
Method Article
Объединяя анализ ДНК в добыче сырой вириона с анализом РНК от зараженных листьев, чтобы обнаружить новый вирус геномов
В этой статье
Резюме
Здесь мы представляем новый подход для выявления вирусов растений с двойной нити ДНК геномов. Мы используем стандартные методы для извлечения ДНК и РНК из зараженных листьев и осуществлять секвенирование нового поколения. Инструменты bioinformatic собрать последовательности в Сахалинской, определить Сахалинской, представляющих геном вируса и назначить геномов таксономических групп.
Аннотация
Этот подход metagenome используется для идентификации вирусов растений с круговой геномов ДНК и их стенограммы. Часто завод ДНК вирусов, которые происходят в низкие титры в принимающих их или не может быть механически привиты на другой хост трудно распространить достижения большей титра инфекционного материала. Зараженные листья шлифуются в мягкий буфера с оптимальным pH и ионный состав рекомендуется для очистки большинства бацилловидные пункт ретровирусы. Карбамид используется сломать вверх включения органов, которые ловушку вирионов и распустить клеточных компонентов. Дифференциального центрифугирования обеспечивает дальнейшее разделение вирионы от загрязнений завод. Затем лечение протеиназы K удаляет capsids. Затем вирусной ДНК сконцентрировано и используется для виртуализации нового поколения (НГС). NGS данные используются для сборки Сахалинской, которые принимаются в NCBI-BLASTn определить подмножество вирус последовательностей в созданном наборе данных. В параллельных конвейера РНК изолирован от зараженных листьев с использованием стандартного метода извлечения РНК на основе столбцов. Затем рибосома истощения осуществляется для обогащения для подмножества мРНК и вирус стенограмм. Собрал последовательности, производные от РНК последовательности (РНК seq) были представлены NCBI-BLASTn определить подмножество вирус последовательностей в этом наборе данных. В нашем исследовании мы определили два смежных полнометражные badnavirus геномов в двух наборах данных. Этот метод является предпочтительным для другой распространенный подход, который извлекает совокупного населения малых РНК последовательности для воссоздания растений вирус genomic последовательностей. Этот вирус восстанавливает последний метагеномных конвейера связанных последовательностей, ретро переписывание элементы вставляются в геном растений. Это сочетается в биохимических и молекулярных анализов для дальнейшего различить активно инфекционных агентов. Подход, задокументированы в данном исследовании, восстанавливает последовательности представитель репликацию вирусов, которые вероятно указывают активной вирусной инфекции.
Введение
Возникающие болезни растений диск исследователям развивать новые инструменты для определения правильного причинной агент(ы). Первоначальные сообщения о новых или повторяющиеся вирусных заболеваний основаны на часто встречающиеся симптомы, такие как мозаика и пороки развития листьев, очистка вен, карликовость, увядание, поражения, некроз, или другие симптомы. Стандарт для представления новый вирус возбудителя болезни заключается в том, чтобы отделить его от других загрязняющих патогенов, распространить его в подходящий хост и воспроизвести заболевание, прививки в здоровых растений видов исходного узла. Ограничение этого подхода является, что многие родов вирусов растений зависит от насекомых или других векторов для передачи подходящей принимающей или вернуться к оригинальной видов хост. В этом случае поиск соответствующий вектор может быть продлен, могут возникнуть трудности для создания колонии лаборатории вектора, и необходимы дальнейшие усилия для разработки протокола для экспериментальной передачи. Если условия для успешной передачи лабораторных не может быть достигнута, то работа отстает от стандарт для представления новых вирусных заболеваний. Для вирусов, которые происходят в их естественной хостов на очень низкие титры исследователи должны определить альтернативные хосты для распространения поддерживать достаточные запасы инфекционных для проведения научных исследований. Для видов вирус заражает только несколько растений это также может быть препятствием для выращивания запасов культур1.
В последние годы ученые все чаще применяют NGS высокой пропускной способности и метагеномных подходы для выявления вируса последовательностей, которые присутствуют в окружающей среде, которые могут существовать отношения к известной болезни, но может быть назначен таксономических видов и родов 2 , 3 , 4. такие подходы к открытию и категоризации генетических материалов в различных условиях предоставляют способ описать разнообразие вируса в природе или их присутствие в определенные экосистемы, но не обязательно подтвердить основы для определения причинных агентов для явно заболеваний.
Род Badnavirus принадлежит к семейству Caulimoviridae pararetroviruses. Эти вирусы являются бацилловидные в форме с круговой двойной нити ДНК геномов около 7 до 9 КБ. Все pararetroviruses реплицировать через промежуточное РНК. Pararetroviruses существуют как episomes и реплицировать независимо от завода хромосомной ДНК5,6. Полевые исследования вируса населения указывают, что эти популяции вируса генетически сложны. Кроме того информация, полученная по широкому геномах растений путем sequencing высокой пропускной способности обнаружили многочисленные примеры фрагментов генома badnavirus вставлены события незаконной интеграции в геномов растений. Эти эндогенные badnavirus последовательности не обязательно связаны с инфекцией7,8,9,10,11. Впоследствии использование NGS для идентификации новых badnaviruses в качестве возбудителя заболевания осложняется субпопуляция разнообразия episomal геномов, а также появление эндогенных последовательности12,13.
Хотя есть не один оптимальный конвейер для открытия Роман pararetrovirus геномов, есть два общих подхода к идентифицировать эти вирусы как причинных агентов для болезни. Одним из методов является обогатить для малых РНК последовательности из зараженных листьев и затем собрать эти последовательности для воссоздания вирус genome(s)14,,1516,17. Другой подход — подвижного круга амплификации (RCA) для того чтобы усилить круговой ДНК вируса геномов18. Успех RCA зависит от возраста листа и титр вируса в выбранной ткани. RCA продукты подвергаются пищеварение ограничения и клонирования в плазмиды для прямого секвенирования19,,2021.
Канна желтый крапчатости вирус (CaYMV) является badnavirus и описывается как этиологической причиной заболевания желтой пятнистости в Канны, хотя только 565 bp фрагмент генома был ранее изолированные от зараженных Каннас22. Современные исследования выявлены CaYMV в purpurata Альпиния (цветение имбиря; CaYMV-Ap)23. Целью данного исследования было восстановить полный badnavirus последовательностей генома от зараженных Канна лилии. Мы описать протокол для очистки вируса от завода загрязнений и затем изолировать вирусной ДНК от этого препарата и подготовить библиотеки ДНК для использования в NGS. Этот подход устраняет необходимость в промежуточных молекулярной амплификации шаги. Мы также изолировать mRNA от инфицированных растений для РНК след NGS, которая включает в себя РНК seq осуществлялась с помощью подготовки каждого нуклеиновых кислот. Собрал Сахалинской были найдены относиться к Badnavirus таксонов в обоих наборах данных, с помощью национального центра для биотехнологии и информации (NCBI) основные местные выравнивание инструмент для поиска нуклеиновые кислоты (BLASTn). Мы определили геном badnavirus двух видов24.
протокол
1. общий вирус очистки от дифференциального центрифугирования, с помощью стандартного метода, Кови et al. 25
- Во-первых, вырезать 80-100 г листьев от больного растения и притирают Варинга блендер на 4 ° C, с помощью 200 мл, шлифовка буфера (0,5 М NaH2PO4, 0,5 М Na2HPO4 (рН 7,2). и 0,5% (w/v) Na2SO3). Носите лабораторный халат и перчатки для всех шагов данной процедуры.
- Затем перенесите огневки (300 мл) на стакан емкостью 1,0 Л. Добавьте 18 g мочевины и 25 мл 10% неионогенных моющего средства (t-Oct-C6H4-9(OCH2CH2) OH) огневки внутри химические вытяжки.
Примечание: Этот шаг лучше носить защитные очки и простые дыхательные маски для личной защиты. - Перемешать с магнитной мешалкой кратко в капюшоне и охватывают стакан с фольгой. Затем передать покрыты фольгой стакан холодной комнате и перемешать с магнитной мешалкой на ночь при 4 ° C.
- Передача огневки центрифуга ротор бутылки (250 мл-контейнеры) и центрифуги в фиксированный угол ротора на 4000 x g 10 мин при 4 ° C. Химический вытяжной шкаф восстановить супернатант и процеживают через 4 слоя Марли.
