
Method Article
Мукоцилиальные эпителиальные органоиды из эмбриональных клеток Ксенопы: Поколение, Культура и живая визуализация высокого разрешения
В этой статье
Резюме
Мы описываем простой протокол для разработки мукоцилитарных эпителиальных органоидов из глубоких эктодермальных клеток, изолированных от эмбрионов Xenopus laevis. Мультипотентные прародители регенерируют прекурсоры эпителиальных клеток кубков и позволяют в реальном времени отслеживать инициирование и прогрессирование клеточных переходов на поверхности органоидов.
Аннотация
Мукоцилиарный эпителий обеспечивает первую линию обороны путем удаления иностранных частиц через действие производства слизи и реснички опосредованного зазора. Многие клинически значимые дефекты слизистой оболочки эпителия выведены, поскольку они происходят глубоко в организме. Здесь мы представляем уротворяемую 3D-модель для мукококилиарного эпителия, генерируемого из мультипотентных прародителей, которые были микрохирургически изолированы от эмбрионов Xenopus laevis. Слизистые эпителиальные органоиды покрыты недавно сгенерированным эпителием из глубоких эктодермальных клеток, а затем украшены различными узорчатыми многоцилионированными клетками, секреторными клетками и слизистыми клетками, которые неотличимы от родного эпидермиса в пределах 24 ч. Полные последовательности динамических клеточных переходов от мезенхимальных к эпителиальным, которые возникают на апической поверхности органоидов, можно отследить с помощью живой визуализации высокого разрешения. Эти в пробирке культурных, самоорганизовых слизистых эпителиальных органоидов предлагают явные преимущества в изучении биологии слизистой эпителия с высокой эффективностью в генерации, определенные условия культуры, контроль над числом и размером, и прямой доступ для живой визуализации во время регенерации дифференцированного эпителия.
Введение
Травмы, инфекции и болезни слизистой оболочки эпителия связаны с нарушением производства и очистки слизи, которая часто встречается при легочныхрасстройствах,таких как хроническая обструктивная болезнь легких, астма, муковисцидоз, бронхиэктаз, и первичной цилиарнойдискинезии 1,2,3,4. Недавнее продвижение в органоидной технологии, например, базально-клеточной выведенной органоид легких называется трахеосфера, которая recapitulates регенерации слизистой эпителия возникают в качестве перспективной модели стерапевтическим потенциалом 1,5,6. Однако его использование в настоящее время ограничено, отчасти из-за отсутствия определенных культурных условий и низкой эффективности в органоидных производствах. Мукоцилийный эпителий в дыхательных путих человека и эпидермис лягушки удивительно похожи по морфологии тканей, клеточному составу иего функции 7,8,9,10,11,12. В обоих организмах мукокоцилярный эпителий обеспечивает защиту первой линии, высвеняя слизь и противомикробные вещества и очищает вредные частицы и патогенные микроорганизмы путем синхронного действия ресничок.
Здесь мы описываем простой протокол для генерации мукокоцилевых эпителиальных органоидов с использованием мультипотентных прародителей эмбрионов Xenopus laevis 13,14. Ранее мы сообщали14 о том, что при отсутствии экзогенных факторов роста и внеклеточной матрицы микрохирургически изолированные глубокие клетки из ранней гаструловой стадии эктодерма спонтанно собираются в агрегаты, регенерируют эпителий на его поверхности и созревают в слизистую эпителий путем интеркаляции многоцилионных и других соточенных клеток в пределах 24 ч. В дополнение к быстрому развитию, этот протокол предлагает различные возможности для непосредственного доступа к переходам мультипотентных глубоких эктодермальных клеток в эпителиальных прародителей, которые резюмируют шаги регенерации нарушенногоэпителия 14, которые не доступны из нетронутых эмбрионов и эктодермов (также известный как животное крышка)15,16,17. Количество и размер производимых органоидов масштабируемы с высокой эффективностью, контролируя стартовые материалы эмбрионов Xenopus. Органоиды в плавающей культуре могут быть легко отсортированы и переданы на желаемом этапе для дальнейшего анализа, включая визуализацию с высоким разрешением, механическое тестирование, лечение наркотиками и генетическуюхарактеристику 14. Эта спонтанная, тканевая механика, обусловленная регенерацией эпителия на поверхности эмбриональных клеточных агрегатов, приводит к мукокоцилиальным эпителиальным органоидам и обеспечивает новую трехмерную (3D) модель для изучения биологии слизистой эпителия.
протокол
Протоколы использования животных и экспериментальные протоколы были утверждены институциональным комитетом по уходу за животными (IACUC) Института фундаментальных наук (IBS 18-01) и Корейским передовым институтом науки и техники (KA2017-22).
1. Эмбрионы
- Получить X. laevis эмбрионов с помощью стандартной процедуры: вручную собирать яйца из стимулированных самок лягушек и выполнять экстракорпоральноеоплодотворение 18,19.
- Де-желе оплодотворенных эмбрионов с нежным возбуждением около 5 мин в 2% цистеина в 1/3x модифицированный солевой Барта (MBS; см. рецепт 1X MBS ниже) при рН 819.
- Дополнительный шаг для живой визуализации: флуоресцентно маркировать специфические белки и наблюдать их динамику в органоиде, переходить к разделу 5.
- (По желанию) Для мониторинга загрязнения поверхностных эктодермальных клеток в органоидах, этикетка апической поверхности эмбрионов с NHS-родамин на стадии 914. Инкубация эмбрионов в 1 мг/мл NHS-родамина в 1/3x MBS (рН 9,0) в течение 30 мин с нежным гайкой. Вымойте эмбрионы три раза, переведя в чашку Петри заполненную 1/3x MBS в течение 15 минут.
