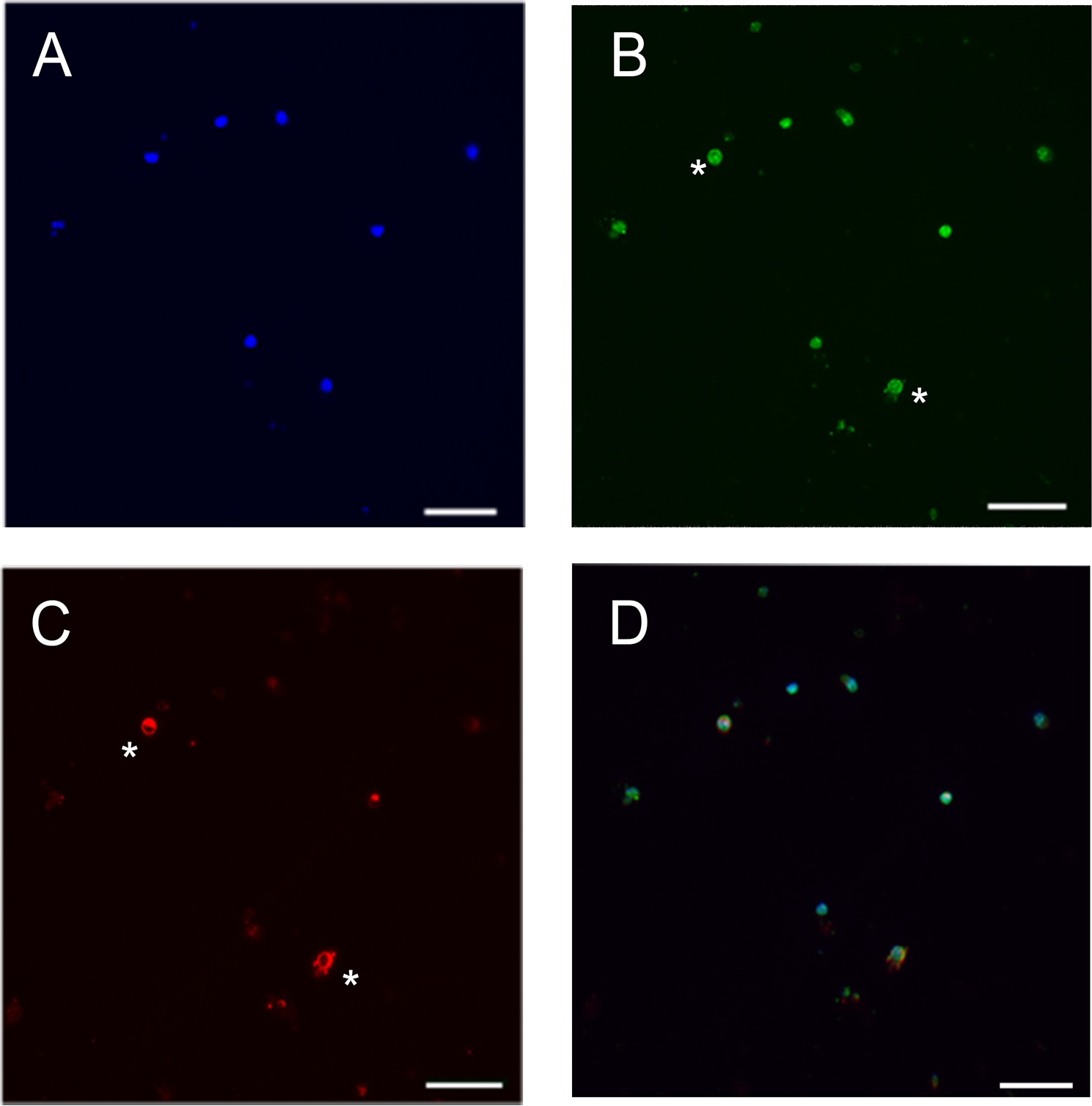
Помимо трансплантации, фоторецептора слои также были использованы для изучения клеточной сигнализации путем посева клеток для изготовления фоторецепторов культур 12,22. Кроме того, они используются для изучения экспрессии генов и циркадный ритм 23,24. Мы использовали клетки фоторецептора слой из мыши дикого типа в послеродовой день 8, чтобы подготовить фоторецепторов культуры в отсутствие FCS и клетки поддерживали в течение 5 дней при 37 ° С в инкубаторе с 5% CO 2 (фиг 1E ). Клетки затем фиксировали с использованием 4% параформальдегиде, а затем продолжалась иммуногистохимического анализа с использованием методов, представленных поставщиками антител либо с мышиным моноклональным анти-RHO (1: 250, Millipore Mab5316) или кроличьей поликлональной анти-СА (1: 200, щедрым Дар Игаль Геры и Дэвида Хикса). Мы использовали анти-RHO и анти-SAG (хотя анти-SAG этикеток колбочек и палочек) в связи с тем, что анти-провисания ранее маркер, чем анти-Ро. Доля челLs положительные для антитела анти-RHO ниже, чем у позитивным для анти-SAG антител (рисунок 2). Это можно объяснить тем фактом, что СА, стержень arrestin также выраженную конусов, которые делают 3% от фоторецепторов в изолированном слое или, альтернативно, с тем, что в ходе постнатального созревания сетчатки, выражение СА предшествует Ро 25,26.
Фоторецептор слой также используется для мониторинга специфическую экспрессию генов путем фоторецепторов дикого типа сетчатки и головного мозга животных в возрасте 35 дней РНК получали с использованием CsCl ультрацентрифугирования 27 и гибридизовали с ДНК-чип мыши массива. В мессенджеры для родопсина (Rho), S-arrestin (SAG) и конус-трансдуцин (Gnat2) выражены в частности, в сетчатке по сравнению с мозга (фиг.3А). Выражение видное место в