- Разделите огневки среди 38,5 мл полипропиленовые пробирок и центрифуги для 2,5 ч на 40000 x g при 4 ° C. Как правило для проверки наличия зеленых гранул в нижней части трубки и белые гранулы по длине трубки. Слить супернатант и сохранить оба гранул; Поместите образцы на льду.
Примечание: Зеленые гранулы содержит хлоропластов, крахмал и другие органеллы. - Работая в химические вытяжки, используйте для разделения гранулы резиновыми полицейский. Ресуспензируйте белые гранулы в каждой бутылке ротора в 1 мл ddH2O в течение 1-2 ч при сохранении суспензий на ночь при 4 ° C, чтобы позволить материалы полностью раствориться в раствор. Центрифуга для подвески на 6000 x g и 4 ° C на 10 мин для удаления оставшихся мусора.
- Центрифуга для концентрированных подвеска на 136 000 x g за 2 ч при 4 ° C до Пелле вирионов. Ресуспензируйте гранулы в 1 мл буфера (50 мм трис-HCl, pH 7.5, 5 мм MgCl2).
Примечание: Это необязательный шаг является для лечения вирионы с DNAse I (10 мкг/мл) для 10 минут при 37 ° C, чтобы удалить не encapsidated ДНК, т.е., загрязняя хлоропластов и митохондриальной ДНК. Затем, деактивируют DNAse I путем добавления ЭДТА в 1 мм. - Нарушить вирионы с 40 мкл 2 мкг/мкл протеиназы K при 37 ° C 15 мин.
- Работа внутри химические вытяжки восстановить вириона ДНК методом экстракции органических. Одежда защитная маска, перчатки и пальто лаборатории во время извлечения для защиты от потенциальных последствий острого здоровья. Добавить 1 объем фенол хлороформ изоамилового спирта (49:50:1) в образце и встряхнуть рукой для 20 s. центрифуги при комнатной температуре за 5 мин на 16000 x g. удалить верхний водной фазе и передачи новой трубки. Повторите это добыча два или более раз. Распоряжаться органические фазы, поместив его в стеклянной бутылке отходов для утилизации химических институциональных26.
- Концентрат ДНК, используя высыпание этанола. Используйте 0,3 М конечная концентрация натрия ацетата (рН 5.2) и 2,5 объемами этанола 95%. Поместите образцы при-20 ° C за 30-60 мин и центрифуги на 13 000 x g 10-20 мин в Пелле ДНК26.
- Работая на лабораторном столе, Ресуспензируйте Пелле ДНК в 1 мл 0,1 мм TE буфера (рН 8,0). Заполнителя подвеска через колонку фильтрации коммерческих гель (обычно используется для полимеразной цепной реакции (ПЦР) очистки) отказаться от соли и низкий молекулярный вес материала, которые могли бы помешать NGS.
- Анализ образцов электрофорезом геля агарозы 1%, с использованием бромид ethidium, пятнать для просмотра качества подготовки. Оцените качество ДНК с помощью nanodrop спектрофотометром.
Примечание: Коэффициент поглощения образцов на 260 λ и 280 λ между 1.85 и 2.0 обычно указывает, что подготовка является «чистым» от примесей и желаемого качества. - Анализ качества ДНК (используйте 5 ПГ-10 нг) с помощью чипа на основе капиллярного электрофореза инструмент.
Примечание: Качество вывода показывает чистый вершины, представляющие фрагментов ДНК, распределенных по размеру вдоль оси x. Высота пика указывает обилие фрагмента. Зубчатые пики указывают частично деградировавших фрагменты или химических загрязнений. Круглая кривые представляют мазок ДНК, указывающее низкого качества
2. Библиотека подготовка использование ДНК и эмульсионной основе клонального амплификации (emPCR усилители)
Примечание: Библиотека обычно подготовлен Фондом NGS, который осуществляет клиенто ориентированной работы.
- Сдвига решения ДНК (> 200 ng) с помощью распылителя, который преобразует ДНК фрагментов. Перевязать коммерческих адаптеры согласно инструкциям руководства27.
- Проводить emPCR амплификации ДНК образца согласно производителя инструкции28,29,30. Мыть Повторите три раза и после каждого мытья, Пелле бусины в minicentrifuge для 10 s. отменить надосадке после каждой стирки.
Примечание: Процедура начинается с подготовки захвата бусины промывкой в буфере коммерческих мыть, комплект. emPCR обычно используется для усиления шаблон для NGS. - Тепла денатурировать ДНК или РНК при 95 ° C за 2 мин, а затем 4 ° C до готовой к использованию. Использование 200 - миллионов молекул ДНК/РНК до 5 миллионов захвата бусины в окончательный объем 30 мкл. подготовить пробный образец наряду с образца ДНК/РНК и выполняют следующие шаги с образцов нуклеиновых кислот, а также макет образца.
- Выполнять эмульгирование, vortexing трубки эмульсии нефти для 10 s на максимальной скорости, затем залить все содержимое (4 мл) в пластиковой помешивая трубки, который совместим с платформой гомогенизатор. Место перемешивания трубки на платформе для смешивания эмульсии при 2000 об/мин за 5 мин.
- Отказаться от 100 мкл аликвоты эмульсии в 8-газа крышка трубки или 96-луночных тарелку. Крышка трубки или печать пластину и осуществляют emPCR, используя производителя рекомендованные программы28.
Примечание: После завершения ПЦР проверьте скважин, чтобы увидеть, если эмульсия нетронутыми и затем перейти. Отменить все хорошо, если эмульсия нарушается. - Носите пальто лаборатории и работы в химической капот для сбора амплифицированного ДНК бусины (АБР). Вакуум аспирацию эмульсии из скважин и собирать шарики в 50 мл трубки. Промыть лунки дважды с 100 мкл изопропанола и аспирационная промыть, чтобы же Тюбик 50 мл.
- Вихревой собранные эмульсии и Ресуспензируйте АБР изопропанолом окончательный объем 35 мл. Пелле АБР в 930 x g для 5 минут удалить супернатант и добавляют 10 мл расширения буфера. Вихревой АБР и затем вымыть, добавляя изопропиловый спирт 40мл окончательный объем центрифуги и удалить супернатант после каждого мытья и повторите шаг мыть дважды.
- Провести окончательный мыть с помощью этанола вместо изопропиловый спирт. Добавьте расширение буфера окончательный объем 35 мл, вихрь и Пелле бусы на 930 x g для 5 минут удалить супернатант но оставить 2 мл расширения буфера.
- Передать пробки microcentrifuge подвеска и кратко центрифуги для пеллет АБР. После отмены супернатанта, промойте АБР Пелле дважды с 1 мл раствора расширения буфера. Центрифуга и отбросить надосадке после каждой стирки.
- Подготовить для обогащения шарик библиотеки ДНК, добавьте 1 mL 1 N NaOH в бисер. Вихревой АБР и затем проинкубируйте 2 мин при комнатной температуре. Центрифуга и удалить супернатант. Повторите этот шаг мыть раз.
- Добавьте 1 mL отжига буфера, затем вихревой АБР и инкубировать в течение 2 минут при комнатной температуре. Кратко центрифуги и удалить супернатант. Повторите этот шаг, снова используя 100 мкл отжига буфера.
- Для отжига грунтовки секвенирования ДНК, добавьте 15 мкл Seq Primer A и 15 мкл Seq Primer B комплекта. Кратко смешать, vortexing и место пробки microcentrifuge в блоке тепла в 65 ° C для 5 минут передать льда на 2 мин.
- Вымойте три раза с 1,0 мл отжига буфера. Вихрь 5 s и удалить супернатант каждый раз.
- До последовательности, Измерьте количество бисера с использованием коммерческих шарик счетчика. Там должно быть по крайней мере 500 000 обогащенного бусины.
Примечание: Шарик счетчик является специальное устройство, которое меры бусины в предоставленный microcentrifuge трубку.
