
Method Article
Генотипирование и количественная оценка в Ситу Гибридизации Окрашивание в зебрафиш
В этой статье
Резюме
Технологии редактирования генов позволили исследователям генерировать мутантов зебры для исследования функции генов с относительной легкостью. Здесь мы предоставляем руководство для выполнения параллельных генотипирования эмбрионов и количественной оценки сигналов гибридизации на месте у зебры. Такой объективный подход обеспечивает большую точность в фенотипивных анализах, основанных на гибридизации in situ.
Аннотация
На месте гибридизации (ISH) является важным методом, который позволяет исследователям изучать распределение мРНК на месте и был критическим методом в биологии развития на протяжении десятилетий. Традиционно, большинство исследований экспрессии генов опирались на визуальную оценку сигнала ISH, метода, который склонен к предвзятости, особенно в тех случаях, когда идентификационные данные образцов известны априори. Ранее мы сообщали о методе, чтобы обойти эту предвзятость и обеспечить более точную количественную оценку сигналов ISH. Здесь мы представляем простое руководство по применению этого метода для количественной оценки уровней экспрессии генов, представляющих интерес для эмбрионов, окрашенных ISH, и соотносить это с их соответствующими генотипами. Этот метод особенно полезен для количественной оценки пространственно ограниченных сигналов экспрессии генов в образцах смешанных генотипов и обеспечивает объективную и точную альтернативу традиционным методам визуального скоринга.
Введение
Внедрение технологий редактирования генома (ЗФН, TALENs и совсем недавно, CRISPR/Cas9) привело к массовому увеличению числа лабораторий по всему миру, которые используют эти системы для изучения функции конкретных генов in vivo. Зебрафиш, в частности, поддаются генетическим манипуляциям, и многие мутанты были созданы в недавнем прошлом1,2. Для биологов в области развития, один из наиболее распространенных методов для оценки фенотипических последствий генных мутаций в эмбриональном развитии находится в situ гибридизации (ISH). При отсутствии очевидных морфологических дефектов, отделяющих гомозиготных мутантов от их дикого типа или гетерозиготных братьев и сестер, важно уметь точно идентифицировать различные генотипы.
Классический ISH опирается на качественный анализ интенсивности сигнала для получения выводов о регулятивных взаимодействиях между геном интереса и выбранными генами маркеров. Хотя эти анализы являются полезными, они могут быть предвзятыми ожиданиями исследователей. Таким образом, был разработан метод количественной оценки экспрессии генов после визуализации эмбрионов, окрашенных ISH, без предварительного знания соответствующего генотипа. За этим последовала эффективная экстракция ДНК и генотипирование, что позволило нам количественно соотнести генотип с экспрессией генов3. В то время как генотипирование эмбрионов после ISH был использован до4,5, изображение на основе количественной оценки ISH моделей не был широко использован, кроме нескольких исследований6,7. Наиболее популярные альтернативы полагаются на визуальный скоринг или подсчет МГ-окрашенных клеток8,9,10, как склонны к плохой воспроизводимости и исследователь смещения. Этот метод особенно полезен для изучения изменений в генах с экспрессией моделей, которые пространственно ограничены, такие как runx1 или gata2b, как выражается в ограниченном подмножество клеток аорты пола называется гемогенный эндотелий11,12.
Здесь мы стремимся предоставить практическое руководство по осуществлению количественной оценки с помощью анализа изображений с использованием Фиджи13, а также днк-экстракции и генотипирования протокола. Это предназначено для визуальной иллюстрации нашего ранее опубликованного метода3. Наш метод позволяет точно едить вариации экспрессии генов, обнаруженные ISH, и объективное назначение уровней экспрессии генов на конкретные генотипы.
протокол
Процедуры, касающиеся животных, регулируются Законом 1986 года о животных (научные процедуры) и были утверждены министерством внутренних дел и местным органом по защите животных и этическому обзору.
1. Изображение ISH-окрашенных эмбрионов
- Подготовьте раствор глицерола (50%-80% в буфере 1x PBS) и смешайте для гомогенизации раствора (например, оставьте в ролике не менее 5 мин). Это решение может храниться в течение нескольких месяцев при комнатной температуре.
- После на месте гибридизации14,15,16,17, передача эмбрионов в раствор глицерола с 3 мл Пастер пипетка и оставить, чтобы поселиться, по крайней мере 5 мин. Если изображения эмбрионов старше 24 л.с. (часы после оплодотворения), они могут быть отбелены, как описано18.