фоторецепторов слоя (PR), чем в целом по сетчатке, которая охватываетВнутренняя слоя сетчатки, показывая, что эти гены экспрессируются Действительно, по фоторецепторов. Для Gnat2, относительно выражение по сравнению с Ро и провисать показывает, что выражается конусов, содержащихся в изолированной фоторецепторов слоя. Выражение рековерина (Rcvrn) в слое фоторецептора по сравнению с всей сетчатки увеличивается (фигура 3В).
Фоторецепторов слой также используется для отслеживания выражение Ро и GNAT2 с помощью Вестерн-блоттинга следующих способов, предусмотренных поставщиков антител и сравнить с внутренней слоя сетчатки (рис 4). Обратите внимание на отсутствие ГУР и GNAT2 28 маркеров палочек и колбочек, соответственно, в целом сетчатки и в сетчатке RD1 мыши на послеродовой день 35.

Fiфигура 1. Схематическое изображение vibratome секционирования мыши сетчатки. () Установка плоской монтажа сетчатки с фоторецепторов вниз на желатиновой среза. (B) Раздел внутреннего сетчатки с vibratome лезвия. (C) Раздел внешней сетчатки добавил желатина. (D) Контроль Наличие фоторецептора на краю фрагмента сетчатки после выделения фоторецептора слой. Масштабная линейка используется для зеленой флуоресценции используется белого светового микроскопа. Фоторецепторы расположены на краю изображения. (E) Искусственный клетки фоторецепторов (пост-пять дней). (F) Высшее Увеличение панели Е.

Рисунок 2. Дифференциальная экспрессия ГУР и СА в мышь фоторецепторов культуры. () Окрашивание ядер (синий), (В) СА окрашивание (зеленый), (С) RHO окрашивание (красный). (D), объединенного изображения. Потому что окрашивание СА выражается раньше, чем окрашивание RHO только две ячейки двойной помечены СА и Ро (см *). Масштабная линейка:. 23 мкм Пожалуйста, нажмите здесь, чтобы увидеть увеличенную версию этой цифры.