3. Общие мРНК изоляции и dsDNA синтеза начиная с инфицированных Canna листья что тест-ПЦР для CaYMV с помощью сообщили диагностических праймеры
- Носите лабораторный халат и перчатки для личной защиты во всех последующих шагах. Работая на лабораторном столе, собирать 12 образцов из листьев и ввергнуть образцы в жидкий азот. Использование мельницы для гомогенизации. Использование коммерческих комплект, который предоставляет стандартный метод на основе столбцов для всего завода РНК изоляции. Добавьте буфер lysis гуанидина Изотиоцианаты, представленной в комплект к земле образца и трясти за 20 s.
- Добавление этанола и перемешать, согласно инструкции по установке. Добавьте каждый огневки спин столбец, который связывает РНК мембраны. Вымойте три раза и элюировать РНК в восстановления трубки24.
- Количественного определения РНК, используя спектрофотометр для измерения коэффициента поглощения на 260 λ и 280 λ. проверять РНК целостность с помощью электрофореза геля агарозы 1% окрашенных бромидом ethidium.
Примечание: Поглощение соотношение между 1.85 и 2.0 указывает, что подготовка желаемого качества. Лечить РНК с DNase I (10 мкг/мл) для 10 минут при 37 ° C. Используйте столбец коммерческих спин сконцентрировать РНК в РНКазы свободной воды31. Бассейн РНК образцы перед продолжением. - Использование набора для удаления рРНК для удаления завод рибосомной РНК. Аликвота магнитные бусы пробки microcentrifuge и мыть дважды с РНКазы свободной воды. Вихревой трубе аликвота Ресуспензируйте, поместите трубку на магнитную подставку и ждать жидкость для очистки. Отменить супернатант и заменить решение ресуспендирования магнитный шарик. Вихрь Ресуспензируйте и 1 мкл АБС битор РНКазы.
Примечание: Такие комплекты использовать oligo-dT привязан к магнитной бусины, которые гибридизировать к мРНК. Данный метод использует стандартные магнит шарик сепарационные технологии для восстановления стенограммы24. - Комбинат 500 нг 1,25 мкг РНК, RNase бесплатная вода и реакция буферы, предоставляемые в комплект. Поместите смесь для 10 мин при 50 ° C. Удалить из тепла и Добавить промытый магнитные шарики в РНКазы свободной воды. Вихревой кратко и набор при комнатной температуре в течение 5 мин.
- Место на магнитную подставку и ждать жидкость для очистки. Супернатант передать свежие microcentrifuge трубки. Установите на льду.
- Используйте метод на основе решения захвата для обогащения exosomes и 200 нг РНК подготовить кДНК библиотеки.
Примечание: Двойной нити кДНК библиотеки обычно подготовлен Фондом NGS, который осуществляет клиенто ориентированной работы. - Фрагмент РНК с помощью коммерческого решения фрагментации РНК (0,136 г ZnCl2 и 100 мм трис-HCl рН 7,0). 2 мкл раствора до 18 мкл РНК (200 ng всего). Спин трубы кратко в microcentrifuge, поместите образцы при 70 ° C 30 s, и передачи в лед. Остановите реакции с использованием 2 мкл 0,5 М ЭДТА pH 8.0 и 28 мкл 10 мм трис-HCl рН 7,5.
- Связать магнитные бусы РНК путем смешивания при комнатной температуре на 10 минут использования магнитных концентратор собирать шарики и удалить супернатант. Вымойте бусины три раза с 200 мкл на 70% спирте. Отменить каждый мыть и затем воздух сухой гранулированных бусы при комнатной температуре 3 мин Ресуспензируйте в 19 мкл 10 мм трис-HCl рН 7,5.
- Отжиг случайных праймеров для раздробленной РНК путем нагревания до 70 ° C для 10 мин и затем поместите трубку на льду Стрэнд 2 мин подготовить первый и второй cDNA стренги, используя набор стандартных коммерческих cDNA синтеза.
- Очищайте двухручьевой cDNA, используя концентратор магнитный шарик. Вымойте с 800 мкл 70% этанола в три раза. Отменить каждый мыть и подсушите воздухом гранулы при комнатной температуре 3 мин Ресуспензируйте в 16 мкл 10 мм трис-HCl рН 7,5. Используйте концентратор магнитный шарик для разделения бисер от двунитевая cDNA, который в настоящее время в растворе. Удалите cDNA, дозирование в новой ПЦР-пробирку 200 мкл.
- Осуществляют фрагмент конец ремонт с использованием полимеразы Taq и смесь deoxyribonucleotides, предоставляемый коммерческая библиотека подготовка комплекта. Коммерческих комплект предоставляет предварительно разбавленного адаптеры для добавления к каждому концу двухручьевой cDNA, используя коммерческие лигаза при 25 ° C для 10 мин.
4. NGS ДНК библиотека готовят из сырой вирус подготовки и dsDNA библиотека подготовила от мРНК
- Используйте инструмент пиросеквенирования стандарта высокой пропускной способности и следуйте все рекомендованные производители протоколы для создания прямых показаний последовательностей ДНК. Использование коммерческих последовательности реагентов, включая дневно обозначенные нуклеотидов.
Примечание: Подробнее инструкциям производителя с инструментом. - Выполняют после последовательности анализа с использованием генома Ассамблеи программное обеспечение, которое автоматически собирает читает производить первый набор Сахалинской со средней длиной < 700 bp. Использование FastQC программного обеспечения на веб-сайте iPlant/CyVerse, который выполняет контроль качества Проверка на сырой последовательности данных32. Выберите последовательности с Phred баллов ≥ 30 продолжить реконструировать более последовательностей из небольших последовательности читает24 с помощью сопоставления и amplicon программного обеспечения.
Примечание: Подробнее см. инструкции производителя. - Представить эти собраны Сахалинской NCBI-BLASTn анализ с использованием МЕГАВЗРЫВ по умолчанию модуль, а также Viridplantae (TaxID: 33090) и вирусов (TaxID: 10239) как имена ограничения Организменное33. Соберите субпопуляция Сахалинской, которые показывают высокое сходство сообщил Badnavirus геномов в отчет.
- Убедитесь, что присоединились леса, которые представляют собой один или более кандидата полнометражного вирус геномов, правильно производить в кадр последовательности, которые имеют ту же организацию стандартного badnavirus геном. Для этого, ввод генома вируса полная длина кандидат в плазмиду рисунок программное обеспечение. Затем подтвердите первые 15 нуклеотидов состоит из tRNAвстретил (TGGTATCAGAGCGAG) которая высоко сохраняется среди badnaviruses. Найти потенциальных сплайсингу сигнал в конце 3' генома. Аннотируйте полный геном определить наличие двух небольших ORFs и один большой ORF, кодировку polyprotein. Затем используйте портал ExPASy перевод инструмент для определения badnavirus ORF3, ORF1 и ORF2 перевод продуктов34.
Примечание: Это научное программное обеспечение является бесплатным и будет генерировать круговой ДНК, определить все открытые чтения рамы и обеспечивает немедленного вывода, чтобы убедиться, что последовательность представляет полную длину круговой ДНК генома. - Используйте несколько инструментов сравнения последовательности, мышц и CLUSTALW, открытым исходным кодом для сравнения геномов вируса, полученные от35,анализ ДНК и РНК36.
- Поиск в базе данных нуклеотидов NCBI получить полный геном последовательности 30 badnavirus видов и экспортировать их в виде документа в формате .fasta. Загрузите программное обеспечение, которое проводит эволюционный генетический анализ последовательностей наряду с последовательности генома вируса, полученные NGS последовательности. Генерировать несколько выравниваний последовательности и максимальная вероятность деревья с помощью мышц37.
5. качество оценки De Novo последовательности по амплификации PCR геномов вирусов из зараженных растений
- Входной последовательности генома вновь выявленных полнометражные badnavirus (формат .fasta) в бесплатные онлайн инструмент Primer3 для получения ПЦР праймеры38. Идентифицировать грунтовка наборы, которые будут производить поэтапно продукции 1000-1500 bp по всей длине genome(s) вирус. Отправьте последовательности сервисный центр, который будет синтезировать и доставить праймеры PCR.
