
Method Article
Один анализ культуры миофибры для оценки функциональности взрослых мышечных стволовых клеток Ex Vivo
В этой статье
Резюме
В этом протоколе описан метод культивирования и функционального анализа мышечных стволовых клеток in vitro, который сохраняет большую часть их взаимодействий с их эндогенной нишей.
Аннотация
Взрослая скелетная мышечная ткань содержит популяцию стволовых клеток, которая необходима для ее способности к регенерации. При повреждении мышц мышечные стволовые клетки выходят из состояния покоя и активируют миогенную программу, что в конечном итоге приводит к восстановлению поврежденных тканей, сопутствующему пополнению пула мышечных стволовых клеток. Различные факторы влияют на активность мышечных стволовых клеток, среди них внутренние стимулы, а также сигналы из прямой среды мышечных стволовых клеток, ниши стволовых клеток. Выделение и культивирование одиночных миофиберов с связанными с ними мышечными стволовыми клетками сохраняет большую часть взаимодействия стволовой клетки с ее нишей и, следовательно, является ближайшей возможностью для изучения функциональности мышечных стволовых клеток ex vivo. Здесь представлен протокол выделения, культивирования, трансфекции siRNA и иммуноокрашивания мышечных стволовых клеток на их соответствующих миофибрах из мышц мыши EDL(extensor digitorum longus). Экспериментальные условия, изложенные здесь, позволяют изучать и манипулировать мышечными стволовыми клетками ex vivo, включая исследование миогенной активности без врожденной необходимости в экспериментах in vivo на животных.
Введение
Скелетные мышцы у взрослых представляют собой постмитотическую ткань, в основном состоящую из многоядерных миофиберов, которые являются эффекторными клетками для произвольных движений. Он обладает замечательной способностью к регенерации, процесс, который напоминает эмбриональный миогенез и подвергается нарушениям в возрасте и болезни1. Эта поразительная регенеративная способность скелетных мышц зависит от мышечных стволовых клеток (MuSCs), которые также называются сателлитными клетками из-за их расположения между сарколеммой и базальной пластинкой миофиберов2,3. В условиях покоя MUSC находятся в состоянии покоя и характеризуются экспрессией транскрипционного фактора Pax7 и маркерами покоя, такими как Sprouty14,5,6,7,8. При активации, например, после травмы, MuSC покидают состояние покоя и повышают миогенный регуляторный фактор MyoD9. Двойные положительные MuSC Pax7/MyoD пролиферируют и дифференцируются, тем самым генерируя миогенные клетки-предшественники, которые также часто называют миобластами. Затем эти миобласты далее дифференцируются в удлиненные миоциты, процесс, совпадающий с молекулярными и морфологическими изменениями, например, потерей Pax7 и повышением регуляции экспрессии миогенина10. Затем миоциты в конечном итоге сливаются друг с другом или с существующими миофибрами, тем самым восстанавливая поврежденную ткань. Важно отметить, что небольшая часть мышечных стволовых клеток возвращает повышение регуляции MyoD и способна самообновляться11. Состояние дифференцировки MuSC и миогенной прогрессии можно легко наблюдать при исследовании миогенных маркеров, таких как Pax7, MyoD и Миогенин10.
Культура одиночных миофиберов с их смежными MuSC является отличным методом для исследования функциональности MuSC в условиях ex vivo, поскольку MuSC остаются в своей эндогенной нише12,13. Поведение MuSC регулируется внутренними сигналами, а также внешними сигналами, предоставляемыми нишей, специализированным анатомическим местом, включающим компоненты внеклеточного матрикса (ECM), окружающего MuSC, и само миофибра. Например, одним из внешних регуляторов покоя MuSC является сигнализация Notch. Здесь сигнальные сигналы принимаются MuSC как от миофибра, так и от ECM14,15,16. Кроме того, ниша MuSC важна для управления осью деления MuSC, тем самым регулируя судьбу дочерних клеток MuSC17,18. Разумно, что такие параметры, как асимметричные деления MuSC, миогенная прогрессия и самообновление, могут быть однозначно оценены в этой экспериментальной установке. Например, многоклеточный кластер может образовываться из одного MuSC после 72-часового периода культивирования, который может быть исследован на предмет возникновения и процента различных миогенных популяций, таких как самообновляющиеся, пролиферирующие и далее дифференцированные MuSC8,19,20,21. Состояние дифференцировки MuSC может быть определено путем исследования экспрессии/коэкспрессии Pax7, MyoD и миогенина. После 72 ч культуры клетки в кластере могут быть дискриминированы по следующим параметрам: только клетки Pax7 являются самообновляющимися MuSC, в то время как Pax7 / MyoD двойные положительные клетки пролиферируют / активируют MuSC, а дальнейшие дифференцированные миогенные клетки являютсямиогенин-положительными 22. Кроме того, числа MuSC или повторное вхождение в клеточный цикл/активация могут быть исследованы в дополнение к миогенной прогрессии, например, с помощью иммунофлуоресцентного анализа на основе параметров, описанных выше.
Здесь описаны уникальные особенности протокола выделения и культивирования миофибры, например, сохранение взаимодействия MuSC с его нишей. Мышиные целые мышцы EDL(extensor digitorum longus)тщательно рассекаются, перевариваются коллагеназой и физически тритурируются для получения одиночных миофиберов с их ассоциированными MuSC для дальнейшего культивирования. Кроме того, протокол разграничивает шаги по трансфекции MuSC с siRNA для функционального анализа генов-кандидатов и последовательных анализов на основе иммунофлуоресценции без необходимости трансгенных животных.
