
Для просмотра этого контента требуется подписка на Jove Войдите в систему или начните бесплатную пробную версию.
Method Article
Измерение термогенеза скелетных мышц у мышей и крыс
В этой статье
Резюме
Мышам и крысам хирургически имплантируют удаленные температурные транспондеры, а затем привыкают к тестовой среде и процедуре. Изменения мышечной температуры измеряются в ответ на фармакологические или контекстуальные стимулы в домашней клетке или во время предписанной физической активности (т. Е. Ходьбы на беговой дорожке с постоянной скоростью).
Аннотация
Термогенез скелетных мышц обеспечивает потенциальный путь для лучшего понимания метаболического гомеостаза и механизмов, лежащих в основе расхода энергии. На удивление мало доказательств того, что нейронные, миоклеточные и молекулярные механизмы термогенеза напрямую связаны с измеримыми изменениями температуры мышц. В этой статье описывается метод, в котором температурные транспондеры используются для получения прямых измерений температуры скелетных мышц мыши и крысы.
Удаленные транспондеры хирургически имплантируются в мышцы мышей и крыс, и животным дается время на восстановление. Затем мыши и крысы должны быть повторно приучены к тестовой среде и процедуре. Изменения мышечной температуры измеряются в ответ на фармакологические или контекстуальные раздражители в домашней клетке. Мышечная температура также может быть измерена во время предписанной физической активности (то есть ходьбы по беговой дорожке с постоянной скоростью), чтобы учесть изменения в активности как факторы, способствующие изменениям температуры мышц, вызванным этими стимулами.
Этот метод был успешно использован для выяснения механизмов, лежащих в основе мышечного термогенного контроля на уровне мозга, симпатической нервной системы и скелетных мышц. Приведены демонстрации этого успеха с использованием запаха хищника (PO; запах хорька) в качестве контекстуального стимула и инъекций окситоцина (Oxt) в качестве фармакологического стимула, где запах хищника вызывает термогенез мышц, а Oxt подавляет мышечную температуру. Таким образом, эти наборы данных показывают эффективность этого метода в обнаружении быстрых изменений температуры мышц.
Введение
В рамках метаболических исследований исследование термогенеза скелетных мышц является многообещающим новым способом исследования гомеостаза массы тела. Опубликованная литература поддерживает идею о том, что термогенные реакции одной из крупнейших систем органов организма — скелетных мышц — обеспечивают путь для увеличения расхода энергии и других метаболических эффектов, тем самым эффективно перебалансируя системы при таких заболеваниях, как ожирение 1,2,3. Если мышцу можно считать термогенным органом, исследования должны использовать практическую методологию для изучения термогенных изменений в этом органе. Желание понять эндотермическое воздействие скелетных мышц и полезность этой методологии для изучения недрожащего термогенеза мышц не являются специфическими для метаболических исследований. Дисциплины, включая эволюцию4, сравнительную физиологию5 и экофизиологию 6,7, проявили личную заинтересованность в понимании того, как термогенез мышц может способствовать эндотермии и как этот механизм адаптируется к окружающей среде. Представленный протокол предоставляет критические методы, необходимые для решения этих вопросов.
Предоставленный метод может быть использован при оценке как контекстуальной, так и фармакологической модуляции мышечной температуры стимулов, включая уникальную технику обеспечения запаха хищника (PO) для смещения контекста для воспроизведения угрозы хищника. Предыдущие отчеты продемонстрировали способность PO быстро индуцировать значительное увеличение мышечного термогенеза8. Кроме того, фармакологические стимулы также могут изменять температуру мышц. Это было продемонстрировано в контексте PO-индуцированного мышечного термогенеза, где фармакологическая блокада периферических β-адренорецепторов с использованием надолола ингибировала способность PO индуцировать термогенез мышц, не оказывая существенного влияния на сократительный термогенез во время ходьбы на беговой дорожке8. Центральное введение агонистов рецепторов меланокортина у крыс также использовалось для распознавания механизмов мозга, изменяющих термогенез 9,10.
