
Method Article
Визуализация узлов и Notochordal пластины в Gastrulating эмбрионов мыши, используя растровая электронная микроскопия и вся гора иммунофлуоресценции
В этой статье
Резюме
Notochordal пластины и узлов являются временными, сигнализации организаторы в развитии эмбрионов мыши, которые могут быть визуализированы с помощью нескольких методов. Здесь, мы подробно описывают, как выполнять два из методов, чтобы изучить их структуру и Морфогенез: 1) растровая электронная микроскопия (SEM); и 2) вся гора иммунофлюоресценции (WMIF).
Аннотация
Мыши после имплантации эмбриона претерпевает изменения основных формы после начала гаструляция и морфогенеза. Отличительной чертой морфогенеза является формирование переходных организаторов, узел и notochordal плиты, из клеток, которые прошли через примитивных полоска. Надлежащее формирование этих сигналов центров имеет важное значение для разработки плана тела и методы, чтобы визуализировать их представляют большой интерес для развития биологов мыши. Узел и notochordal плиты лежат на вентральной поверхности gastrulating эмбрионов мыши вокруг эмбриональных день (E) 7.5 развития. Узел является структурой Чашеобразная, клетки которых имеют одну стройную ресничку. Правильной субцеллюлярные локализации и вращение реснички в яму узла определяет асимметрия слева направо. Notochordal плита клетки также обладают одной реснички хотя короче, чем те из узла клеток. Notochordal плита образует хорда, которое выступает организатором важных сигналов для somitogenesis и нейронных патронирования. Потому что клетки узла и notochordal пластины временно присутствуют на поверхности и обладают ресничек, они могут быть визуализированы с помощью растровая электронная микроскопия (SEM). Среди других методов, используемых для визуализации этих структур на клеточном уровне является вся гора иммунофлюоресценции (WMIF) с использованием антител против белки, которые сильно выражены в узел и notochordal пластины. В настоящем докладе мы описываем наши оптимизированные протоколы для выполнения SEM и WMIF узла и notochordal пластины в развивающихся эмбрионов мыши чтобы помочь в оценке формы тканей и клеточных Организации в одичал типа и мутантов гаструляция эмбрионов.
Введение
Решающее значение для формирования зародыша мыши1гаструляция и сопровождающих морфогенетических движений. Изменения в клеточных фигуры и Организации в ходе морфогенеза диктовать информацию для регулирования судьбу клеток и также позволяют последовавшей сигнальные пути точно выполнять свои функции по диверсификации новообразованной зародышевых1. Формирование переходных структур Организации и сигнализации центров, таких как узел и хорда имеет важное значение для выполнения программы развития2. Развития биологов использовали различные методы для изучения морфогенез этих структур, наиболее заметным из которых является использование сотовой репортеров и живой ex vivo изображений следовать динамику в клеточном и субклеточном поведение2 ,3,4. В настоящем докладе, мы ориентируемся на с подробными сведениями о нашем оптимизирован протоколов для двух из этих методов: сканирование электронная микроскопия (SEM) и вся гора иммунофлюоресценции (WMIF), которые были и по-прежнему важную роль в изучении морфогенез узла и notochordal плиты, предшественник хорда.
Мыши эмбриональные узел является каплевидные Кубок клеток, который расположен на вентральной поверхности эмбриона мыши вокруг начала в конце головы фолд стадии во время гаструляция и морфогенез (эмбриональных день, E7.5-E8)2,5, 6,7. Notochordal плита морфологически кпереди исходит от узел3. Каждая ячейка в узел и notochordal плиты характеризуется одну ресничку, что выступает наружу, который длиннее в клетках узел, но длина которого изменяется в зависимости от стадии развития2. Вращение реснички в узел яму было показано, быть важным для сигнализации, определяющий левый правый асимметрия4. Notochordal пластина является прекурсором хорда, сигнальный центр, который имеет важное значение для патронирования соседние сегменты и вышележащих Нервная трубка3.