протокол
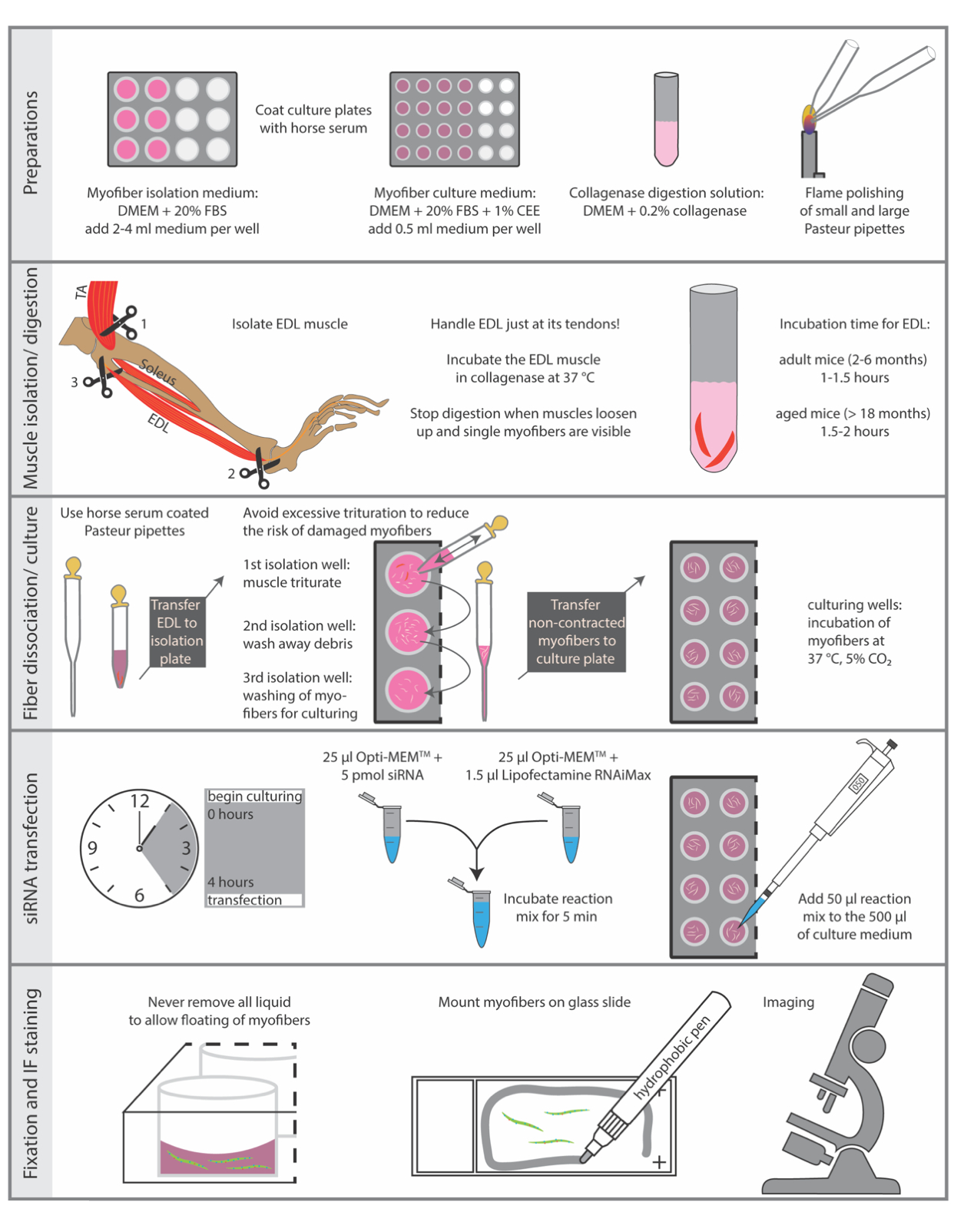
Жертвоприношение животных должно осуществляться в соответствии с национальными правилами проведения экспериментов на животных. Протокол, описанный здесь, был выполнен в соответствии с руководящими принципами Института Лейбница по проблемам старения - Института Фрица Липмана и директивой Европейского союза (ЕС) 2010/63/EU (номер лицензии на извлечение органов: O_JvM_18-20). Основные этапы протокола обобщены на рисунке 1.
1. Подготовка культуральных пластин, сред и пипеток Пастера
ПРИМЕЧАНИЕ: Все материалы и оборудование, необходимые для выделения и культивирования отдельных миофибров, должны быть как можно более стерильными. Поэтому рекомендуется выделять одиночные миофибры под полустерильным рассеченным капюшоном.
- Используйте стерильный HS (конская сыворотка) для покрытия тканевых культуральных пластин. Покрытие предотвращает прикрепление одиночных миофибров к пластиковой поверхности. Для каждой мыши требуется 4 скважины из 12 скважин для изоляции и специальное количество скважин из 24-луночной плиты для культивирования. Инкубируйте колодцы из 12-луночной плиты с 1 мл и колодцы из 24-луночной плиты с 0,5 мл HS в течение 5 мин при RT (комнатная температура), затем удалите HS и дайте плитам высохнуть еще 5 мин.
ПРИМЕЧАНИЕ: HS может быть собран и повторно использован для целей нанесения покрытия несколько раз, если его сохранить стерильным. - Подготовьте изоляционную среду миофибры, дополнив DMEM (модифицированную среду Dulbecco Eagle с 4,5 г / л глюкозы и пирувата натрия) 20% FBS (фетальная бычья сыворотка), фильтруйте через фильтр 0,22 мкм. Добавьте среду к предварительно покрытым изоляционным пластинам (12-луночная пластина, 2-4 мл изоляционной среды на скважину) примерно за 30 мин до изоляции и уравновешивайте в увлажненном инкубаторе с температурой 37 °C с 5% CO2.
- Приготовьте культуральную среду миофибры, добавив DMEM (модифицированную среду Dulbecco Eagle's с 4,5 г / л глюкозы и пирувата натрия) с 20% FBS (фетальная бычья сыворотка) и 1% экстракт куриного эмбриона, фильтруйте через фильтр 0,22 мкм. Добавляют 0,5 мл среды в каждую лунку предварительно покрытых культуральных пластин (24-луночная пластина) примерно за 30 мин до выделения и уравновешивают в увлажненном инкубаторе при 37 °C с 5% CO2.
- Приготовьте раствор для переваривания коллагеназы путем растворения 0,2% (мас./об.) коллагеназы типа 1 (из Clostridium histolyticum)в DMEM (модифицированная среда Dulbecco Eagle's с 4,5 г/л глюкозы и пирувата натрия), фильтр через фильтр 0,22 мкм. Для двух мышц EDL(extensor digitorum longus)используют 2,5 мл раствора коллагеназы в стерильной реакционной трубке 15 мл. Кроме того, предварительно нагрейте раствор ~10 мин на циркуляционной водяной бане при 37 °C перед началом выделения.
- Приготовьте стерильные пипетки Пастера для тритурации переваренных коллагеназой мышц. Используйте алмазную ручку для резки пипеток Пастера.
- Для каждой мыши используют одну большую отверстие пипетки с отверстием около 0,3 см и длиной около 10-12 см и вторую стеклянную пипетку с небольшим отверстием около 0,1 см и длиной примерно 22 см(рисунок 2F).
- Сгладьте края обеих пипеток путем тепловой полировки пламенем горелки Бунзена. Держите наконечник пипетки в течение 5-10 с в пламя легким движением, чтобы обеспечить равномерное распределение тепла до тех пор, пока острые края стекла не сгладятся.
- Непосредственно перед использованием покройте оба типа стеклянных пипеток стерильным HS, заполнив всю пипетку ~ 2 мл HS в течение 5 мин, после чего выбросьте HS и дайте пипеткам высохнуть в течение 5 минут при RT.
2. Выделение мышц EDL и переваривание коллагеназы
- Опрыскивайте все оборудование 70% этанолом, чтобы избежать загрязнения.
- Приносите в жертву мышь в соответствии с национальными правилами экспериментов на животных.
- Опрыскивайте задние конечности мыши 70% этанолом. Используйте закаленные тонкие изогнутые ножницы (режущая кромка 24 мм) и тонкие щипцы (Dumont 7, изогнутые или прямые), чтобы удалить кожу и обнажить нижележащие мышцы. Избегайте любого контакта нижележащих мышц с мехом (увеличивает риск загрязнения).
- Удалите окружающие фасции тонкими изогнутыми щипцами, не повреждая нижележащие мышцы(рисунок 2A). Закройте щипцы, чтобы избежать изгиба.
- Используйте изогнутые щипцы для обнажения дистальных сухожилий ТА(большеберцовой кости передней)и EDL мышцы. Чтобы удалить ТА, захватите дистальное сухожилие ТА щипцами и вырежьте тонкими пружинными ножницами Vannas (режущая кромка 5 мм, диаметр наконечника 0,35 мм). Удерживая ТА на сухожилии, потяните его к колену и разрежьте мышцу близко к колену(рисунок 2B),мышца EDL теперь обнажается.
