
Method Article
Многомасштабный анализ роста бактерий при стрессовом лечении
В этой статье
Резюме
Этот протокол позволяет улавливаться описанием роста бактерий в стрессовых условиях на одноклеточных и клеточных уровнях населения.
Аннотация
Анализ бактериальной способности расти и выживать в стрессовых условиях имеет важное значение для широкого спектра микробиологических исследований. Уместно охарактеризовать реакцию бактериальных клеток на стрессообразующие методы лечения, такие как воздействие антибиотиков или других противомикробных соединений, облучение, нефизиологический рН, температура или концентрация соли. Различные методы лечения стресса могут нарушить различные клеточные процессы, включая деление клеток, репликацию ДНК, синтез белка, целостность мембраны или регулирование клеточного цикла. Эти эффекты, как правило, связаны с конкретными фенотипами в клеточном масштабе. Поэтому понимание масштабов и причинно-следственной связи вызванных стрессом недостатков роста или жизнеспособности требует тщательного анализа нескольких параметров, как на одноклеточном, так и на уровне населения. Экспериментальная стратегия, представленная здесь, сочетает в себе традиционные оптические мониторинга плотности и анализы покрытия с одноклеточными методами анализа, такими как цитометрия потока и микроскопическая визуализация в реальном времени в живых клетках. Эта многомасштабная структура позволяет своевременно описывать влияние стрессовых условий на судьбу бактериальной популяции.
Введение
Общая цель этого протокола заключается в анализе поведения бактериальных клеток, подверженных стрессовому лечению в популяции и на одноклеточном уровне. Бактериальный рост и жизнеспособность традиционно решаются на уровне популяции с помощью мониторинга оптической плотности (OD600nm),который является прокси синтеза бактериальной массы клеток, или путем покрытия анализы для определения концентрации жизнеспособных клеток в культуре (колонии формирования единицы на миллилитр, CFU / mL). При нормальных (без стресса) условиях выращивания, OD600nm и CFU/mL измерения строго коррелируют, потому что бактериальное удвоение время напрямую зависит от увеличения массы клеток1,2. Однако, эта корреляция часто нарушается в условиях, которые влияют на синтез клеточной массы3, клеточное деление4, или что вызвать лиза клетки. Простой пример обеспечивается стресс лечения, которые подавляют деление клеток, которые приводят к образованию нитевидных бактериальных клеток5,6. Пятковые клетки удлиняются обычно, потому что синтез клеточной массы не влияет, но они не в состоянии разделить на жизнеспособные клетки. Оптическая плотность культуры, следовательно, будет увеличиваться с течением времени с нормальной скоростью, но не концентрацией жизнеспособных клеток, определяемых анализами покрытий (CFU/mL). В этом случае, как и во многих других, оптическая плотность и измерения покрытия являются информативными, но не обеспечивают всестороннего понимания наблюдаемого стрессового эффекта. Эти ансамбль анализы должны быть объединены с одноклеточными методами анализа, чтобы позволить углубленной характеристики стресс-индуцированных недостатков роста.
Здесь описывается процедура, которая сочетает в себе четыре взаимодополняющих экспериментальных подхода: (1) традиционные анализы напольное покрытие и базовый мониторинг оптической плотности для мониторинга жизнеспособности клеток и синтеза клеточной массы, соответственно; (2) цитометрия потока для оценки размера клеток и параметров содержания ДНК на большом количестве клеток; (3) микроскопическая снимка для анализа морфологии клеток; и (4) замедленного одноклеточного изображения в микрофлюидных камерах для изучения временной динамики судьбы клеток. Эта многомасштабная структура позволяет интерпретировать глобальное воздействие на рост клеток и жизнеспособность в свете поведения отдельных клеток. Эта процедура может быть применена для расшифровки реакции различных видов бактерий практически на любой стресс, представляющий интерес, включая рост в особых условиях (т.е. среду роста, рН, температуру, концентрацию соли), или воздействие антибиотиков или других противомикробных соединений.
протокол
1. Клеточная культура, индукция напряжения и процедура отбора проб
ПРИМЕЧАНИЕ: Используйте стерильную культурную стеклянную посуду, советы пипетки и средний рост, отфильтрованный на 0,22 мкм, чтобы избежать фоновых частиц. Здесь, культуры клетки выросли в низкой autofluorescence богатой определенной среде (см. Таблица материалов)7,8.
- Полоса бактериального штамма интереса от замороженных глицерола фонда на Лурия-Брот (LB) агарозной пластины (с селективным антибиотиком, если это необходимо) и инкубировать при 37 КС ночь (17 ч).
ПРИМЕЧАНИЕ: Пример эксперимента, представленный здесь использует Escherichia coli MG1655 hupA-mCherry. Этот штамм производит флуоресцентно помечены субединицу no ИЗ HU нуклеоидного связанного белка, что позволяет свет микроскопии визуализации хромосомы в живых клетках9. - Прививать 5 мл среды с одной колонией и расти при 37 градусов по Цельсию с встряхиванием при 140 вращения в минуту (об/ ч) на ночь (17 ч). Для обеспечения удовлетворительной аэрации взволнованной культуры необходимо использовать пробирки большого диаметра (2 см).
- На следующее утро измеряют оптическую плотность на уровне 600 нм (OD600nm)и разбавляют культуру в пробирку, содержащую свежую среду до OD600nm 0.01. Общий объем культуры должен быть скорректирован в зависимости от количества точек времени, которые будут проанализированы в ходе эксперимента.
- Загрузите образец культуры объемом 200 л (0,2 мл на хорошо работающий объем с прозрачным дном) и поместите его в автоматический считыватель пластин (см. Таблица материалов)для мониторинга OD600nm во время инкубации при 37 градусах Цельсия.
- Инкубировать привит пробирку при 37 градусах Цельсия с тряской (140 об/мин) до ОД600нм и 0,2, что соответствует полной экспоненциальной фазе в богатой среде.
ПРИМЕЧАНИЕ: Очень важно расти клетки, по крайней мере 4-5 поколений, прежде чем достичь надлежащего экспоненциального роста. Первоначальный инокулум, используемый в шаге 1.3 (OD600nm и 0.01) должен быть адаптирован в случае роста в более бедной среде (т.е. когда экспоненциальная фаза достигается ниже ОД 0,2), или если требуется больше поколений (например, для конкретных физиологических исследований или расширенного лечения стресса). - На OD600nm и 0.2, возьмите следующие образцы культуры, соответствующие точке времени t0 (экспоненциально растущие клетки до индукции напряжения): (1) образец 150 л, который будет немедленно загружен в микрофлюидный аппарат для визуализации микроскопии замедленного времени (см. раздел 2); (2) Образец 200 л для проверки разбавления и покрытия (см. раздел 3); (3) образец 250 л л, который следует поставить на лед для анализа цитометрии потока (раздел 4); (4) Образец 10 л, который должен быть немедленно отложен на агарозе установлен слайд для микроскопии снимок изображения (см. раздел 5).