- Культура эмбриона в 1/3x MBS при предпочтительной температуре (14-26 градусов по Цельсию) до тех пор, пока не будут обнаружены первые признаки 10-й стадии (т.е. появление темных пигментированных клеток вокруг блатопора на вегетативном представлении).
2. Подготовка микрохирургических инструментов, решений и культурных сосудов
- Подготовь инструменты, необходимые в том числе пару хирургических стиппов класса и волос инструменты (петля волос и нож для волос) длямикрохирургии 20.
- Подготовка следующих средств культуры для эмбрионов: 1/3X MBS, где 1X MBS сделан с NaCl (88 mM), KCl (1 mM), NaHCO3 (2.4 mM), MgSO4 (0.82 mM), Ca(No3)2 (0.33 mM), CaCl2 (0.41 mM), и HEPES (10 mM). Отрегулируйте рН до 7,4 с 10 M NaOH.
ПРИМЕЧАНИЕ: Дополнительно: добавить капли фенола красный, чтобы указать рН. - Подготовь следующие средства культуры для эмбриональных тканей и органоидов: Данильчик для Эми (DFA)21 дополнен свежим 1% антибиотиком и антимикотическим раствором. Подготовка DFA с NaCl (53 мМ), Na2CO3 (5 мМ), глюконат калия (4,5 мМ), глюконат натрия (32 мМ), CaCl2 (1 мМ) и MgSO4 (1 мМ). Отрегулируйте рН до 8,3 с гранулированным бицином. Фильтр DFA (фильтр с бутылкой 0,2 мкм), aliquot и хранить его при -20 градусов по Цельсию.
- Подготовка кальция и магния, свободной DFA для отделения глубоких клеток от эктодерма с помощью рецепта выше и опуская CaCl2 и MgSO4. Aliquot и хранить при -20 градусов по Цельсию.
- Подготовка неклейвых ПЦР труб для агрегации эмбриональных клеток.
- Чтобы вызвать спонтанную агрегацию изолированных эмбриональных клеток, подготовьте неклейвые ПЦР трубки, покрывая трубы ПЦР круглым дном при температуре 200 МКЛ 1% BSA (1 г BSA в 100 мл дистиллированной воды) на ночь при температуре 4 градусов по Цельсию или 2 ч при комнатной температуре. Каждая трубка будет использоваться для сборки одного органоида.
- Промыть BSA покрытием ПЦР трубки с DFA три раза, чтобы удалить любые остаточные BSA.
- Заполните ПЦР трубки с 200 йл DFA.
3. Изоляция глубоких эктодермальных клеток
- Выберите и соберите эмбрионы по мере их достижения на ранней стадии 10 с помощью инструментов для волос под стереоскопом.
- Используя одноразовый переносной пипетки, перенесите отобранные эмбрионы в заполненную DFA чашку Петри.
- Удалите вителлиновую мембрану эмбрионов с помощью острых типсов с вегетативной стороны, не нарушая сторону эмбриона.
ПРИМЕЧАНИЕ: Будьте осторожны, чтобы избежать разоблачения эмбрионов в воздухе. Введение пузырьков воздуха в раствор или вывод эмбрионов на поверхность приведет к взрыву эмбриона. - Чтобы изолировать крышку животного, распоить животную сторону эмбриона вверх.
- Визуально оцените размер крышки животного, которая будет вырезана, и сделайте первый разрез вдоль края крышки животного с помощью ножа для волос. Вытяните нож для волос наружу, чтобы сделать разрез.
- Повторите шаг 3.5, чтобы создать цепочку небольших порезов, чтобы акцизную крышку животного.
- Обрезать толстый слой края крышки животного с помощью ножа для волос, чтобы предотвратить включение мезодерм прекурсоров.
ПРИМЕЧАНИЕ: Чтобы предотвратить заживление и агрегацию изолированных животных шапки, перейти к следующему шагу в течение 10 минут. Как правило, мы изолируют 5-10 шапки животных в то время, чтобы собрать несколько органоидов. - Чтобы отделить глубокие эктодермальные клетки от крышки животного, перенесите вырезанные колпачки животных в чашку Петри, наполненную без кальция и магния DFA с одноразовой трубкой передачи. Будьте осторожны, чтобы не ввести пузырьки воздуха во время передачи.
- Чтобы сохранить достаточно места для следующих шагов, используя инструменты для волос, положение животных шапки лицом к животному стороне вверх и поддерживать щедрое расстояние от других explants.
- Подождите 5-10 минут, а затем контролировать explants под стереоскопом. После того, как ослабленные глубокие клетки были найдены от края темного пигментного поверхностного слоя, начать поднимать поверхностный слой от светлых глубоких клеток эктодерма с помощью волосяного ножа под стереоскопом.
- Аккуратно отсоедините (отсоедините) поверхностный слой ножом для волос, начиная с края.
- Соберите глубокие эктодермальные клетки с минимальным устремлением (10-u201215 L), чтобы ограничить количество без кальция и магния DFA, которое передается в агрегации средств массовой информации на следующем этапе.
ПРИМЕЧАНИЕ: Отдельные поверхностные клетки могут быть удалены из средств массовой информации, чтобы предотвратить загрязнение оставшихся глубоких клеток эктодерма.