Поскольку атрибуты расположения (поверхности), формы (Кубок) и обладающие собственный внешний клеточных структур (реснички) SEM традиционно используется для визуализации узла и notochordal пластины и изучить их формирования и структуру2, 7. МДж также используется для изучения изменений в структуре самого узла или ресничек на клетки в мутаций, затрагивающих гаструляция, морфогенез, а также реснички формирования8,9,10. SEM это техника, которая использует фокусированный луч электронов допрос топологических Ультраструктура внешней поверхности материалов, таких как биологические образцы11. Этот образец обычно фиксированной, сушат и затем распыления покрытием с металлами для наблюдения под сканирующий электронный микроскоп, как мы описали в шаге 1.
WMIF является методом окрашивания визуализировать гена продуктов, таких как белки, в трех измерениях (3D). WMIF тканей, органов или даже целых организмов обеспечивает пространственной информации о распределении сигнала и форму результирующей структуры в 3D. Методика основана на фиксации образца, затем окрашивание с флуоресцентные конъюгатов. Мышь эмбрионов ~ E7.5 являются небольшими и прозрачным и поэтому идеально подходит для WMIF протоколы для визуализации узла и notochordal пластины. К примеру транскрипционный фактор, который Barchyury (T) выражается в ядрах узла и notochordal плиты и в меньшей степени в примитивных полоска, вокруг E7.5-E8 эмбрионального развития и хорошей рабочей антител против T WMIF являются коммерчески доступные и сделать возможным пятная процедуру. Клетки узла и notochordal пластины характеризуются свёрнутые апикальной поверхности, которые сталкиваются снаружи и таким образом могут быть окрашены с конъюгированных флуоресценции Фаллоидин знак F-актина в апикального сужения. Используя в качестве примеров этих реагентов, комбинация T и F-актина, окрашивание, WMIF обеспечивает представление узла и notochordal пластины в 3D в gastrulating эмбрионов мыши как мы показываем в шаге 8. Однако маркеры реснички, таких как ARL13B или ацетилированный тубулин, а также другие маркеры узла и notochordal плиты, таких как FOXA2, могут также использоваться для выполнения WMIF мыши развивающиеся эмбрионы3,4.
Мы показали, что striatin взаимодействия протеин 1 (STRIP1) имеет важное значение для нормального гаструляция и морфогенез в эмбриона мыши8. STRIP1 — это основной компонент striatin взаимодействующих фосфатаз и киназы комплексов (STRIPAK), которые мы и другие причастны актина цитоскелета организации8,12. Дефект Strip1 мутант эмбрионов в основных находится в формировании осевой мезодермы (узел и notochordal плиты) и расширение тела Антеро задней оси. Мы использовали SEM и WMIF для анализа узел и notochordal пластины в одичал тип (WT) и Strip1 мутант эмбрионов, как мы покажем в Представитель результаты и соответствующие показатели.
протокол
Все эксперименты с участием эксперименты на животных были утверждены соответствующими властями в Северный Рейн-Вестфалия (LANUV-NRW).
1. сканирующая электронная микроскопия мыши эмбриональные узла
- Жертву беременных женщин мыши на ~ E7.5 (2-4 Сомит стадии), шейки матки дислокации. Подробное объяснение с диаграммами шаги 1.1 - 1.7 доступен в мыши эмбриона лабораторных руководства13.
- Откройте живота через кожу и мезентерии и удаление матки с помощью ножниц и тонкой щипцами.
- Промойте матки кратко в дистиллированной воде и поместите его в небольшой чистый Петри (6 см) содержащий 1 x фосфат амортизированное saline (PBS).
- Под рассекает микроскопом и с помощью тонкой щипцы удалите мышцы матки, чтобы освободить отдельных deciduae или имплантации сайтов.
- Держите каждого decidua с одной парой щипцов и использовать другие пары, чтобы сделать продольный разрез полный толщина между красную часть (будущая плацента) и белая часть (где расположен эмбриона). Сделайте поверхностные перфорации вертикально вдоль белой частью decidua граничащей с разрез. Вытяните decidua друг от друга по горизонтали на две половинки и тщательно зачерпнуть из эмбриона в белую часть decidua.
- Передавать эмбриона новой Петри (35 мм) с свежими стерильной фильтрации PBS. Повторите для всех deciduae/эмбрионы.