ПРИМЕЧАНИЕ: Убедитесь, что на этом шаге захватывается только сухожилие TA, иначе основной EDL может быть поврежден. При отрезании мышцы ТА убедитесь, что сухожилия в колене можно легко увидеть впоследствии. - Поднимите дистальное сухожилие EDL тонкими изогнутыми щипцами и вырежьте тонкими пружинными ножницами Vannas(рисунок 2C). Обнажите проксимальное сухожилие EDL, осторожно потянув EDL к колену. Вырежьте проксимальное сухожилие тонкими пружинными ножницами Vannas. Переместите мышцу EDL на 37 °C предварительно нагретых 2,5 мл раствора для переваривания коллагеназы в реакционной трубке объемом 15 мл со стадии 1,4. (Рисунок 2D).
ПРИМЕЧАНИЕ: Сделайте небольшой разрез на внешней соединительной ткани колена, чтобы полностью обнажить проксимальное сухожилие EDL. Убедитесь, что вы только захватываете сухожилия и не растягивайте EDL слишком сильно. - Повторите шаги 2.3. до 2.6. со вторым EDL. Добавьте обе мышцы EDL в одну и ту же реакционную трубку объемом 15 мл, заполненную раствором для переваривания коллагеназы 2,5 мл.
- Инкубируют мышцы EDL в реакционной трубке при 37 °C на плавающей водяной бане.
ПРИМЕЧАНИЕ: Время инкубации зависит от нескольких факторов, таких как активность коллагеназы, возраст мыши и количество фиброзной ткани. Типичное время инкубации для мышц EDL взрослых мышей (2-6 месяцев) составляет 1-1,5 ч, а для пожилых мышей (18-месячный возраст) 1,5-2 ч. - Чтобы избежать чрезмерного переваривания мышц, проверяйте мышцы во время пищеварения. Остановите пищеварение, когда мышцы ослабнут и видны одиночные миофибры(рисунок 2E). Аккуратно перенесите мышцы с помощью пипетки с большим отверстием в первую лунку предварительно выровненной 12-луночной пластины, содержащей изоляционную средумиофибры (рисунок 2G).
3. Диссоциация и культура миофибры
- Для следующих шагов используйте стереобинокулярный микроскоп, предпочтительно оснащенный нагревательной пластиной (37 °C). Используйте пипетку с большим отверстием, чтобы промыть мышцы теплой изоляционной средой до тех пор, пока не высвободятся одиночные миофибры. Диссоциируйте мышцы с помощью пипетки с большим отверстием до тех пор, пока желаемое количество миофиберов не будет свободно плавать в растворе.
ПРИМЕЧАНИЕ: Избегайте чрезмерной тритурации, чтобы снизить риск повреждения миофибров. Настоятельно рекомендуется использовать нагревательную пластину для диссоциации миофибры, так как температура падает во время процесса выделения, что приведет к гибели миофибры. - Перенос неконтактных миофибров(рисунок 2H)с покрытой HS стеклянной пипеткой с малым отверстием во вторую скважину, заполненную изоляционной средой для смывания мусора и законсервированных (поврежденных) миофибров(рисунок 2I).
ПРИМЕЧАНИЕ: Чтобы избежать чрезмерного движения изолированных миофибров, их можно перенести во вторую лунку, а затем продолжить процесс тритурации. - Перенесите незаконтрактованные миофибры в следующую(3-ю)хорошо заполненную изоляционной средой для повторной промывки.
- Используйте стеклянную пипетку с малым отверстием, покрытую HS, для переноса примерно 50-100 незаконтактных миофибер на 24-луночную пластину, содержащую питательную среду миофибры.
- Инкубируйте миофибры при 37 °C, 5% CO2 в течение выделенного времени (рекомендуется максимум 96 ч).
ПРИМЕЧАНИЕ: MuSC делят один раз либо плоский, либо апикально-базальный через 42 ч культуры. Кроме того, после 72 ч культуры MuSC образуют многоклеточные кластеры, состоящие из самообновляющихся, пролиферирующих или совершенных (дифференцированных) MuSC.
4. трансфекция siRNA
- Через 4 ч после выделения миофибры трансфектируют миофибр-ассоциированные MuSCs (50-100 незаконтактованных миофибер в одной лунке 24-луночной пластины, заполненной 500 мкл культуральной среды) трансфекционным реагентом на основе липидов, например, RNAiMAX по протоколу производителя с конечной концентрацией 5 пмол siRNA. Поэтому добавляют 25 мкл Opti-MEM с соответствующим объемом siRNA к 25 μL Opti-MEM, содержащим 1,5 мкл трансфекционного реагента. Инкубируют реакционную смесь в течение 5 мин и добавляют ее затем к 500 мкл питательной среды.
ПРИМЕЧАНИЕ: Вторая трансфекция через 24 ч или 48 ч рекомендуется для более длительных периодов культивирования, например, более 48 ч. Смена среды после трансфекции не требуется.
5. Фиксация и окрашивание ПЧ
- Для иммуноокрашивания используют стерео бинокулярный микроскоп. Все объемы на следующих этапах регулируются на одну скважину из 24-луночной плиты. Выполните все следующие действия с помощью пипетки с покрытием HS.
- Осторожно отбросьте культуральную среду миофибры, оставив в лунке раствор (примерно 100 мкл на 24 лунку). Сделайте это для всех дальнейших шагов, если не указано иное, чтобы разрешить плавание миофибров. Добавьте 500 мкл 2% PFA для фиксации миофибров соседними с ними MuSC, инкубируйте в течение 5 мин при комнатной температуре (RT).
- Осторожно удалите супернатант и трижды промойте миофибры PBS (рН 7,4, 500 мкл в течение 5 мин при РТ каждый).
- Добавляют 500 мкл пермеабилизационного раствора (0,1% тритона Х-100, 0,1 М глицина в PBS, рН 7,4), инкубируют в течение 10 мин при РТ.
- Удалить раствор для пермеабилизации и добавить 500 мкл блокирующего раствора (5% HS в PBS, pH 7,4) в течение 1 ч при RT.
ПРИМЕЧАНИЕ: Проверьте рекомендуемый блокирующий раствор для первичных антител, чтобы избежать неспецифического связывания. - Удалите блокирующий раствор и добавьте 300 мкл первичного разведения антител (например, анти-Pax7 (PAX7, DSHB, неразбавленный), анти-MyoD (клон G-1, Santa Cruz, 1:200)) на лунку. Инкубировать при 4 °C в течение ночи.
- Промыть три раза с 500 мкл PBS на лунку (5 мин при RT).
- Удалить PBS и добавить 300 мкл вторичного разведения антител (например, антимышечного IgG1-546 и антимышечного IgG2b-488 1:1000) на лунку. Инкубировать 1 ч на RT, защищенном от света. Для следующих шагов рекомендуются условия пониженного освещения.
- Промыть два раза с 500 мкл PBS на лунку (5 мин при RT).
- Выполняют окрашивание DAPI с использованием 500 мкл раствора на лунку (конечная концентрация DAPI 10 мкг/мл) в течение 5 мин при РТ.