4. Поколение мукоцилиальных эпителиальных органоидов
- Передача собранных глубоких эктодермальных клеток в неклейвую ПЦР-трубку, содержащую 200 МКЛ DFA. Аккуратно пипетка средств массовой информации (2'u20123 раз) для разгона переданных ячеек в трубке ПЦР.
ПРИМЕЧАНИЕ: Сроки, обозначенные как часы после агрегирования (hpa) начинается с этого шага. Размер полученных органоидов контролируется количеством глубоких эктодермальных клеток, добавленных в ПЦР-трубку. Глубокие эктодермальные клетки из одного или более колпачков животных могут быть использованы, в зависимости от желаемого размера органоида. - Закройте трубку ПЦР. Держите ПЦР трубки вертикально, чтобы вызвать спонтанную агрегацию в нижней части.
- Мониторинг процесса агрегации под стереоскопом. Клетки обычно собираются в нижней части трубки ПЦР в течение часа и собираются в сферические агрегаты в пределах 2-3 ч, в зависимости от размера.
- Для проведения живой визуализации или тестирования на наркотики во время разработки мукокоцилиальных эпителиальных органоидов, собирать агрегаты на 2 л.с. с помощью трубы 200 л, оснащенной увеличенными наконечниками (нарезать стерилизованными ножницами), чтобы избежать повреждения агрегатов во время сбора.
- Чтобы агрегаты развивались в слизистую эпителиальный органоид в культуре, собирайте сферические агрегаты из трубки ПЦР при 5 л.с. и перенесите их в заполненную DFA чашку Петри.
- Распоить агрегаты далеко от других, чтобы предотвратить их слияние. В пределах 24 ч культуры при комнатной температуре без каких-либо дополнительных факторов, зрелые слизистые эпителиальные органоиды могут наблюдаться, чтобы вращаться с действием избиения ресничок, покрывающих поверхность дифференцированного эпителия
5. (Необязательно) высокое разрешение живой визуализации развивающихся органоидов
- Подготовка мРНК для микроинъекции.
- Чтобы визуализировать эпителиализацию, которая происходит на начальной стадии органоидного образования, подготовить мРНК для эпителиальной специфической зонулы occludens белка-1 (ЗО-1) и для излагания клеточных мембран путем усиления pCS2-ЗО1-RFP и pCS2-mem-GFP плазмидов (подарок от Лэнса Дэвидсона).
- Извлекайте и вышифровывайте плазмидную ДНК, а затем транскрибируйте ограниченную мРНК с помощью комплекта транскрипции SP6/T7 in vitro.
- Aliquot транскрибируется мРНК и хранить его при -80 градусов по Цельсию.
- Микроинъекционная мРНК в оплодотворенный эмбрион
- Поместите оплодотворенные эмбрионы в 3% Ficoll в 1x MBS.
- Загрузите 3-4 МКЛ мРНК с помощью наконечника микрозагрузщика в вытянутую стеклянную иглу (длинный и тонкий конические кончики иглы с внутренним диаметром 10-201230 мкм).
- Прикрепите иглу к микроинъектору и отрегулируйте время и давление, чтобы обеспечить постоянный объем мРНК для микроинъекции.
- Ввимите мРНК прямо под апическую поверхность полюса животного. Различные бледно-цветные круговые патч, который вызван расширением коры видна во время микроинъекции.
- Перенесите вводимые эмбрионы на 1/3X MBS и свяжете их до стадии 9.5.
- Соберите флуоресцентно помеченные эмбрионы под стереоскопом флуоресценции (настройки возбуждения/выброса для GFP (488/510) и RFP (532/588)).
- Продолжить шаг 1.3.
- Соберите и культуру органоидов (разделы 3 и 4) до желаемой стадии развития.
- Выполните живую визуализацию.
- Подготовьте камеру визуализации со стеклянным дном, приклеив крышку стекла к специально измельченной акриловой камере с использованием кремниевой смазки.
ПРИМЕЧАНИЕ: Плотно запечатать камеру, чтобы предотвратить утечку культурных средств массовой информации. - Заполните камеру визуализации DFA.
- Выберите одну шестиугольную электронную микроскопию передачи (TEM) сетки (75 сетки) из контейнера с помощью типсов и применить небольшое количество жира к краю сетки.
ПРИМЕЧАНИЕ: Размер сетки должен быть меньше диаметра агрегата, так что агрегат находится на сетке. - Нажмите вниз слегка, чтобы обеспечить сетку TEM в нижней части камеры изображения.
- Перенесите агрегаты в камеру визуализации и распоите их в сетке.
ПРИМЕЧАНИЕ: Избегайте позиционирования агрегатов рядом с смазкой. На протяжении всего эксперимента агрегаты должны сидеть на сетке TEM, не контактируя с нижней частью камеры, чтобы предотвратить физическое сжатие. - Заполните камеру изображения с DFA и запечатать его с крышкой стекла и жира.
ПРИМЕЧАНИЕ: Камера должна быть герметичной без пузырьков воздуха, чтобы предотвратить турбулентность или движение во время изображения. - Чтобы следить за прогрессированием слизистой оболочки эпителиального органоидного образования, соберите замедленное z-стек изображения агрегатов (от 2 л.с.) с помощью конфокального микроскопа.
ПРИМЕЧАНИЕ: Мы обычно собираем z-стеки толщиной 120 мкм каждые 15 минут, используя цель 20X, чтобы следить за динамическим поведением клеток, но эти спецификации должны быть оптимизированы для целей экспериментов.
- Подготовьте камеру визуализации со стеклянным дном, приклеив крышку стекла к специально измельченной акриловой камере с использованием кремниевой смазки.
