
Method Article
Стратегический скрининг и характеристика визуального GPCR-мини-G Белковый сигнальный комплекс для успешной кристаллизации
В этой статье
Резюме
В настоящем докладе описывается скрининг различных моющих средств для подготовки визуального GPCR, родопсин, и его комплекс с мини-Go. Демонстрируются биохимические методы, характеризующие качество комплекса на разных этапах очистки. Этот протокол можно обобщить к другим комплексам протеина мембраны для их будущих структурных изучений.
Аннотация
Ключом к определению кристаллических структур мембранных белковых комплексов является качество образца до кристаллизации. В частности, выбор моющего средства имеет решающее значение, так как влияет как на стабильность, так и на монодисперсность комплекса. Недавно мы определили кристаллическую структуру активного состояния родопсина крупного рогатого скота в сочетании с инженерным g белка, мини-Go, на 3,1 й резолюции. Здесь мы подробно описываем процедуру оптимизации подготовки родопсино-мини-Г-о комплекса.o Темное состояние родопсин был подготовлен в классических и неопентильных гликоля (NPG) моющих средств, а затем комплексное образование с мини-GO под воздействием света. Стабильность родопсина оценивалась ультрафиолетовой (УФ-ВИС) спектроскопией, которая отслеживает восстановление в родопсин светочувствительного лиганда, 9-cis retinal. Автоматизированная хроматография размеров (SEC) была использована для характеристики монодисперсности родопсина и родопсина-мини-Go комплекса. Электрофорус СДС-полиакриламид (SDS-PAGE) подтвердил образование комплекса, определив соотношение моляров 1:1 между родопсином и мини-Go после окрашивания геля синим цветом Coomassie. После перекрестной проверки всех этих аналитических данных мы ликвидировали неподходящие моющие средства и продолжили использование лучшего кандидата моющего средства для крупномасштабной подготовки и кристаллизации. Дополнительная проблема возникла из-за неоднородности N-гликозилирования. Гетерологовыраженный родопсин наблюдался на SDS-PAGE, чтобы иметь две различные N-гликозилатные популяции, которые, вероятно, препятствовали кристаллогенезу. Поэтому были протестированы различные ферменты дегликозилации, а эндогликосида F1 (EndoF1) производила родопсин с одним видом N-гликозилирования. Благодаря этому стратегическому конвейеру для характеристики качества белка, подготовкаo комплекса родопсин-мини-Г была оптимизирована для доставки кристаллической структуры. Это была лишь третья кристаллическая структура комплекса сигнализации белка GPCR-G. Этот подход также может быть обобщен для других мембранных белков и их комплексов для облегчения подготовки образцов и определения структуры.
Введение
Определение кристаллических структур мембранных белков и их комплексов всегда было сложным из-за трудностей в получении хорошо дисфрасдействующих кристаллов. В отличие от растворимых белков, интегральные мембранные белки составляют гидрофобное ядро, которое охватывает клеточную мембрану. Для удаления мембранных белков из клеточной мембраны в водиной буфер, моющие средства должны быть использованы для формирования моющего средства-белка мицелле, тем самым заменяя липиды вокруг гидрофобного ядра мембранных белков. Стабильность, активность и целостность мембранных белков напрямую зависят от химических и структурных свойств моющего средства1,а свойства моющего средства также определяют размер мицелле. Большой моющий мицелл может окклюзии гидрофильных поверхностей небольшой мембранный белок, тем самым предотвращая кристаллизацию из-за отсутствия кристаллических контактов при использовании метода диффузии пара. Небольшой моющий мицелл является выгодным для кристаллографии, но короткоцепочечные моющие средства, как правило, жестче и, следовательно, приводят к дестабилизации и агрегации мембранного белка. Поэтому, перед кристаллизацией, дополнительная процедура скрининга моющего средства незаменима, типично пристреливая более короткие моющие средства которые все еще поддерживают стабилность протеина.
G белковые рецепторы (GPCRs) являются интегральными мембранными белками, содержащими семь трансмембранных селиков. GPCRs существуют в двух основных состояниях, либо неактивное состояние, стабилизированное обратными агонистами или антагонистами, или активное состояние, связанное с агонистом и стабилизированное белком G, хотя вполне вероятно, что между этими двумя крайностями существует множество субгосударств. Структурное определение GPCRs первоначально сосредоточено на неактивных государствах, связанных с обратными агонистами и антагонистами из-за их более высокой стабильности, чем активные государства2. Когда GPCRs активируются при агонисте связывания, рецепторы очень динамичны, и расщелина формы переходно на цитоплазмамическое лицо рецептора для G белка соединения. Считается, что именно этот динамизм является причиной того, что GPCRs, связанные с агонистом, часто более нестабильны, чем неактивное состояние. Таким образом, становится необходимым для проверки моющих средств, подходящих для конформационного состояния исследуемого рецептора, потому что вполне вероятно, что для изучения активного состояния потребуются более мягкие моющие средства по сравнению с неактивным состоянием.
В этом отчете мы используем визуальный GPCR, бычий родопсин3, и его комплекс с мини-Go белок4,5 для моющего средства скрининга экспериментов, представляющих неактивное состояние и активное состояние, соответственно. Скрининг моющего средства был сосредоточен на классических алкил-мальтосайдных и глюкоцидных моющих средствах и неопентиновых гликолях (NPG). В этом контексте, классическое моющее средство построено из сахарной головы группы и алкил цепи, в то время как NPG типа моющих средств содержит два одинаковых классических моющих средств, которые сливается с четырехместный углерод на стыке между сахарами и алкил цепи6,7,8.
Экспериментальный рабочий процесс был разработан, начиная от очистки родопсина в различных моющих средств, а затем формирование родопсина-мини-Go комплекса и заканчивая характеристикой комплекса с использованием нескольких методов (Рисунок 1). Для неактивного состояния родопсина, восстановление светочувствительной лиганд9-цис жевательной сети контролировалась ультрафиолетовой видимой (УФ-ВИС) спектроскопией. Спектр показывает физико-химическое состояние сопредила и указывает на ее окружающую среду в связывающем кармане родопсина. Для оценки монодисперсности очищенного родопсина, а также формирования родопсина-мини-Г-о-комплексаo использовалась хроматография размеров исключения (SEC). По мере того как SEC дифференцирует молекулы протеина их размером и формой, агрегированная популяция протеина можно определить по мере того как они elute в томе пустоты. Для подтверждения сложного образования фракции из SEC были оценены натрием dodecyl сульфат-полиакриламид гель электрофорес (SDS-PAGE), чтобы подтвердить наличие как родопсина и мини-Go.