{kind=link}

Рисунок 3. Сравнение экспрессии трех генов в мозгу и во всей сетчатки и фоторецепторов у животных дикого типа в возрасте тридцати пяти дней. (А) Родопсин (Ро), S-arrestin (SAG) и конус-трансдуцин (Gnat2). (Б) кальбиндин (Calb1) и рековерина (Rcvrn). Данные показаны в виде относительной экспрессии. Относительная выражение значение выраженияполучены из данных микрочипов после normaliszation с помощью программного обеспечения Прочная Multi-массива Средняя (RMA), имеющихся в базе знаний KBaSS (http://kbass.institut-vision.org/KBaSS/).

Рисунок 4. Выражение ГУР и GNAT2 в космическом сетчатки с дикого типа сетчатке послеродовой день 35. Отсутствие их выражения во внутреннем сетчатки и в сетчатке RD1 мыши на послеродовой день 35. ACTB, бета актина. Пожалуйста, нажмите здесь, чтобы увидеть увеличенную версию этой цифры.
{kind=link}
Обсуждение
Сетчатка модель органом в биологии. Исследование сетчатки привело к 6 главных открытий в биологии. Именно в происхождении гена супрессора RB1 первый опухоли. Это показывает молекулярную связь между рецептором тирозин киназ и МАР-киназ через взаимодействие с Сыном sevenless. Это был вовлечен в открытии PAX6, первого гена мастер управления для органов морфогенеза. Он находится в центре генетической связи фактора комплемента H (CFH) с возрастной макулярной дегенерации (ВМД), первый ген восприимчивости болезни определены экране генома широкий ассоциации (GWAS). Наконец, это привело к первой успешной генной терапии для Leber врожденной амаврозом, первый судебный процесс по устранению генная терапия у человека любого наследственные заболевания. Структура этого органа, сохраняется в большинстве видов позвоночных. Его доступность для манипуляций в естественных условиях призвала на ранней стадии функциональной геномики исследований по этой составной части процентовRAL нервную систему.
Здесь мы показываем, как отделить фоторецепторов слой из внутреннего слоя сетчатки vibratome срезов. Этот шаг имеет решающее значение для получения чистых фоторецепторов культур. Наш протокол рассечение делает загрязнение клетках ПЭС очень маловероятно.
Одним из основных проблем является уплощение сетчатки, что необходимо в разделе должным образом внутренней и внешней сетчатки. Срезы сетчатки лучше на сетчатке глаза с малым диаметром, как те, от грызунов, и это диаметр ограничение метода с действующим имеющегося материала.
Желательно перед началом биологически значимые эксперимент, на практике на основе выборки из сетчатки. Проиллюстрируем метод представительными результатами, полученными из культивируемых клетках фоторецепторов, используя материал для выполнения экспрессии изучение обеих мРНК и белков. Исследования экспрессии также может быть выполнена в качестве альтернативы на Sectioнс при лазерном захвата микродиссекции, но культуры лучше всего выполнять с помощью vibratome секционирования. Мы могли бы использовать технику микродиссекции лазера с совершенно иной стратегии. Но, чтобы собрать фоторецепторов слой с микродиссекции устройства для культуры, было бы необходимо, чтобы избежать фиксатор, и это усложнило бы нынешнюю методологию значительно.
Мы разрабатываем протокол, направленный на изучение кинетики и экспрессии генов белка в vibratome разделах течение дегенерации фоторецепторов в моделях пигментный ретинит. Мы считаем, что подробное описание протокола будет полезна для исследователей в области сетчатки биологии, и особенно для протеомическим и метаболомики исследований.
Раскрытие информации
The authors have nothing to disclose.
Благодарности
Igal Gery and David Hicks for anti-SAG antibodies. Ram Fridlich for reading the manuscript.