- Подготовка и этикетка достаточно ПЦР трубки для передачи ISH-окрашенных эмбрионов после изображения.
- Добавьте 100% глицерол на дно колодца в стеклянной депрессии слайд с 3 мл Пастер пипетка.
- Используя пипетку Pasteur объемом 3 мл, перенесите один эмбрион, окрашенный ISH, на стеклянную горку и ориентируйся по мере необходимости под стереомикроскоп, оснащенный цифровой камерой и нижней и верхней подсветкой.
ПРИМЕЧАНИЕ: Используйте гель загрузки пипетки советы для размещения эмбрионов для визуализации, но другие инструменты (например, щипцы, вскрытие иглы) будет в равной степени адекватным. - Используя первый эмбрион, отрегулируйте освещение и время экспозиции при нужном увеличении. Используйте эти условия для ВСЕХ эмбрионов в том же эксперименте (т.е. если визуализация 40 эмбрионов из гетерозиготного мутанта incross, убедитесь, что освещение, время экспозиции и увеличение одинаковы для всех).
- Изображение столько эмбрионов, сколько требуется. Наметьте каждое изображение уникальным номером. После визуализации перенесите эмбрион в трубку ПЦР/пластину с одинаковым номером.
ПРИМЕЧАНИЕ: Изображения должны быть сохранены как файлы TIF, но другие форматы также являются адекватными.- При необходимости удалите избыток глицерола в пЦР-трубках/пластинах.
ПРИМЕЧАНИЕ: На данный момент эмбрионы могут храниться в трубках ПЦР в течение нескольких недель при комнатной температуре.
- При необходимости удалите избыток глицерола в пЦР-трубках/пластинах.
2. Извлеките ДНА и генотип зародить ISH-окрашенные
ПРИМЕЧАНИЕ: Здесь используйте надежный и недорогой метод изоляции геномной ДНК на основе метода HoTSHOT19 с эффективностью извлечения ДНК 95%-100%3.
- После завершения изображения добавьте 40-75 qL щелочным лисисовым буфером (например, HoTSHOT) к каждой трубке.
- Инкубировать при 95 градусах по Цельсию в течение примерно 30 минут и охладить трубки до 4 градусов по Цельсию, прежде чем добавить равный объем буфера нейтрализации. Ночная инкубация при 4 градусах По Цельсию может повысить эффективность ПЦР.
ПРИМЕЧАНИЕ: На данный момент геномная ДНК может быть использована для генотипирования или храниться при -20 градусов по Цельсию до тех пор, пока это не потребуется. - Образцы генотипа с соответствующим методом (например, HRMA, RFLP)3,20,21, как требуется для мутации интереса.
- Обратите внимание на генотип, соответствующий каждому образцу (например, с использованием программного обеспечения электронной таблицы).
3. Количественная интенсивность пикселей окрашенных ISH эмбрионов (анализ изображений с использованием программного обеспечения Фиджи)
- Чтобы количественно количественно на месте гибридизации (ISH) окрашивания интенсивности сигнала, преобразовать все изображения в 8-битный серый масштаб, как описано3. Если изображения были сохранены как . TIF файлы, используйте макрос Фиджи для преобразования партии3. Кроме того, конвертировать изображения в других форматах (например, . JPG) до . TIF с использованием соответствующего программного обеспечения, а затем преобразовать в 8-битный серый с помощью Фиджи. Для удобства, вот пошаговая процедура, которая была опубликована ранее3, с некоторыми изменениями.
- Откройте изображения на Фиджи и инвертировать изображение с Edit Затем измените тип изображения на 8-битный(Изображение
- Используя инструмент выбора полигона, нарисуйте область интереса (ROI) вручную на изображении вокруг области, содержащей сигнал.
- Нажмите т, чтобы открыть рентабельность инвестиций менеджера. Используйте команду «Мера» менеджера ROI для измерения интенсивности рентабельности инвестиций. Копирование средней стоимости из окна результатов в программное обеспечение электронной таблицы.
- Переместите ту же рентабельность инвестиций, обеспечивая тот же размер и форму, что и исходный регион, в область зебры, не содержащую окрашивания. Повторите шаг 3.4 для измерения фона.
- Чтобы получить среднее пиксельную интенсивность сигнала ISH, вычесть среднее значение интенсивности фоновой области из запятнанной области для каждого эмбриона.
- Присвоить каждому значению интенсивности генотип (от шага 2.3).
4. Анализ результатов с помощью соответствующих статистических тестов
- Участок все значения на одном участке, чтобы определить любые отклонения от нормального распределения.