6. (Необязательно) Изображение развивающихся органоидов путем фиксации и иммуностимулирования
- Исправить органоиды на желаемой стадии развития, передав их в стеклянный флакон, наполненный фиксаторным раствором.
ПРИМЕЧАНИЕ: Добавьте объем фиксативного решения, который в 20 раз больше, чем у образцов, чтобы обеспечить полную фиксацию. Выполните следующие процессы на nutator, если не указано иное. В целом, органоиды фиксируются с 4% параформальдегида (PFA) в PBS. Тем не менее, различные фиксаторы могут потребоваться для обнаружения конкретных белков. Например, мы использовали 4% PFA с 0,25% глутаралдегида в PBS для обнаружения F-актина и ацетилированного тубулина. Для обнаружения инсектина (ИТЛН) и ЗО-1 органоиды фиксируются раствором ледяного Дента (4:1 метанол: диметилсульфоксид) на ночь при -20 градусов по Цельсию. Фиксированные органоиды Дента должны быть серийно обезвожены перед стиркой (шаг 6.3). Длительность инкубации и мытья антител может быть оптимизирована для конкретных нужд. - Исправить органоиды в течение 15 мин при комнатной температуре (RT) или на ночь при температуре 4 градусов по Цельсию.
- Вымойте 3 раза с PBST (PBS с 0,1% Тритон X-100) в течение 15 минут на RT.
- Блок неспецифической привязки с 10% козьей сыворотки в PBST (PBSGT) в течение 1 ч на RT.
- Инкубация с первичными антителами (1:200) в PBSGT ночь при 4 градусов по Цельсию.
- Вымойте 3 раза с PBST в течение 15 минут на RT.
- Инкубация вторичными антителами (1:200) в PBSGT ночь при 4 градусах Цельсия.
- Вымойте 3 раза с PBST в течение 15 минут на RT.
- Перенесите фиксированные и иммунотелированные органоиды в камеру визуализации и приступайте к конфокальные изображения.
Результаты
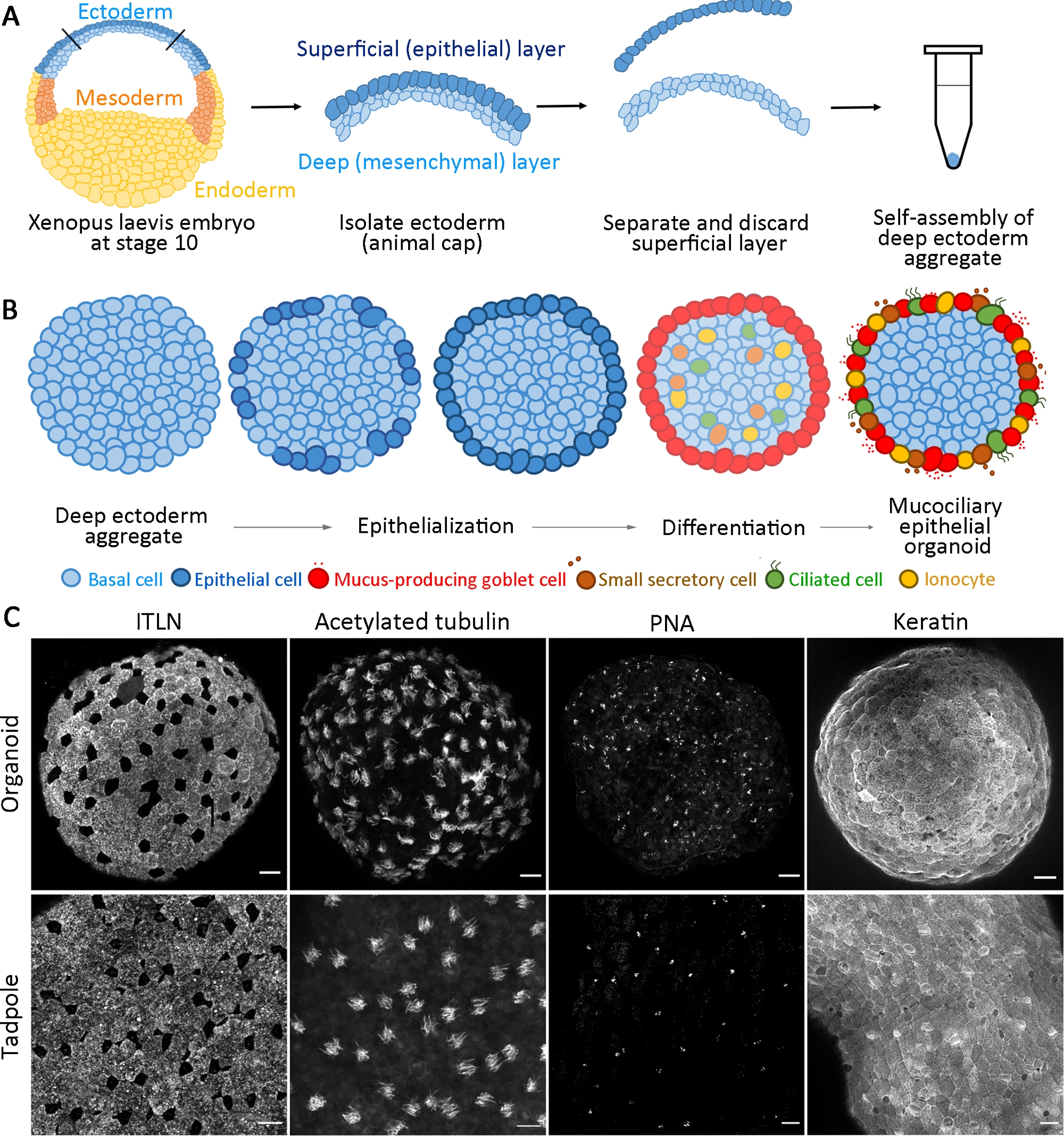
Этот стандартизированный протокол генерирует слизистую эпителиальный органоид из мультипотентных прародителей, изолированных от ранней гаструловой стадии X. laevis эмбрионов в пределах 24 ч откультивирования 14. Собранные глубокие эктодермальные клетки самосборляться, чтобы сформировать агрегат в неклейной трубке ПЦР и пройти поверхностную эпителиализацию и дифференциацию клеток кубков. Недавно эпителиализированная поверхность агрегатов обеспечивает субстрат, похожий на родной эпителий, найденный в виво для интеркаляции внутренних клеток (например, многоцилионированных и других аксессуаров) и развивается, чтобы сформировать мукоцилиарные эпителиальные органоиды(рисунок 1A,B). В течение 24 ч после агрегации самоорганизованные слизистые эпителиальные органоиды регенерируют зрелый эпидермис, неотличимый от эпидермиса головастика. Органоиды состоят из полностью дифференцированного эпителия (кератина), слизи- секретированных клеток кубок (ITLN), мультицилиалированных клеток (ацетилированный тубулин), и небольшие секреторные клетки (арахис агглютинин, PNA) (Рисунок 1C).