- Разоблачить клеточной культуры, оставшихся в пробирке для конкретного лечения стресса вы хотите исследовать и инкубировать при 37 градусов по Цельсию с встряхивания (140 об/ ч. ).
ПРИМЕЧАНИЕ: Культура, растущая в автоматизированном считывателе пластин для мониторинга OD600nm, также должна подвергаться стрессовому лечению. - В соответствующие временные моменты после лечения стресса (т1,т2,т3и т.д.), возьмите следующие образцы клеток из подчеркнутой культуры: (1) Образец 200 Л для разбавления и прокладки (см. раздел 2); (2) образец 250 л л для анализа цитометрии потока (раздел 3); (4) Образец 10 л, который должен быть немедленно отложен на агарозе установлен слайд для микроскопии снимок изображения (см. раздел 4).
ПРИМЕЧАНИЕ: Каждое стресс-индуцирующее лечение имеет эффективность, которая зависит от дозы и времени. Таким образом, может потребоваться провести предварительные тесты, чтобы определить дозу и продолжительность лечения, которые будут использоваться для оптимальных результатов. Это может быть сделано путем выполнения мониторинга OD с помощью автоматизированного считывателя пластин (потенциально связанных с покрытием анализы) клеточной культуры, обработанной с диапазоном доз и время экспозиции. В эксперименте, представленном здесь, клеточная культура была обработана с клеточного деления ингибирующих антибиотикцексин (Цеф.) на 5 мкг /мл конечной концентрации в течение 60 мин. Cephalexin затем смыло путем гралета клеток в 15 мл трубки с помощью центрифугирования (475 г, 5 мин), удаление супернатанта, resuspending клеточной гранулы в равном объеме свежей трубки. Промытые клетки были инкубированы при 37 градусах Цельсия с тряской (140 об/мин), чтобы обеспечить восстановление. Образец был взят на т60 (60 мин после цефалексина дополнение, соответствующее "цефалексин-60мин-обработанный" образец), т120 (60 мин после мытья), и т180 (120 мин после мытья).
2. Плата асссе
ПРИМЕЧАНИЕ: Тестирование покрытия позволяет измерять концентрацию клеток, способных генерировать CFU в образцах культуры. Эта процедура показывает скорость, с которой одна клетка делится на две жизнеспособные клетки и позволяет обнаружить обнаружение арестов деления клеток (например, увеличение времени генерации бактериального поколения клеточного лиза).
- Приготовьте 10-кратное серийное разбавление до10-7 из 200 л образца культуры в свежей среде. Плита 100 л соответствующего разбавления на неселективных пластинах агарозы LB для получения между 3-300 колониями после ночной инкубации при 37 градусах Цельсия.
ПРИМЕЧАНИЕ: Серийное разбавление в свежей среде должно быть выполнено быстро, чтобы ограничить бактериальные деления. Кроме того, исследователи могли бы рассмотреть вопрос об использовании соленового раствора без источника углерода для предотвращения деления клеток в процессе разбавления. - На следующий день подсчитайте количество колоний, чтобы определить концентрацию жизнеспособных клеток (CFU/mL) в каждой образце культуры. Участок CFU/mL как функция времени для необработанных и обработанных культур клетки.
3. Цитометрия потока
ПРИМЕЧАНИЕ: В следующем разделе описывается подготовка образцов клеток для анализа цитометрии потока. Этот метод анализа показывает распределение размера клеток и содержания ДНК для большого количества клеток. По возможности рекомендуется немедленно обработать пробы цитометрии потока. Кроме того, образцы могут храниться на льду (до 6 ч) и анализироваться одновременно в конце дня, после того, как были выполнены промывоц и микроскопическая визуализация.
- Разбавить 250 л культурных образцов для получения 250 qL при концентрации 15 000 клеток/Л (соответствующей ОД600нм 0,06) в свежей среде при 4 градусах Цельсия.
ПРИМЕЧАНИЕ: Инкубация на льду ограничит рост и морфологическую модификацию клеток. Кроме того, пользователь может рассмотреть возможность выполнения фиксации клеток в 75% этанола, как это рекомендуется для потока цитометрии10. - Для окрашивания ДНК смешайте бактериальный образец с раствором 10 мкг/мл флуоресцентного красителя ДНК (коэффициент 1:1) и инкубировать в темноте в течение 15 мин, прежде чем анализировать образец.
- Передайте образец в цитометр потока с частотой потока в 120 000 ячеек/мин. Приобретайте вперед-рассеянный (FSC) и side-scattered (SSC) свет также, как сигнал флуоресценции красителя динары (FL-1) с соотвествующие установками.
- Участок FSC и FL-1 плотность клеток гистограммы представлять распределение размера клеток и содержания ДНК в популяции клеток.
4. Снимок микроскопии изображения
ПРИМЕЧАНИЕ: Следующая часть описывает подготовку слайдов микроскопии и приобретение изображений для анализа снимков населения. Эта процедура предоставит информацию о морфологии клеток (длина клеток, ширина, форма) и внутриклеточной организации нуклеоидной ДНК.
- Разогреть термодепональную камеру микроскопа при температуре 37 градусов по Цельсию, чтобы стабилизировать температуру перед началом наблюдений. Эта камера позволяет модуляцию температуры микроскопической оптики и этап уобразцов во время экспериментов замедленного действия.
- Подготовьте агарозные слайды, как описано в Лестерлине и Дюбарри7.
- Удалите пластиковую пленку из нижней части синей рамы (см. Таблица материалов),оставляя выдолбленную пластиковую пленку на другой стороне. Придерживайтесь синей рамы на слайде из стекла микроскопа.
- Пипетка 150 л расплавленного 1% агарозного среднего раствора и залить в синем каркасном отсеке. Быстро накройте крышкой, чтобы удалить лишнюю жидкость и подождать несколько минут для агарозной площадки, чтобы затвердеть при комнатной температуре.
- Когда образец ячейки будет готов, снимите крышку и пластиковую пленку из синей рамы. Налейте 10 л образца клеток на агарозную площадку и осторожно наклоните стеклянную горку, чтобы распространить каплю. Когда вся жидкость была адсорбирована, наденьте чистый ковер на синюю рамку, чтобы запечатать образец. Микроскопия слайд теперь готов к микроскопии.