ПРИМЕЧАНИЕ: Нормальное распределение также может быть проверено с помощью теста Колмогорова-Смирнова или теста Шапиро-Вилк. Однако для больших размеров выборки существует высокий риск ложных срабатываний в этих тестах. - При наличии сильных отклонений от нормального распределения преобразуйте все значения (с использованием функций ln или sqrt), чтобы убедиться, что они обычно распределены перед началом работы.
- Проанализируйте различия между значениями (преобразованными при необходимости), назначенными каждому генотипу (wt vs. heterozygote vs. mutant) с помощью 2-хвостой ANOVA с 95% уровнем достоверности, что свидетельствует о равенстве дисперсий с тестом Левена и коррекцией Уэлча. Для парных сравнений между каждой парой генотипов используйте пост-специальный тест Tukey's (равные отклонения) или Games-Howell (неравные отклонения).
- Если значения обычно не распределены, несмотря на преобразование, используйте непараметрический тест (Kruskall-Wallis) для анализа различий между ранжированными значениями и тестом нескольких сравнений После Хок Данна с коррекцией Bonferroni для парной Сравнения.
- Участок непреобразованных значений (от шага 3.6) в качестве точка участков для лучшего представления результатов.
Результаты
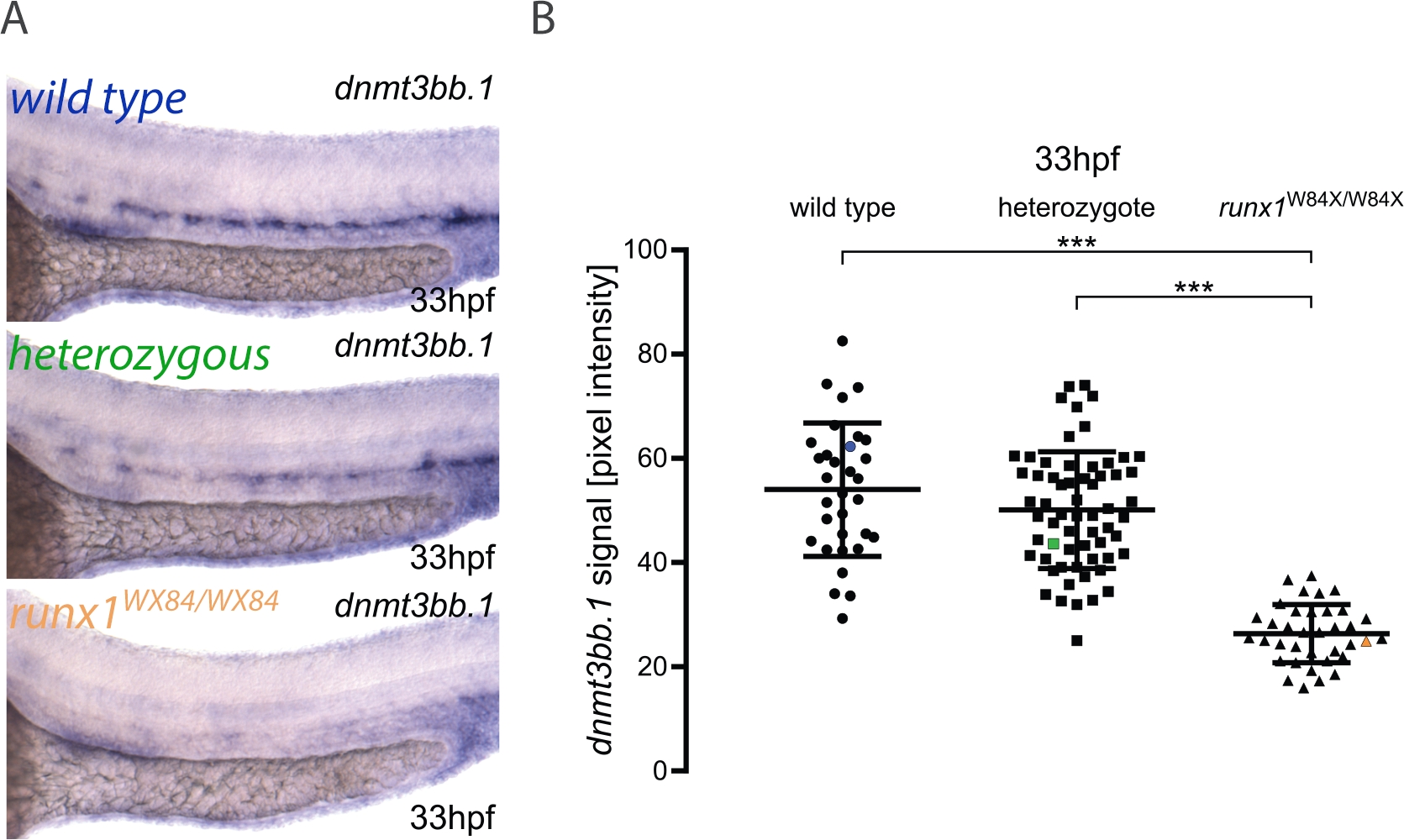
Здесь мы описываем практическое применение трубопровода для количественной оценки изображений и генотипирования эмбрионов, опубликованных в другом месте3. Рабочий процесс для метода отображается на рисунке 1. Чтобы проиллюстрировать, как использовать этот метод, ISH был выполнен для dnmt3bb.1 в 33 л.с. эмбрионов из runx1W84X / 22 incross (Рисунок 2). 130 эмбрионов были изображены с использованием тех же условий освещения, которые описаны в протоколе, и помечают их уникальным номером. После визуализации каждый эмбрион был перенесен в трубку ПЦР для генотипирования. На этом этапе был проведен анализ изображения, чтобы отнести значение интенсивности пикселя к каждому изображению. Затем генотип был назначен соответствующему изображению, а значения интенсивности пикселей сгруппированы в соответствии с их генотипом для статистического анализа. Снижение экспрессии dnmt3bb.1 было обнаружено в runx1W84X/W84X мутантов (Рисунок 2A,B)3, в согласии с предыдущими наблюдениями5. Интересно, что runx1W84X / гетерозиготных эмбрионов показали никаких значительных различий в dnmt3bb.1 выражение (Рисунок 2A,B) по сравнению с его диким типом братьев и сестер, предполагая, что одна копия Runx1 достаточно для поддержания dnmt3bb.1 выражение на соответствующих уровнях.