Примечание: Вывода определяет приемлемые грунтовка пар с общей и приемлемой температуры плавления и точная грунтовка местах вдоль введенной последовательности. - Работая на лабораторном столе и носить пальто лаборатории и перчатки, изолируйте 5 мкг ДНК с листьями инфицированных вирусом и здорового управления, используя автоматизированный метод, который включает в себя стандартные парамагнитных целлюлозы частиц для изолировать дна от завода материала39 . Замораживание в жидком азоте в пробки microcentrifuge листовой материал (20-40 мг) и измельчить с помощью бисерная мельница. Объединить образец с литического буфера в пробки microcentrifuge и добавить РНКазы A для каждого образца. Вихревой образца для 10-20 s и кратко спина образца для удаления твердых частиц.
Примечание: Парамагнитных целлюлозы частицы обладают высоким потенциалом ДНК связывающих и изолировать высокие урожаи чистого ДНК. Методы стандартных коммерческих кремнезема столбец для изоляции ДНК не эффективно извлечь ДНК из различных видов растений. Следовательно десятки методы существуют, которые являются изменения этих процедур для повышения эффективности отдельных растений. Метод частиц автоматизированных парамагнитных целлюлозы был выбран потому что он дает больше и более высокого качества ДНК из более чем 25 видов травянистые растения40. - Используйте коммерческие реагент картриджи для автоматизированного парамагнитных изоляции ДНК. Добавьте 300 мкл нуклеиназы свободной воды в каждой коммерческой реагент картриджа и передачи завод lysate же картридж. Поместите картридж в стойку картридж, место поршень в колодец ближе к трубе элюции и место Элюирующий буфер в элюции трубку. Загрузить картриджи в изоляции машина автоматизированных нуклеиновой кислоты и запустить завод ДНК изоляции протокол41,42.
- Осуществляют ПЦР извлечь набор перекрывающихся продуктов PCR. Используйте 5 мкм каждого прямого и обратного праймера с 35 циклов амплификации PCR. Используйте следующие условия Велоспорт: Денатурация при 95 ° C 60 s, отжиг при 50 ° C для 45 s и расширение в 72 ° C для 1-2 мин с расширением окончательный при 72 ° C для 7-10 минут использования фильтрации столбца расфасованных гель для устранения солей и низкий молекулярный вес материала в шаге 1.231.
- Рассчитать молярное соотношение 3:1 продукта ПЦР для векторных определить количество продукта PCR чтобы перевязать 50 нг линеаризованного pGEM плазмиды43. Использование элемента управления Вставьте ДНК для определения, является ли перешнуровок работать эффективно. Выполняют лигирование, на ночь с помощью T4 ДНК лигаза (3 U/мкл) при 4 ° C. Затем преобразование коммерчески подготовленные JM109 сведущее Escherichia coli клетки. Использование управления 100 ПГ режиссерский плазмидной ДНК как позитивный элемент управления для эффективного преобразования. Пластина 100 мкл трансформированных клеток на LB-агар пластин с антибиотиком и синий/белый выбора восстановить перевязаны плазмид26. Инкубировать пластины для 16-24 ч при 37 ° C.
Примечание: PGEM вектор имеет ген lacZ, который кодирует β-галактозидазы. Преобразованные бактерий, выращенных на тарелку содержит 100 мкг/мл ампициллин, 0,5 мм ИПТГ, 80 мкг/мл 5-bromo-4-chloro-3-indoyl-β-D-galactopyranosidase (X-gal) станет синим за активность β-галактозидазы. PGEM плазмиды линеаризованных в манере, которая нарушает ген lacZ . Колонии, которые содержат вставки продукт PCR нарушить ген lacZ и не усваивать X-Гал. Эти колонии являются белыми. Таким образом колоний с insert можно отличить от тех, кто без вставки по цвету колонии (белый и синий)26. - Изолируйте ДНК из трех колоний, с использованием стандарта на основе столбцов плазмида изоляции комплект39. Последовательности трех плазмид продукта преобразования. Сравнение каждой последовательности ДНК с геномов вируса de novo собрал, производимые NGS. Используйте CLUSTALW для выравнивания последовательностей и обеспечить, что они надлежащим образом отсортированы.
Результаты
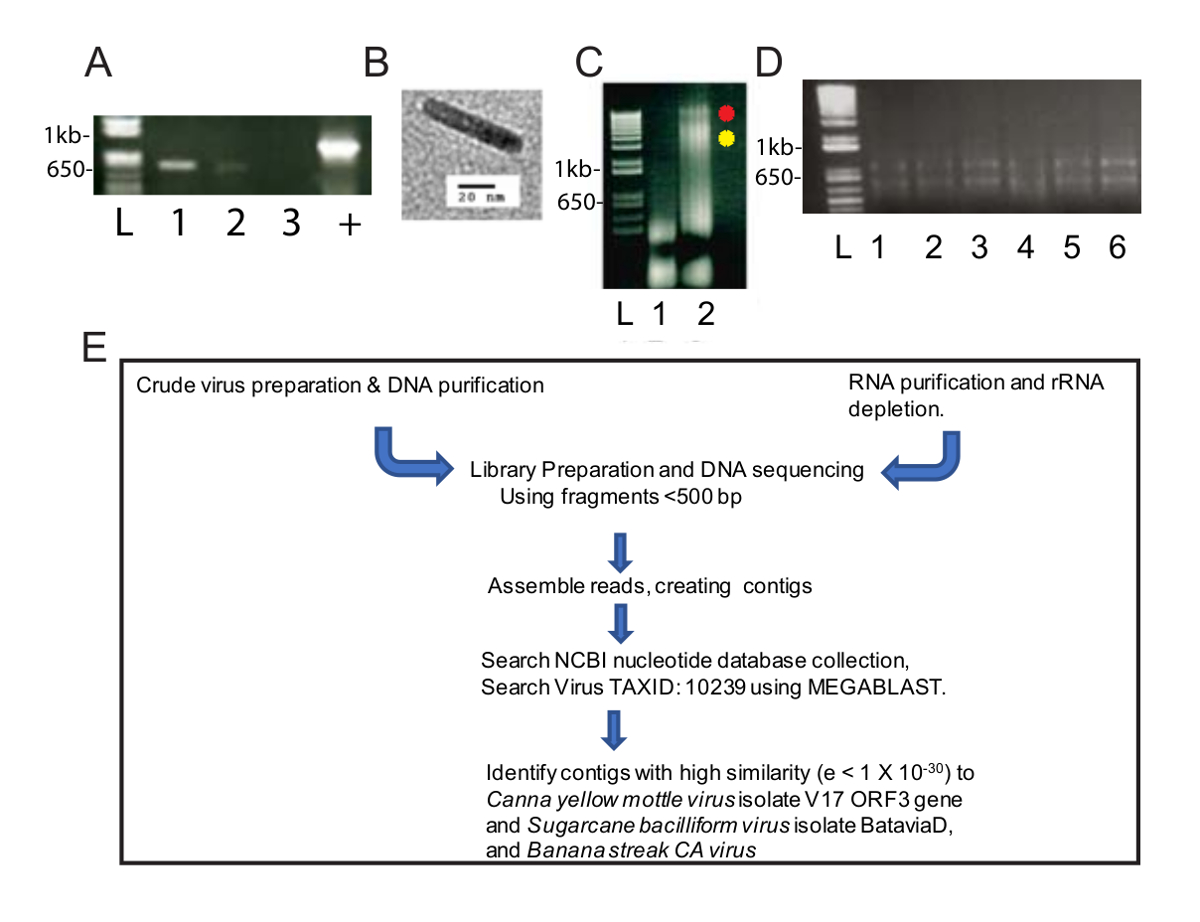
Этот метод очистки измененных вирусов предусмотрено обогащение вируса ННО полезно для выявления двух видов вируса NGS и биоинформатики. После огневки был центрифугируется на 40000 x g для 2,5 ч, там был зеленый Пелле в нижней части трубки и белые гранулы по длине. Зеленые гранулы был высокомобильна в одну трубу microcentrifuge и белые гранулы был высокомобильна в две пробирки microcentrifuge. ПЦР была проведена с использованием стандартных диагностических праймеры CaYMV PCR, и продукты были обнаружены в растворимых гранул белого и не зеленые гранулы (рис. 1А). Образец подготовки нефти был осмотрен просвечивающей электронной микроскопии и мы наблюдали бацилловидные частиц измерения 124-133 Нм в длину (рис. 1Б). Это в пределах предсказал модальной длины большинства badnaviruses. ДНК было извлечено из белых и зеленых гранул и высокомобильна отдельно. Рисунок 1Cмы загрузили 5 мкл ДНК, извлеченные из зеленого и белые гранулы образца (1,6 мкг ДНК для зеленых фракции) и 3,1 мкг ДНК для белых фракция 0,8% агарозном электрофорез геля и анализ ДНК, после бромид ethidium пятнать. Зеленая часть содержащихся низкомолекулярных ДНК в то время как белая часть производства две полосы выше молекулярный вес ДНК, а также нижней молекулярный вес ДНК (рис. 1C). Гель, представлены на рисунке 1C был 40 мин на 100 V и мазок в переулок 3 свидетельствует о том, что гель напряжения должен быть понижен для получения четкого полос. Эти данные позволяют предположить, что белые гранулы был обогащен для вирионов. Концентрация (0,6 мкг/мл) ДНК, извлеченные из белого образца была низкой, но достаточный для NGS, который требует минимум 10 нг ДНК, чтобы продолжить. Фрагментированные ННУ были использованы для подготовки библиотека для NGS.