- Поместите слайд на стадии микроскопа и выполните приобретение изображения с использованием передаваемого света (с целью фазового контраста) и с возбуждением источника света на соответствующих длинах волн (560 нм для mCherry).
- Выберите поля зрения, содержащие изолированные ячейки, чтобы облегчить автоматическое обнаружение клеток во время анализа изображений. Убедитесь, что по крайней мере 300 клеток имеют изображение, чтобы обеспечить надежный статистический анализ популяции клеток.
5. Микрофлюидика замедленного микроскопического изображения
ПРИМЕЧАНИЕ: Следующая часть объясняет подготовку микрофлюидных пластин (см. Таблица Материалов), загрузка клеток, программа микрофлюики и замедленное приобретение изображений. Эта процедура визуализации показывает поведение отдельных клеток в режиме реального времени.
- Удалите консервативную раствор из микрофлюидной пластины и замените его свежей средней разогретой до 37 градусов по Цельсию, как описано в руководстве пользователя микрофлюидного программного обеспечения.
- Печать микрофлюидной пластины к многообразной системе и нажмите на кнопку Priming.
- Поместите микрофлюидную пластину с многообразной системой на стадии микроскопа и разогревайте при температуре 37 градусов по Цельсию в течение 2 ч, прежде чем начать приобретение микроскопии.
ПРИМЕЧАНИЕ: Этот шаг по разогрева имеет решающее значение, чтобы избежать расширения микрофлюидной камеры, которая изменила бы фокусировку микроскопа во время эксперимента замедленного действия и компрометирующего приобретения изображения. - Уплотнение микрофлюидной пластины. Замените среду из скважины 8 на 150 л образца культуры и замените среду от 1 до 5 желаемой средой с реагентом или без нее.
- Запечатайте микрофлюидную пластину и поместите ее на сцену микроскопа.
- На микрофлюидном программном обеспечении (см. Таблица Материалов)запустите процедуру загрузки клеток. Убедитесь, что загрузка клеток является удовлетворительной, глядя под микроскопом в передаваемом свете. Запустите процедуру загрузки ячейки во второй раз, если плотность клеток в камере недостаточна.
- Выполняйте тщательно фокусироваться в режиме передаваемого света и выберите несколько полей зрения, которые показывают изолированные бактерии. Важно выбрать поля, которые не переполнены, чтобы иметь возможность контролировать рост изолированных клеток с течением времени (рекомендуется 10-20 ячеек на 100 мкм2). Это также облегчит обнаружение клеток во время анализа изображений.
- На микрофлюидном программном обеспечении нажмите на кнопку «Создать протокол». Программа инъекций среды роста для 1'2 эквивалентов времени поколения, чтобы позволить клеткам адаптироваться (по желанию). Затем запрограммируйте инъекцию стрессообразующей среды в течение 10 мин при 2 пси, а затем инъекцию в 1 пси для требуемой продолжительности стрессового лечения. Если вы собираетесь проанализировать восстановление клеток после стресса, запрограммируйте инъекцию свежей среды роста на требуемую продолжительность.
ПРИМЕЧАНИЕ: В эксперименте, представленном здесь, цефалексин был введен в течение 10 мин при 2 пси, а затем 50 мин на 1 пси. Затем, свежий рост среды был введен в 2 пси в течение 10 минут, а затем 3 ч в 1 пси. - Выполняйте микроскопические изображения в режиме замедленного выполнения с 1 кадром каждые 10 минут, используя фазовый контраст в передаваемом свете и 560 нм источник света возбуждения для сигнала mCherry, если это необходимо.
ПРИМЕЧАНИЕ: Важно начать микроскопическое приобретение изображения одновременно с началом протокола микрофлюидных инъекций.
6. Анализ изображений
ПРИМЕЧАНИЕ: В этом разделе кратко описаны ключевые этапы обработки и анализа снимков и замедленной микроскопии. Открытие и визуализация изображений микроскопии осуществляется с открытым исходным кодом ImageJ/Fiji (https://fiji.sc/)11. Количественный анализ изображений проводится с использованием программного обеспечения ImageJ/Fiji с открытым исходным кодом вместе с бесплатным плагином MicrobeJ12 (https://microbej.com). Этот протокол использует версию MicrobeJ 5.13I.
- Откройте программное обеспечение Фиджи и плагин MicrobeJ.
- Для анализа моментальных снимков отбросьте все изображения, соответствующие одному слайду микроскопа (один образец) в панель загрузки MicrobeJ, чтобы совпьировать изображения и сохранить полученный файл стеков изображений. Для данных замедленного времени просто перебросьте стек изображения в панель загрузки MicrobeJ.
- Запустите автоматическое обнаружение контуров клеток на основе сегментации фазового контрастного изображения и, если это уместно, ядер на основе сегментации окрашенного сигнала флуоресценции ДНК. Проверьте точность обнаружения ячейки визуально и использовать инструмент редактирования MicrobeJ для коррекции, если это необходимо. Сохранить полученный файл результата.
ПРИМЕЧАНИЕ: Параметры, используемые для обнаружения клеток кишечной палочки, указаны в таблице материалов (см. Колонку Комментариев/Описание MicrobeJ). Для других видов бактерий (особенно для бактерий, не входящих в форму), пользователь должен уточнить настройки перед обнаружением (см. учебник MicrobeJ). Для замедленного изображения, запуск полуавтоматического обнаружения клеток с помощью инструмента редактирования MicrobeJ может быть предпочтительным, чтобы позволить сосредоточиться на судьбе отдельных клеток (см. учебник MicrobeJ). - Нажмите на значок ResultJ, чтобы завершить анализ и получить окно ResultJ. С этой точки можно сгенерировать множество различных типов выходных графиков. Участок нормализованных гистограмм клеточной формы / длины и среднего числа нуклеоидов на клетку.
Результаты
Описанная процедура была использована для анализа поведения клеток Escherichia coli K12 во время преходящего воздействия цефалексина, антибиотика, который специально подавляет деление клеток(рисунок 1А)13. HupA-mCherry E. coli штамм, который производит флуоресцентно помечены HU белка, связанного с хромосомной ДНК был использован для исследования динамики хромосомы на протяжении всего этого лечения8,9. Экспоненциально растущие клетки hupA-mCherry E. coli были проанализированы ранее (т0)и 60 мин после инкубации с цефалексином (т60). Затем антибиотик смыло и было проанализировано восстановление популяции клеток после 1 ч(т 120)и 2 ч (т180)(рисунок 1В).