Многие мутанты зебры не показывают эмбриональный фенотип, который в противном случае может быть обнаружен с помощью других технологий потери функций, таких как олигонуклеотиды морфолино (МО). Это несоответствие можно отнести к ряду причин, включая вне цели эффекты, материнский белок компенсации, гипоморфный аллель23 или недавно обнаруженный феномен генетической компенсации24,25,26,27. В этом примере мы спросили, было ли уменьшено или потеряно выражение runx1 в мутантах lmo4auob100, так как ранее опубликованные данные с помощью lmo4a MO предположили, что runx1 уменьшается в морфантах lmo4a 28. Здесь, анализ показал никаких существенных различий в выражении runx1 между диким типом и lmo4auob100 гомозиготных мутантов3 (Рисунок 3A,B). Дальнейший анализ одного эмбриона qPCR показал, что было небольшое, но значительное снижение экспрессии runx1 в lmo4auob100 мутантов(рисунок 3C). Таким образом, возможно, что количественная оценка изображения не сможет обнаружить небольшие различия в уровнях выражения. Кроме того, отсутствие различий между генотипами, которые мы обнаружили, реально, и эксперименты qPCR обнаруживают изменения в экспрессии runx1 в других тканях, таких как теленчефалон, где выражены как lmo4a, так и runx1. Исследователи всегда должны проверить свои результаты с помощью независимого метода, как qPCR, но в идеале обогащения для ткани интерес с помощью цитометрии потока, например.
В редких случаях, когда ИСЗ имеет высокий фон(рисунок 3D),значение интенсивности пикселей этой области настолько высока, что вычитание из значения сигнала производит отрицательное число, и в таких случаях эти эмбрионы будут исключены из анализа. По нашему опыту, это произошло примерно в 0,4% из runx1-проверенныхэмбрионов3, но может варьироваться между экспериментами, зондами или партиями реагентов. Хотя это может быть ограничением метода, низкая частота высокого фона вряд ли повлияет на общие результаты.