- Промыть два раза с 500 мкл PBS на лунку (5 мин при RT).
- Используйте гидрофобную ручку, чтобы нарисовать круг на микроскопическом стеклянном слайде, чтобы создать водоотталкивающий барьер, который предотвратит разлив жидкости, содержащей миофибры. Перенесите миофибры в минимально возможном объеме на микроскопический стеклянный слайд и рассейте их на стеклянной горке.
ПРИМЕЧАНИЕ: Избегайте физического вытягивания миофибров на микроскопический стеклянный слайд, так как это может вызвать трение, приводящее к отделению кластеров от миофибров. - Удалите остаточную жидкость с помощью пипетки Пастера с небольшим отверстием или пипетки 200 мкл.
- Используйте две капли водной монтажной среды и накройте миофибры крышкой. Дайте слайдам высохнуть в течение времени, рекомендованного производителем. Храните слайды при температуре 4 °C в темноте.
Результаты
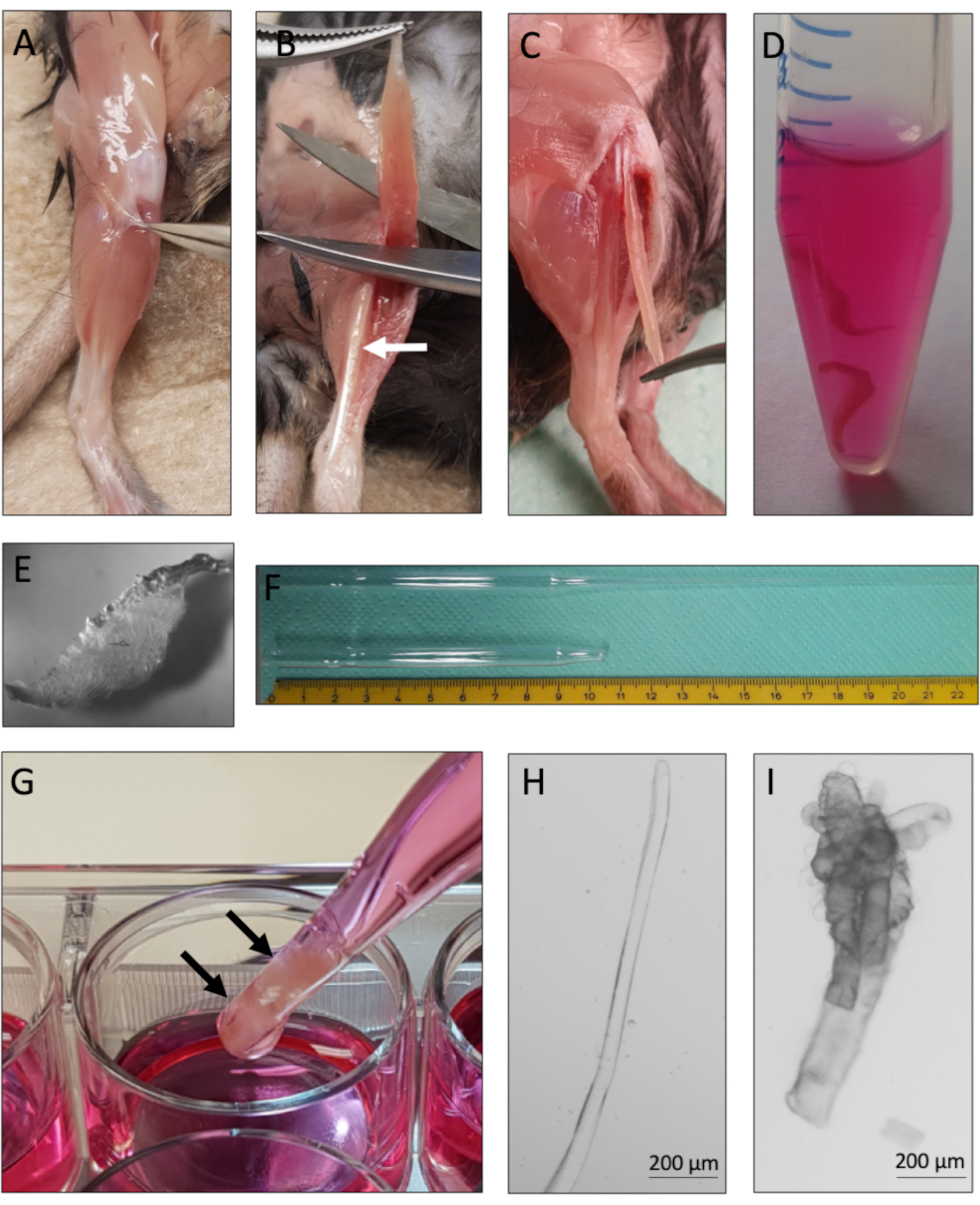
Этот протокол предоставляет инструкции для успешного получения и культивирования одиночных миофибров с их ассоциированными MuSC из мышиных мышц EDL. Основные этапы протокола обобщены на рисунке 1. Тщательное рассечение мышц EDL от сухожилия к сухожилию(рисунок 2A-C)имеет решающее значение для высокого выхода жизнеспособных миофиберов. Диссоциация мышц достигается сначала путем переваривания коллагеназы(рисунок 2D),затем физической тритурации(рисунок 2G). Интактные миофибры(рисунок 2H)культивируются, тогда как гиперконтрактные и мертвые миофибры(рисунок 2I)должны быть исключены из культуры и анализа.