Рисунок 1: Процедура анализа бактериальной реакции на стрессовое лечение. (A) Схема метода. (B) Мультфильм, иллюстрирующие морфологию клеток во время нормального роста в богатой среде и во время преходящего воздействия цефалексина (Цеф.), от добавления на (т0) и после стирки цефалэксина от (t60) до (t180). Пожалуйста, нажмите здесь, чтобы просмотреть большую версию этой цифры.
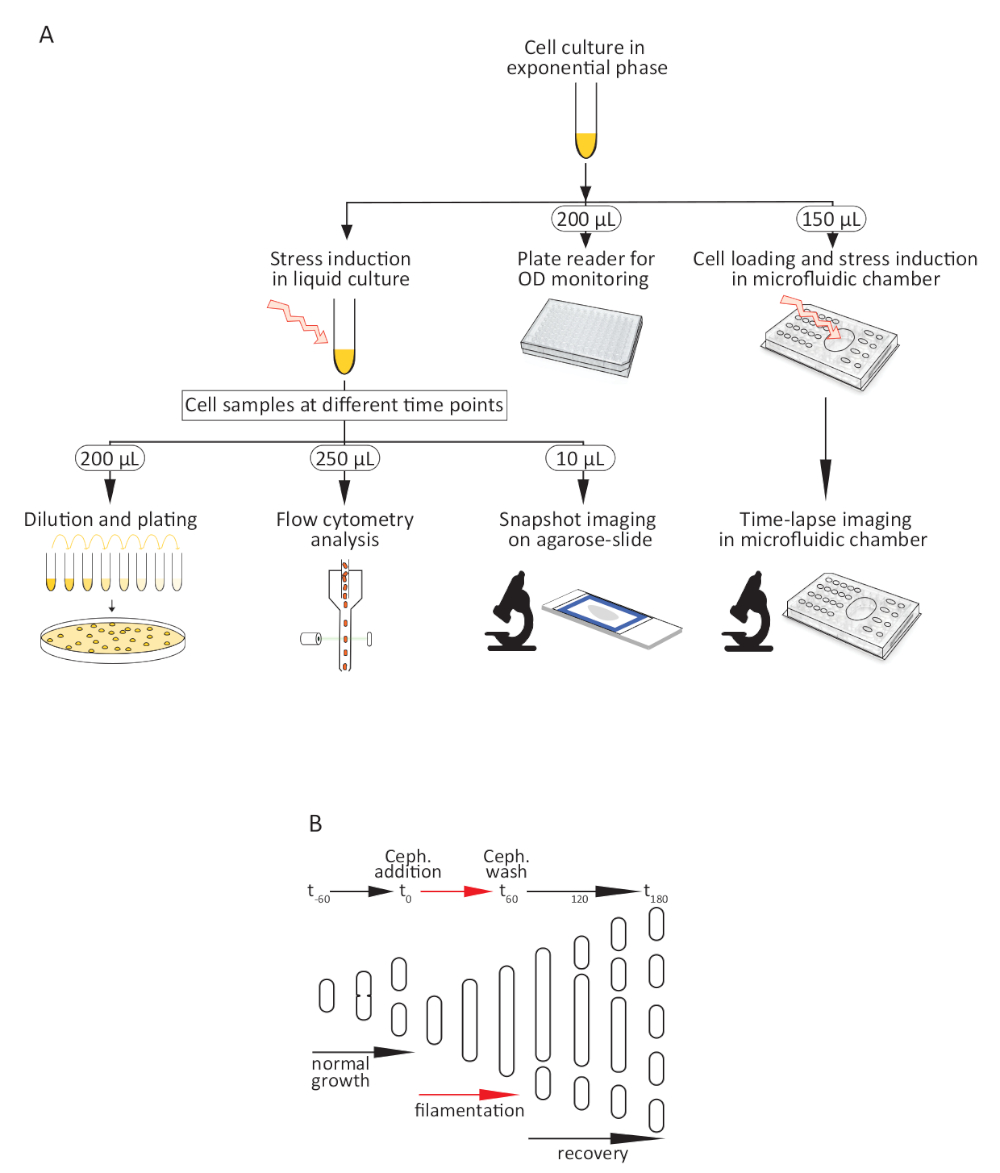{kind=link}
Параллельная эволюция OD и CFU/mL является первым индикатором, который помогает понять эффект лечения стресса. Эти два параметра строго коррелируются во время невозмутимого роста, но часто разъединяются и развиваются независимо под стрессом. Клеточные культуры, растущие в присутствии цефалексина выставлены аналогичные OD600nm увеличивается, как без стресса культур(Рисунок 2A), показывая, что препарат не влияет на синтез клеточной массы. Однако концентрация жизнеспособных клеток не увеличивалась при цефалексине из-за строгого ингибирования деления клеток(рисунок 2В). Клетки начали делитьснова когда cephalexin извлекли и окончательн достигли концентрации эквивалентную к unstressed культуре на (t180). Эти результаты отражают бактериостатический эффект цефалеста, который вызывает полностью обратимое ингибирование деления клеток. Различные напряжения приведут к различному разъединению кривых OD и CFU/mL, в зависимости от индуцированного эффекта (например, модификации морфологии клеток, такой как нити или выпуклые, гибель клеток с или без лисиса). Неполный список возможных результатов, свидетельствуют о различных стресс-индуцированных эффектов представлен на рисунке 2C.

Рисунок 2: Мониторинг роста бактерий необработанных и цефалэксинобработанных клеток на уровне популяции. (A) Мониторинг оптической плотности (OD600nm/mL). (B) Концентрация жизнеспособных клеток (CFU/mL) в необработанных и цефалексин-60мин обработанных культур. Бары ошибок указывают на стандартное отклонение для экспериментального тройного. (C) Схема возможных результатов и связанных с ними стрессовых эффектов. Пожалуйста, нажмите здесь, чтобы просмотреть большую версию этой цифры.