- Удаление Райхерт мембраны, сравнительно непрозрачный мембраны, охватили эмбриона, от каждого эмбриона дразнить его прочь как носок, начиная ectoplacental конус (красноватые имплантации сайт). Для генотипирования, взять маленький кусочек (~ 0,1 мм2) из желточного мешка на данном этапе.
- Химические вытяжки и ношение надлежащей защиты (перчатки) передать эмбрионы EM класс фиксатором, состоящий из глютаральдегид 2,5% в стерильной фильтрации PBS в пробки microcentrifuge (1,5 мл) при комнатной температуре. Исправить эмбрионы на ночь при 4 ° C.
- Тщательно Удалите фиксатор глютаральдегид из трубки без касатьться эмбрионы и отменить в надлежащий контейнер для отходов. Вымойте эмбрионы три раза в стерильной фильтрации PBS, за 15 мин при комнатной температуре.
- Обезвоживает эмбрионов в серии этанола за 5 мин: 50%, 70%, 85% и 100% или абсолютного этанола в три раза. Хранение эмбрионов в −20 ° C в этанол или перейти сразу к следующему шагу.
- Трансфер эмбрионов в этаноле корзины для критической точки, сушка (ДСП) в критической точке сушилка машины. Заполните камеры с этанолом для покрытия корзины полностью.
- Обмен этанола, перемещая тщательно с жидкие CO2 по десять раз на 10 ° C. Слить жидкость CO2 после последнего шага, до тех пор, пока Палата наполовину. Нагрейте до 40 ° C до тех пор, пока давление достигает 80 баров (критическая точка) и жидкие CO2 изменяется на газ. Подождите 10 минут, а затем медленно Удар подачу газа в течение примерно 45 минут.
- Альтернативой является легче ДСП для сушки добавьте Гексаметилдисилазан (ГМДО) в соотношении 1:1 для эмбрионов в этанол 30 мин. Затем передать эмбрионы чисто HMDS 30 мин удалить эмбрионов жидкости с помощью пипетки и оставляют для просушки на 30 мин.
Примечание: Оба сушки методы работали одинаково хорошо в наших руках. - Использование тонкой кистью для монтирования сушеные эмбрионов с вентральной стороне (узел) вверх на SEM заглушки с двухсторонней ленты.
- Вставьте заглушки с эмбрионов в распыления покрытия машины для золота частиц покрытия, которое предпочитают брать длинные тонкие реснички. Нанесите слой 120-150 Å; время зависит от тока, который будет меняться с каждой выборки.
- Поместите покрытием заглушки с эмбрионами в Микроскоп SEM, применять вакуум и соблюдать эмбриональных узел и notochordal плиты клетки с реснички в увеличениях, начиная от 1000 X 15, 000 X.
2. Вся гора иммунофлюоресценции узел мыши и Notochordal плиты
- С помощью ледяной PBS с 0,05% Tween-20 (PBSTw), выполните шаги 1.1-1.7 выше чтобы удалить эмбрионов в E7.75 и поместить их в PBSTw в 35 мм Петри на льду.
- Химические вытяжки и ношение надлежащей защиты (перчатки) передать фиксирующие решение параформальдегида 4% в PBS в пробки microcentrifuge эмбрионов. Исправить эмбрионы на ночь при 4 ° C.
- Осторожно снимите фиксатор параформальдегида из трубки без касатьться эмбрионы и отменить в надлежащий контейнер для отходов. Вымойте эмбрионы три раза в PBS, содержащие 0,2% Тритон X-100 (PBSTr) за 5 мин при комнатной температуре. Выполните все мыть и последующих шагов инкубации на шейкере nutating.
- Удалить последний мыть и добавить блокирование раствор, содержащий PBSTr тепло инактивированной сывороткой 10% (от видов принимающих вторичное антитело). Блок с 2 часов ночи (или более) на nutator при 4 ° C.
- Снять блокировку и добавить ~ 1 мл основное антитело, разбавленный преграждая разрешение, например антитела анти T при разбавлении 1: 500. Инкубируйте на ночь (или больше) на nutator при 4 ° C.