Еще одним фактором, который необходимо учитывать, являются посттрансляционные изменения (ПТМ) на мембранных белках. ПТМ, такие как N-гликозилирование часто наблюдаются на эукариотических мембранных белков, вырабатываемых в системах экспрессии клеток млекопитающих и насекомых. Ограниченный Штамм N-гликозилирования клеток эмбриональной почки человека 293 (HEK293) был разработан путем удаления гена, кодирующего N-ацетилглюкоза I (GnTI), в результате чего однородная N-гликозилация glcNAc2Man5 на сайте консенсуса Asn-X-Ser/Thr. Хотя N-гликозилирование можно предотвратить путем мутации аминокислотных остатков в месте консенсуса, это также может изменить функцию белка или эффективность складывания. В бычьем родопсине мутация N-гликозитилированных остатков Asn15 приводит к неправильному сворачиванию и уменьшению активации белка G9,,10. Родопсин, используемый в этом докладе, был выражен в HEK 293 GnTI-дефицитной клеточной линии. Тем не менее, SDS-PAGE показал наличие двух видов родопсина. Эта неоднородность может предотвратить образование кристаллов и, следовательно, дегликозилирование с использованием пептида-N-гликозидаза F (PNGase F) и эндогликосидаза F1 (Endo F1) был протестирован. Дегликозилированный продукт характеризовался SDS-PAGE и жидкой хроматографией-масс-спектрометрией (LC-MS) для определения уровня гликозилирования и ее однородности.
протокол
ПРИМЕЧАНИЕ: Этот протокол для скрининга моющего средства подробно описан для 30 г клеточных гранул HEK293 в качестве исходного материала.
1. Материалы, химические вещества и реагенты
ПРИМЕЧАНИЕ: Все решения готовятся с использованием аналитических реагентов класса и ультрачистой воды, которая очищается от деионизированной воды, чтобы достичь резистоспособность 18,2 МЗ см при 25 градусах Цельсия.
- Решения для буферных акций
- Подготовка 10x фосфат буферного солья (10x PBS).
- Подготовка БУФЕРа HEPES: 1 M, тиснено до рН 7,5 с NaOH.
- Подготовка 5 M NaCl.
- Подготовка 2 M MgCl2.
ПРИМЕЧАНИЕ: Все биржевые решения проходят через фильтр 0,22 мкм для поддержания их бесплодия.
- Решения для акций Детергента
- Подготовка dodecyl maltoside (DDM), 10% (w/v).
- Подготовка децил мальтосайд (DM), 10% (w/v).
- Подготовка 6-циклогексил-гексил мальтоид (Cymal-6), 10% (w/v).
- Подготовка 5-циклогексил-пентил мальтоид (Cymal-5), 10% (w/v).
- Подготовка неил глюкоцид (C9G), 10% (w/v).
- Подготовка лаурилсолоза неопентилгликоль (LMNG), 5% (w/v).
- Подготовка децил мальтоза неопентилгликоль (DMNG), 10% (w/v).
- Подготовка cymal-6 неопентилгликоль (C6NG), 10% (w/v).
- Подготовка cymal-5 неопентилгликоль (C5NG), 10% (w/v).
- Подготовка октил глюкозы неопентилгликоль (OGNG), 10% (w/v).
ПРИМЕЧАНИЕ: Для 10% моющего средства бульона раствор, растворить 1 г моющего порошка в ультрачистой воде с нежным раскачиванием, а затем настроить окончательный объем до 10 мл. Раствор детергента должен храниться при -20 градусов для длительного хранения и на льду во время работы.
ВНИМАНИЕ: Бутилированные моющие средства, как правило, рекомендуется хранить на -20 градусов морозильник. Бутылки, содержащие порошок моющего средства, следует нагревать до комнатной температуры перед открытием. Порошок детергента гигроскопичен, поэтому уравновешивание температуры предотвратит образование конденсации, которая смочит моющее средство.
- Другие химические вещества и реагенты
- Приготовьте 1D4 иммуноаффинити агарозной минины: 10 мл 50% шлама.
ПРИМЕЧАНИЕ: 1D4 иммуноаффинити агарозы являются агарозные бусы связаны с моноклональным Rho1D4 антитела, который связывает последние 9 аминокислот крупного рогатого рода TETS-VAPA как эпитоп. Агароза иммуноаффинити 1D4 работает как материал очистки сродства для захвата белков, содержащих последовательность C-терминала 1D4. Этот материал очистки может быть подготовлен9,,11 или приобретены. - Приготовьте 9-cis раствор для стинальной кисти: 1 мМ, растворенный в 100% этаноле.
ПРИМЕЧАНИЕ: Предотвращение воздействия света на ретину во время подготовки и хранения. - Приготовьте 1D4 пептид (последовательность ТЭЦВАПА): 800 мкм, растворенный в воде.
- Приготовьте 1D4 иммуноаффинити агарозной минины: 10 мл 50% шлама.
- Буферов
ПРИМЕЧАНИЕ: Все буферы смешиваются от фондовых решений до желаемой концентрации. Все буферы охлаждаются до 4 градусов цельсия перед использованием.- Подготовка буфера A: PBS, 0.04% DDM.
- Подготовка Буфер B: 20 мМ HEPES pH 7.5, 150 mM NaCl, 0.04% DDM.
- Подготовка буфера C: 20 мМ HEPES pH 7.5, 150 мМ NaCl, и моющее средство при их рабочей концентрации, перечисленных в таблице 1.
- Подготовка Буфер D: 20 мМ HEPES pH 7.5, 150 mM NaCl.
- Подготовка elution буфера: 20 мМ HEPES pH 7.5, 150 mM NaCl, 80 мкм 1D4 пептид, и моющее средство в их рабочей концентрации.
- Подготовка буфера SEC: 20 мМ HEPES pH 7.5, 150 мМ NaCl, 0.025% DDM; фильтруется через фильтр 0,22 мкм.
- Растворитель для LC-MS
- Подготовка растворителя A: ацетонитрила, содержащего 0,1% formic кислоты.
- Подготовка растворителя B: ультрачистая вода, содержащая 0,1% для модной кислоты.
- Подготовка растворителя C: изо-пропанол.
2. Растворить клеточную мембрану и экстракцию белка
- Оттепель 30 г HEK293 GnTI- клеточные гранулы, выражающие бычий родопопсин мутант N2C/M257Y/D282C3,9 до комнатной температуры, добавьте 120 мл 1x PBS буфера, содержащего коктейль-ингибитор протеазы, и гомогенизируют с помощью гомогенизатора Dounce или электрического гомогенизатора (13 000 r.00) для 3.00. Соберите гомогенизированную клеточную подвеску в стакан и отрегулируйте объем до 150 мл.
ПРИМЕЧАНИЕ: 30 г клеточных гранул эквивалентно 3 l клеточной культуры при плотности 2 х 106 клеток/мл. - Аккуратно добавьте 10% ДДМ в гомогенизированные клетки, чтобы дать окончательную концентрацию 1,25%. Перемешать на льду 1 ч.
- Центрифуга лизата клетки при 4 кв и 150000 х г в течение 45 минут, чтобы удалить нерастворимый мусор.
- Перенесите супернатант в бутылку 500 мл и добавьте 10 мл 1D4 иммуноаффинити агарозной оси (50% суспензии). Аккуратно перемешайте растворимый клеточный лизат и сеятину в течение 4 ч или на ночь при 4 градусах Цельсия.
- Загрузите смесь lysate/resin к открытому столбце для того чтобы собрать участь.
- Вымойте с помощью 10 объемов колонки (CV) мытья Буфер А.
ПРИМЕЧАНИЕ: Объем столбца — это объем упакованных (100%) агароуз агарозы используется. В этом случае 1 CV составляет 5 мл. - Приостановить с исправления с 2 CV буфера А.