Здесь представлено предварительное исследование способности нейрогормона окситоцина (Oxt) изменять термогенез мышц у мышей. Подобно угрозе хищника, социальные встречи с однополыми конспецифичными повышают температуру тела, явление, называемое социальной гипертермией11. Учитывая отношение Oxt к социальному поведению12, было высказано предположение, что Oxt является посредником социальной гипертермии у мышей. Действительно, антагонист рецептора окситоцина уменьшает социальную гипертермию у мышей11, а у детенышей мышей, лишенных Oxt, наблюдается дефицит поведенческих и физиологических аспектов терморегуляции, включая термогенез13. Учитывая, что Harshaw et al. (2021) не нашли доказательств, подтверждающих термогенез β3-адренергической рецептор-зависимой коричневой жировой ткани (BAT) с социальной гипертермией11, было высказано предположение, что социальная гипертермия может быть вызвана индункцией мышечного термогенеза Oxt.
Для измерения термогенеза скелетных мышц следующий протокол использует имплантацию предварительно запрограммированных транспондеров IPTT-300, прилегающих к интересующей мышце в мыши иликрысе 8,10,14,15. Эти транспондеры представляют собой стеклянные инкапсулированные микрочипы, которые считываются с помощью соответствующих транспондерных считывателей. Практически никакие исследования не использовали эту технологию в этом качестве, хотя исследования показали необходимость специфичности, обеспечиваемой этим методом 16,17. Предыдущие исследования показали надежность этого метода и разнообразие способов, с помощью которых могут быть использованы температурные транспондеры по сравнению с другими методамитемпературного тестирования 18 или в сочетании с хирургическими методами (например, канюляция19). Тем не менее, исследования такого рода полагаются на различные стратегические места для измерения общей температуры тела 20,21,22 или определенных тканей, таких как BAT 23,24,25.
Вместо того, чтобы измерять температуру в этих местах или при использовании ушных или ректальных термометров26, способ, описанный здесь, обеспечивает специфичность для интересующей мышцы. Способность нацеливаться на участок путем непосредственной имплантации транспондеров, прилегающих к интересующим мышцам, более эффективна для зондирования термогенеза мышц. Он предоставляет новый путь в дополнение к тем, которые обеспечиваются поверхностной инфракрасной термометрией27,28 или измерениями кожной температуры с помощью термопары 29. Кроме того, данные, предоставленные с помощью этого метода, предлагают ряд направлений исследований, избегая необходимости в большом, дорогостоящем, высокотехнологичном оборудовании и программном обеспечении, таком как инфракрасная термография 30,31,32.
Этот метод успешно используется для измерения температуры в квадрицепсах и икроножной крупемии, либо в одностороннем, либо в двустороннем порядке. Этот метод также был эффективен в сочетании со стереотаксической хирургией14,15. В пределах ~ 7-10 см от конечности транспондера портативные считыватели транспондера (DAS-8027 / DAS-7007R) используются для сканирования, измерения и отображения температуры. Это расстояние было критическим и ценным для предыдущих исследований 8,9,10, поскольку оно сводит к минимуму потенциальные стрессоры и изменяющие температуру переменные, такие как обращение с животными во время процедур тестирования. Используя таймеры, измерения могут быть записаны и собраны в течение определенного периода времени без прямого взаимодействия с животными.