- Удаление основного антитела и сохранить его для последующего использования, добавив азид натрия до конечной концентрации 0,02% (1 мкл 20% акций в 1 мл раствора антител). Антитела могут быть повторно использованы ~ 10 раз. Промойте эмбрионов дважды с PBSTr и затем промойте их в три раза по 30 мин на nutator при 4 ° C.
- Заменить мыть конъюгированных флуоресценции вторичное антитело, против видов хост основного антитела, разводят на ~ 1: 1000 на ночь (или больше) на nutator при 4 ° C.
- Удаление вторичных антител и дважды промыть PBSTr, а затем вымыть три раза по 30 мин с PBSTr.
- Замените последний мыть PBSTr содержит конъюгированных флуоресценции Фаллоидин 1: 500, чтобы пятно F-актина и 1: 1000 DAPI, пятно ядер, за 1 ч при комнатной температуре.
- Промойте дважды в PBSTr и мыть раз с PBSTr за 30 мин при комнатной температуре.
- Заменить PBSTr с PBS и оставить эмбрионы на льду. Подготовьте чистый положительно заряженных слайды (60 x 24 мм) и coverslips (24 x 24 мм) и водной основе глицерина монтаж средств массовой информации, например, 90% глицерина в 1xPBS и antifade реагент, чтобы смонтировать эмбрионов.
- Положите два куска прозрачной клейкой ленты на расстоянии ~ 15 мм друг от друга на прозрачную часть слайда. Это позволит создать достаточно 3D пространстве (в измерении Z), которая позволила бы уплощение эмбрионов, но не полностью squishing них.
- Под микроскопом рассечения тщательно перемещение эмбрионов, с помощью отрезока Р200 пипетки (обеспечить достаточное пространство для эмбриона быть передана и не повреждены) в слайд.
- С помощью тонкой щипцы, сделайте два полное сокращений на боковых гранях желточного мешка разворачиваться эмбриона. Место эмбриона с вентральной стороне (узел и notochordal плиты) до (спинной невральной трубки вниз на слайде).