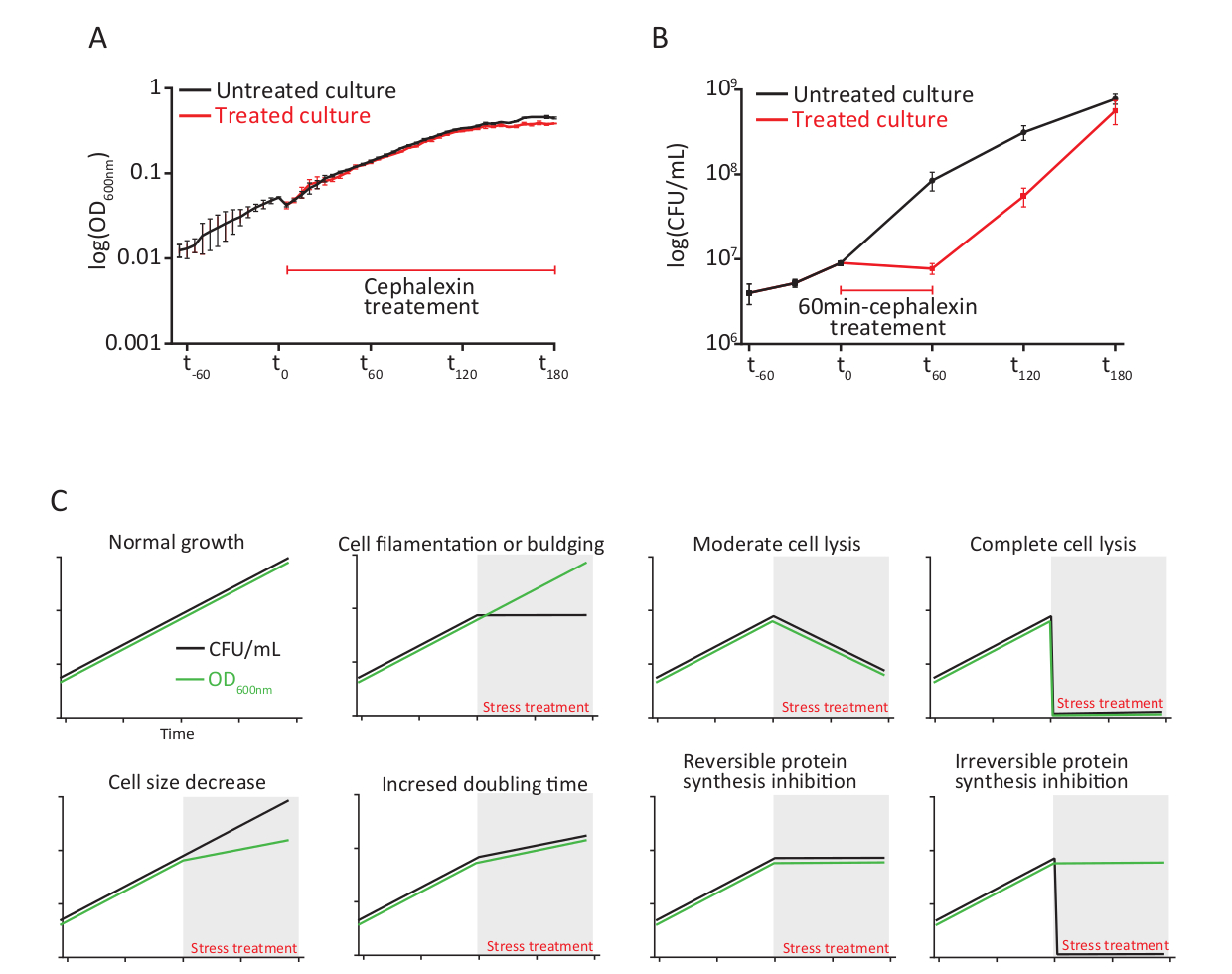{kind=link}
Одноклеточный анализ необходим для точной интерпретации реакции на стресс, наблюдаемой на уровне населения. Цитометрия потока позволяет изучить размер клетки и содержание ДНК нескольких тысяч клеток14,15 (рисунок 3). Воздействие цефалексина спровоцировало параллельное увеличение размера клеток и содержания ДНК (т60). Когда цефалексин был удален, размер клеток популяции и содержание ДНК постепенно уменьшались, чтобы стать похожими на ненапряженную популяцию наt 180. Эти результаты показывают, что цефалексин не препятствовал репликации ДНК и спровоцировал образование нитевидных клеток, содержащих несколько хромосомных эквивалентов. Эти нити делятся на клетки с нормальным размером клеток и содержанием ДНК, когда препарат был смыт. Результаты цитометрии потока будут очень разными для стрессов, которые подавляют синтез ДНК, которые приводят к образованию нитевидных клеток, содержащих только одну нереплицирующую хромосому. В этом случае размер клеток будет увеличиваться аналогичным образом, но не будет связан с увеличением содержания ДНК.

Рисунок 3: Представитель ный анализ цитометрии потока необработанных и цефалэксин-60мин обработанных клеток. (A) Гистограммы распределения размеров клеток (FSC-H). (B) Гистограммы содержания ДНК (FL1-SYTO9). n - проанализировано 120 000 клеток. Пожалуйста, нажмите здесь, чтобы просмотреть большую версию этой цифры.