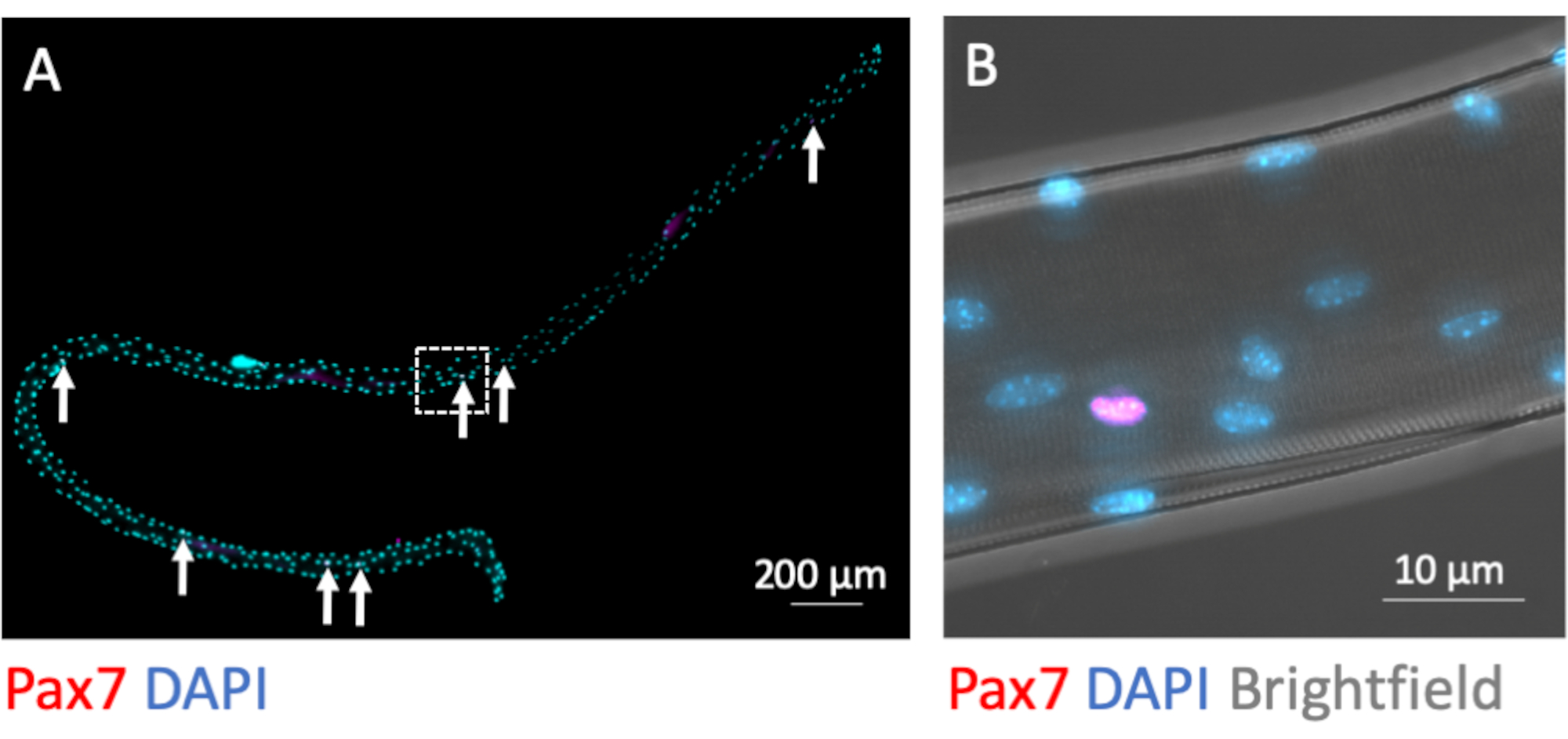
MuSC остаются связанными с миофибером во время процесса изоляции. Иммунофлуоресцентное окрашивание для транскрипционного фактора Pax7 идентифицирует и отличает 7-9 ядер MuSC от множества мионуклеев на миофибру при фиксации непосредственно после завершения процесса выделения(рисунок 3A). На фиг.3В показана увеличенная область из Фиг.3А с дополнительным каналом яркого поля, который обнажает субклеточную миофибриллярную структуру миотрубы и демонстрирует сигнал иммунофлуоресценции Pax7 в ядре MuSC.
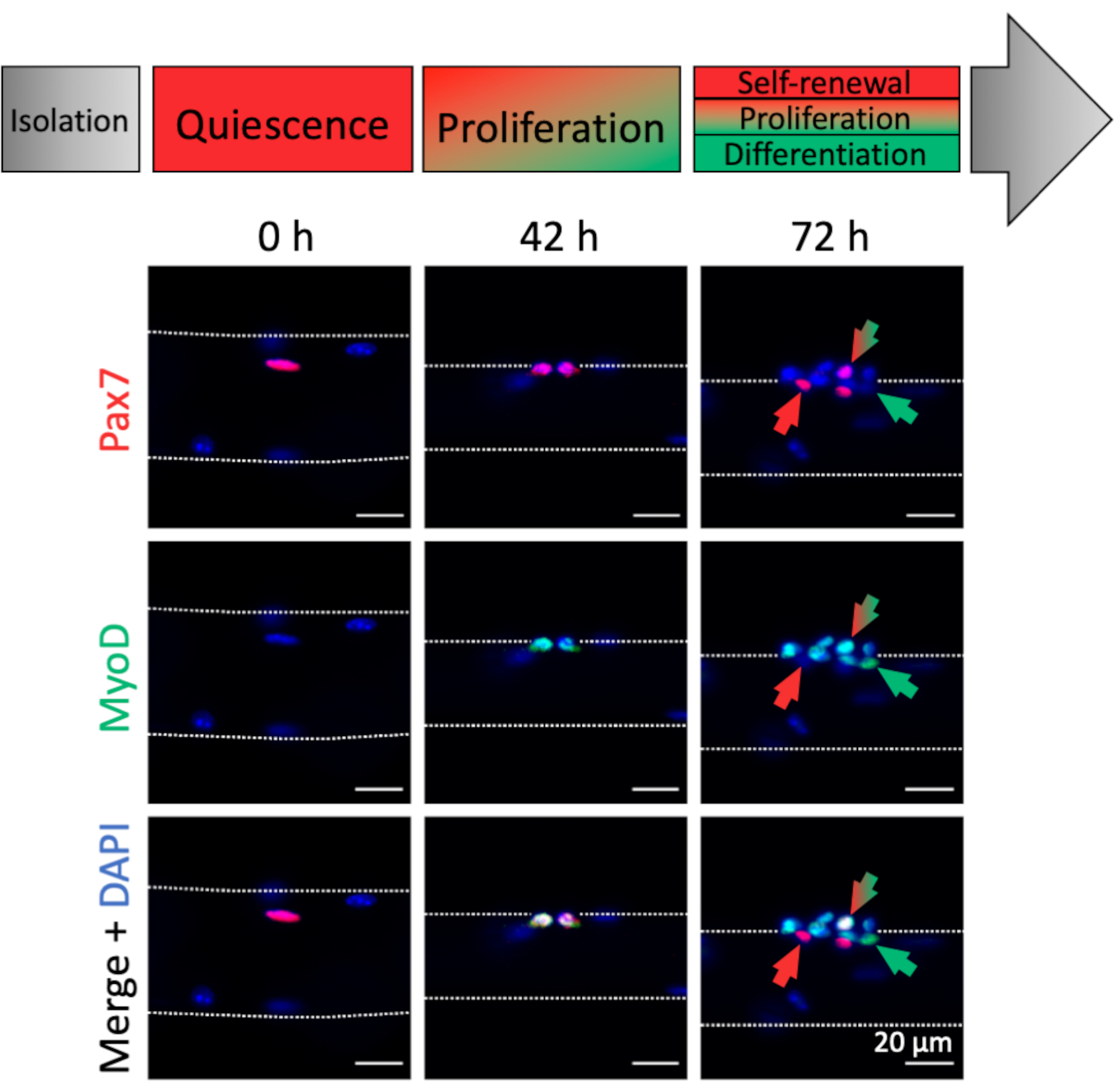
Активация и миогенная прогрессия миофибр-ассоциированных MuSCs могут быть проанализированы по экспрессии миогенных маркеров. Связанные со свежеизолированными миофибрами (0 ч), в основном покоятся MUSC, которые характеризуются экспрессией Pax7 и отсутствием экспрессии MyoD, тем самым напоминая in vivo гомеостатическое состояние(рисунок 4,0 ч). Благодаря процедуре рассечения/диссоциации и составу среды культивирования миофибры, MuSC быстро активируют и повышают регуляцию транскрипционного фактора MyoD для облегчения пролиферации, что можно наблюдать через 42 часа, когда MuSC подверглись их первому делению(Рисунок 4,42 ч). После 72 часов культивирования MuSC образуют кластеры потомств с различными миогенными судьбами, что сопровождается паттернами экспрессии различных миогенных маркеров(рисунок 4,72 ч). Только клетки Pax7+ сопротивляются дифференцировке и становятся самообновляющимися стволовыми клетками. Двойные положительные клетки Pax7+ и MyoD+ являются пролиферативными, тогда как клетки только MyoD+ продвинулись вдоль миогенной линии и будут дифференцироваться.