- Добавьте 50 мкл монтажа СМИ на эмбрион. Место 4-5 эмбрионов на слайд. Добавьте каплю монтажа СМИ в сторону coverslip, которая будет сначала коснитесь слайда (верхней или нижней стороне), затем поместите его трансграничным двух кусков ленты и медленно опустите его на эмбрионов, с помощью тонкой щипцами или изогнутых тонкой иглой избегая создания пузырьков воздуха.
- Очистите избыток монтажа СМИ с использованием абсорбирующего стереть. Будьте осторожны, чтобы не перемещать coverslip в процессе.
- С помощью щедрой количество лак, печать по бокам coverslip без его перемещения.
- Соблюдайте под Сканирующий конфокальный микроскоп.
Результаты
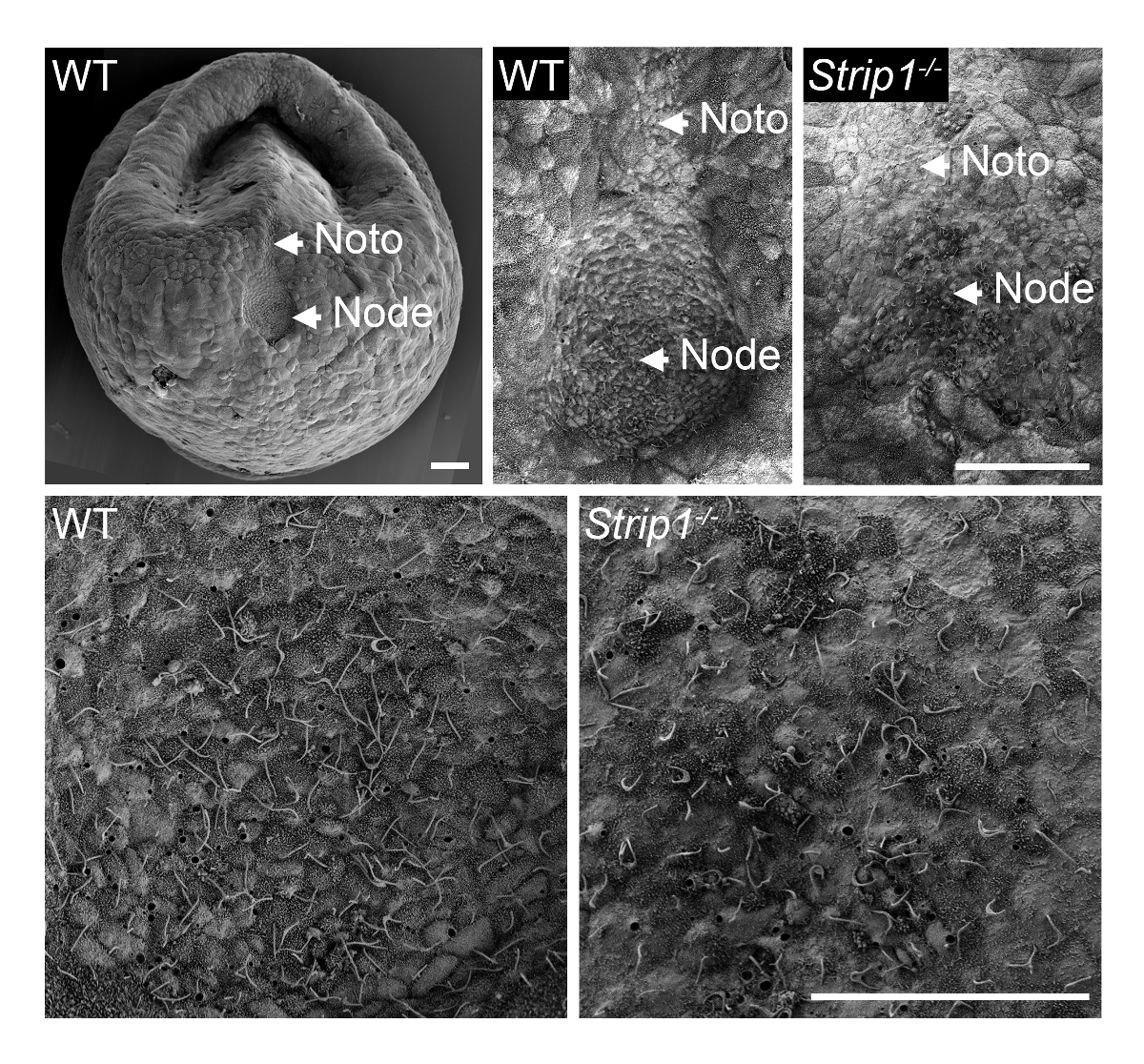
С целью изучения формирования узла в WT и Strip1 мутант эмбрионов на ~ E7.5, мы использовали SEM, как описано в шаге 1 и показано на рисунке 18. Ультраструктурные детали вне топологии с помощью SEM были весьма информативным и было сразу ясно, что в отличие от Пит образный узел в WT эмбрионов, мутант эмбрионов плоский и нерегулярных узел. Увеличение эмбрионов показали характерные ресничек на узел клетки, которые определены их однозначно. Насыпная плотность нижней реснички в мутанта может объясняться с потерей узла структуры ямы и кривизны или меньшее количество клеток узел. Notochordal пластина, которая появляется вытекающих из узла был также нерегулярные мутант эмбрионов. Они были идентифицировать их короче ресничек. Таким образом SEM важно выявить дефекты морфогенеза узел в Strip1 мутантов8. Мы также использовали SEM в предыдущих исследованиях показать отсутствие реснички в узле эмбриональных мутантов, которые не имеют centrioles, которые предоставляют шаблон для ресничек9.