Параллельно РНК было извлечено от зараженных Канна растений (рис. 1D) для высокой пропускной способности РНК seq. Стандартный рабочий процесс был проведен для библиотеки подготовки, NGS, создавая Сахалинской и выявления вирусного генома последовательности (рис. 1E). Сравнивались результаты от использования ДНК и РНК в качестве исходных материалов.
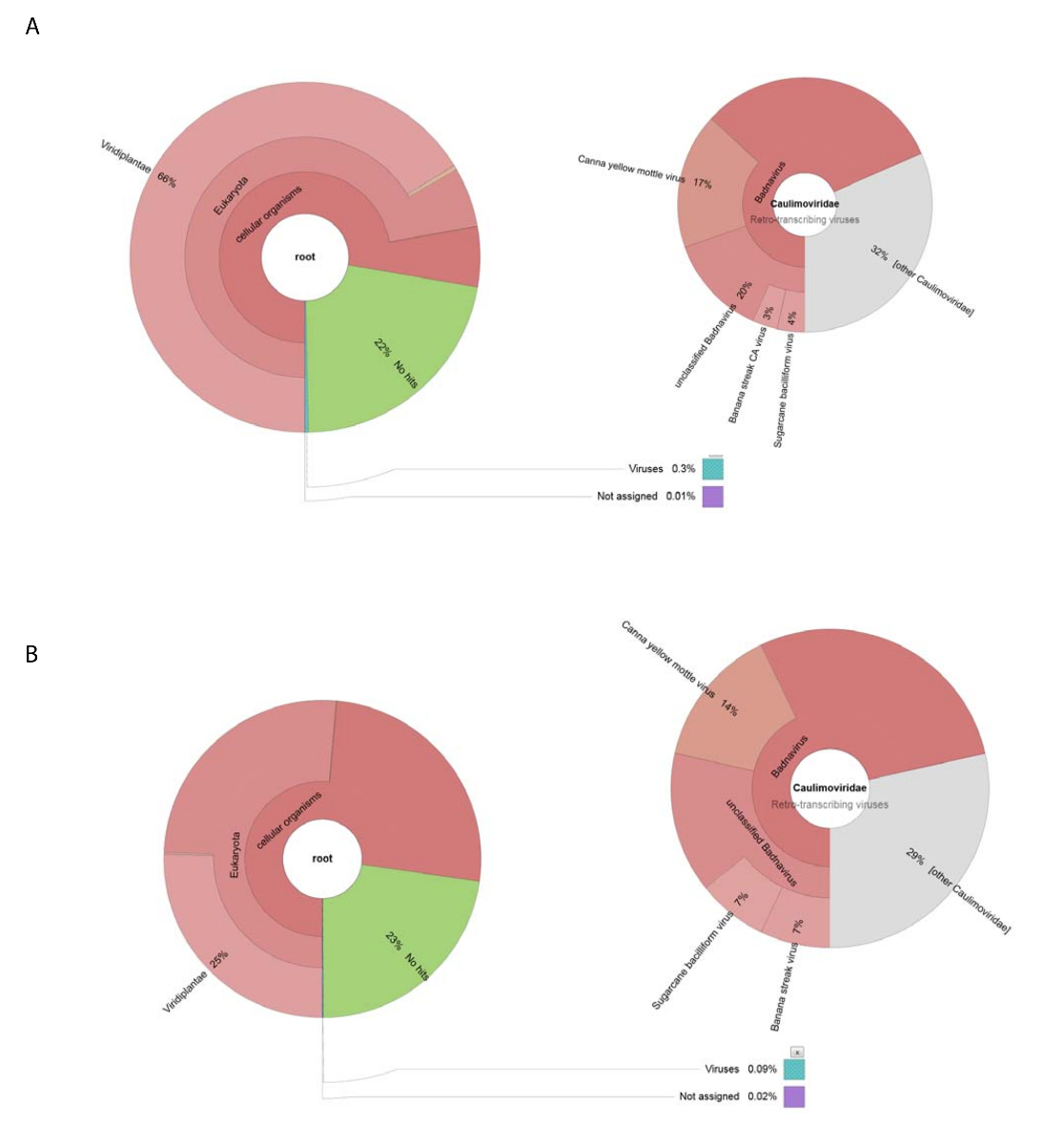
Мы получили 188,626 сырье читает ДНК, NGS с помощью ДНК, изолированные от подготовки сырой вирус. Чтений были собраны в 13,269 Сахалинской и BLASTn был использован для поиска набора данных NCBI нуклеотидных последовательностей (с помощью Viridplantae TaxID: 33090 и вируса TaxID: 10239 как ограничивающие организмов) (рис. 1E). Результаты NCBI-BLASTn показало, что 93% de novo собрал Сахалинской были клеточных последовательностей, 22% были неизвестны, и 0,3% были вирус Сахалинской (рисA). Большинство из Сахалинской классифицированы как клеточных последовательностей были определены как митохондриальных или хлоропласта ДНК. В пределах набора данных вируса Сахалинской, 32% вирус Сахалинской были связаны с членами Caulimoviridae (которые были не Badnavirus последовательности) и 58% из них были связаны с Badnavirus. Вирус Сахалинской 29% были очень похожие (е < 1 x 10-30) к CaYMV, изолировать V17 ORF3 генов (EF189148.1), тростниковый сахар бацилловидные вирус изолировать Батавии D, полный геном (FJ439817.1), и банан полоска CA вирус полный геном ( KJ013511). В рамках этой группы населения были длинные Сахалинской, которая напоминала два полнометражных геномов.
Высок объём РНК seq производится 153,488 уборка отдельных последовательностей читает в среднем читать Длина < 500 bp. Contig Ассамблеи сократить это до 8,243 Сахалинской. Они были представлены NCBI-BLASTn (с помощью Viridplantae TaxID: 33090 и вируса TaxID: 10239 как ограничивающие организмов) и выходы включены 76% Сахалинской в категорию растений клеточных последовательностей, 23% были неизвестны, и 0.1% были классифицированы как вирус Сахалинской ( Рисунок 2 B). более внимательное изучение населения 0,1% населения Сахалинской вирус определено, что 68% из них были назначены в Caulimoviridae (рис. 2B). Три больших Сахалинской внутри этой группы населения были определены с высоким подобия (е < 1 X 10-30) чтобы изолировать V17 ORF3 гена CaYMV (EF189148.1), тростниковый сахар бацилловидные вирус изолировать Batavia D, полный геном (FJ439817.1) и банан полоска CA вирус полный геном (KJ013511). Изучение трех Сахалинской, мы вручную присоединились два из них производят полнометражного вирус генома.
Мы сравнили Сахалинской длины генома вируса, производимые ДНК и РНК последовательности как взаимной эшафот чтобы подтвердить наличие двух полнометражных вирус геномов. Один полнометражный вирус геном 6,966 bp предварительно был назван Canna желтый крапчатости связанных вирус 1 (CaYMAV-1) (рис. 3А). Геном второй был 7,385 bp и вариант CaYMV, заражая Альпиния purpurata (CaYMV-Ap01) (рис. 3А).