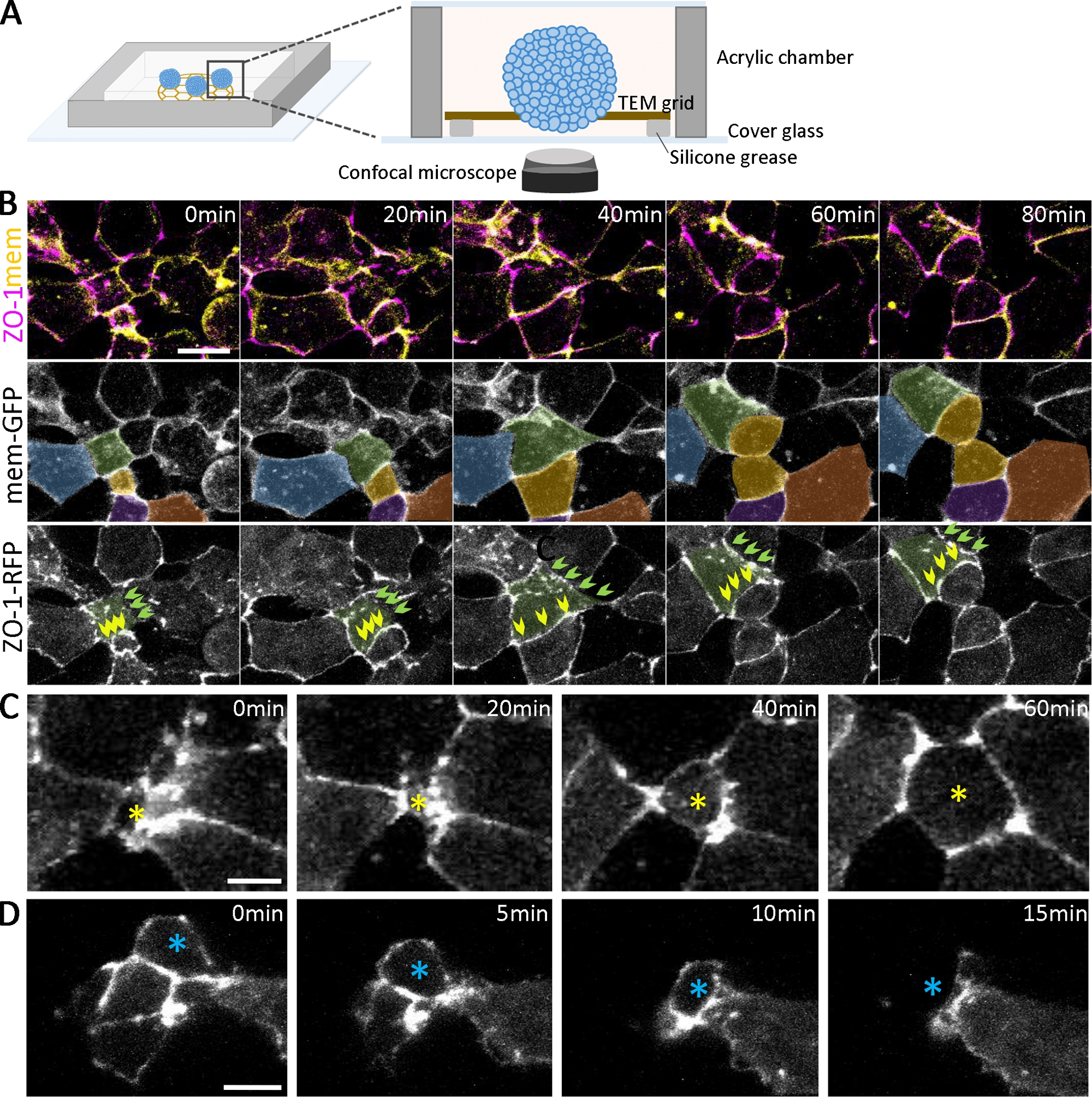
Помимо подтверждения развития различных типов клеток с иммуностимулированием, за динамикой развития органоидов может последовать живая визуализация(рисунок 2A). Для изучения эпителиализации, которая возникает на ранней стадии органоидногообразования (рисунок 1B),мы помечены эмбрионов, выражая флуоресцентно помечены жесткой соединения белков (ЗО-1-RFP) и мембраны локализации белков (mem-GFP). При двойной маркировке последовательные шаги образования плотных соединений ЗО-1 могут быть отмечены и количественно проанализированы во время эпителиализации(рисунок 2). Например, для клеток(рисунок 2B,зеленого цвета) на разных стадиях эпителиализации (на 0 мин), некоторые области клеточной адгезии разбросаны puncta ЗО-1(рисунок 2B, зеленые стрелки). В отличие от этого, другие области полностью собраны, смежные выражения ЗО-1(рисунок 2B, желтые стрелки). Со временем панкта сливаются и соединяются, чтобы сформировать смежные плотные соединения(рисунок 2B, зеленыестрелки), а смежные плотные соединения поддерживают свою морфологию даже во время деления клеток(рисунок 2B, желтые стрелки). По мере созревания плотных соединений клетки динамически перемещаются в поверхность и из нее вдоль апических плоскостей органоидов(рисунок 2C,D). Кроме того, путем отслеживания клеток spatiotemporally на поверхности дифференцируя органоидов(рисунок 2B, цветные клетки), многомасштабный анализ возможен, начиная от отдельных puncta смежных жесткие соединения, клеточно-клеточные границы, и подмножества популяций клеток в органоидах.