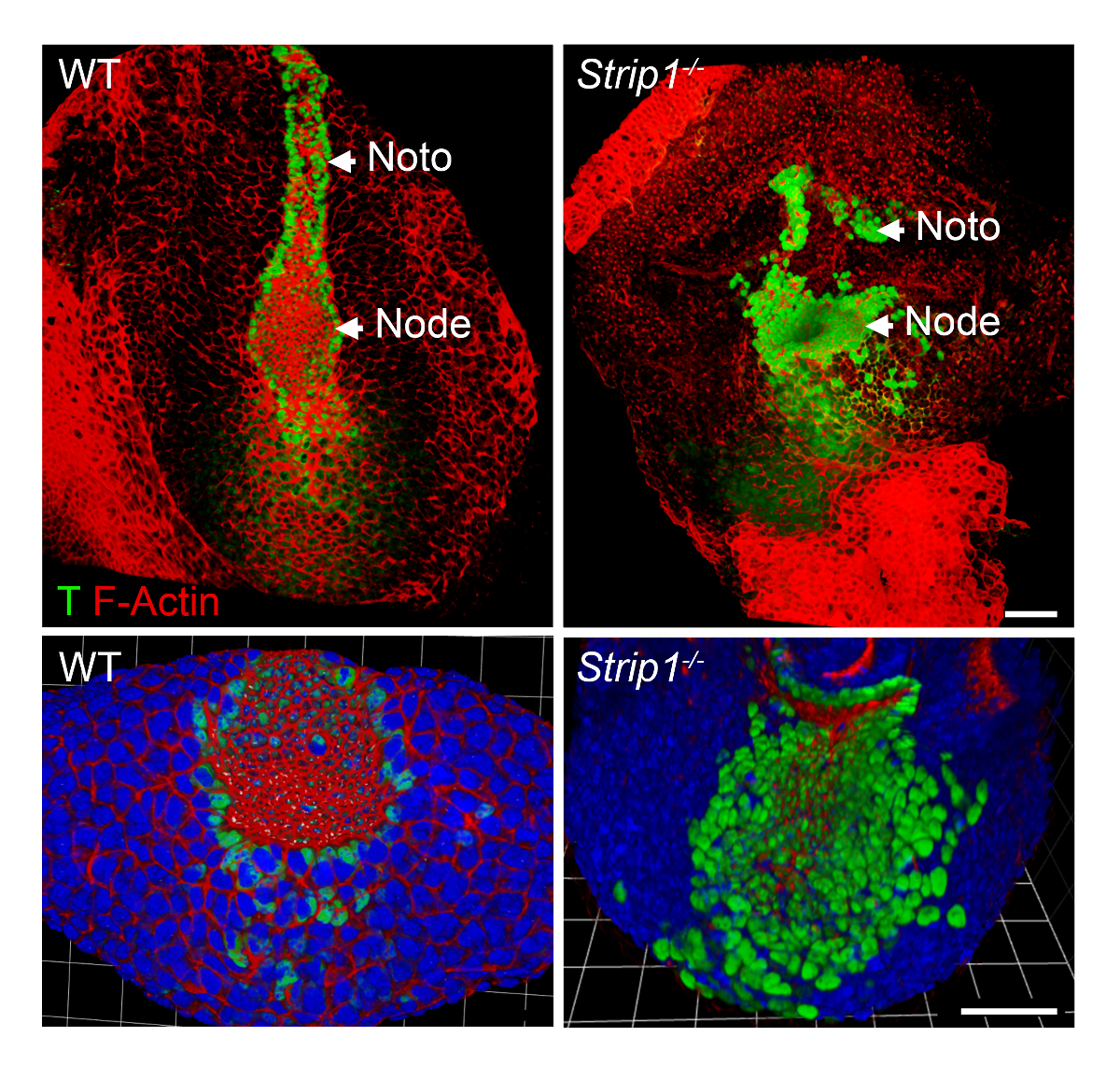
Для изучения дефектов формирования осевой мезодермы в Strip1 мутант эмбрионов на клеточном уровне, мы использовали WMIF как описано в шаге 2 и показано на рисунке 2. Используя эту технику, узел и notochordal пластины были легко идентифицируется F-актина и T окрашивания. WT узел и notochordal плиты клетки constricted апикальной домены, где F-актина обогатилась, и ядерной T окрашивание было очевидно. Notochordal плита рострально продлен в WT, но была короткой и нерегулярных в мутанта. Эти данные показали, что F-актина Организации является ненормальным в различных зародышевых мутант эмбрионов, включая осевой мезодермы8. Таким образом WMIF сыграл изучать дефекты в узел и формирование notochordal пластину в Strip1 мутант эмбрионов.