Наконец праймеры PCR, которые были разработаны для клон ~ 1000 bp фрагмент каждого вируса, были использованы для дифференциально обнаружить оба геномов в популяции 227 Канна растений, представляющих девять коммерческих сортов. Во многих случаях отдельные растения были инфицированы обоих вирусов. Мы предоставляем пример обнаружения ПЦР-CaYMAV-1 и CaYMV-Ap01 в 12 растений. Три из них были положительными только для CaYMV-Ap01 и девять были позитивными для обоих вирусов (рис. 3B).

Рисунок 1 : Подготовка нуклеиновой кислоты вируса и NGS рабочего процесса. Электрофорез 565 bp ПЦР фрагментов CaYMV геномов геля агарозы (A) (1,0%). Два продукты PCR были обнаружены в пробах, приготовленный из белых гранул (дорожки 1, 2), но не в образце зеленой Пелле (пер. 3). Положительный контроль (+) представляет собой продукт PCR, усиливается от зараженных растений ДНК, который был изолирован с помощью автоматизированных метода с участием стандартных парамагнитных целлюлозы частиц. Лейн L содержит лестница ДНК, используется в качестве стандарта для измерения размера полос линейной ДНК в образце полос. (B) пример частицы вируса, осмотрено просвечивающей электронной микроскопии в белые гранулы оправился от сырой фракционирование зараженных Канна листьев. (C) агарозы (0,8%) гель-электрофореза ДНК, оправился от зеленого (полоса 1) и белый (полоса 2) гранул что проверенные позитивные методом ПЦР в группе A. Красные и желтые точки рядом с пер 2 определить две полосы ДНК высокой молекулярной массой, которые происходят в белом фракции. Электрофорез всего РНК, восстановлены на основе столбцов РНК очищение геля агарозы (D) (1%). Лейн L содержит лестница ДНК, используется в качестве стандарта для измерения размера линейной полос в образце полос. Переулок 1-6 содержит РНК, изолированных от зараженных Канна листья, которые были объединены в один образец Рибо истощение и РНК след (E) схема конвейера изоляции нуклеиновых кислот, Библиотека подготовки, последовательности, contig Ассамблеи и геном вируса Открытие. Пожалуйста, нажмите здесь, чтобы посмотреть большую версию этой фигуры.
{kind=link}

Рисунок 2 : Крона графики визуализации таксономических категорий Сахалинской. (A) диаграммы на слева показывает, изобилия и таксономические распределение Сахалинской, собранный из сырой вирус подготовки. Правый график изображает пропорции вирус Сахалинской, связанные с семьей Caulimoviridae , род Badnavirus и трех тесно связанных между собой видов. (B) на левой панели показывает, что обилие Сахалинской, производный от РНК seq на основе их таксономических распределения. Справа — граф, изображающие обилие Сахалинской населения Сахалинской вирус, связанные с семьей Caulimoviridae , род Badnavirus и трех тесно связанных между собой видов. Пожалуйста, нажмите здесь, чтобы посмотреть большую версию этой фигуры.
{kind=link}

Рисунок 3 . Характеристика CaYMAV-1 и CaYMV-Ap01 геномов. (A) схематическое представление Canna желтый крапчатости связать вирус 1 (CaYMAV) и Канна желтый крапчатости вирус похож на геном, изолированный от Альпиния purpurata (CaYMV-Ap01). Нуклеотидной позиции 1-10 идентифицируется как начало генома и содержит типичные большинства badnavirus геномов сайт антикодонвстретил тРНК. Остановка и запуск позиции для перевода открытой чтении кадра (ORF) 1 и 2 являются смежными. Эти белки имеют неизвестные функции. ORF3 является polyprotein, содержащие цинковый палец (ZnF), протеазы (Pro), обратной транскриптазы (RT) и H РНКазы доменов. 3' poly(A) последовательность сигнала сохраняется для обоих геномов вируса. (B) ПЦР-анализ был проведен с использованием РНК, изолированный от инфицированных вирусом листья и грунтовки, которые обнаружить CaYMAV и CaYMV-Ap01. В том же населения 12 растений три были заражены CaYMV-Ap01 только, тогда как остальные были заражены CaYMAV и CaYMV-Ap01. (+) указывает положительный контроль и (-) указывает на отрицательный контроль. Эта цифра, воспроизводится/изменение от Wijayasekara и др. 24 с разрешения. Пожалуйста, нажмите здесь, чтобы посмотреть большую версию этой фигуры.
{kind=link}
Обсуждение
В последние годы различные методы были использованы для изучения вируса биоразнообразия растений в природных средах, которые включают обогащает вирусоподобных частиц (VLP) или вирусом конкретных ДНК или РНК2,3,44, 45,46 . Эти методы следуют NGS и bioinformatic анализа. Целью данного исследования было найти возбудителя широко распространенным заболеванием в культивируемых растений. Заболевание было сообщено в результате неизвестного вируса, который имеет не охватило бацилловидные частиц, и для которых только фрагмент 565 bp был клонированных47. Эта информация была достаточной для предварительного исследователей назначить гипотетически вирус в род Badnavirus в семье Caulimoviridae. В то время как предыдущие доклады предположил, что Канна крапчатости болезни в Канна лилии был результатом одного badnavirus, используя метагеномики подход, изложенный в этом исследовании, мы определили, что болезнь была вызвана двумя предварительное badnavirus видов24. Таким образом, сила с использованием подхода metagenome обнаружить возбудителя болезни является, что мы теперь можем определить ситуации там, где может быть более чем одной причиной.
Наш подход, сочетая ДНК и РНК последовательности данных является тщательное и также показывает, что результаты, с использованием двух подходов приносят устойчивые результаты и подтвердили наличие двух смежных вирусов. Мы заняты измененная процедура для изоляции caulimoviruses и подготовил образец, который был обогащен для связанных вирус нуклеиновых кислот и которые были защищены в рамках капсид вирус. Служба лаборатории был заключен контракт для секвенирования ДНК. Основные концепции для секвенирования de novo является ДНК-полимеразы включает люминесцентные, помечаются нуклеотидов в ДНК шаблон нити во время последовательных циклов синтеза ДНК. Сахалинской собрал следуют NGS были представлены в производство несколько Сахалинской, которые были определены как вирус Сахалинской bioinformatic рабочий процесс. Дальнейшее подтверждение двух вирусных геномов10,24,48,49,50 был получен путем анализа bioinformatic РНК seq данных, полученных от обедненного Рибо РНК препаратов. Один из интересных результатов состоял в том, чтобы узнать, что населения последовательностей восстановления, ДНК и РНК последовательности представили аналогичные дистрибутивов-вирусных и вирусных нуклеиновых кислот. Для секвенирования ДНК и РНК, < 0,5% последовательностей были происхождения вируса. В рамках населения вируса последовательности 78-82% принадлежал семье Caulimoviridae. Сравнивая Сахалинской собрал вирус от ДНК и РНК последовательности, мы подтвердили, произошедшую два сборных геномов в обоих наборах данных.
Проблемой использования только секвенирования ДНК для идентификации новых вирусных геномов, что геном badnavirus является открытым циркуляр ДНК. Мы предположили, что перекрытия несплошностей в геном последовательности может представлять препятствия для Ассамблеи генома от Сахалинской. Первоначальный анализ результатов секвенирования ДНК выявили два аналогичных геномов вируса. Мы предположили, что эти геномы либо представлены генетическое разнообразие видов, которые не были изучены, или представлены два вида совместно заражение же завод24. Таким образом анализ коллективных bioinformatic наборов данных, полученных NGS ДНК и РНК последовательности, позволил подтверждение наличия двух полнометражных геномов.
Есть еще один доклад, который разработал альтернативный метод для извлечения VLP и нуклеиновые кислоты из растений гомогенатах для метагеномных исследований, на основе процедур для восстановления ДНК вируса мозаики цветной капусты (CaMV; caulimovirus)3. Этот подход, обозначенный Роман РНК и ДНК вируса последовательностей в-культивируемых растений. Действия, производные от процедуры изоляции caulimovirus, используемые в данном исследовании для обнаружения возбудителя болезни культивируемых растений являются в отличие от шагов, полученных для извлечения VLP естественно зараженных растений24. Успех обеих модифицированных методов свидетельствует о том, что рамки процедуры для caulimovirus изоляции может быть ценной отправной точкой для исследования метагеномных вирусов растений в целом.
Раскрытие информации
Авторы не имеют ничего сообщать.