ВНИМАНИЕ: Начиная со шага 2.8 и далее, шаги, которые должны быть выполнены в состоянии тусклого красного света, помечены как «Темный» в начале описания. - (Темный) Добавьте 9-cis ретину к повторной мисе до конечной концентрации 50 мкм. Аккуратно перемешайте при 4 градусах по Цельсию 4-16 ч в темноте.
ПРИМЕЧАНИЕ: Более короткое время инкубации может привести к неполному восстановлению ретины. - (Темный) Удалите поток через столбец. Вымойте с мелисомой с 20 CV Буфер А, а затем 15 CV Буфер B.
- (Темный) Приостановите действие мелины в 2 CV Buffer B, а затем разделите суспензию с ресиней поровну на 10 10-мл колонн.
- (Темный) Удалить поток через из столбца, а затем повторно гофы в 1 мл буфера C. Инкубировать в течение 1 ч при 4 c.
- (Темный) Повторите шаг 2.11.
- (Темный) Удалите поток через столбец, а затем повторно приостановить посевную в 0,8 мл Elution Buffer для каждого столбца. Аккуратно перемешать по 2 ч.
- (Темный) Соберите elution от колонки в трубку 2 мл.
- (Темный) Отрежь по ресину в 0,7 мл буфера Elution для каждой колонки. Аккуратно перемешать по 1 ч.
- (Темный) Соберите elution из колонны в ту же трубку.
3. спектроскопия УФ-ВИС
- Подготовьте спектрофотометр, чтобы покрыть диапазон измерений 250-650 нм. Запись базовой линии с помощью воды или Elution Buffer.
- (Темный) Загрузите элютированный белок в кварцевый кювет. Измерьте спектр протеинового образца.
- (Темный) Осветите белок непосредственно в кюветте в течение 2 мин со светом, проехав через 495 нм долгопроходя фильтр.
- Измерьте спектр освещенного образца.
- Выполните одно и то же измерение для всех образцов белка, очищенных в других 9 моющих средствах, как темных, так и освещенных состояниях.
- Участок кривых (абсорбция против длины волны) в X-Y рассеяния диаграммы.
4. Автоматизированная хроматография размеров родопсина и родопсина-мини-Go комплекса
- (Темный) Концентрируйте белок до 100 кЛ при центрифугации с помощью спинового концентратора с молекулярным отсечкой веса (MWCO) в 30 кДа при 4 градусах Цельсия. Переконцентрированные образцы можно разбавлять с помощью потока через концентратор или буфер C. Чтобы определить концентрацию протеина образца, измерьте абсорбцию на уровне 280 нм с помощью спектрофотометра.
ПРИМЕЧАНИЕ: Начиная со шага 4.2 и далее, эксперимент не требует темной среды, и поэтому образцы могут быть подготовлены при нормальном свете. - Приготовьте 100 л родопсина при 0,7 мг/мл для каждого состояния моющего средства.
- Приготовьте смесь родопсина (0,7 мг/мл) и мини-Go4,,12 (0,2 мг/мл) смеси для каждого моющего средства. Дополнить смесь 1 мМ MgCl2. Осветите смесь светом от фильтра длиной 495 нм и инкубировать в течение 30 минут.
- Установите 24 мл геля фильтрации колонки с диапазоном фракционирования 10-600 кДА шарового белка на жидкой очиститель хроматографии. Равновесие столбца с буфером SEC.
ПРИМЕЧАНИЕ: очиститель жидкой хроматографии оснащен автосэмпером, детектором длины волны и фракционным коллектором. - Перенесите образцы на флаконы автосэмпера и поместите их в поднос образца. Программа метод файл для автоматизации последовательных SEC работает для каждого образца, с автосэмпером загрузки 77 л образца в столбец, и очиститель eluting 24 мл буфера SEC на скорости потока 0,5 мл / мин за пробег. Запись абсорбции на 280 нм и 380 нм.
- Соберите пиковые фракции родопсина иo родопсина-мини-G-O комплекса при объеме удержания около 12,9 мл.
- Проанализируйте левые образцы родопсина со ступени 4.2 иo пиковые фракции родопсина-мини-G-комплекса на 4-12% SDS-денатурный градиент гели с Coomassie синий окрашивания.
- Участок elution хроматограмма(A 280 илиA 380 против удержания объема).
5. Исследование дегликосии и LC-MS
- Для исследования LC-MS используйте только образец родопсина, очищенный в моющих средствах LMNG.
- Приготовьте смесь родопсина в 200 л при 1 мг/мл и PNGase F13 при 0,01 мг/мл. Хорошо перемешать и инкубировать при 4 градусах Цельсия на ночь.
- Приготовьте смесь родопсина в 200 л при 1 мг/мл и Эндо F113 при 0,01 мг/мл. Хорошо перемешать и инкубировать при 4 градусах Цельсия на ночь.
- Проанализируйте результат пищеварения с помощью SDS-PAGE и Coomassie синего окрашивания.
- Концентрируйте необработанные и обработанные Эндо F1 образцы родопсина и при условии очистки SEC в Буфере D.
ПРИМЕЧАНИЕ: Это подготовить образец с минимальным количеством моющего средства для исследования LC-MS. Буфер D не содержит моющего средства, но из-за медленной нескоростной Скорости LMNG из мембранного белка14, родопсин не будет агрегировать. - Соберите пиковую фракцию при удержании объема около 12,9 мл. Концентрата до 1 мг/мл с помощью спинового концентратора (MWCO 30 kDa).
- Введите 10 мкг белка в столбец Reprosil 200 C18-A и ущелье столбца с помощью линейного метода градиента с растворителем состава и настроек, перечисленных в таблице 2. Поток делится до 25% для масс-спектрометра и 75% для обнаружения УФ-излучения.
Результаты
Экспериментальный рабочий процесс по подготовке и анализу образцов обобщен на рисунке 1. Использование открытых столбцов для мелкомасштабной очистки сродства позволило нам параллельно готовить образцы в различных моющих условиях(рисунок 1А). Такая мелкомасштабная установка очистки дала достаточно белка для дальнейшего анализа с использованием спектроскопии УФ-ВИС, SEC и SDS-PAGE(рисунок 1B-C).