Рисунок 1: Поколение мукоцилиальных эпителиальных органоидов.
(A)Схема, показывающая протокол для сборки глубоких эктодермальных агрегатов из эмбрионов X. laevis. (B)Схема для модели слизистой оболочки эпителиального органоидного образования, возникают из многопотентных глубоких эктодермальных клеток (поперечный вид). Клетки, расположенные на поверхности, переходят в эпителиальные клетки и дифференцируются в клетки кубков. Дифференциация цилиированных клеток, секреторных клеток и ионоцитов радиально интеркалируется в поверхность и регенерирует зрелый эпидермис. (C)Максимальная z-проекция слизистой оболочки эпителия иммунотензия для ITLN (слизисто-продуцированные клетки кубок), ацетилированный тубулин (цилиалированные клетки), PNA (малые секретонные клетки), и кератин (эпителиальные клетки) в органоидах на 24 hpa (верхняя панель) и tadpole epidermis (нижняя панель). Шкала бар 30 мкм. Пожалуйста, нажмите здесь, чтобы просмотреть большую версию этой цифры.
{kind=link}

Рисунок 2: Живая визуализация развивающихся органоидов.
(A)Схема камеры визуализации живых органоидов (не для масштабирования). (B)Временных последовательностей конфокальных стеков, собранных из глубоких эктодермальных клеточных агрегатов, выражают ЗО-1-RFP и mem-GFP от 2,5 л.с. Шкала бар 20 мкм. Клетки псевдо-цветные для отслеживания с течением времени. Клетки зеленого цвета имеют различные статусы адгезии клеток, в том числе один постепенно развивающийся положительный адгезии ЗО-1 (зеленые стрелки) и один поддержания смежных ЗО-1 положительные адгезии (желтые стрелки) с течением времени. (C, D) Покадровые конфокальные изображения глубокого эктодермального клеточного агрегата ЗО-1-RFP показывают радиально интеркалирующие клетки, движущиеся к поверхности (C, желтая звезда) и движущиеся внутри агрегатов (D, голубая звезда). Шкала баров 10 мкм. Пожалуйста, нажмите здесь, чтобы просмотреть большую версию этой цифры.
{kind=link}
Обсуждение
Мукоцилиальные эпителиальные органоиды, генерируемые из глубоких эктодермальных клеток эмбриона X. laevis, являются мощным инструментом для изучения эпителиализации и дифференциации мультипотентных прародителей in vitro. В отличие от широко принятого животного колпачкаанализ 16 используется для in vitroорганогенез 13 и развитие слизистой эпителии15 , 17,22, которые используют нетронутыми эктодерм, глубокие эктодерм полученных органоидов, представленных в этом протоколе предлагают явную возможность контролировать ткани механики управляемых фаз регенерацииповерхностного эпителия 14. Примерно на 2 л.с., вновь генерируемых ЗО-1 положительные эпителиальныеклетки (рисунок 2) начинают появляться на апической поверхности органоидов и увеличить их популяцию, чтобы покрыть весь органоид, как ткань затвердевать или снижаетсоответствие 14. Регенерация эпителия и последующие спецификации линии для слизистых клеток кубок проходят спонтанно в химически определенных культурных средств массовой информации в течение дня. Эти быстро развивающиеся слизистые эпителиальные органоиды обеспечивают платформу для изучения динамического поведения клеток в режиме реального времени, в высоком разрешении, во время прогрессивных этапов эпителиальной регенерации. Они также позволяют изутовить фундаментальные вопросы, возникающие при развитии мукокоциарной эпителия, гомеостаза и связанных с нимзаболеваний2,9,23. В частности, механическая чувствительность глубоких клеток-предшественников во время перехода к эпителиальным прекурсорам клеток кубков,выявленным в органоидах 14, может служить для связи респираторных заболеваний, связанных с аномальной базальной дифференциацией, где слизисто-секретные клетки кубок являются чрезмерно- или недостаточнопроизведенными 23.