Чтобы проверить влияние выбора различных областей для коррекции фона, мы сначала измерили интенсивность пикселей сигнала runx1 ISH в 28 л.с. эмбрионов, используя различные области для коррекции фона(рисунок 4). Были выбраны четыре различных региона: два в области ствола (R1 и R2), один в области желтка (неокрашенные, но, вероятно, накапливают фоновое окрашивание) и меньшую площадь передного к рентабельности инвестиций (R4, рисунок 4B). Измерение интенсивности пикселей в этих регионах показало относительно стабильную разницу в интенсивности между рентабельностью инвестиций и любой фоновой областью(рисунок 4C). Тем не менее, R3 всегда показывал очень высокие значения (выше тех, в рентабельности инвестиций). После инверсии и преобразования в 8-битную область желтка выглядит очень яркои и, таким образом, не подходит для использования в качестве фоновой коррекции. R2 был ближе к рентабельности инвестиций, но содержал некоторый сигнал ISH, и использование его для коррекции снизило среднее интенсивность пикселей по сравнению с R1 (расположен дальше дорсально, вдали от сигнала ISH) или R4. Таким образом, R1 или R4 являются подходящими областями, которые могут быть использованы для коррекции фона (несмотря на то, что площадь R4 меньше, чем у R1). Далее мы хотели сравнить, как использование R1 или R4 повлияло на результаты при сравнении выражения runx1. Для этого мы перекрещены dll4/- heterozygotes29 и проанализировали выражение runx1 в случайно выбранном диком типе и dll4-/-эмбрионах (Рисунок 4E). Хотя использование R1 или R4 для фоновой коррекции повлияло на индивидуальные значения, средняя интенсивность пикселей в рамках одного и того же генотипа существенно не отличалась(рисунок 4E). Кроме того, сравнение выражения runx1 по-прежнему дает аналогичные значения средней интенсивности между генотипами, использующими области R1 или R4 в качестве фоновой коррекции(R1No16,3 иR418,2, соответственно). Взятые вместе, мы пришли к выводу, что, хотя выбор фоновой области имеет важное значение, основным критерием является то, что она не включает желтки регионов (склонны к накоплению фонового окрашивания) и что она не должна содержать каких-либо (конкретных) окрашивания, которые могут исказить значения интенсивности пикселей фона.

Рисунок 1: Рабочий процесс параллельной количественной оценки изображения и протокола генотипирования. Эмбрионы, собранные из инкросса рыбы гетерозигот для мутантного аллеля, исследуются на измеренный ген со стандартным протоколом ISH. После визуализации геномная ДНК извлекается с помощью протокола HotSHOT путем добавления буфера лисиса непосредственно к эмбриону в 0,2 мл ПЦР трубки, а затем 30 мин инкубации при 95 градусов по Цельсию. Эта ДНК используется для генотипирования эмбрионов ПЦР, ПЦР и полиморфизмом длины фрагмента ограничения (RFLP), анализами KASP или любым другим подходящим методом. Параллельно изображения для каждого эмбриона переворачиваются и преобразуются в 8-битную серую шкалу. ROIs одинаковой формы и размера, содержащие сигнал ISH (желтый) и фон (синий) вручную подобраны и измерены. Измерения, присвоенные соответствующим генотипам, статистически анализируются. Рисунок адаптированы из Dobrzycki и др.3Пожалуйста, нажмите здесь, чтобы просмотреть большую версию этой фигуры.
{kind=link}

Рисунок 2: Количественная оценка изображения в runx1 мутантах показывает снижение уровней выражения dnmt3bb.1 ISH. (A) Пример изображения ISH в 33 л.с. дикого типа (синий), runx1q/W84X (зеленый) и runx1W84X/W84X (оранжевый) эмбрионов, показывая dnmt3bb.1 выражение в дорсальной аорты. (B) Значения интенсивности пикселей dnmt3bb.1 mRNA в эмбрионах runx1W84X/W84X(n'36) значительно снижены по сравнению с дикими типами (n-32) и гетерозиготами (n-62) (ANOVA, p qlt; 0.001). Коэффициенты вариации составляют 24%, 22% и 21% для диких групп типа, гетерозигот и мутантов соответственно. Синий, зеленый и оранжевый точки данных соответствуют примеру изображений с панели А. Бары представляют собой среднее s.d.pslt;0.001 (Games-Howell после специального теста). Рисунок адаптированы из Dobrzycki и др.3Пожалуйста, нажмите здесь, чтобы просмотреть большую версию этой фигуры.