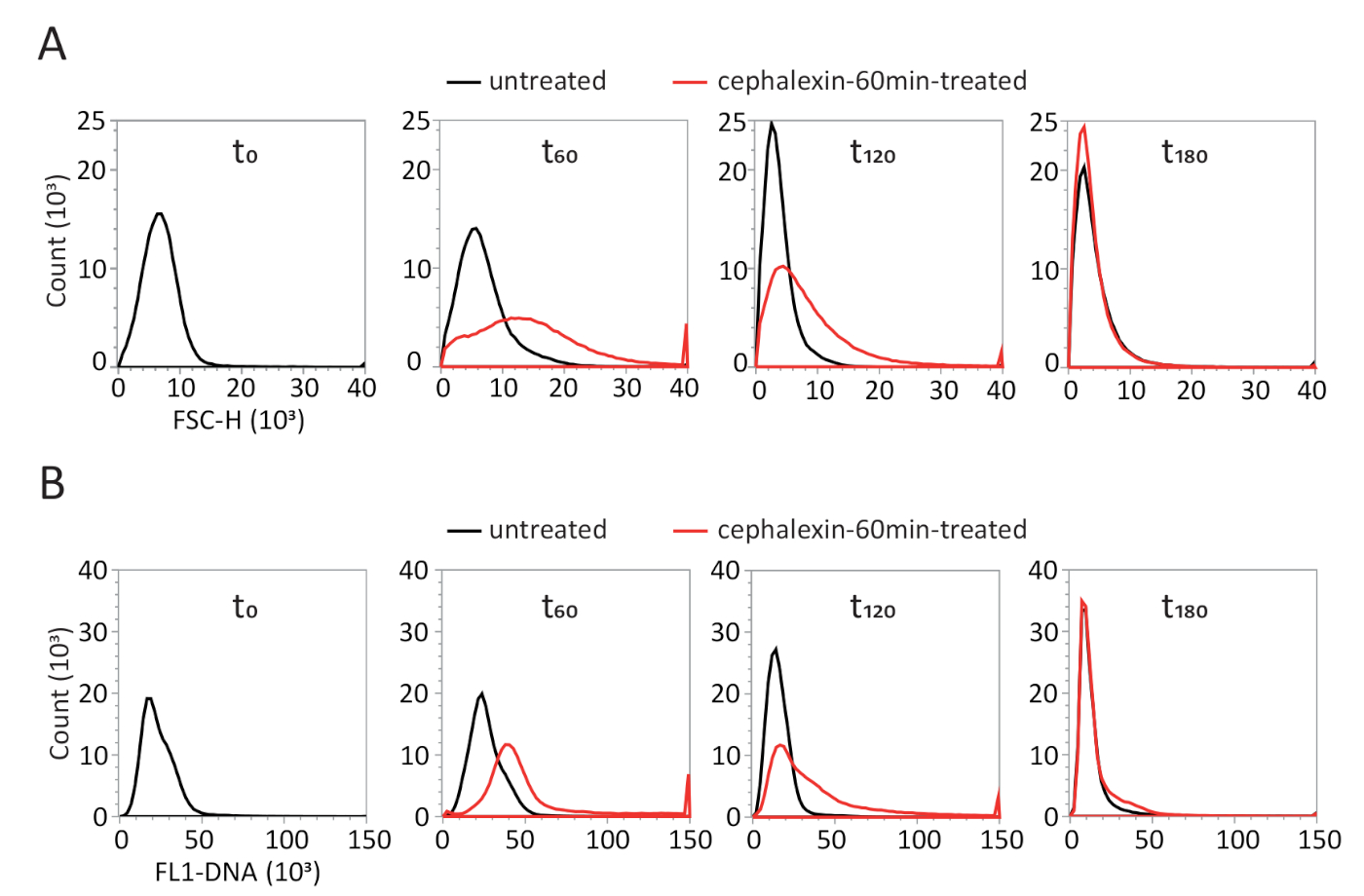{kind=link}
Снимок микроскопии изображения был использован для изучения морфологии клеток и внутриклеточной организации ДНК показано HU-mCherry локализации (Рисунок 4A). Цефалексин спровоцировал образование длинных клеток с нормальной шириной клеток и отсутствием делениессты. Эти гладкие нити содержали регулярно расположенные структуры ДНК, называемые нуклеоидами, подтверждая, что цефалексин не влияет на репликацию и сегрегацию хромосом. Количественный анализ изображений в значительной степени подтвердил размер клетки и увеличение содержания ДНК, ранее наблюдаемое с цитометрией потока(рисунок 4B,C). Результаты будут очень разными для стрессов, которые вызывают повреждение ДНК, которые приводят к образованию нитевидных клеток, в которых репликация продолжается, но сегрегация нарушается. В этом случае размер клеток и содержание ДНК будут увеличиваться аналогичным образом, но клетки будут питать единую неструктурированную массу ДНК. Снимки могут также выявить другие виды аномальных форм клеток или наличие мини, ануклиата или лисердных клеток (клеток-призраков).

Рисунок 4: Микроскопия снимок анализа необработанных и цефалексин-60min обработанных клеток. (A) Представитель микроскопии изображения, показывающие фазовый контраст (серый) и HU-mCherry сигнал (красный). (B) Гистограммы распределения длины клетки. Шкала бар No 5 мкм. (C) Гистограммы числа нуклеоидов на клетку. Для каждого образца было проанализировано от 800 до 2000 клеток. Пожалуйста, нажмите здесь, чтобы просмотреть большую версию этой цифры.
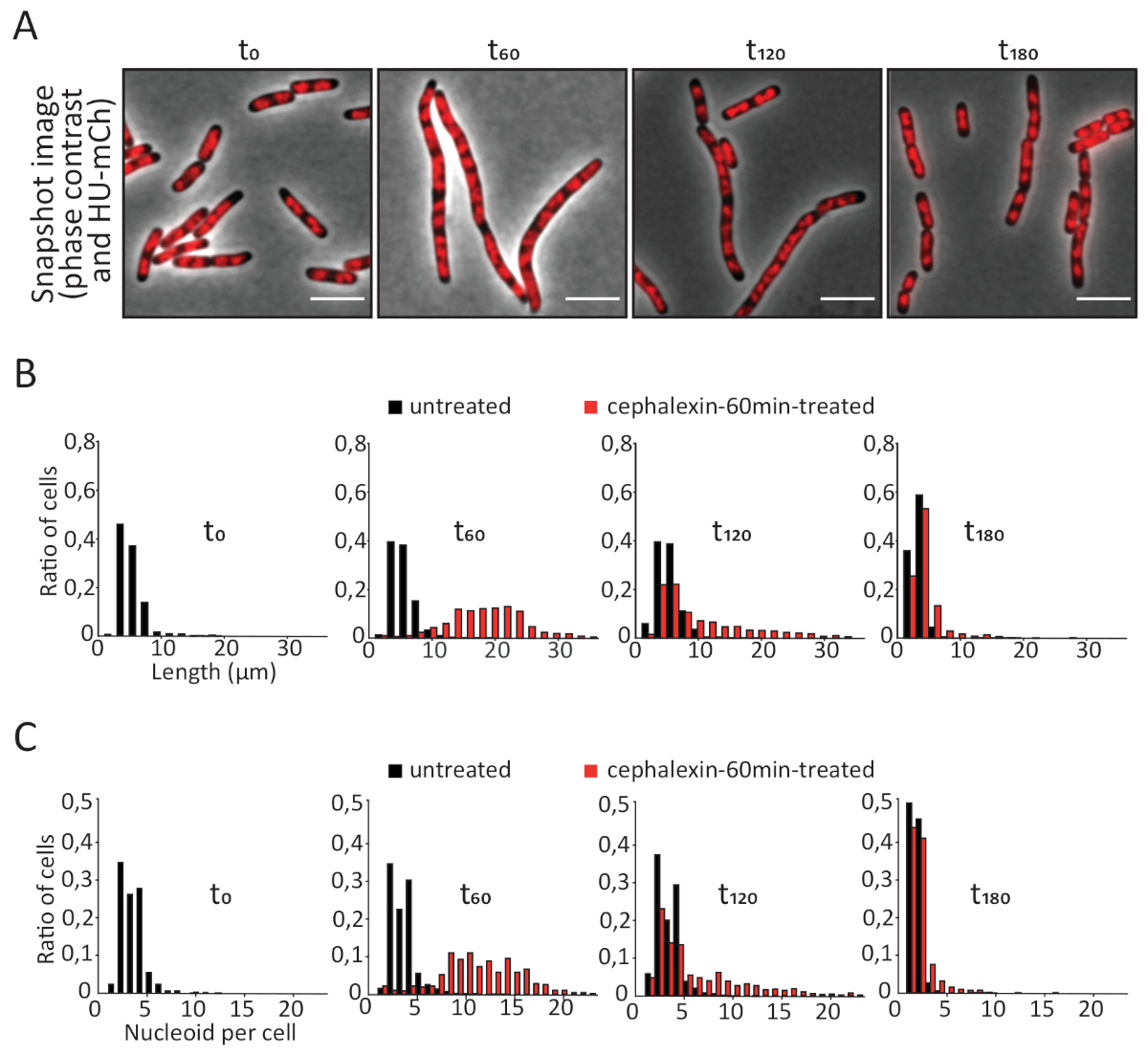{kind=link}
Микроскопия, связанная с микрофлюидным аппаратом16, помогла определить ранее наблюдаемые фенотипы и дает дополнительные сведения о развитии и причинно-следственной связи дефицита роста. Time-lapse изображения(Рисунок 5A и фильм 1) подтвердили, что удлинение клеток (синтез клеточной массы), и хромосома репликации и сегрегации не были ингибированы воздействием цефалексина. Кроме того, он показал процесс восстановления, когда цефалексин был удален. Анализ нитевидной линии клеток показал, что деление клеток перезапускается на 20 мин после смывания препарата(рисунок 5В). В результате разделенные клетки были жизнеспособными, потому что они, в свою очередь, разделились, что в конечном итоге привело к образованию 33 клеток, демонстрирующих нормальный размер и содержание ДНК. Это позволило просчитать общее время генерации в размере 31 мин в течение 180 мин эксперимента, что аналогично времени генерации, рассчитанной для неподчеркнутой популяции из измерений CFU/mL (33 мин).

Рисунок 5: Микроскопия покадрового анализа цефалэксин-60мин обработанных клеток. (A) Представитель микроскопии изображения, показывающие фазовый контраст (серый) и HU-mCherry сигнал (красный). Контролируемой нитевидной ячейки указывается белый контур, и разделенные клетки разными цветами. Шкала бар No 5 мкм. (B) Схематическое представление нитевидной линии клеток, соответствующих панели (A) и фильм 1. Пожалуйста, нажмите здесь, чтобы просмотреть большую версию этой цифры.
{kind=link}

Фильм 1: Микрофлюидический фильм E. coli HU-mCherry лечится цефалексином. Cephalexin был введен после 60 мин, а затем инъекции свежей среды RDM в течение 3 ч. Время, указанное желтым (1 кадр каждые 10 мин). Шкала бар No 5 мкм. Пожалуйста, нажмите здесь, чтобы посмотреть это видео (право нажмите, чтобы скачать).