Хотя этот протокол предлагает простой подход к генерации этих органоидов, Есть несколько важных шагов для успеха в экспериментах. Чтобы предотвратить загрязнение поверхностных эпителиальных клеток во время изоляции глубоких эктодермальных клеток от крышки животного, следует следить за крышкой животного, помещенной в DFA без кальция и магния, под стереоскопом и обнаруживать нужное время, чтобы инициировать разделение темно-пигментного поверхностного слоя крышки животного. Если ткань хранится в кальция и магния, свободной DFA слишком долго, вся ткань будет диссоциировать и различение между глубокими и поверхностными клетками будет невозможно для глубоких эктодермовых агрегатов. Чтобы подтвердить отсутствие поверхностных клеток в глубоких эктодермовых агрегатах, мы рекомендуем флуоресцентно маркировать апиакальной поверхность эмбриона с помощью NHS-родамина (шаг 1.414) до микрохирургии; это позволило бы легко идентифицировать поверхностные клетки, если они существуют в результате органоидов. Так как эпителиальная регенерация регулируется механикойтканей 14,важно, чтобы избежать непреднамеренного генерации силы для самоорганизовых органоидов. В частности, мы предлагаем избегать контакта со стеклянным дном камеры визуализации во время живой визуализации, размещая агрегаты по краям сеток TEM, так как это позволяет свободно контактировать с окном изображения живых агрегатов (шаг 5.1.2.). Этот in vitro- культурная,самоорганизованная 3D-модель для мукокоциального эпителия будет служить в качестве уротворяемого инструмента для ответа на фундаментальные вопросы, возникающие во время регенерации эпителия и спецификации линии клеток кубок.
Раскрытие информации
Авторов нечего раскрывать.
Благодарности
Мы благодарим сотрудников лаборатории Ким и Лэнса Дэвидсона за их комментарии и поддержку. Эта работа была поддержана стипендией молодого ученого в HYK из Института фундаментальных наук (IBS-R0250Y1).
Материалы
| Name | Company | Catalog Number | Comments |
| Equipment | |||
| Dual-stage Glass Micropipette Puller | Narishige | PC-100 | |
| Picoliter microinjector | Warner Instruments | PLI-100A | |
| Confocal Laser Microscope | |||
| Stereoscope | |||
| Tools | |||
| Forcep | Dumont | Dumont #5 | |
| Hair knife | Reference (Kay, B.K.; Peng, H.B., 1991) | ||
| Hair loop | Reference (Kay, B.K.; Peng, H.B., 1991) | ||
| hCG injection | |||
| human chorionic gonadotropin | Sigma | cg10-10vl | |
| MBS solution | |||
| 10M Sodium hydroxide | Sigma | 72068 | |
| Calcium chloride | Sigma | C3881 | |
| Calcium nitrate | Sigma | C1396 | |
| HEPES | Sigma | H4034 | |
| Magnesium sulfate | Sigma | 230391 | |
| Phenol-red | Sigma | P0290 | |
| Potassium chloride | Sigma | 7447-40-7 | |
| Sodium bicarbonate | Sigma | S6014 | |
| Sodium chloride | Sigma | S9625 | |
| Sodium hydroxide reagent grade, 97%, powder-25g | Sigma | 655104 | |
| dejellying solution | |||
| L-Cysteine hydrochloride monohydrate | Sigma | C7880 | |
| Sodium hydroxide 10M | Sigma | 72068 | |
| Ficoll solution | |||
| Ficoll | Sigma | F4375 | |
| DFA solution | |||
| Sodium chloride | Sigma | S9625 | |
| 0.22mm Filter | Millipore | S2GPT05RE | |
| Antibiotic Antimycotic Solution | Sigma | A5955 | |
| Bicine | Sigma | B3876 | |
| Calcium chloride | Sigma | C3881 | |
| Magnesium sulfate | Sigma | 230391 | |
| Potassium gluconate | Sigma | G4500 | |
| Sodium carbonate | Sigma | 222321 | |
| Sodium gluconate | Sigma | G9005 | |
| mRNA in vitro transcription | |||
| SP6/T7 in vitro transcription kit | Invitrogen | AM1340 | |
| mRNA microinjection | |||
| Borosilicate glass capillary tubes | Harvard Apparatus | GC100-10 | |
| Eppendorf microloader pipette tips | ThermoFisher | A25547 | |
| Mineral oil | Sigma | M5904 | |
| PCR tube coating | |||
| BSA | Thermofisher | 26140079 | |
| PCR tubes | SSI | SSI-3245-00 | |
| Imaging | |||
| Custom-milled acrylic chamber | |||
| Coverglass 24mmX50mm | Duran | B01_001650 | |
| SPI Hexagonal TEM Grids, Gilded Nickel (50mesh) | SPI | 275HGN-XA | |
| SPI Hexagonal TEM Grids, Gilded Nickel (75mesh) | SPI | 2775GN-XA | |
| Silicone grease | Shinetsu | HIVAC-G | |
| Fixation | |||
| 20ml screw top-cap vial | Wheaton | WH.986580 | |
| 2ml screw top-cap vial | |||
| Benzyl alcohol | Sigma | 305197 | |
| Benzyl benzoate | Sigma | B6630 | |
| Dimethyl sulfoxide (DMSO) | Sgima | D4540 | |
| Glutaraldehyde 10% EM GRADE | Electron Microscopy Sciences | 16120 | |
| Goat serum | Jackson | 005-000-121 | |
| Methanol | Sigma | 322415 | |
| Paraforlamdehyde | Sigma | P6148 | |
| Phosphate-buffered saline (PBS) | LPS Solution | CBP007B | |
| Triton X-100 | Sigma | T8787 | |
| Primary antibody (1:200) | |||
| acetylated tubulin | Sigma | clone 6-11B-1 | |
| Itln1 | Proteintech | 11770-1-AP | |
| Keratin | Developmental Studies Hybridoma Bank | 1h5 | |
| ZO1 | Invitrogen | 402200 | |
| Vectors | |||
| pCS2-mem-GFP | Gift from Dr. Lance Davidson | ||
| pCS2-ZO1-RFP | Gift from Dr. Lance Davidson |
Ссылки
- Barkauskas, C. E., et al. Lung organoids: current uses and future promise. Development. 144 (6), 986-997 (2017).