Спектроскопия УФ-ВИС показала стабильность родопсина
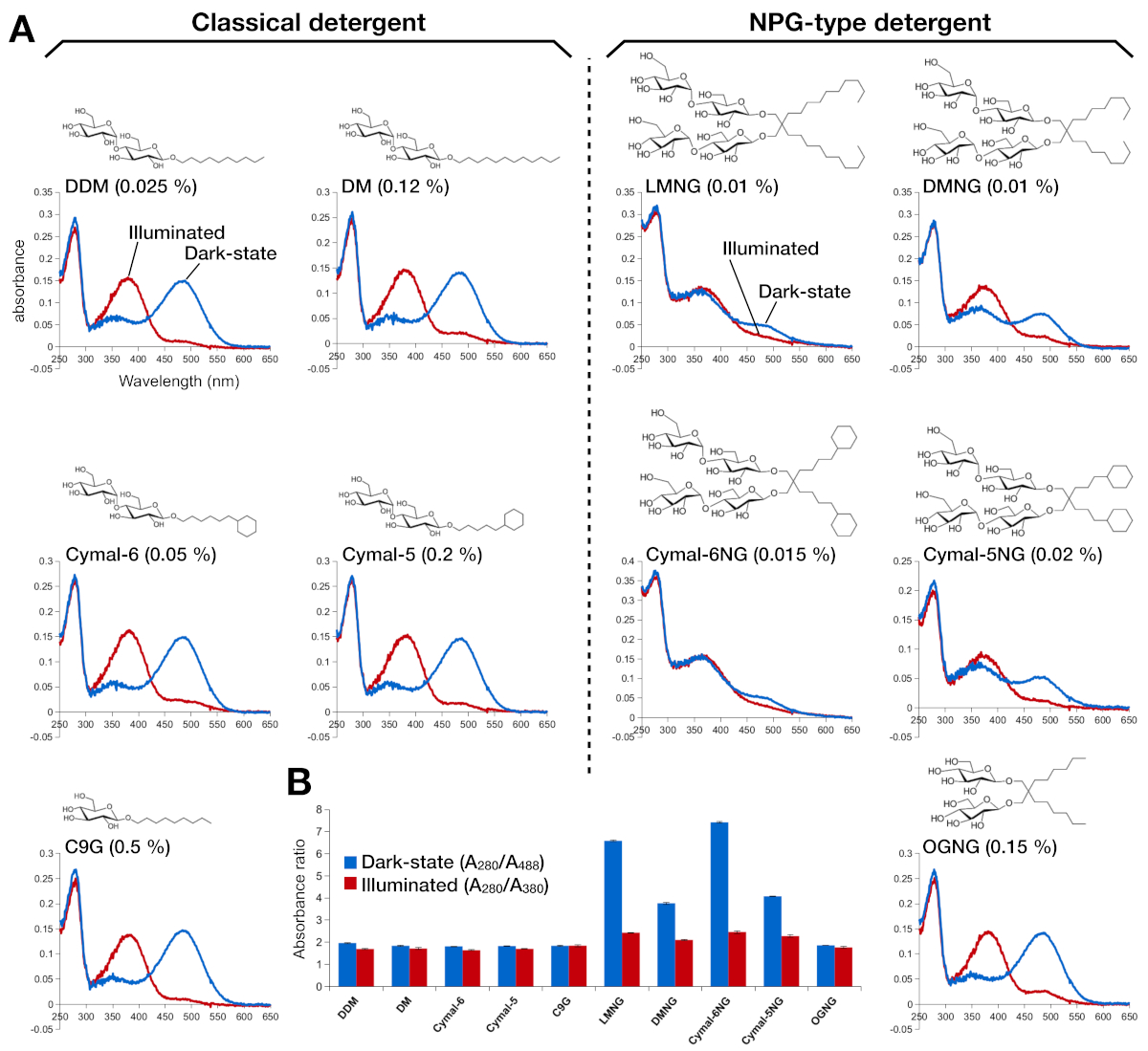
Стабильность восстановленного с ыветной социера родопсина оценивалась по его оптическому абсорбции(рисунок 2). В темном состоянии, 9-cis жеренная корыт ковальц ковалентно связана с Lys296 как протонная база Шифф. После освещения, 9-cis сетчинеизируются до все-транс изоформы и Шифф базовое звено депротонным. Протонная 9-cis жевательная подчет дает пик поглощения на 488 нм, в то время как депротонированный все-транс жетины имеет пик в 380 нм. СПЕКТРы УЛЬТРА-VIS родопсина в ДДМ показали типичное поглощение 9-cis сцепления связаны и свет-активированный родопсин, где синий сдвиг 108 нм с примерно такой же оптической плотностью было четко наблюдается(Рисунок 2А, верхняя левая панель). Когда родопсин дестабилизируется, а затем связывающийся карман для изменения сытины, что приводит к депротонации ретины и, возможно, диссоциации. Если это произойдет, и тогда спектр показывает вклад от депротонации, а также свободную форму ретины15. Таким образом, мы определили эффективность восстановления реконституции сытины в родопсин по коэффициенту абсорбции между белком (280 нм) и женейной (488 нм для протонатированной 9-cis ретины, 380 нм для депротонного всетрансового жетины) (Рисунок 2B). Образцы родопсина, очищенные в классических моющих средствах (DDM, DM, Cymal-6, Cymal-5, C9G), показывают тот же оптический профиль. Тем не менее, образцы, очищенные в моющих средствах NPG (LMNG, DMNG, Cymal-6NG, Cymal-5NG) показывают оптические профили, указывающие на неоптимальную привязку к шее, за исключением образца OGNG, который дал тот же оптический профиль, что и образец DDM.
Хроматография, исключенная из размера, показала чистоту образца и монодисперсность белка.
SEC является эффективным и надежным аналитическим инструментом для оценки образцов белка во время подготовки и скрининга. Он проверяет чистоту образца от предыдущего шага очистки, а также монодисперсность белковых молекул. Для родопсина и его мини-Go комплекса, качество образца было интерпретировано из кривых поглощения на 280 нм и 380 нм(рисунок 3A). 280 нм следы показали наличие белка, и 380 нм след показал наличие ретины. Любые сигналы, появляющиеся в объеме пустоты (около 8 мл при использовании этой колонки), приписывались белковым агрегатам. Таким образом, результаты показали, что образцы, подготовленные в классических моющих средств были в монорасходном состоянии, за исключением C9G, где некоторая часть агрегата появились. В отличие от этого, образцы, подготовленные с использованием моющих средств типа NPG, содержали гораздо больше агрегатов, чем образец C9G; Наиболее совокупное образование привело к образованию LMNG и Cymal-6NG, но в DMNG и Cymal-5NG было отмечено меньше агрегатов. Исключением был OGNG, который показал аналогичный профиль DDM. Белковые агрегаты, eluting на объеме пустоты также имели более плохую заполняемость сеток, как показано в соотношении A280/A380, которое увеличилось по сравнению с пиком при объеме удержания в 12,9 мл, соответствующем 135 kDa. Еще одной особенностью мы наблюдали, что родопсин и родопсин-мини-Go eluted вокруг того же объема удержания(Рисунок 3B). Это неудивительно, потому что кажущийся молекулярный вес моющего средства родопсин был 120 kDa и родопсин-мини-Go 144 kDa. Поэтому мы не смогли определить комплексное образование только на основе данных SEC, поэтому SDS-PAGE был использован для дальнейшего анализа очищенной SEC выборки.
SDS-PAGE подтвердила комплексное образование
SDS-PAGE является стандартным методом для идентификации белковых компонентов в образце. Концентрированный родопсин (до очистки SEC) были проанализированы SDS-PAGE, чтобы подтвердить его чистоту, и показали две полосы около 37 kDa и смазанную полосу выше 50 kDa(рисунок 4A). Более низкие 2 полосы более поздно были подтвержены для того чтобы иметь по-разному положения N-гликозилации. Полоса выше 50 kDa была интерпретирована как агрегированные родопсиновые олигомеры, индуцированные буфером образца SDS-PAGE, поскольку эти агрегаты не были соблюдены в SEC или любых других методах обнаружения. Поскольку данные SEC не могли подтвердить комплексное образование, с помощью SDS-PAGE были проанализированы eluted фракции SEC из образцов родопсина-мини-Go. SDS-PAGE показал белковые полосы как родопсина, так и мини-Go во всех моющих условиях, предполагая, что комплекс был сформирован независимо от выбора моющего средства(рисунок 4B).