Благодарности
Исследование финансировалось Оклахома центр для развития науки и технологии применения исследований программы этапа II AR 132-053-2; и Оклахома Департамента сельского хозяйства специальность культур исследования Грант программы. Мы благодарим д-р HongJin Хван и ОГУ биоинформатики основного объекта, который был поддержан грантов от NSF (EOS-0132534) и низ (2P20RR016478-04, 1P20RR16478-02 и 5P20RR15564-03).
Материалы
| Name | Company | Catalog Number | Comments |
| NaH2PO4 | Sigma-Aldrich St. Louis MO | S5976 | Grinding buffer for virus purification |
| Na2HPO4 | Sigma-Aldrich | S0751 | Grinding buffer for virus purification |
| Na2SO3 | Thermo-Fisher Waltham, MA | 28790 | Grinding buffer for virus purification |
| urea | Thermo-Fisher | PB169-212 | Homogenate extraction |
| Triton X-100 | Sigma-Aldrich | X-100 | Homogenate extraction |
| Cheesecloth | VWR Radnor, PA | 21910-107 | Filter homogenate |
| Tris | Thermo-Fisher | BP152-5 | Pellet resuspension& DNA resuspension buffers |
| MgCl2 | Spectrum, Gardena, CA | M1035 | Pellet resuspension buffer |
| EDTA | Spectrum | E1045 | Stops enzyme reactions |
| Proteinase K | Thermo-Fisher | 25530 | DNA resuspension buffer |
| phenol:chloroform:isoamylalcohol | Sigma-Aldrich | P2069 | Dissolve virion proteins |
| DNAse I | Promega | M6101 | Degrade cellular DNA from extracts |
| 95% ethanol | Sigma-Aldrich | 6B-100 | Virus DNA precipitation |
| Laboratory blender | VWR | 58984-030 | Grind leaf samples |
| Floor model ultracentrifuge &Ti70 rotor | Beckman Coulter, Irving TX | A94471 | Separation of cellular extracts |
| Floor model centrifuge and JA-14 rotor | Beckman Coulter | 369001 | Separation of cellular extracts |
| Magnetic stir plate | VWR | 75876-022 | Mixing urea into samples overnight |
| Rubber policeman | VWR | 470104-462 | Dissolve virus pellet |
| 2100 bioanalyzer Instrument | Agilent Genomics, Santa Clare, CA | G2939BA | Sensitive detection of DNA and RNA quality and quantity |
| 2100 Bioanalyzer RNA-Picochip | 5067-1513 | Microfluidics chip used to move, stain and measure RNA quality in a 2100 Bioanalyzer | |
| 2100 Bioanalyzer DNA-High Sensitive chip | 5067-4626 | Microfluidics chip used to move, stain and measure DNA quality in a 2100 Bioanalyzer | |
| Nanodrop spectrophotometer | Thermo-Fisher | ND-2000 | Analysis of DNA/RNA quality at intermediate steps of procedures |
| Plant total RNA isolation kit | Sigma-Aldrich | STRN50-1KT | Isolate RNA for RNA-seq |
| RNase-free water | VWR | 10128-514 | Resuspension of DNA and RNA for NGS |
| RNA concentrator spin column | Zymo Research, Irvine, CA | R1013 | Prepare RNA for RNA-seq |
| rRNA removal kit | Illumina, San Diego, CA | MRZPL116 | Prepare RNA for RNA-seq |
| DynaMag-2 Magnet | ThermoFisher | 12321D | Prepare RNA for RNA-seq |
| RNA enrichment system | Roche | 7277300001 | Prepare RNA for RNA-seq |
| Agarose | Thermo-Fisher | 16500100 | Gel analysis of DNA/RNA quality at intermediate steps of procedures |
| Ethidium bromide | Thermo-Fisher | 15585011 | Agarose gel staining |
| pGEM-T +JM109 competent cells | Promega, Madison, WI | A3610 | Clone genome fragments |
| pFU Taq polymerase | Promega | M7741 | PCR amplify virus genome |
| dNTPs | Promega | U1511 | PCR amplify virus genome |
| PCR oligonucleotides | IDT, Coralvill, IA | Custom order | PCR amplify virus genome |
| Miniprep DNA purification kit | Promega | A1330 | Plasmid DNA purification prior to sequencing |
| PCR clean-up kit | Promega | A9281 | Prepare PCR products for cloning |
| pDRAW32 software | ACAClone | Computer analysis of circular DNA and motifs | |
| MEGA6.0 software | MEGA | Molecular evolutionary genetics analysis | |
| Primer 3.0 | Simgene.com | ||
| Quant-iT™ RiboGreen™ RNA Assay Kit | Thermo-Fisher | R11490 | Fluorometric determination of RNA quantity |
| GS Junior™ pyrosequencing System | Roche | 5526337001 | Sequencing platform |
| GS Junior Titanium EmPCR Kit (Lib-A) | Roche | 5996520001 | Reagents for emulsion PCR |
| GS Jr EmPCR Bead Recovery Reagents | Roche | 5996490001 | Reagents for emulsion PCR |
| GS Junior EmPCR Reagents (Lib-A) | Roche | 5996538001 | Reagents for emulsion PCR |
| GS Jr EmPCR Oil & Breaking Kit | Roche | 5996511001 | Reagents for emulsion PCR |
| GS Jr Titanium Sequenicing kit* | Roche | 5996554001 | Includes sequencing reagents, enzymes, buffers, and packing beads |
| GS Jr. Titanium Picotiter Plate Kit | Roche | 5996619001 | Sequencing plate with associated reagents and gaskets |
| IKA Turrax mixer | 3646000 | Special mixer used with Turrax Tubes | |
| IKA Turrax Tube (specialized mixer) | 20003213 | Specialized mixing tubes with internal rotor for creating emulsions | |
| GS Nebulizers Kit | Roche | 5160570001 | Nucleic acid size fractionator for use during library preparations |
| GS Junior emPCR Bead Counter | Roche | 05 996 635 001 | Library bead counter |
| GS Junior Bead Deposition Device | Roche | 05 996 473 001 | Holder for Picotiter plate during centrifugation |
| Counterweight & Adaptor for the Bead Deposition Devices | Roche | 05 889 103 001 | Used to balance deposition device with picotiter plate centrifugation |
| GS Junior Software | Roche | 05 996 643 001 | Software suite for controlling the instrument, collecting and analyzing data |
| GS Junior Sequencer Control v. 3.0 | Roche | (Included in item 05 996 643 001 above) | |
| GS Run Processor v. 3.0 | Roche | (Included in item 05 996 643 001 above) | |
| GS De Novo Assembler v. 3.0 | Roche | (Included in item 05 996 643 001 above) | |
| GS Reference Mapper v. 3.0 | Roche | (Included in item 05 996 643 001 above) | |
| GS Amplicon Variant Analyzer v. 3.0 | Roche | (Included in item 05 996 643 001 above) |
Ссылки
- Dijkstra, J., Jager, C. P. Practical Plant Virology : Protocols and Exercises. , Springer-Verlag. Berlin Heidelberg. 1 edn (1998).
- Roossinck, M. J. Plant virus metagenomics: biodiversity and ecology. Annu Rev Genet. 46, 359-369 (2012).
- Melcher, U., et al. Evidence for novel viruses by analysis of nucleic acids in virus-like particle fractions from Ambrosia psilostachya. J Virol Methods. 152 (1-2), 49-55 (2008).
- Stobbe, A. H., Schneider, W. L., Hoyt, P. R., Melcher, U. Screening metagenomic data for viruses using the e-probe diagnostic nucleic Acid assay. Phytopathology. 104 (10), 1125-1129 (2014).
- Borah, B. K., et al. Bacilliform DNA-containing plant viruses in the tropics: commonalities within a genetically diverse group. Mol Plant Pathol. 14 (8), 759-771 (2013).
- Bousalem, M., Douzery, E. J., Seal, S. E. Taxonomy, molecular phylogeny and evolution of plant reverse transcribing viruses (family Caulimoviridae) inferred from full-length genome and reverse transcriptase sequences. Arch Virol. 153 (6), 1085-1102 (2008).
- Geering, A. D., et al. Banana contains a diverse array of endogenous badnaviruses. J Gen Virol. 86, Pt 2 511-520 (2005).