Материалы
| Name | Company | Catalog Number | Comments |
| Cytidine 5′-diphosphocholin* | Sigma-Aldrich | C0256 | 4.7 µM Cytidine 5′-diphosphocholin |
| Cytidine 5′-diphosphoethanolamine* | Sigma-Aldrich | C0456 | 2.7 µM Cytidine 5′-diphosphoethanolamine |
| Linoleïc acid/ bovine serum albumin (BSA)* | Sigma-Aldrich | L8384 | 100 µg/ml linoleïc acid/ bovine serum albumin (BSA) |
| Triiodo-L-thyronine* | Sigma-Aldrich | T6397 | 0.03 µM Triiodo-L-thyronine |
| 96-well plates | Greiner bio-one | 655-095 | |
| Binocular microscope | Leica | MZ-75 | |
| CO2 independent (CO2-i) | Life Technologies | 18045054 | |
| DMEM | Life Technologies | 41966029 | |
| Forceps n°5 Dumont | Bionic | 11254-20 | |
| Gelatin from porcin skin type A | Sigma-Aldrich | G2500 | |
| GeneChip | Affymetrix | U74v2 | |
| Gentamicin solution | Life Technologies | 15710049 | |
| Hydrocortison* | Sigma-Aldrich | H0888 | 0.55 µM hydrocortison |
| Insulin* (I) | Sigma-Aldrich | I1884 (ITS) | 0.86 µM insulin (I) |
| Papain | Worthington-biochem | WOLS03124 | |
| Poly-D-lysine | Sigma-Aldrich | P-6407 | |
| Progesterone* | Sigma-Aldrich | P7556 | 2.0 µM progesterone |
| Prostaglandin* | Sigma-Aldrich | P5172 | 0.28 µM prostaglandin |
| Putrescine* | Sigma-Aldrich | P5780 | 182 µM putrescine |
| Scalpel Albion | EMS | 72000 | |
| Scissor | Moria | 15396-00 | |
| Sodium pyruvate* | Sigma-Aldrich | S8636 | 1 mM sodium pyruvate |
| Sodium selenite* (S) | Sigma-Aldrich | I1884 (ITS) | 0.29 µM Na2SeO3 (S) |
| Taurine* | Sigma-Aldrich | T8691 | 3 mM taurine |
| Transferrin* (T) | Sigma-Aldrich | I1884 (ITS) | 0.07 µM transferrin (T) |
| Vibratome apparatus | Leica | VT1000-S | |
| * Supplements | |||
Ссылки
- The Organization of the Retina and Visual System. Webvision. Kolb, H., Nelson, R., Fernandez, E., Jones, B. , (2014).
- Perez De Sevilla Muller, L., Shelley, J., Weiler, R. Displaced amacrine cells of the mouse retina. J Comp Neurol. 505 (2), 177-189 (2007).
- Mohand-Said, S., et al. Photoreceptor transplants increase host cone survival in the retinal degeneration (rd) mouse. Ophthalmic Res. 29 (5), 290-297 (1997).
- Daiger, S., Sullivan, L., Bowne, S. RetNet, the Retinal Information Network. , The University of Texas Health Science Center. Houston, TX. Available from: https://sph.uth.edu/retnet (2014).
- Mohand-Said, S., Hicks, D., Dreyfus, H., Sahel, J. A. Selective transplantation of rods delays cone loss in a retinitis pigmentosa model. Arch Ophthalmol. 118 (6), 807-811 (2000).
- MacLaren, R. E., et al. Retinal repair by transplantation of photoreceptor precursors. Nature. 444 (7116), 203-207 (2006).
- Yang, Y., et al. Transplantation of photoreceptor and total neural retina preserves cone function in P23H rhodopsin transgenic rat. PLoS One. 5 (10), (2010).
- Mohand-Said, S., et al. Normal retina releases a diffusible factor stimulating cone survival in the retinal degeneration mouse. Proc Natl Acad Sci USA. 95 (14), 8357-8362 (1998).
- Fintz, A. C., et al. Partial characterization of retina-derived cone neuroprotection in two culture models of photoreceptor degeneration. Invest Ophthalmol Vis Sci. 44 (2), 818-825 (2003).