Спектрометрия LC-MS определила n-гликозилирование в родопсине
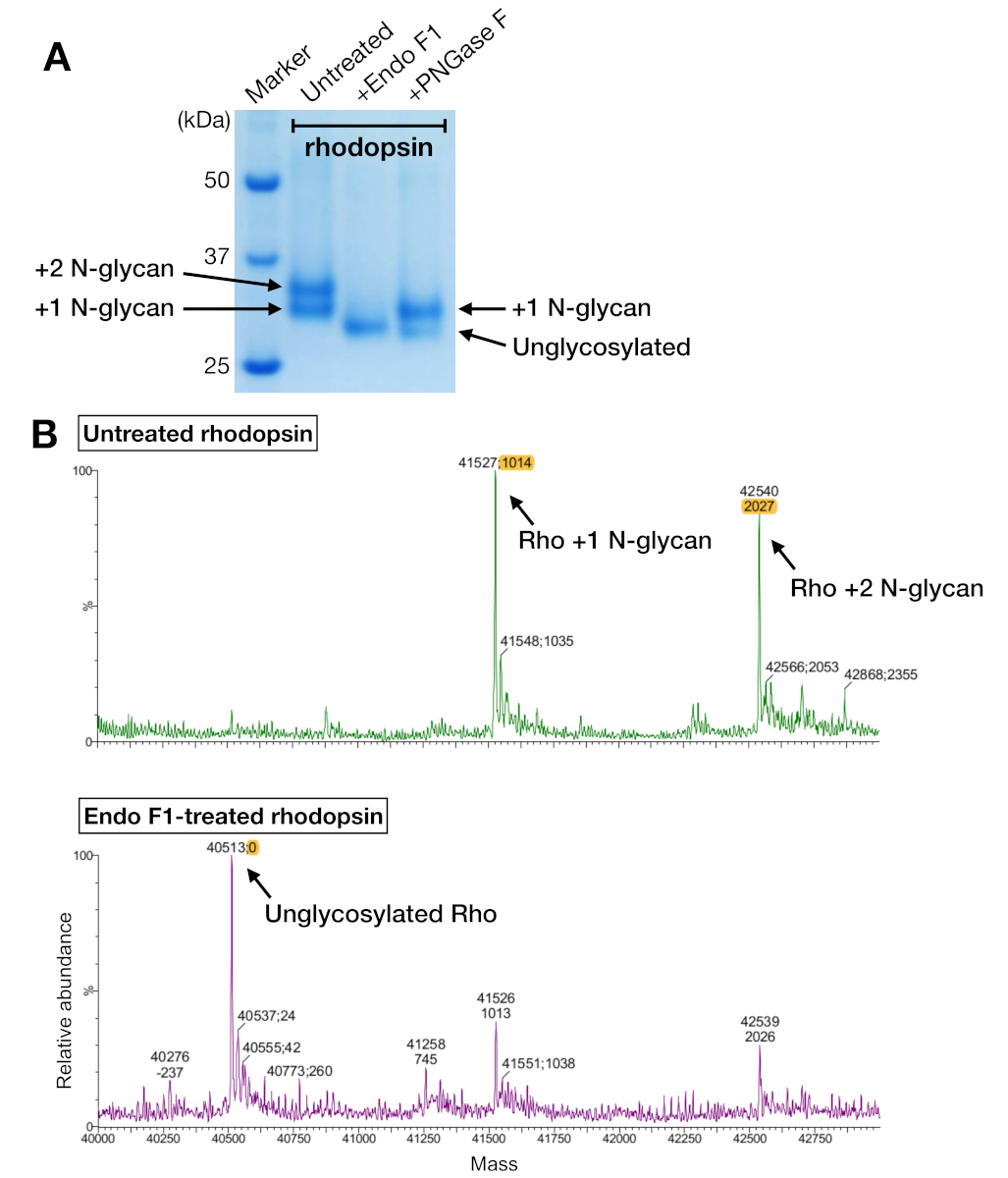
Образцы родопсина как из очистки сродства, так и SEC показали две белковые полосы, которые мигрировали с видимым молекулярным весом около 37 кДа на геле SDS-PAGE, который не мог быть отделен SEC при использовании 24-мл колонки. Различные модели N-гликозилирования на гетерологло-выраженном родопсине от HEK 293 GnTI- клетки были наиболее вероятным объяснением. Поэтому два фермента, PNGase F и Endo F1, были протестированы на их способность к дегликозилату родопсина. Из данных SDS-PAGE, Endo F1 уменьшил молекулярный вес обеих белковых полос в один продукт, в то время как переваривание PNGase F все еще давало две популяции(рисунок 5A). Непереваренные и Эндо F1 обработанных образцов были проанализированы с помощью СПЕКТРометрии LC-MS для выявления масс различных видов. Данные показали, что родопсин, произведенный в HEK 293 GnTI- клетки, содержащие ся один или два N-гликанов, с разницей в массе 1014-1 Da. Эндо F1-обработанный родопсин не содержит N-гликанов и имел массовую разницу 2027-1 Da по сравнению с родопсином, содержащим два N-ггликанов. Эти результаты согласуются с отсутствием фермента N-acetylglucosaminyltransferase I в клеточной линии, используемой для экспресс-родопсина, что приводит к тому, что все N-гликаны имеют структуру GlcNAc2Man5, (масса 1014 Da).

Рисунок 1: Подготовка образца и характеристика для эксперимента по скринингу моющих средств. ()Подготовка образцов родопсина в различных моющих средствах во время очистки. (B) Методы, используемые в протоколе: УФ-VIS спектроскопия, размер исключения хроматографии (SEC), SDS-PAGE и жидкой хроматографии-масс-спектрометрии (LC-MS). (C) Экспериментальный рабочий процесс для характеристики родопсина, родопсина-мини-Go, и дегликозилационный продукт родопсина. Пожалуйста, нажмите здесь, чтобы просмотреть большую версию этой цифры.