Обсуждение
Необходимо обратить внимание на состояние роста клеток во время процедуры. Расти клетки в течение нескольких поколений, прежде чем достичь полной экспоненциальной фазы. Для успеха этого метода важно, чтобы все образцы клеток собирались одновременно, и лучше всего анализировать только одну обработанную и одну необработанную культуру одновременно. Образцы клеток для микроскопии должны поддерживаться при экспериментальной температуре на протяжении всей процедуры. Затем необходимо разогреть камеру микроскопа и микрофлюидную камеру до начала эксперимента. Если образцы клеток для цитометрии потока не могут быть проанализированы легко, они могут храниться на льду до 6 ч. Инкубация на льду ограничит рост и морфологическую модификацию клеток. Кроме того, пользователь может рассмотреть вопрос об использовании фиксации клеток в этаноле 75%, который обычно рекомендуется для потока цитометрии10. Если протокол требует мытья стресс индуктора из среды, центрифуги и пипетки клетки очень тщательно, чтобы избежать повреждения потенциальных аномальных клеток.
Как цитометрия потока, так и анализ моментальных снимков дают доступ к размеру клеток и параметрам содержания ДНК, при этом снимки обеспечивают дополнительное наблюдение за морфологией клеток. Окрашивание ДНК DAPI10 (4',6-diamidino-2-phenylindole) или другими стабильными красителями ДНК может быть выполнено, если не доступно флуоресцентное слияние для наблюдения ядер в интересуемом организме организма. Если анализ цитометрии потока не может быть выполнен, важно, чтобы изображение и анализ большого количества клеток с помощью микроскопии.
Микроскопическая визуализация также может быть выполнена с использованием штаммов, несущих флуоресцентные слияния белков, участвующих в конкретных путей интереса. Это помогло бы выявить влияние стресса на различные клеточные процессы, такие как репликация, транскрипция, синтез клеточной стенки, или деление клеток. Метод может быть применен к ряду бактериальных видов, единственным требованием является то, что микрофлюидный аппарат должен быть совместим с морфологией клеток. Стандартные микрофлюидные пластины удобны для бактерий стержня с шириной клеток между 0,7 мкм и 4,5 мкм. Тем не менее, кокки, ovococci, или другие бактериальные штаммы с своеобразными формами должны быть проверены. Кроме того, если микрофлюидные эксперименты не могут быть выполнены из-за отсутствия оборудования или несовместимых бактериальных штаммов, замедленное изображение может быть выполнено на агарозных слайдах в течение максимальной продолжительности 2ч.
Общее преимущество этого многомасштабного анализа заключается в том, чтобы обеспечить глобальное видение влияния индукции стресса на несколько аспектов способности роста бактерий (т.е. синтез массы, жизнеспособность клеток, морфология клеток, целостность мембраны, содержание ДНК) и способ они развиваются со временем в бактериальной популяции растет в стрессовых условиях. Это также позволяет анализировать восстановление нормального роста на одноклеточном уровне и уровне популяции. Этот подход применим к широкому кругу видов бактерий и практически к любому виду лечения стресса, например, к воздействию антибиотиков или других противомикробных соединений, анализу влияния взаимодействия с другими организмами на многовидовые населения, или эффект генетической мутации.
Раскрытие информации
Авторы заявили об отсутствии конкурирующих интересов.