{kind=link}

Рисунок 3: Измерение уровней экспрессии runx1 по уровню ISH в lmo4auob100мутантов. (A) Представитель изображения ISH для runx1 в 28 л.с. дикого типа (синий), heterozygous (зеленый) и lmo4auob100/uob100 (оранжевый) эмбрионов, показывая выражение в спинной аорты. (B) Количественная оценка сигнала runx1 mRNA, обнаруженные ISH, от 28 л.с. дикого типа (n'15), heterozygous lmo4aq/- (het) (n'34) и lmo4auob100/uob100 эмбрионов из одного сцепления не показывает существенной разницы в интенсивности пикселей runx1 между различными генотипами (ANOVA,sgt; p. Синий, зеленый и оранжевый точки данных соответствуют примеру изображений с панели А. Бары представляют среднее значение s.d. (C) Boxplots, показывающие нормализованные уровни runx1 mRNA (2-ЗТ)в одном диком типе (синий; n'12) и lmo4auob100/uob100 (мут, оранжевый; n'12) эмбрионы, измеренные qRT-PCR, показывая снижение уровня runx1 в mutants. р-р злт; 0,05(т тест). (D) Пример эксперимента ISH на 28 л.с. эмбриона (окрашенные для runx1, желтые наконечники стрел), показывающие высокий фон. Рисунок адаптированы из Dobrzycki и др.3Пожалуйста, нажмите здесь, чтобы просмотреть большую версию этой фигуры.
{kind=link}

Рисунок 4: Влияние коррекции фоновой интенсивности на исходы измерений. (A) Представитель изображение runx1 ISH окрашивания в диком типе эмбриона на 28 л.с. (B) То же изображение после инверсии и преобразования в 8-битный. Область интереса (ROI) выделена зеленым цветом, а четыре различных области, используемые для коррекции фона (R1-R4), выделены желтым цветом. (C) Измерения интенсивности сырьевых пикселей во всех регионах, показанных в панели B. Обратите внимание, что интенсивность в R3 (желтке) последовательно выше, чем фактический сигнал ISH в рентабельности инвестиций (n'11). (D) Уровни экспрессии Runx1 в рентабельности инвестиций с использованием фоновых областей R1, R2 и R4. Области для рентабельности инвестиций, R1, R2 и R3-28500 пикселей; R4-8500 пикселей. Обратите внимание, что фон R3 не использовался для этого сравнения, так как фоновая коррекция (ROI-R3) последовательно давала отрицательные значения. (E) Уровни экспрессии Runx1 в диком типе и dll4-/- мутанты, использующие r1 или R4 для фоновой коррекции (n'10 для каждого образца). Статистический анализ в группах D и E проводился с использованием непараметрического теста Kruskal-Wallis, предполагая, что значения интенсивности пикселей обычно не распределяются. Пожалуйста, нажмите здесь, чтобы просмотреть большую версию этой цифры.
{kind=link}
Обсуждение
При использовании этого метода следует учитывать несколько факторов для количественной оценки экспрессии генов. Условия визуализации должны поддерживаться на протяжении всего эксперимента (например, освещение, время экспозиции и позиционирование эмбриона), чтобы уменьшить изменчивость между измерениями. Критический момент заключается в том, чтобы избежать чрезмерного окрашивания образцов, так как различия в окрашивании между образцами могут быть замаскированы. Например, снижение экспрессии VegfA при отсутствии Eto2 в эмбрионах Xenopus laevis 30 может быть обнаружено только путем тщательного мониторинга окрашивания в течение 24-часового периода. Таким образом, это хорошая практика, чтобы эмпирически определить адекватные уровни окрашивания для каждого гена, которые лучше всего представляют его выражение, не достигая насыщения. Чрезмерное погревение также искусственно увеличит интенсивность фоновых пикселей в преобразованных 8-битных серых изображениях и исказит результаты количественной оценки. В крайних случаях фоновый уровень эмбриональных тканей может быть выше, чем сигнал ISH в выбранной рентабельности инвестиций, и эти образцы должны быть исключены из анализа. Аналогичное явление наблюдалось при проверке пригодности неокрашенного желтка для коррекции фона(рисунок 4). После инверсии и преобразования в 8-битные темные пиксели в желтоковой области становятся ярче, чем сигнал ISH в эмбрионе и делают фон исправленные значения отрицательными. Таким образом, избегайте использования желтка для коррекции фона. Измерение фонового сигнала в пигментированных областях эмбриона (например, глаза или тонченная часть ствола от 26/28 л.с. и далее) будет в равной степени искажать результаты количественной оценки и также следует избегать. Существуют протоколы для отбеливания эмбрионов зебры, либо до или после ISH18 и отбеливания эмбрионов старше 24 л.с. до визуализации рекомендуется.