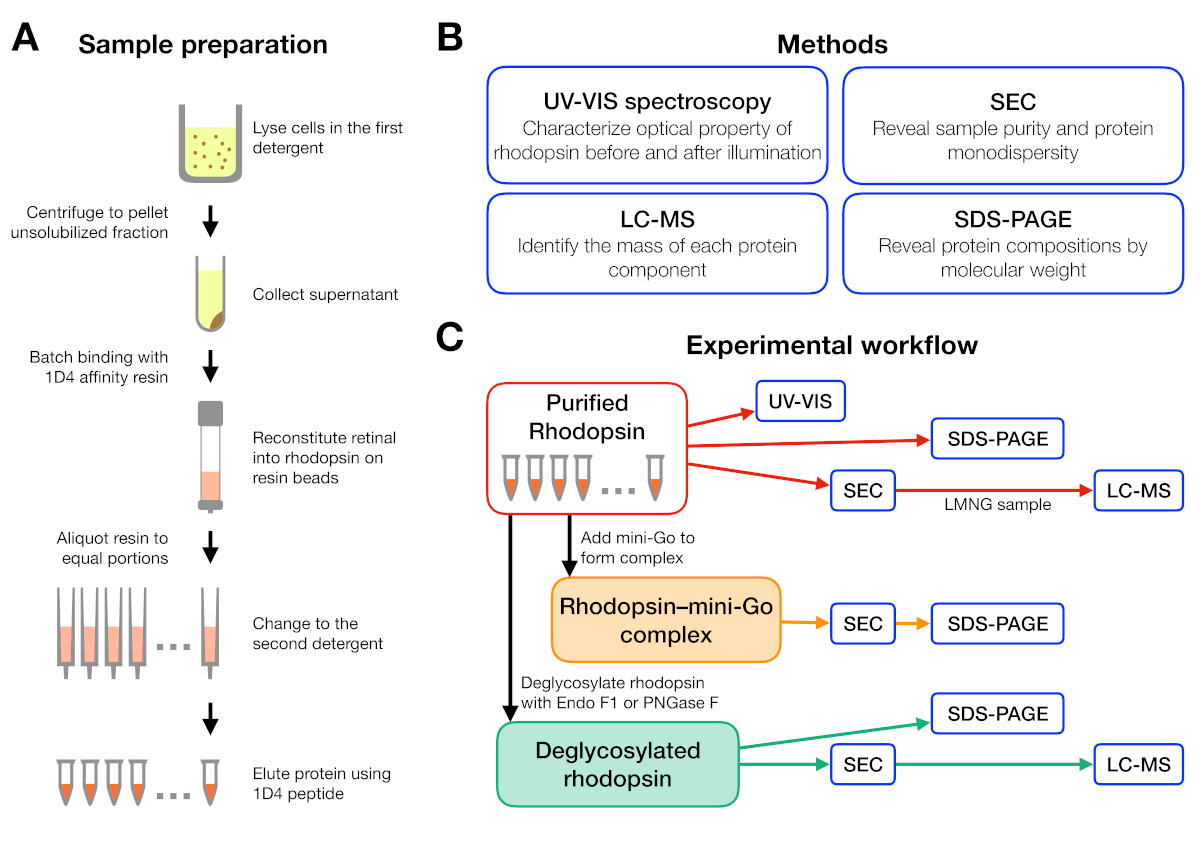{kind=link}

Рисунок 2: спектроскопия родопсина УФ-ВИз. (A)УФ-VIS спектры родопсина. Спектры темного состояния, 9-cis связанных с говтанием, родопсин показаны синими кривыми. После освещения 9-cis сетчатый сетчатый цвет раздевят и изорезы во все транс сетчатые, а спектры освещенного родопсина показаны как красные кривые. Химическая структура каждого моющего средства показана как всетра. (B) Соотношения A280/A488 (синий бар) и A280/A380 (красный бар) изображают стабильность родопсина в темном состоянии и светлое состояние, соответственно. Пожалуйста, нажмите здесь, чтобы просмотреть большую версию этой цифры.
{kind=link}

Рисунок 3: Размер исключения хроматографии профилей родопсина и родопсина-мини-Go комплекс очищенв в 10 различных моющих средств. (A) Левая панель показывает SEC профили образцов очищены в классических моющих средств. Правая панель представляет профили SEC образцов, очищенных в моющих средствах типа NPG. Профиль стандартных маркерных белков отображается как наложение вместе с образцом DDM. Интерпретация пиковых профилей показана для DMNG, с идеальным сценарием (без агрегатов), наблюдаемым для DDM, DM, Cymal-6, Cymal-5 и OGNG. (B) Увеличенный профиль образца OGNG в объеме удержания 12-14 мл. Все образцы были проанализированы с помощью колонки Superdex200 Увеличение 10/300 GL. Пожалуйста, нажмите здесь, чтобы просмотреть большую версию этой цифры.
{kind=link}

Рисунок 4: SDS-PAGE анализ родопсина и родопсина /мини-Го комплекса. (A) Образцы родопсина очищены в моющих средствах. Размазанная полоса выше 50 кДА приписывается агрегированным олигомерам родопсина, индуцированным буфером образца SDS-PAGE. (B) SEC очищенные образцы родопсина / мини-Go комплекса. Родопсин с 1 и 2 N-гликан и мини-GO изображены. Пожалуйста, нажмите здесь, чтобы просмотреть большую версию этой цифры.
{kind=link}

Рисунок 5: Выявление гликозилирования у родопсина. (A) SDS-PAGE анализ дегликозилированных родопсин с использованием PNGase F и Эндо F1. (B) СПЕКТРы LC-MS родопсина без (верхняя панель) и с deglycosylation Endo F1 (нижняя панель). Для подготовки родопсина-мини-Go комплекса для кристаллизации, мы выбрали Endo F1 вместо PNGase F, потому что Эндо F1 поставил один однооднородный вид родопсина. Пожалуйста, нажмите здесь, чтобы просмотреть большую версию этой цифры.
{kind=link}
| Моющего средства | Концентрация рабочей работы (%) | Критическая концентрация мицелле (%) |
| Ddm | 0.025 | 0.0087 |
| Dm | 0.12 | 0.087 |
| Кималь-6 | 0.05 | 0.028 |
| Кималь-5 | 0.2 | 0.12 |
| C9G | 0.5 | 0.2 |
| LMNG | 0.01 | 0.001 |
| DMNG | 0.01 | 0.0034 |
| Cymal-6NG | 0.015 | Недоступно; должно быть ниже 0,056 |
| Cymal-5NG | 0.02 | 0.0056 |
| ОГГ | 0.15 | 0.058 |
Таблица 1: Концентрация моющего средства буфера C.
| Время (мин) | Солунт А (%) | Солунт B (%) | Солунт C (%) | Скорость потока (мл/мин) |
| 0 | 0 | 95 | 5 | 0.5 |
| 1 | 0 | 95 | 5 | 0.5 |
| 5 | 20 | 75 | 5 | 0.6 |
| 25 | 85 | 10 | 5 | 0.6 |
| 26 | 90 | 5 | 5 | 0.6 |
| 30 | 90 | 5 | 5 | 0.6 |
Таблица 2: Параметры выражания столбцов.
Обсуждение
Успех в кристаллизации белка сильно зависит от протеина образца, особенно мембранных белков и их комплексов из-за осложнения, вызванного моющими средствами. Данный отчет демонстрирует скрининг моющих средств и оценку качества образцов для белковых сигнальных комплексов GPCR-mini-G. Различные методы были широко использованы для изучения биохимического свойства мембранных белков, например, термостабильность анализ с использованием флуоресцентных красителей16,17, связывание анализ для обнаружения комплексного образования путем измерения изменения в триптофан флуоресценции сигнал18 или резонансной передачи энергии с биосенсорами19. Тем не менее, химическая среда, используемая в этих методах, сильно отличается от тех, для подготовки образца кристаллизации, либо белки находятся в тысячу раз ниже концентрации для измерения на основе флуоресценции, или белки встроены в липидные бислои или в одном фиксированном состоянии моющего средства. В этом протоколе используемые методы также стандартизируются в крупномасштабной подготовке образцов перед кристаллизацией. Таким образом, оптимизированные параметры могут быть легко переданы для подготовки по шкале кристаллизации без дальнейшего крупного скрининга и оптимизации.