Система культивирования миофибры позволяет эффективно интерференцировать активность MuSC различными вмешательствами, одним из которых является трансфекция siRNA, как подробно описано в этом протоколе. Для мониторинга эффективности трансфекции миофибровых ассоциированных MUSCs была трансфектирована флуоресцентно меченая нецелевая siRNA. Pax7-положительные MuSC аккумулировали цитоплазматическую siRNA гранулоподобным образом, что указывает на эффективное поглощение(рисунок 5A). В цитоплазме миофиберов не наблюдалось флуоресцентных гранул, что свидетельствует о естественном барьере поглощения в миофибрах через 4 ч после выделения и о том, что трансфекция siRNA специфически нацелена на MuSC. Количественная оценка положительно трансфектированных клеток Pax7+ на миофибру показала, что более половины всех MuSC поглощали видимые количества флуоресцентно меченой siRNA непосредственно перед завершением первого раунда деления через 24 часа. Количество трансфектированных клеток Pax7+ увеличилось до 74% через 30 часов(рисунок 5B). Кроме того, не было никакой разницы в количестве клеток Pax7+ на миофибр трансфектированных или нетрансфектированных состояний в обеих временных точках, демонстрируя отсутствие неблагоприятного воздействия на количество стволовых клеток из-за процедуры трансфекции(рисунок 5C).
Таким образом, протокол предоставляет подробное описание выделения и культивирования одиночных миофибров EDL с прилегающими к ним мышечными стволовыми клетками. Это позволяет изучать активность мышечных стволовых клеток ex vivo, связанных с миофибрами, например, путем иммунофлуоресцентного анализа. Манипуляции мышечными стволовыми клетками путем трансфекции siRNA эффективны и обеспечивают методологическую основу для функционального анализа.

Рисунок 1:Схематическая сводка основных шагов. Основные этапы процедуры изоляции и иммуноразрашивания обобщены на этом рисунке. Сокращения: DMEM, модифицированный орлиный медиум Дульбекко; FBS, Фетальная бычья сыворотка; CEE, экстракт куриного эмбриона; ТА, большеберцовая кпереди; EDL, extensor digitorum longus Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
{kind=link}

Рисунок 2:Рассечение EDL мыши и выделение одного миофьбера. (A) Кожа задней конечности мыши удаляется, а мышца, окружающая фасцию, оттягивается, чтобы обнажить ТА(большеберцовую переднюю)мышцу. (B) Мышца TA удаляется, чтобы обнажить мышцу EDL(extensor digitorum longus),отмеченную белой стрелкой. (C) Мышца EDL рассекается путем разрезания сухожилий. (D) Две мышцы EDL переваривают коллагеназу. (E) Появление переваренной коллагеназой мышцы с видимыми одиночными миофибрами, ослабляющимися из ядра ткани. (F) Пипетка Пастера с большим и малым отверстием с масштабом. (G) Мышцы EDL (отмеченные черными стрелками) физически тритурируются с помощью пипетки Пастера с большим отверстием. (H) Одиночные неповрежденные миофибры тонкие и блестящие и могут быть индивидуально собраны для культуры и анализа. (I)Гиперконтрактные и мертвые миофибры непригодны для культуры и анализа. Микроскопические изображения 2H и 2I были получены с помощью микроскопа с использованием объектива N-Achroplan 5x. Шкала 200 мкм. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
{kind=link}

Рисунок 3:Микроскопическое изображение целого отдельного миофибры с связанными с ним MuSC. Одиночные миофибры из EDL молодых мышей C57BL/6 были получены и зафиксированы PFA после изоляции (0 ч). (A)Иммунофлуоресцентное окрашивание Pax7 и DAPI идентифицирует мышечные стволовые клетки, ассоциированные с миофиброй (MuSC, отмеченные стрелками). Микроскопическое изображение было получено с использованием плитки и функции z-стека микроскопа, оснащенного масляным объективом Plan-Apochromat 40x. Шкала бара составляет 200 мкм.(B)Увеличение от(A),показывающее мионуклеи, одно положительное ядро Pax7 и ярко-темный рисунок от миофибриллярных структур в ярком поле. Шкала составляет 10 мкм. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
{kind=link}

Рисунок 4:Иммунофлуоресцентные изображения активации MuSC и миогенной прогрессии во время одной культуры миофибры. Мышечные стволовые клетки (MuSC) свежеизолированных миофиберов (0 ч) представляют собой гомеостатическое состояние, близкое к покою in vivo, характеризующееся экспрессией Pax7 и отсутствием экспрессии MyoD. Во время одной культуры миофибры большинство MuSC повышают уровень MyoD и повторно входят в клеточный цикл для деления и пролиферации (42 ч). После 72 ч культуры судьба MuSC может быть дискриминирована на основе экспрессии миогенных маркеров. Клетки с экспрессией Pax7 будут самообновляться (красная стрелка), тогда как двойные положительные клетки Pax7 и MyoD (красная / зеленая стрелка) продолжают размножаться. Только положительные клетки MyoD (зеленая стрелка) совершили миогенную дифференцировку. Микроскопические изображения были получены с помощью микроскопа с объективом Plan-Apochromat 20x (0 ч, 42 ч) или LD Plan-Neofluar 40x объективом (72 ч). Шкала 20 мкм. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
{kind=link}

Рисунок 5:Флуоресцентная трансфекция siRNA MuSC и анализ поглощения. Одиночные миофибры EDL были получены и трансфектированы флуоресцентно меченой siRNA (siGLO) в соответствии с этапами, предусмотренными этим протоколом. (A)Цитоплазматическое накопление гранул siGLO конкретно в pax7-положительной мышечной стволовой клетке (MuSC) на одном миофибре через 30 ч культуры. Микроскопическое изображение было получено с использованием z-стека и апотомной функции микроскопа с масляным объективом Plan-Apochromat 100x. Шкала стержней составляет 5 мкм.(B)Количественная оценка поглощения siRNA положительными мышечными стволовыми клетками Pax7 через 24 или 30 ч культуры. (C)Количество pax7-положительных клеток на одно миофибра, сравнивающее нетрансфектированные и трансфектированные состояния через 24 или 30 часов культивирования. Данные показаны как среднее со стандартным отклонением n = 4 мыши. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
{kind=link}
Обсуждение
Здесь представлен метод функционального исследования роли конкретного гена в MuSC с использованием подхода in vitro. Важно отметить, что в описанной здесь системе MuSC культивируются в условиях, которые максимально напоминают ситуацию in vivo, сохраняя большую часть взаимодействий MuSC со своей нишей. Это достигается путем культивирования изолированных миофибров с прилегающими к ним MuSC в плавающих условиях и последовательной трансфекции siRNA. Описаны процедуры выделения миофибры, трансфекции siRNA и исследования популяций MuSC в течение 72 ч культуры с помощью иммунофлуоресцентного анализа. Кроме того, было продемонстрировано, что около 74% всех MuSC были трансфектированы флуоресцентно меченой контрольной siRNA после 30 ч культуры.