Поскольку этот метод опирается на измерение интенсивности пикселей в определенной области на фоне интенсивности пикселей в эквивалентной неокрашенной области, он не подходит для количественной оценки вездесущих или почти вездесущих генов как есть. Вместо этого, он хорошо подходит для измерения экспрессии генов с пространственно ограниченным распределением, где область для измерения интенсивности фоновых пикселей может быть легко идентифицирована. Наш дополнительный анализ теперь показывает, что использование меньшей площади (3-4x меньше) для коррекции фона дает аналогичные результаты с использованием эквивалентной площади рентабельности инвестиций. Это расширяет применимость метода к генам, выраженным в более широких пространственных областях (и, таким образом, требующих больших рентабельности инвестиций для измерения интенсивности), до тех пор, пока можно использовать четко неокрашенные области эмбриона для фоновой коррекции.
Наконец, мы предлагаем, чтобы генотипирование выполнялось параллельно или после количественной оценки изображения, чтобы свести к минимуму предвзятость экспериментатора. Просить второго экспериментатора повторить количественную оценку на анонимизированных образцах и сравнить его с первым набором измерений также поможет уменьшить предвзятость экспериментаторов. Если изображения, которые будут количественно от сравнения между лечения, которые не требуют генотипирования (например, дикий тип против химического ингибитора или дикий тип против MO нокдаун), экспериментатор выполнения измерений должны быть ослеплены к личности Образец.
Раскрытие информации
Авторы заявляют, что у них нет конкурирующих финансовых интересов.
Благодарности
Мы хотели бы поблагодарить сотрудников биомедицинских служб в Оксфорде и Бирмингеме за отличное животноводство. T.D. финансировалась стипендией Wellcome Trust Chromosome and Developmental Biology PhD Scholarship (#WT102345/13/13/) R.M. и M.K. были профинансированы Британским фондом сердца (BHF IBSR Fellowship FS/13/50/30436) и благодарны за их щедрую поддержку. R.M. подтверждает поддержку центра передового опыта BHF (RE/13/1/30181), Оксфорд.
Материалы
| Name | Company | Catalog Number | Comments |
| 0.2 mL PCR tubes (8-strips with lids) | StarLab | A1402-3700 | 96-well plates are equally appropriate for sample handling but beware of cross contamination between samples |
| 3 mL Pasteur pipettes | Alpha Laboratories | LW4114 | |
| Cavity slides | Brand | BR475535-50EA | |
| Digital Camera (Qimaging Micropublisher 5.0) | Qimaging | ||
| Eppendorf Microloader tips | Eppendorf | 10289651 | the tips are used to orient the embryos for imaging in glycerol |
| Excel | Microsoft | ||
| F3000 Fiber Optic Cold Light Source | Photonic | ||
| Fiji | |||
| Glycerol | Sigma | G5516-1L | |
| Graphpad Prism 8.01 | GraphPad Software, Inc. | we prefer to use Graphpad, but other statistics software packages are also suitable (e.g. SigmaPlot or SPSS) | |
| HotSHOT alkaline lysis buffer | 25 mM NaOH, 0.2 mM disodium EDTA, pH 12 | ||
| HotSHOT neutralization buffer | Tris HCl 40 mM, pH 5 | ||
| PBS (10X) pH 7.4 | Thermofisher | 70011044 | |
| Stereomicroscope with illumination stand (Nikon SMZ800N) | Nikon | ||
| Thermocycler | Thermofisher |
Ссылки
- Varshney, G. K., et al. High-throughput gene targeting and phenotyping in zebrafish using CRISPR/Cas9. Genome Research. 25 (7), 1030-1042 (2015).
- Varshney, G. K., et al. CRISPRz: a database of zebrafish validated sgRNAs. Nucleic Acids Research. 44 (D1), D822-D826 (2016).
- Dobrzycki, T., Krecsmarik, M., Bonkhofer, F., Patient, R., Monteiro, R. An optimised pipeline for parallel image-based quantification of gene expression and genotyping after in situ hybridisation. Biology Open. 7 (4), bio031096 (2018).
- Bresciani, E., et al. CBFbeta and RUNX1 are required at 2 different steps during the development of hematopoietic stem cells in zebrafish. Blood. 124 (1), 70-78 (2014).
- Gore, A. V., et al. Epigenetic regulation of hematopoiesis by DNA methylation. Elife. 5, e11813 (2016).
- Fan, Y., et al. Tissue-Specific Gain of RTK Signalling Uncovers Selective Cell Vulnerability during Embryogenesis. PLoS Genetics. 11 (9), e1005533 (2015).
- Wen, B., et al. GATA5 SUMOylation is indispensable for zebrafish cardiac development. Biochimica et Biophysica Acta. 1861 (7), 1691-1701 (2017).