Целью данного протокола является оптимизация подготовки стабильного и однородного белкового комплекса GPCR-mini-G для кристаллизации и определения структуры рентгеновской кристаллографией. Протокол интегрирует набор методов для качественной оценки воздействия моющего средства и дегликозилирования при подготовкеo родопсино-мини-Г-о комплекса. Родопсин в неактивном состоянии и светоактивированном состоянии, связанном с и без предуцинового пептида, кристаллизуется при очищении в моющих средствах октилговый глюкозид (C8G)20,21,22 и C9G23,24. Поскольку родопсин-мини-G-комплекс, очищенный в C8G и C9G, не давал кристаллов (данные не показаны), мы затем исследовали более широкий спектр других моющих средств, используя описанную стратегию (Рисунок 1).o Воспользовавшись светочувствительностью родопсина, мы вполне могли бы следовать восстановлению ретины на длинах волн, кроме 280 нм. В спектроскопии УФ-ВИС и SEC мы обнаружили ретину на уровне 380 нм или 488 нм. Однако большинство мембранных белков не имеют такого удобного хромофора, чтобы следовать функциональности во время очистки. Другие варианты были бы сделать ligand обнаруживаемым путем добавлять свет-обнаруживаемый хромофор или путем использование radioligand-связывающих и термических анализов переноса25.
Родопсин имеет молекулярный вес 40 кДа. Из-за массы моющего средства он связывает, его очевидный молекулярный вес на SEC составляет около 120 kDa. Поэтому неудивительно, что связывание мини-Go (24 kDa) не было легко обнаружено на SEC, так как это потребовало бы дифференциации белков с явными массами 120 kDa и 144 kDa. Таким образом, для подтверждения чистоты и сложного образования был использован анализ фракций SEC по SDS-PAGE. Даже если профили SEC показывают четкий сдвиг при комплексном образовании, все равно рекомендуется проводить анализ SDS-PAGE, чтобы подтвердить комплексное образование с правильными связывающими партнерами, а не с другими со-очищенными белковыми загрязнителей.
Родопсин и мини-Go были очищены в миллиграммовых количествах, что позволило использовать низкой чувствительности обнаружения комплексов, таких как УФ-VIS поглощения во время SEC и Commassie Blue окрашивания SDS-PAGE гелей. Там, где образцы ограничены, следует использовать более чувствительные обнаружения, такие как очиститель LC, оснащенный детектором флуоресценции для отслеживания сигналов триптофана от белка (280 нм возбуждение, 350 нм выбросов) и серебра для гелей SDS-PAGE. Другой подход заключается в том, чтобы сплавить флуоресцентный белок, такой как зеленый белок флуоресценции (GFP) к интересуемому белку, который также позволит обнаружить даже во время выражения белка26, но он должен быть удален до кристаллизации.
Важно обеспечить, чтобы очищенный белок также был свободен от неоднородности, возникающей в результате переменных ПТМ. В случае, описанном здесь, две популяции родопсина, наблюдаемые на гелях SDS-PAGE, были охарактеризованы как имеющие один или два N-гликанов. Переменная модификация белка потенциально предотвратит образование хорошо дисфракционных кристаллов, поэтому мы поэтому дегликозаливом родопсина. Эндогликосидаза Эндо F1 был наиболее эффект эндогликосидазе испытания и лечение привело к одному виду унгликозилированных рецепторов, в то время как PNGase F только частично удалены гликанов на родопсин и в результате смеси родопсин полностью unglycosylated или с одним N-гликан остался. Родопсин без лечения дегликозилазы был успешно кристаллизуется3,27,28, и N-гликан на родопсин Asn15 важно сформировать кристаллический контакт в тех случаях. В случае родопсина-мини-Go,необходимо удалить N-гликанов Эндо F1 для получения кристаллов. Существует нет стандартизированного правила дегликозируется белков, представляющих интерес до кристаллизации, но удаление неоднородных ПТМ следует рассматривать, когда белки не кристаллизуется после обширных испытаний кристаллизации.
Описанные здесь данные и методология привели нас к выбору OGNG в качестве наиболее предпочтительного моющего средства для кристаллизациикомплекса родопсин-мини-Г из-за его небольшого размера мицелле и его способности стабилизировать комплекс. Мы также использовали Endo F1 для обеспечения очищенного родопсина был однородным видом. Кристаллы были впоследствииполучены,и мы определили кристаллическую структуру до 3,1 й4, который был только третьей кристаллической структурой GPCR-G белка сигнального комплекса14,29.
Для мембранных белков, связанных с и без белка партнера, они должны рассматриваться как два различных белка. Белок в разных функциональных состояниях имеет различные конформации и находится на разном энергетическом уровне. Поэтому рекомендуется оптимизировать протокол подготовки для каждого функционального состояния, так как параметр для неактивного состояния не может быть полностью перенесен в активированное состояние. Также, не упоминать изменение в свойстве протеина осложненного связыванием протеина соучастника. Протокол использует методы, которые стандартизированы для подготовки образца кристаллизации для подготовки неактивного мембранного белка в различных моющих средствах, с последующим активацией белка и сложным образованием, а также для характеристики качества белка. Таким образом, этот протокол можно легко обобщить с другими мембранными белками и их комплексами для структурных исследований с незначительными изменениями.
Раскрытие информации
CGT является консультантом и членом Научно-консультативного совета Сосея Гептареша. Всем остальным авторам нечего раскрывать.
Благодарности
Мы благодарим профессора д-ра Гебхарда Ф. Х. Шертлера за его долгосрочную поддержку в этом проекте, д-ра Роджера Дж.П. Доусона и Хоффмана Ла Роша за поддержку в культуре клеток. Эта работа была организована Швейцарским национальным научным фондом (гранты 210030-153145 и 310030B_173335 GFXS), а также финансированием CGT от Европейского исследовательского совета (EMPSI, 339995) и Совета медицинских исследований (MRC U105197215). FP признает ETH Цюрих через Национальный центр компетенций в области исследований молекулярных ультрабыстрых науки и техники (NCCR MUST) и ETH Femtosecond и Аттосекунда науки и техники (ETH FAST) программ. FP, JM, AB и CJT признают долгосрочную финансовую поддержку со стороны Института Пола Шеррера.