Особое внимание следует сосредоточить на тщательном рассечении мышцы EDL, так как обширное растяжение, защемление или сдавливание приведет к сокращению и последовательной гибели миофибров. Кроме того, важно исследовать кластеры MuSC по крайней мере из 20 различных миофиберов на реплику для каждого условия. Это необходимо, поскольку числа и свойства MuSC варьируются из-за существования субпопуляций MuSC. При исследовании влияния специфической siRNA на MuSC с использованием метода плавающей культуры миофибры сравнение состояния с нацеливающей siRNA на нецелевой контроль должно проводиться в пределах одной мыши и мышцы. Это рекомендуется, чтобы избежать специфических для мышей различий, которые могут покрывать или усиливать эффекты siRNA. Эффективность нокдауна может быть определена иммунофлуоресцентным анализом с антителами, направленными против гена-мишени, с использованием одиночных миофиберов с прилегающими к ним MuSC. Если это не вариант, можно проверить эффективность нокдауна siRNA в первичных миобластах с последующим количественным анализом ОТ-ПЦР или иммуноблота. Эффективность siRNA должна быть определена до анализа влияния siRNA на MuSC на одиночные миофибры. Использование интеллектуального пула, состоящего из 4 различных siRNAs по сравнению с одним, увеличивает эффективность нокдауна, но также увеличивает риск неспецифического таргетинга. В качестве контроля следует использовать нецелевую siRNA. Для непосредственного мониторинга эффективности трансфекции можно использовать флуоресцентно меченую нецелевую siRNA, как это выполнено здесь. Временная точка для трансфекции с siRNA составляет около 4 часов после выделения, точка времени, когда базальная пластина, окружающая MuSC, уже проницаема для siRNA. Если следует исследовать влияние специфической siRNA на MuSCs после 72 или 96 ч, рекомендуется выполнять вторую трансфекцию siRNA через 24 ч или 48 ч для поддержания высокой эффективности нокдауна.
Анализ культуры миофибры демонстрирует различные преимущества по сравнению с исследованием MuSC обычными методами культивирования клеток. MuSC остаются привязанными к миофибрам в течение всего процесса изоляции, тем самым сохраняя решающее взаимодействие MuSC с его нишей19,23,24,25. Сохраненное взаимодействие MuSC с миофиброй является предпосылкой для изучения нишевых зависимых эффектов на функциональность MuSC, которые не могут быть повторены в обычных 2D-культурах миобластов. Например, во время старения MuSCs проявляют нарушенную миогенную способность, что приводит к снижению эффективности регенерации мышечной ткани после повреждения20,26. Это обесценение, по меньшей мере, частично связано с изменениями в нише MuSC, в частности изменениями в составеECM 27,28. Протокол культивирования миофибр позволяет изучать и вмешиваться в эти аберрантные изменения ниши.
В отличие от метода, описанного здесь, очистка MuSC методами иммуномаркировки и сортировки, такими как FACS (флуоресцентная активированная сортировка клеток) или MACS (магнитная сортировка клеток), включает удаление MuSC из их ниши. Интересно, что 2D-культуры изолированных MuSC из старых мышц теряют свои внешние сигналы и ведут себя аналогично MuSC, изолированным от молодых мышц, тем самым не повторяя ситуацию in vivo соответствующим образом29. Кроме того, полная диссоциация мышечной ткани и маркировка MuSC поверхностными маркерами приводит к транскриптомным изменениям и активации клеток30,31,32. Еще одним преимуществом системы культивирования миофибры является возможность вмешательства в функциональность MuSC на различных уровнях. Манипуляции с MuSC на культивируемых миофибрах могут быть эффективно достигнуты путем нокдауна гена, опосредованного siRNA, как подробно описано здесь. Аналогичным образом, применение химических соединений или доставка рекомбинантных белков очень эффективны для вмешательства в пути стволовых клеток20,28. Кроме того, ретро- или лентивирусные векторы экспрессии позволяют вводить экзогенные гены, т.е. конститутивные активные мутанты33. Кроме того, влияние внешних факторов на функциональность MuSC может быть изучено в системе, описанной здесь, например, условия культивирования могут быть дополнены супернатантом из различных физиологических или патологических источников для моделирования различных состояний, таких какраковая кахексия 34,35.
Одним из ограничений метода, описанного здесь, является тот факт, что единая система культивирования миофибры не может полностью повторить влияние всех системных факторов или влияние других типов клеток на MuSC. Кроме того, время, в течение которого миофибры могут поддерживаться жизнеспособными в культуре, ограничено, и поэтому изучение процессов, связанных с MuSC, фокусируется на ранних событиях, таких как активация и миогенная приверженность. Кроме того, исследование взаимодействия MuSC с другими нишевыми клетками, такими как иммунные клетки или фибро-адипогенные клетки-предшественники, невозможно. Для исследования системного влияния на функциональность MuSC можно либо выполнить эксперименты по повреждению мышц с последующим анализом регенерации мышц in vivo, либо выполнить эксперименты по трансплантации36,37.
Взятый вместе, протокол выделения и культивирования миофибры предоставляет большие возможности для генетических или механистических исследований на взрослых MuSC без требования трансгенных моделей мышей и потенциально может уменьшить эксперименты на животных.
Раскрытие информации
Авторы заявляют об отсутствии конкурирующих финансовых интересов.
Благодарности
Мы благодарим Кристину Позер и Кристину Пикер за отличную техническую помощь и критическое прочтение рукописи. Эта работа была поддержана грантом Deutsche Forschungsgemeinschaft JvM (MA-3975/2-1), фонда Carl Zeiss и Deutsche Krebshilfe (DKH-JvM-861005).
Материалы
| Name | Company | Catalog Number | Comments |
| Alexa Fluor 488 goat anti-rabbit IgG 2b | ThermoScientific | A-21141 | use 1:1000 for IF |
| Alexa Fluor 546 goat anti-mouse IgG1 | ThermoScientific | A-21123 | use 1:1000 for IF |
| chicken embryo extract | Seralab | CE-650-J | chicken embryo extract containing growth factors etc. |
| collagenase type 1 | Sigma | C0130 | |
| DMEM (Dulbecco’s modified Eagle’s medium with 4.5 g/l glucose and sodium pyruvate) | GibCo | 41966029 | cell culture medium |
| fetal bovine serum | Gibco | 10270-106 | fetal bovine serum |
| horse serum | Gibco | 26050-088 | |
| Lipofectamine RNAiMax | ThermoScientific | 13778150 | transfection reagent |
| MyoD antibody clone G-1 | Santa Cruz | sc-377460 | dilute 1:200 for IF |
| Pax7 antibody | DSHB | PAX7 | use undiluted |
| siGLO Red Transfection Indicator | horizon discovery | D-001630-02-05 | non targeting siRNA |
Ссылки
- Henze, H., Jung, M. J., Ahrens, H. E., Steiner, S., von Maltzahn, J. Skeletal muscle aging - Stem cells in the spotlight. Mechanisms of Ageing and Development. 189, 111283 (2020).