- Espin-Palazon, R., et al. Proinflammatory signaling regulates hematopoietic stem cell emergence. Cell. 159 (5), 1070-1085 (2014).
- Peterkin, T., Gibson, A., Patient, R. Redundancy and evolution of GATA factor requirements in development of the myocardium. Developmental Biology. 311 (2), 623-635 (2007).
- Genthe, J. R., Clements, W. K. R-spondin 1 is required for specification of hematopoietic stem cells through Wnt16 and Vegfa signaling pathways. Development. 144 (4), 590-600 (2017).
- Kalev-Zylinska, M. L., et al. Runx1 is required for zebrafish blood and vessel development and expression of a human RUNX1-CBF2T1 transgene advances a model for studies of leukemogenesis. Development. 129 (8), 2015-2030 (2002).
- Butko, E., et al. Gata2b is a restricted early regulator of hemogenic endothelium in the zebrafish embryo. Development. 142 (6), 1050-1061 (2015).
- Schindelin, J., et al. Fiji: an open-source platform for biological-image analysis. Nature Methods. 9 (7), 676-682 (2012).
- Lleras Forero, L., et al. Segmentation of the zebrafish axial skeleton relies on notochord sheath cells and not on the segmentation clock. Elife. 7, (2018).
- Jowett, T., Yan, Y. L. Double fluorescent in situ hybridization to zebrafish embryos. Trends in Genetics. 12 (10), 387-389 (1996).
- Thisse, C., Thisse, B. High-resolution in situ hybridization to whole-mount zebrafish embryos. Nature Protocols. 3 (1), 59-69 (2008).
- Narayanan, R., Oates, A. C. Detection of mRNA by Whole Mount in situ Hybridization and DNA Extraction for Genotyping of Zebrafish Embryos. Bio-protocol. , e3193 (2019).
- Monteiro, R., Pouget, C., Patient, R. The gata1/pu.1 lineage fate paradigm varies between blood populations and is modulated by tif1gamma. EMBO JOURNAL. 30 (6), 1093-1103 (2011).
- Truett, G. E., et al. Preparation of PCR-quality mouse genomic DNA with hot sodium hydroxide and tris (HotSHOT). Biotechniques. 29 (1), 52-54 (2000).
- Wilkinson, R. N., Elworthy, S., Ingham, P. W., van Eeden, F. J. A method for high-throughput PCR-based genotyping of larval zebrafish tail biopsies. Biotechniques. 55 (6), 314-316 (2013).
- Parant, J. M., George, S. A., Pryor, R., Wittwer, C. T., Yost, H. J. A rapid and efficient method of genotyping zebrafish mutants. Developmental Dynamics. 238 (12), 3168-3174 (2009).
- Jin, H., et al. Definitive hematopoietic stem/progenitor cells manifest distinct differentiation output in the zebrafish VDA and PBI. Developement. 136 (4), 647-654 (2009).
- Stainier, D. Y. R., et al. Guidelines for morpholino use in zebrafish. PLoS Genetics. 13 (10), e1007000 (2017).
- El-Brolosy, M. A., Stainier, D. Y. R. Genetic compensation: A phenomenon in search of mechanisms. PLoS Genetics. 13 (7), e1006780 (2017).
- Rossi, A., et al. Genetic compensation induced by deleterious mutations but not gene knockdowns. Nature. 524 (7564), 230-233 (2015).
- Ma, Z., et al. PTC-bearing mRNA elicits a genetic compensation response via Upf3a and COMPASS components. Nature. 568 (7751), 259-263 (2019).
- El-Brolosy, M. A., et al. Genetic compensation triggered by mutant mRNA degradation. Nature. 568 (7751), 193-197 (2019).
- Meier, N., et al. Novel binding partners of Ldb1 are required for haematopoietic development. Development. 133 (24), 4913-4923 (2006).
- Kettleborough, R. N., et al. A systematic genome-wide analysis of zebrafish protein-coding gene function. Nature. 496 (7446), 494-497 (2013).
- Leung, A., et al. Uncoupling VEGFA functions in arteriogenesis and hematopoietic stem cell specification. Developmental Cell. 24 (2), 144-158 (2013).
Перепечатки и разрешения
Запросить разрешение на использование текста или рисунков этого JoVE статьи
Запросить разрешениеСмотреть дополнительные статьи
This article has been published
Video Coming Soon
Авторские права © 2025 MyJoVE Corporation. Все права защищены