- Cronin, T., Leveillard, T., Sahel, J. A. Retinal degenerations: from cell signaling to cell therapy; pre-clinical and clinical issues. Curr Gene Ther. 7 (2), 121-129 (2007).
- Wright, A. F. A searchlight through the fog. Nat Genet. 17 (2), 132-134 (1997).
- Leveillard, T., et al. Identification and characterization of rod-derived cone viability factor. Nat Genet. 36 (7), 755-759 (2004).
- Lillig, C. H., Holmgren, A. Thioredoxin and related molecules--from biology to health and disease. Antioxid Redox Signal. 9 (1), 25-47 (2007).
- Fridlich, R., et al. The thioredoxin-like protein rod-derived cone viability factor (RdCVFL) interacts with TAU and inhibits its phosphorylation in the retina. Mol Cell Proteomics. 8 (6), 1206-1218 (2009).
- Yang, Y., et al. Functional cone rescue by RdCVF protein in a dominant model of retinitis pigmentosa. Mol Ther. 17 (5), 787-795 (2009).
- Bennett, J. Strategies for delivery of rod-derived cone viability factor. Retina. 25 (8 Suppl), S47(2005).
- Komaromy, A. M., et al. Transient photoreceptor deconstruction by CNTF enhances rAAV-mediated cone functional rescue in late stage CNGB3-achromatopsia. Mol Ther. 21 (6), 1131-1141 (2013).
- Birch, D. G., Weleber, R. G., Duncan, J. L., Jaffe, G. J., Tao, W. Randomized trial of ciliary neurotrophic factor delivered by encapsulated cell intraocular implants for retinitis pigmentosa. Am J Ophthalmol. 156 (2), 283-292 (2013).
- Cronin, T., et al. The disruption of the rod-derived cone viability gene leads to photoreceptor dysfunction and susceptibility to oxidative stress. Cell Death Differ. 17 (7), 1199-1210 (2010).
- Leveillard, T., Sahel, J. A. Rod-derived cone viability factor for treating blinding diseases: from clinic to redox signaling. Sci Transl Med. 2 (26), 26ps16(2010).
- Fontaine, V., Hicks, D., Dreyfus, H. Changes in ganglioside composition of photoreceptors during postnatal maturation of the rat retina. Glycobiology. 8 (2), 183-190 (1998).
- Fontaine, V., Kinkl, N., Sahel, J., Dreyfus, H., Hicks, D. Survival of purified rat photoreceptors in vitro is stimulated directly by fibroblast growth factor-2. J Neurosci. 18 (23), 9662-9672 (1998).
- Reichman, S., et al. The homeobox gene CHX10/VSX2 regulates RdCVF promoter activity in the inner retina. Hum Mol Genet. 19 (2), 250-261 (2010).
- Sandu, C., Hicks, D., Felder-Schmittbuhl, M. P. Rat photoreceptor circadian oscillator strongly relies on lighting conditions. Eur J Neurosci. 34 (3), 507-516 (2011).
- Dorrell, M. I., Aguilar, E., Weber, C., Friedlander, M. Global gene expression analysis of the developing postnatal mouse retina. Invest Ophthalmol Vis Sci. 45 (3), 1009-1019 (2004).
- Brooks, M. J., Rajasimha, H. K., Roger, J. E., Swaroop, A. Next-generation sequencing facilitates quantitative analysis of wild-type and Nrl(-/-) retinal transcriptomes. Mol Vis. 17, 3034-3054 (2011).
- Delyfer, M. N., et al. Transcriptomic analysis of human retinal surgical specimens using jouRNAI. J Vis Exp. (78), (2013).
- Ying, S., et al. A CAT reporter construct containing 277bp GNAT2 promoter and 214bp IRBP enhancer is specifically expressed by cone photoreceptor cells in transgenic mice. Curr Eye Res. 17 (8), 777-782 (1998).
Перепечатки и разрешения
Запросить разрешение на использование текста или рисунков этого JoVE статьи
Запросить разрешениеThis article has been published
Video Coming Soon
Авторские права © 2025 MyJoVE Corporation. Все права защищены