Рисунок 1 . Сканирующая электронная микроскопия выявляет дефекты в узел морфогенеза в Strip1 мутант мыши эмбрионов. (Вверху) SEM анализ WT и Strip1 мутант вентральной эмбриональных узлов и notochordal пластин (Noto)8. Слева приведен пример изображения малое увеличение WT эмбриона. (Внизу) Более высоких увеличениях центра узлы, показанные на вершине, раскрывая длинный monocilia, выходящий из узла клеток. Передней составляет всех панелей. Масштаб бары: 30 мкм. пожалуйста, нажмите здесь, чтобы посмотреть большую версию этой фигуры.
{kind=link}

Рисунок 2 . Вся гора иммунофлюоресценции показывает аномальные узел и notochordal пластины на клеточном уровне в Strip1 мутант эмбрионов. (Вверху) Вентральной 3D-рендеринга (Volocity программное обеспечение) из WMIF на WT и Strip1 мутант эмбрионов, используя комбинацию конъюгированных флуоресценции Фаллоидин (F-актина, красный) и T Пятнать антитела (зеленый). (Внизу) Дополнительные примеры окрашивания показано выше упором на узел с выше зум и включая DAPI. Передней составляет всех панелей. Масштаб бары: 30 мкм. пожалуйста, нажмите здесь, чтобы посмотреть большую версию этой фигуры.
{kind=link}
Обсуждение
В этой работе мы продемонстрируем выполнять SEM и WMIF для визуализации эмбриональных узел мыши и notochordal пластины. Небольшой размер gastrulating эмбрионов мыши ~ E7.5 и присутствие этих структур на поверхности делают их идеальными для исследования с использованием методов описанных2,7,8. Наличие хорошей антител, такие как маркеры T и реснички, дает отличную 3D информацию, используя WMIF на структуры, Организации и формирования этих основных эмбриональных организаторы8.
Потому что мышь эмбрионального развития продолжается очень быстрыми темпами, и узел и notochordal пластины присутствуют только временно на поверхности эмбриона, время имеет важное значение для успеха этих экспериментов2,3. Например 2-4 Сомит эмбрионов хороши для SEM анализ зрелой узел ПС с длиной ресничек. В гораздо раньше или позже эмбрионов (например, 12 h до или после) узел может присутствовать не на поверхности. WMIF в этом отношении является немного более гибким, но сами структуры также являются временными во время разработки и сроков в этом случае зависит от интересов исследователей.
Чистота реагентов также имеет важное значение для успеха этих методов, особенно в зондирующего Ультраструктура по SEM. Tiny примесей, которые приклеиваются к эмбрионов обычно приводят к огромные артефактов.
Мы протестировали два различных методов фиксации эмбриона для SEM одно используя половину Карновского фиксатором (2,5% глютаральдегид, буфера cacodylate 2% параформальдегида и 0,1 М) и проще глютаральдегид 2,5% в однократном ПБС. Мы предпочитаем использовать в низкотемпературном растворе глютаральдегида и PBS фиксатор, как описано в шаге 1, однако, мы и другие также половину Карновского фиксатором успешно используется для SEM.
Мы также по сравнению двух методов сушки эмбрионы для SEM и нашли никакой разницы в качестве образца с помощью либо критической точки сушилка HMDS как описано в шаге 1 и других сообщили14.
Шаг 2, мы проверили встраивание эмбрионов после окончательной стирки шаги в низких плавления агарозы 1% монтажа на 35 мм стекло дно блюдо и затем долива он с ~ 10 мкл монтажа среды. Этот метод внедрения работает и сохраняет оригинальный 3D структура эмбриона и связанные с ними структуры; Однако, микроскопом multiphoton требуется изображение образца, потому что регулярные Конфокальный микроскоп не может достичь так глубоко в неповрежденной эмбрионов (~ 1 мм).
Мы считаем, что использование этих двух методов дает дополнительную информацию о структуре узла и notochordal плиты во время нормального развития и мутантов, которые показывают дефекты в формировании этих структур.
Раскрытие информации
Авторы не имеют ничего сообщать.