- Kunii, M., et al. Reconstruction of putative DNA virus from endogenous rice tungro bacilliform virus-like sequences in the rice genome: implications for integration and evolution. BMC Genomics. 5, 80(2004).
- Laney, A. G., Hassan, M., Tzanetakis, I. E. An integrated badnavirus is prevalent in Figure germplasm. Phytopathology. 102 (12), 1182-1189 (2012).
- Gambley, C. F., Geering, A. D., Steele, V., Thomas, J. E. Identification of viral and non-viral reverse transcribing elements in pineapple (Ananas comosus), including members of two new badnavirus species. Arch Virol. 153 (8), 1599-1604 (2008).
- Gayral, P., et al. A single Banana streak virus integration event in the banana genome as the origin of infectious endogenous pararetrovirus. J Virol. 82 (13), 6697-6710 (2008).
- Lyttle, D. J., Orlovich, D. A., Guy, P. L. Detection and analysis of endogenous badnaviruses in the New Zealand flora. AoB Plants. 2011, 008(2011).
- Le Provost, G., Iskra-Caruana, M. L., Acina, I., Teycheney, P. Y. Improved detection of episomal Banana streak viruses by multiplex immunocapture PCR. J Virol Methods. 137 (1), 7-13 (2006).
- Singh, K., Talla, A., Qiu, W. Small RNA profiling of virus-infected grapevines: evidences for virus infection-associated and variety-specific miRNAs. Funct Integr Genomics. 12 (4), 659-669 (2012).
- Alfson, K. J., Beadles, M. W., Griffiths, A. A new approach to determining whole viral genomic sequences including termini using a single deep sequencing run. J Virol Methods. 208, 1-5 (2014).
- Kreuze, J. F., et al. Complete viral genome sequence and discovery of novel viruses by deep sequencing of small RNAs: a generic method for diagnosis, discovery and sequencing of viruses. Virology. 388 (1), 1-7 (2009).
- Zheng, Y., et al. VirusDetect: An automated pipeline for efficient virus discovery using deep sequencing of small RNAs. Virology. 500, 130-138 (2017).
- James, A. P., Geijskes, R. J., Dale, J. L., Harding, R. M. Molecular characterisation of six badnavirus species associated with leaf streak disease of banana in East Africa. Annals of Applied Biology. 158 (3), 346-353 (2011).
- Baranwal, V. K., Sharma, S. K., Khurana, D., Verma, R. Sequence analysis of shorter than genome length episomal Banana streak OL virus like sequences isolated from banana in India. Virus Genes. 48 (1), 120-127 (2014).
- Sukal, A., Kidanemariam, D., Dale, J., James, A., Harding, R. Characterization of badnaviruses infecting Dioscorea spp. in the Pacific reveals two putative novel species and the first report of dioscorea bacilliform RT virus 2. Virus Res. 238, 29-34 (2017).
- BÖmer, M., Turaki, A. A., Silva, G., Kumar, P. L., Seal, S. E. A sequence-independent strategy for amplification and characterisation of episomal badnavirus sequences reveals three previously uncharacterised yam badnaviruses. Viruses. 8 (7), (2016).
- Momol, M. T., Lockhart, B. E. L., Dankers, H., Adkins, S. Canna yellow mottle virus detected in Canna in Florida. Plant Health Progress. , August 2-4 (2004).
- Zhang, J., et al. Characterization of Canna yellow mottle virus in a new host, Alpinia purpurata, in Hawaii. Phytopathology. 107 (6), 791-799 (2017).
- Wijayasekara, D., et al. Molecular characterization of two badnavirus genomes associated with Canna yellow mottle disease. Virus Res. 243, 19-24 (2018).
- Covey, S. N., Noad, R. J., al-Kaff, N. S., Turner, D. S. Caulimovirus isolation and DNA extraction. Methods Mol Biol. 81, 53-63 (1998).
- Sambrook, J., Fritsch, E. F., Maniatis, T. Molecular cloning: A laboratory manual. 2nd edn. , Cold Spring Harbor Press. (1989).
- Radford, A. D., et al. Application of next-generation sequencing technologies in virology. J Gen Virol. 93, Pt 9 1853-1868 (2012).
- Kanagal-Shamanna, R. Emulsion PCR: Techniques and Applications. Methods Mol Biol. 1392, 33-42 (2016).
- Getts, D. R., et al. Targeted blockade in lethal West Nile virus encephalitis indicates a crucial role for very late antigen (VLA)-4-dependent recruitment of nitric oxide-producing macrophages. J Neuroinflammation. 9, 246(2012).
- van Dijk, E. L., Jaszczyszyn, Y., Thermes, C. Library preparation methods for next-generation sequencing: tone down the bias. Exp Cell Res. 322 (1), 12-20 (2014).
- Gel filtration principles and methods. GE Healthcare. , (2010).
- Goff, S., et al. The iPlant Collaborative: Cyberinfrastructure for Plant Biology. Frontiers in Plant Science. 2, (2011).
- Lin, Z., et al. Next-generation sequencing and bioinformatic approaches to detect and analyze influenza virus in ferrets. J Infect Dev Ctries. 8 (4), 498-509 (2014).
- Artimo, P., et al. ExPASy: SIB bioinformatics resource portal. Nucleic Acids Res. 40, Web Server issue 597-603 (2012).
- Edgar, R. C. MUSCLE: a multiple sequence alignment method with reduced time and space complexity. BMC Bioinformatics. 5, 113(2004).
- Hung, J. H., Weng, Z. Sequence Alignment and Homology Search with BLAST and ClustalW. Cold Spring Harb Protoc. 2016 (11), (2016).
- Sohpal, V. K., Dey, A., Singh, A. MEGA biocentric software for sequence and phylogenetic analysis: a review. Int J Bioinform Res Appl. 6 (3), 230-240 (2010).
- Untergasser, A., et al. Primer3--new capabilities and interfaces. Nucleic Acids Res. 40 (15), 115(2012).
- Dhaliwa, A. DNA extraction and purification. Mater Methods. 3, 191(2013).
- Moeller, J. R., Moehn, N. R., Waller, D. M., Givnish, T. J. Paramagnetic cellulose DNA isolation improves DNA yield and quality among diverse plant taxa. Appl. Plant Sci. 2 (10), (2014).
- Moeller, J. R., et al. Paramagnetic cellulose DNA isolation improves DNA yield and quality among diverse plant taxa. Appl. Plant Sci. 2 (10), (2014).
- Grooms, K. Review: Improved DNA Yield and Quality from Diverse Plant Taxa. , (2015).
- Nishimori, A., et al. In vitro and in vivo antivirus activity of an anti-programmed death-ligand 1 (PD-L1) rat-bovine chimeric antibody against bovine leukemia virus infection. PLoS One. 12 (4), 0174916(2017).
- Rojas, M. R., Gilbertson, R. L. Plant Virus Evolution. Roossinck, M. J. 1, Springer-Verlag. 27-51 (2008).
- Roossinck, M. J. The big unknown: plant virus biodiversity. Curr Opin Virol. 1 (1), 63-67 (2011).
- Roossinck, M. J., Martin, D. P., Roumagnac, P. Plant Virus Metagenomics: Advances in Virus Discovery. Phytopathology. 105 (6), 716-727 (2015).
- Momol, M. T., Lockhart, B. E. L., Dankers, H., Adkins, S. Plant Health Progress. , Online (2004).
- Eni, A., Hughes, J. D., Asiedu, R., Rey, M. Sequence diversity among badnavirus isolates infecting yam (Dioscorea spp.). Archives of Virology. 153 (12), Ghana, Togo, Benin and Nigeria. 2263-2272 (2008).
- Harper, G., et al. The diversity of Banana streak virus isolates in Uganda. Arch Virol. 150 (12), 2407-2420 (2005).
- Muller, E., Sackey, S. Molecular variability analysis of five new complete cacao swollen shoot virus genomic sequences. Arch Virol. 150 (1), 53-66 (2005).
Перепечатки и разрешения
Запросить разрешение на использование текста или рисунков этого JoVE статьи
Запросить разрешениеСмотреть дополнительные статьи
This article has been published
Video Coming Soon
Авторские права © 2025 MyJoVE Corporation. Все права защищены