Благодарности
Авторы благодарят Ф. Корнеза за предоставление штамма hupA-mCherry, А. Дедье за техническую помощь с цитометрией и А. Дюкре за помощь в MicrobeJ. Финансирование: C. Lesterlin признает Inserm и CNRS учреждений, а также aTIP-Avenir программы, Schlumberger Фонд образования и исследований (FSER 2019), ANR финансирования программы PlasMed исследований (ANR-18-CE35-0008), а также FINOVI для финансирования J. Cayron; La Ligue contre le cancer для финансирования цитометрического оборудования потока. Вклад автора: C.L. и J.C. конструировали процедуру и написали бумагу; J.C. провел эксперименты и проанализировал данные.
Материалы
| Name | Company | Catalog Number | Comments |
| Agarose | BioRad | 1613100 | Certified molecular biology agarose |
| Attune NxT Acoustic Focusing Cytometer | ThermoFisher scientific | A24858 | Cytometer |
| CellASIC ONIX Microfluidic System | Merck Millipore | CAX2-S0000 | Microfluidic system |
| CellASIC ONIX2 FG | Merck Millipore | ONIX2 1.0.1 | Microfluidic software |
| CellASIC ONIX2 Manifold Basic | Merck Millipore | CAX2-MBC20 | Manifold system |
| CytoOne 96-well plate with lid | Starlab | CC7672-7596 | Microplate with 0,2 mL well working volume and clear flat bottom, for automated plate reader |
| E. coli strain carrying a chromosomal insertion for a hupA-mCherry fusion | Created by P1 transduction of hupA-mCherry in E. coli MG1655 | ||
| Fiji | ImageJ | https://fiji.sc/ | Image software. Cite Schindelin et al. if used in publication |
| Gene Frame | Thermo Scientific | AB-0578 | Blue frame (125 μL, 1,7 x 2,8 cm) |
| Luria-Broth agarose medium | MP Biomedicals | 3002232 | Growth medium for plating assay |
| MicrobeJ | Imagej/Fiji plugin | https://www.microbej.com/ | Microscopy image analysis plugin. Cite Ducret et al. If used in publication; Detection settings: For bacteria : Area (μm2): 0,1-max; Length (μm): 0,5-max; Width (μm): 0,6-max; Range (μm): 0,5-max; Angularity (rad): 0-0,3; 0-max for all other parameters. For nucleoid: Tolerance: 500; Threshold: Local; 0-max for all other parameters |
| Microfluidic Plates CellASIC ONIX | Merck Millipore | B04A-03-5PK | Plate for Microfluidic system |
| Microscope Nikon eclipse Ti | Nikon | Fluorescence microscope | |
| MOPS EZ Rich Defined Medium (RDM) | Teknova | M2105 | Growth rich medium, 10x MOPS Mixture, 0,132 M K2HPO4, 10x AGCU, 5x Supplement EZ, 20% Glucose. Filtered at 0,22 μm |
| SYTO9 Green Fluorescent Nucleic Acid Stain | ThermoFisher scientific | S34854 | DNA fluorescent dye |
| TECAN Infinite M1000 | TECAN | 30034301 | Automated plate reader |
Ссылки
- Donachie, W. D., Begg, K. J., Vicente, M. Cell length, cell growth and cell division. Nature. 264 (5584), 328-333 (1976).
- Donachie, W. D., Blakely, G. W. Coupling the initiation of chromosome replication to cell size in Escherichia coli. Current Opinion in Microbiology. 6 (2), 146-150 (2003).
- Patterson, D., Gillespie, D. Effect of Elevated Temperatures on Protein Synthesis in Escherichia coli. Journal of Bacteriology. 112 (3), 1177(1972).
- Begg, K. J., Donachie, W. D. Cell shape and division in Escherichia coli: experiments with shape and division mutants. Journal of Bacteriology. 163 (2), 615-622 (1985).
- Van De Putte, P., Van Dillewijn, J., Roersch, A. The Selection of Mutants of Escherichia Coli with Impaired Cell Division at Elevated Temperature. Mutation Research. 106, 121-128 (1964).
- Walker, A. R. Initial characterization of temperature-sensitive cell division mutants of Escherichia coli. Biochemical and Biophysical Research Communications. 47 (5), 1074-1079 (1972).
- Lesterlin, C., Duabrry, N. Investigating Bacterial Chromosome Architecture. Chromosome Architecture. 1431, http://link.springer.com/10.1007/978-1-4939-3631-1_6 61-72 (2016).
- Stracy, M., et al. Live-cell superresolution microscopy reveals the organization of RNA polymerase in the bacterial nucleoid. Proceedings of the National Academy of Sciences. 112 (32), E4390-E4399 (2015).
- Fisher, J. K., et al. Four-dimensional imaging of E. coli nucleoid organization and dynamics in living cells. Cell. 153 (4), 882-895 (2013).
- Lesterlin, C., Pages, C., Dubarry, N., Dasgupta, S., Cornet, F. Asymmetry of chromosome Replichores renders the DNA translocase activity of FtsK essential for cell division and cell shape maintenance in Escherichia coli. PLoS genetics. 4 (12), e1000288(2008).
- Schindelin, J., et al. Fiji - an Open Source platform for biological image analysis. Nature Methods. 9 (7), (2019).
- Ducret, A., Quardokus, E. M., Brun, Y. V. MicrobeJ, a tool for high throughput bacterial cell detection and quantitative analysis. Nature Microbiology. 1 (7), 16077(2016).
- Rolinson, G. N. Effect of beta-lactam antibiotics on bacterial cell growth rate. Journal of General Microbiology. 120 (2), 317-323 (1980).
- Boye, E., Løbner-Olesen, A. Flow cytometry: illuminating microbiology. The New Biologist. 2 (2), 119-125 (1990).
- Rieseberg, M., Kasper, C., Reardon, K. F., Scheper, T. Flow cytometry in biotechnology. Applied Microbiology and Biotechnology. 56 (3-4), 350-360 (2001).
- Nolivos, S., et al. Role of AcrAB-TolC multidrug efflux pump in drug-resistance acquisition by plasmid transfer. Science. 364 (6442), 778-782 (2019).
- Chao, Y., Zhang, T. Optimization of fixation methods for observation of bacterial cell morphology and surface ultrastructures by atomic force microscopy. Applied Microbiology and Biotechnology. 92 (2), 381-392 (2011).
Перепечатки и разрешения
Запросить разрешение на использование текста или рисунков этого JoVE статьи
Запросить разрешениеСмотреть дополнительные статьи
This article has been published
Video Coming Soon
Авторские права © 2025 MyJoVE Corporation. Все права защищены