Благодарности
H.B. поддерживается запуск финансирование медицинского факультета и SFB829 Кёльнского университета. C.X. поддерживается DFG Грант Ба 5810/1-1. Мы хотели бы поблагодарить визуализации объектов на CECAD научно-исследовательский центр и Мемориал Слоун Kettering Рак центр (Нью-Йорк, США). Мы благодарим Хоакин Грего-Бесса (Испанский Национальный центр сердечно-сосудистых исследований, Мадрид, Испания) за его представление на монтаж эмбрионы для WMIF.
Материалы
| Name | Company | Catalog Number | Comments |
| 1,1,1,3,3,3 Hexamethyldisilazane (HMDS) | Carl Roth | 3840 | |
| Anti-T antibody | R&D Systems | AF2058 | |
| Critical Point Dryer | Blazers Union | CPD 020 | |
| DAPI | AppliChem | A4099,0005 | |
| Glutardialdehyde solution 25% | Merck | 1042390250 | |
| Triton X-100 | Sigma Aldrich | X100-100ML | |
| Tween 20 | AppliChem | A4974,0500 | |
| SEM coating unit PS3 | Agar Aids for Electron Microscopy | PS3 | |
| SEM microscope Quantum FEG 250 | ThermoFisher Scientific (FEI) | Quantum FEG 250 |
Ссылки
- Rivera-Pérez, J. A., Hadjantonakis, A. K. The dynamics of morphogenesis in the early mouse embryo. Cold Spring Harbor Perspectives in Biology. , (2015).
- Lee, J. D., Anderson, K. V. Morphogenesis of the node and notochord: The cellular basis for the establishment and maintenance of left-right asymmetry in the mouse. Developmental Dynamics. 237 (12), 3464-3476 (2008).
- Balmer, S., Nowotschin, S., Hadjantonakis, A. K. Notochord morphogenesis in mice: Current understanding and open questions. Developmental Dynamics. 245 (5), 547-557 (2016).
- Yoshiba, S., et al. Cilia at the node of mouse embryos sense fluid flow for left-right determination via Pkd2. Science. , (2012).
- Jurand, A. Some aspects of the development of the notochord in mouse embryos. Journal of Embryology and Experimental Morpholog. 32 (1), 1-33 (1974).
- Poelmann, R. E. The head-process and the formation of the definitive endoderm in the mouse embryo. Anatomy and Embryology (Berl). , 41-49 (1981).
- Sulik, K., et al. Morphogenesis of the murine node and notochordal plate. Developmental Dynamics. 201 (3), 260-278 (1994).
- Bazzi, H., Soroka, E., Alcorn, H. L., Anderson, K. V. STRIP1, a core component of STRIPAK complexes, is essential for normal mesoderm migration in the mouse embryo. Proceedings of the National Academy of Sciences of the United States of America. 114 (51), 10928-10936 (2017).
- Bazzi, H., Anderson, K. V. Acentriolar mitosis activates a p53-dependent apoptosis pathway in the mouse embryo. Proceedings of the National Academy of Sciences of the United States of America. 111 (15), 1491-1500 (2014).
- Huangfu, D., Liu, A., Rakeman, A. S., Murcia, N. S., Niswander, L., Anderson, K. V. Hedgehog signalling in the mouse requires intraflagellar transport proteins. Nature. 426 (6962), 83-87 (2003).
- McMullan, D. Scanning electron microscopy 1928-1965. Scanning. 17 (3), 175-185 (2006).
- Bai, S. W., et al. Identification and characterization of a set of conserved and new regulators of cytoskeletal organization, cell morphology and migration. BMC Biology. 9, (2011).
- Behringer, R., Gertsenstein, M., Vintersen Nagy, K., Nagy, A. Manipulating the Mouse Embryo: A Laboratory Manual, Fourth Edition. Cold Harb Lab Press. , (2014).
- Braet, F., De Zanger, R., Wisse, E. Drying cells for SEM, AFM and TEM by hexamethyldisilazane: a study on hepatic endothelial cells. Journal of Microscopy. 186, 84-87 (1997).
Перепечатки и разрешения
Запросить разрешение на использование текста или рисунков этого JoVE статьи
Запросить разрешениеСмотреть дополнительные статьи
This article has been published
Video Coming Soon
Авторские права © 2025 MyJoVE Corporation. Все права защищены