Материалы
| Name | Company | Catalog Number | Comments |
| 1D4 peptide | Peptide2.0 | Under request | |
| 9-cis retinal | Sigma-Aldrich | R5754 | |
| Autosampler A-900 | GE Healthcare | Discontinued | |
| C9G | Anatrace | N324 | |
| cOmplete, EDTA-free protease inhibitor coctail | Roche | 5056489001 | |
| Cymal-5 | Anatrace | C325 | |
| Cymal-5NG | Anatrace | NG325 | |
| Cymal-6 | Anatrace | C326 | |
| Cymal-6NG | Anatrace | NG326 | |
| DDM | Anatrace | D310 | |
| DM | Anatrace | D322 | |
| DMNG | Anatrace | NG322 | |
| Econo column | Bio-Rad | 7372512 | |
| Ettan LC | GE Healthcare | Discontinued | |
| FRAC-950 | GE Healthcare | Discontinued | |
| HPLC Water 2795 Separation Module | Waters AG | 720000358EN | |
| InstantBlue Protein Stain | Expedeon | ISB1L | |
| LCT Premier mass spectrometer (ESI-TOF) | Waters AG | - | |
| LMNG | Anatrace | NG310 | |
| Monitor UV-900 | GE Healthcare | 18110835 | |
| Nanodrop 1000 | Witec AG/ThermoFisher | Discontinued | |
| NuPAGE 4-12% Bis-Tris gel 1.0 mm, 15 well | ThermoFisher | NP0323BOX | |
| NuPAGE MES SDS buffer (20x) | ThermoFisher | NP0002 | |
| OGNG | Anatrace | NG311 | |
| PAGEr Minigel Chamber | Lonza | 59905 | |
| Reprosil 200 C18-AQ column | Morvay Analytik GmbH | #s1503 | |
| Superdex 200 Increase GL column | GE Healthcare | 28990944 | |
| Tabletop centrifuge 5424R | Eppendorf | 5404000413 | |
| Ultracentrifuge Optima XE-100 | Beckmann Coulter | A94516 | |
| ULTRA-TURRAX T25 | IKA WERKE | 0003725003 | |
| UV-VIS spectrophotometer | Shimadzu | UV-2401PC | |
| Waters 2487 Dual λ Absorbance Detector | Waters AG | - |
Ссылки
- Tate, C. G. Practical considerations of membrane protein instability during purification and crystallisation. Methods in Molecular Biology. 601, 187-203 (2010).
- Lebon, G., Bennett, K., Jazayeri, A., Tate, C. G. Thermostabilisation of an agonist-bound conformation of the human adenosine A(2A) receptor. Journal of Molecular Biology. 409 (3), 298-310 (2011).
- Deupi, X., et al. Stabilized G protein binding site in the structure of constitutively active metarhodopsin-II. Proceedings of the National Academy of Sciences. 109 (1), 119-124 (2012).
- Tsai, C. -. J., et al. Crystal structure of rhodopsin in complex with a mini-G o sheds light on the principles of G protein selectivity. Science Advances. 4 (9), (2018).
- Carpenter, B., Tate, C. G. Engineering a minimal G protein to facilitate crystallisation of G protein-coupled receptors in their active conformation. Protein Engineering Design and Selection. 29 (12), 583-594 (2016).
- Chae, P. S., et al. Maltose-neopentyl glycol (MNG) amphiphiles for solubilization, stabilization and crystallization of membrane proteins. Nature Methods. 7 (12), 1003-1008 (2010).
- Loll, P. J. Membrane proteins, detergents and crystals: what is the state of the art. Acta Crystallographica Section F Structural Biology Communications. 70 (12), 1576-1583 (2014).
- Chae, P. S., et al. Glucose-neopentyl glycol (GNG) amphiphiles for membrane protein study. Chemical communications. 49 (23), 2287-2289 (2013).
- Standfuss, J., Xie, G., Edwards, P. C., Burghammer, M., Oprian, D. D., Schertler, G. F. X. Crystal structure of a thermally stable rhodopsin mutant. Journal of Molecular Biology. 372 (5), 1179-1188 (2007).
- Kaushal, S., Ridge, K. D., Khorana, H. G. Structure and function in rhodopsin: the role of asparagine-linked glycosylation. Proceedings of the National Academy of Sciences of the United States of America. 91 (9), 4024-4028 (1994).
- Molday, L. L., Molday, R. S. 1D4: a versatile epitope tag for the purification and characterization of expressed membrane and soluble proteins. Methods in Molecular Biology. 1177 (604), 1-15 (2014).
- Carpenter, B., Tate, C. G. Expression and Purification of Mini G Proteins from Escherichia coli. Bio-Protocol. 7 (8), (2017).
- Grueninger-Leitch, F., D'Arcy, A., D'Arcy, B., Chène, C. Deglycosylation of proteins for crystallization using recombinant fusion protein glycosidases. Protein Science. 5 (12), 2617-2622 (1996).
- Rasmussen, S. G. F., et al. Crystal structure of the β2 adrenergic receptor-Gs protein complex. Nature. 477 (7366), 549-555 (2011).
- Loginova, M. Y., Rostovtseva, Y. V., Feldman, T. B., Ostrovsky, M. A. Light damaging action of all-trans-retinal and its derivatives on rhodopsin molecules in the photoreceptor membrane. Biochemistry (Moscow). 73 (2), 130-138 (2008).
- Alexandrov, A. I., Mileni, M., Chien, E. Y. T., Hanson, M. A., Stevens, R. C. Microscale Fluorescent Thermal Stability Assay for Membrane Proteins. Structure. 16 (3), 351-359 (2008).
- Sonoda, Y., et al. Benchmarking Membrane Protein Detergent Stability for Improving Throughput of High-Resolution X-ray Structures. Structure. 19 (1), 17-25 (2011).
- Maeda, S., et al. Crystallization scale preparation of a stable GPCR signaling complex between constitutively active rhodopsin and G-protein. PloS One. 9 (6), 98714 (2014).
- Boute, N., Jockers, R., Issad, T. The use of resonance energy transfer in high-throughput screening: BRET versus FRET. Trends in Pharmacological Sciences. 23 (8), 351-354 (2002).
- Singhal, A., Guo, Y., Matkovic, M., Schertler, G., Deupi, X., Yan, E. C. Y. Structural role of the T 94 I rhodopsin mutation in congenital stationary night blindness. EMBO Report. 17 (10), 1-10 (2016).
- Choe, H. -. W., et al. Crystal structure of metarhodopsin II. Nature. 471 (7340), 651-655 (2011).
- Mattle, D., et al. Ligand channel in pharmacologically stabilized rhodopsin. Proceedings of the National Academy of Sciences of the United States of America. 115 (14), 3640-3645 (2018).
- Okada, T., Fujiyoshi, Y., Silow, M., Navarro, J., Landau, E. M., Shichida, Y. Functional role of internal water molecules in rhodopsin revealed by X-ray crystallography. Proceedings of the National Academy of Sciences of the United States of America. 99 (9), 5982-5987 (2002).
- Blankenship, E., Vahedi-Faridi, A., Lodowski, D. T. The High-Resolution Structure of Activated Opsin Reveals a Conserved Solvent Network in the Transmembrane Region Essential for Activation. Structure. 23 (12), 2358-2364 (2015).
- Magnani, F., et al. A mutagenesis and screening strategy to generate optimally thermostabilized membrane proteins for structural studies. Nature Protocols. 11 (8), 1554-1571 (2016).
- Kawate, T., Gouaux, E. Fluorescence-detection size-exclusion chromatography for precrystallization screening of integral membrane proteins. Structure. 14 (4), 673-681 (2006).
- Standfuss, J., et al. The structural basis of agonist-induced activation in constitutively active rhodopsin. Nature. 471 (7340), 656-660 (2011).
- Singhal, A., et al. Insights into congenital stationary night blindness based on the structure of G90D rhodopsin. EMBO reports. 14 (6), 520-526 (2013).
- Carpenter, B., Nehmé, R., Warne, T., Leslie, A. G. W., Tate, C. G. Structure of the adenosine A(2A) receptor bound to an engineered G protein. Nature. 536 (7614), 104-107 (2016).
Перепечатки и разрешения
Запросить разрешение на использование текста или рисунков этого JoVE статьи
Запросить разрешениеThis article has been published
Video Coming Soon
Авторские права © 2025 MyJoVE Corporation. Все права защищены