- Chang, N. C., Rudnicki, M. A. Satellite cells: the architects of skeletal muscle. Current Topics in Developmental Biology. 107, 161-181 (2014).
- Mauro, A. Satellite cell of skeletal muscle fibers. Journal of Biophysics and Biochemical Cytolology. 9, 493-495 (1961).
- Lepper, C., Partridge, T. A., Fan, C. M. An absolute requirement for Pax7-positive satellite cells in acute injury-induced skeletal muscle regeneration. Development. 138 (17), 3639-3646 (2011).
- Murphy, M. M., Lawson, J. A., Mathew, S. J., Hutcheson, D. A., Kardon, G. Satellite cells, connective tissue fibroblasts and their interactions are crucial for muscle regeneration. Development. 138 (17), 3625-3637 (2011).
- Sambasivan, R., et al. Pax7-expressing satellite cells are indispensable for adult skeletal muscle regeneration. Development. 138 (17), 3647-3656 (2011).
- Seale, P., et al. Pax7 is required for the specification of myogenic satellite cells. Cell. 102 (6), 777-786 (2000).
- Shea, K. L., et al. Sprouty1 regulates reversible quiescence of a self-renewing adult muscle stem cell pool during regeneration. Cell Stem Cell. 6 (2), 117-129 (2010).
- Chal, J., Pourquie, O. Making muscle: skeletal myogenesis in vivo and in vitro. Development. 144 (12), 2104-2122 (2017).
- Schmidt, M., Schuler, S. C., Huttner, S. S., von Eyss, B., von Maltzahn, J. Adult stem cells at work: regenerating skeletal muscle. Cellular and Molecular Life Sciences. 76 (13), 2559-2570 (2019).
- Motohashi, N., Asakura, A. Muscle satellite cell heterogeneity and self-renewal. Frontiers in Cell and Developmental Biology. 2, 1 (2014).
- Huttner, S. S., et al. Isolation and culture of individual myofibers and their adjacent muscle stem cells from aged and adult skeletal muscle. Methods in Molecular Biology. 2045, 25-36 (2019).
- Pasut, A., Jones, A. E., Rudnicki, M. A. Isolation and culture of individual myofibers and their satellite cells from adult skeletal muscle. Journal of Visualized Experiments. (73), e50074 (2013).
- Baghdadi, M. B., et al. Reciprocal signalling by Notch-Collagen V-CALCR retains muscle stem cells in their niche. Nature. 557 (7707), 714-718 (2018).
- Mourikis, P., et al. A critical requirement for notch signaling in maintenance of the quiescent skeletal muscle stem cell state. Stem Cells. 30 (2), 243-252 (2012).
- Pisconti, A., Cornelison, D. D., Olguin, H. C., Antwine, T. L., Olwin, B. B. Syndecan-3 and Notch cooperate in regulating adult myogenesis. Journal of Cell Biology. 190 (3), 427-441 (2010).
- Kuang, S., Kuroda, K., Le Grand, F., Rudnicki, M. A. Asymmetric self-renewal and commitment of satellite stem cells in muscle. Cell. 129 (5), 999-1010 (2007).
- Troy, A., et al. Coordination of satellite cell activation and self-renewal by Par-complex-dependent asymmetric activation of p38alpha/beta MAPK. Cell Stem Cell. 11 (4), 541-553 (2012).
- Chakkalakal, J. V., Jones, K. M., Basson, M. A., Brack, A. S. The aged niche disrupts muscle stem cell quiescence. Nature. 490 (7420), 355-360 (2012).
- Price, F. D., et al. Inhibition of JAK-STAT signaling stimulates adult satellite cell function. Nature Medicine. 20 (10), 1174-1181 (2014).
- Bernet, J. D., et al. p38 MAPK signaling underlies a cell-autonomous loss of stem cell self-renewal in skeletal muscle of aged mice. Nature Medicine. 20 (3), 265-271 (2014).
- Wang, Y. X., Dumont, N. A., Rudnicki, M. A. Muscle stem cells at a glance. Journal of Cell Science. 127, 4543-4548 (2014).
- Bentzinger, C. F., Wang, Y. X., Dumont, N. A., Rudnicki, M. A. Cellular dynamics in the muscle satellite cell niche. EMBO Reports. 14 (12), 1062-1072 (2013).
- Eliazer, S., et al. Wnt4 from the Niche Controls the Mechano-Properties and Quiescent State of Muscle Stem Cells. Cell Stem Cell. 25 (5), 654-665 (2019).
- Goel, A. J., Rieder, M. K., Arnold, H. H., Radice, G. L., Krauss, R. S. Niche cadherins control the quiescence-to-activation transition in muscle stem cells. Cell Reports. 21 (8), 2236-2250 (2017).
- Sousa-Victor, P., et al. Geriatric muscle stem cells switch reversible quiescence into senescence. Nature. 506 (7488), 316-321 (2014).
- Lukjanenko, L., et al. Loss of fibronectin from the aged stem cell niche affects the regenerative capacity of skeletal muscle in mice. Nature Medicine. 22 (8), 897-905 (2016).
- Lukjanenko, L., et al. Aging disrupts muscle stem cell function by impairing matricellular wisp1 secretion from fibro-adipogenic progenitors. Cell Stem Cell. 24 (3), 433-446 (2019).
- Alsharidah, M., et al. Primary human muscle precursor cells obtained from young and old donors produce similar proliferative, differentiation and senescent profiles in culture. Aging Cell. 12 (3), 333-344 (2013).
- Machado, L., et al. In situ fixation redefines quiescence and early activation of skeletal muscle stem cells. Cell Reports. 21 (7), 1982-1993 (2017).
- van Velthoven, C. T. J., de Morree, A., Egner, I. M., Brett, J. O., Rando, T. A. Transcriptional profiling of quiescent muscle stem cells in vivo. Cell Reports. 21 (7), 1994-2004 (2017).
- van Velthoven, C. T. J., Rando, T. A. Stem cell quiescence: Dynamism, restraint, and cellular idling. Cell Stem Cell. 24 (2), 213-225 (2019).
- Judson, R. N., et al. The Hippo pathway member Yap plays a key role in influencing fate decisions in muscle satellite cells. Journal of Cell Science. 125, 6009-6019 (2012).
- He, W. A., et al. NF-kappaB-mediated Pax7 dysregulation in the muscle microenvironment promotes cancer cachexia. Journal of Clinical Investigation. 123 (11), 4821-4835 (2013).
- Schmidt, M., Poser, C., von Maltzahn, J. Wnt7a counteracts cancer cachexia. Molecular Therapy Oncolytics. 16, 134-146 (2020).
- Feige, P., Rudnicki, M. A. Isolation of satellite cells and transplantation into mice for lineage tracing in muscle. Nature Protocols. 15 (3), 1082-1097 (2020).
- Garry, G. A., Antony, M. L., Garry, D. J. Cardiotoxin induced injury and skeletal muscle regeneration. Methods in Molecular Biology. 1460, 61-71 (2016).
Перепечатки и разрешения
Запросить разрешение на использование текста или рисунков этого JoVE статьи
Запросить разрешениеСмотреть дополнительные статьи
This article has been published
Video Coming Soon
Авторские права © 2025 MyJoVE Corporation. Все права защищены