- Puchelle, E., Zahm, J. M., Tournier, J. M., Coraux, C. Airway Epithelial Repair, Regeneration, and Remodeling after Injury in Chronic Obstructive Pulmonary Disease. Proceedings of the American Thoracic Society. 3 (8), 726-733 (2006).
- Tilley, A. E., Walters, M. S., Shaykhiev, R., Crystal, R. G. Cilia dysfunction in lung disease. Annual Review of Physiology. 77, 379-406 (2015).
- Vareille, M., Kieninger, E., Edwards, M. R., Regamey, N. The Airway Epithelium: Soldier in the Fight against Respiratory Viruses. Clinical Microbiology Reviews. 24 (1), 210-229 (2011).
- Rock, J. R., et al. Basal cells as stem cells of the mouse trachea and human airway epithelium. Proceedings of the National Academy of Sciences of the United States of America. 106 (31), 12771-12775 (2009).
- Sachs, N., et al. Long-term expanding human airway organoids for disease modeling. The EMBO Journal. 38 (4), 100300(2019).
- Dubaissi, E., et al. A secretory cell type develops alongside multiciliated cells, ionocytes and goblet cells, and provides a protective, anti-infective function in the frog embryonic mucociliary epidermis. Development. 141 (7), 1514-1525 (2014).
- Hayes, J. M., et al. Identification of novel ciliogenesis factors using a new in vivo model for mucociliary epithelial development. Developmental Biology. 312 (1), 115-130 (2007).
- Walentek, P., Quigley, I. K. What we can learn from a tadpole about ciliopathies and airway diseases: Using systems biology in Xenopus to study cilia and mucociliary epithelia. Genesis. 55 (1-2), (2017).
- Werner, M. E., Mitchell, B. J. Understanding ciliated epithelia: The power of Xenopus. Genesis. 50 (3), 176-185 (2012).
- Dubaissi, E., Papalopulu, N. Embryonic frog epidermis: a model for the study of cell-cell interactions in the development of mucociliary disease. Disease Models & Mechanisms. 4 (2), 179-192 (2011).
- Walentek, P., et al. A novel serotonin-secreting cell type regulates ciliary motility in the mucociliary epidermis of Xenopus tadpoles. Development. 141 (7), 1526-1533 (2014).
- Asashima, M., et al. In vitro organogenesis from undifferentiated cells in Xenopus. Developmental Dynamics. 238 (6), 1309-1320 (2009).
- Kim, H. Y., Jackson, T. R., Stuckenholz, C., Davidson, L. A. Tissue mechanics drives regeneration of a mucociliated epidermis on the surface of Xenopus embryonic aggregates. Nature Communications. 11 (1), 665(2020).
- Haas, M., et al. DeltaN-Tp63 Mediates Wnt/beta-Catenin-Induced Inhibition of Differentiation in Basal Stem Cells of Mucociliary Epithelia. Cell Reports. 28 (13), 3338-3352 (2019).
- Green, J. Molecular Methods in Developmental Biology: Xenopus and Zebrafish. Guille, M. , Humana Press. 1-13 (1999).
- Stubbs, J. L., Davidson, L., Keller, R., Kintner, C. Radial intercalation of ciliated cells during Xenopus skin development. Development. 133 (13), 2507-2515 (2006).
- Nieuwkoop, P. D., Faber, J. Normal table of Xenopus laevis (Daudin) : a systematical and chronological survey of the development from the fertilized egg till the end of metamorphosis. , Garland Pub. (1994).
- Sive, H. L., Grainger, R. M., Harland, R. M. Early development of Xenopus laevis : a laboratory manual. , Cold Spring Harbor Laboratory Press. (2000).
- Joshi, S. D., Kim, H. Y., Davidson, L. A. Microscopy tools for quantifying developmental dynamics in Xenopus embryos. Methods in Molecular Biology. 917, 477-493 (2012).
- Sater, A. K., Steinhardt, R. A., Keller, R. Induction of neuronal differentiation by planar signals in Xenopus embryos. Developmental Dynamics. 197 (4), 268-280 (1993).
- Sedzinski, J., Hannezo, E., Tu, F., Biro, M., Wallingford, J. B. Emergence of an Apical Epithelial Cell Surface In Vivo. Developmental Cell. 36 (1), 24-35 (2016).
- Rock, J. R., Randell, S. H., Hogan, B. L. M. Airway basal stem cells: a perspective on their roles in epithelial homeostasis and remodeling. Disease Models & Mechanisms. 3 (9-10), 545-556 (2010).
Перепечатки и разрешения
Запросить разрешение на использование текста или рисунков этого JoVE статьи
Запросить разрешениеСмотреть дополнительные статьи
This article has been published
Video Coming Soon
Авторские права © 2025 MyJoVE Corporation. Все права защищены