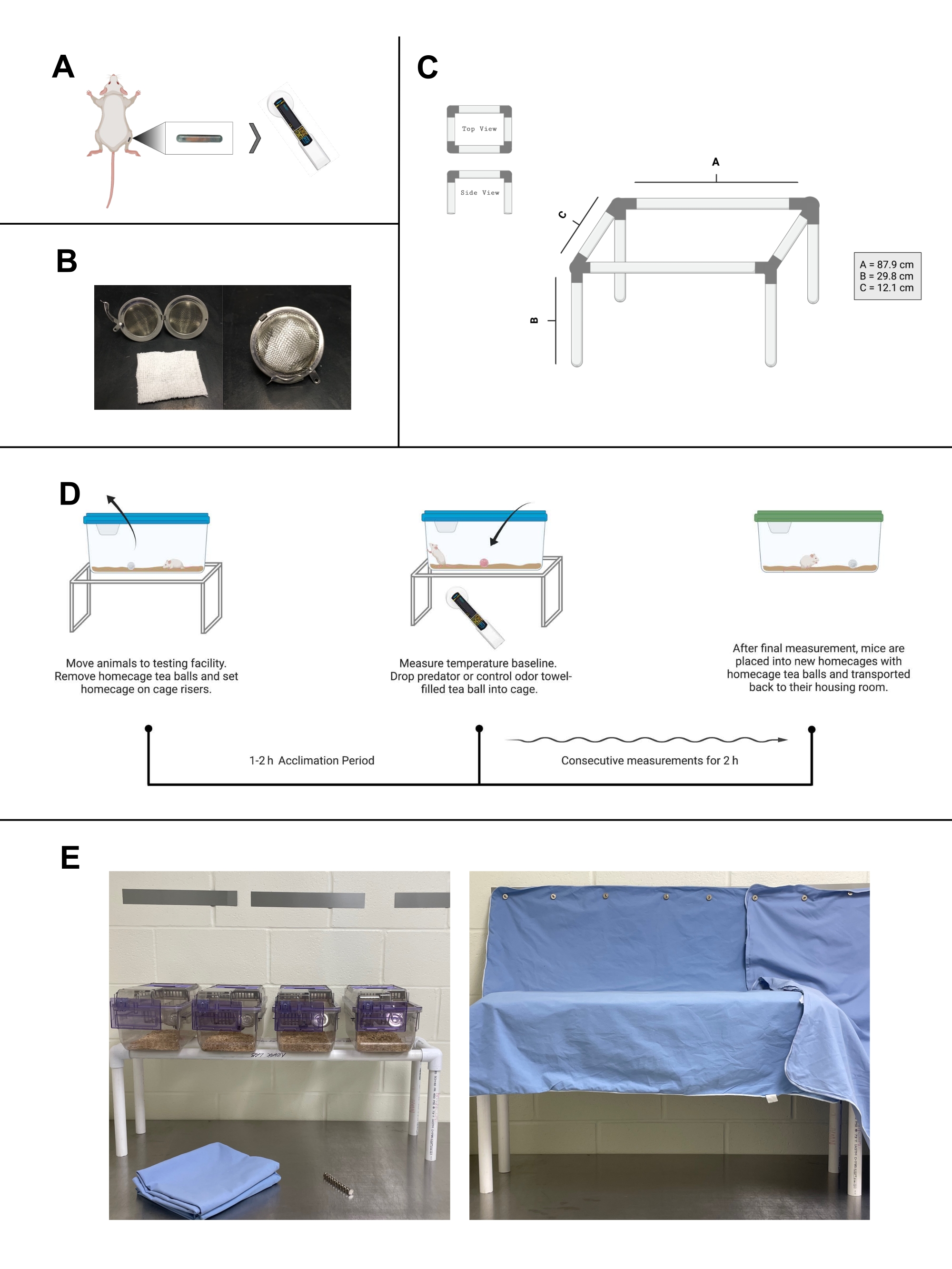
Чтобы еще больше свести к минимуму беспокойство мышей во время тестирования, этот метод описывает сборку и использование стояков из труб ПВХ, чтобы дать экспериментатору доступ к нижней части домашних клеток во время тестирования. Используя стояки в тандеме с цифровым считывателем, измерения температуры конечности транспондера могут быть выполнены без какого-либо взаимодействия с животными после размещения стимула. При минимальных затратах этот метод можно использовать совместно с фармакологическими и контекстуальными раздражителями, что делает его вполне доступным для исследователей. Кроме того, этот метод может быть использован со значительным количеством субъектов (~ 16 мышей или ~ 12 крыс) за раз, экономя время на увеличение общей пропускной способности для любого исследовательского проекта.
В этот метод введен созданный механизм для представления запахов мышам с использованием шариков из нержавеющей стали, называемых «чайными шариками». Хотя эти чайные шарики идеально подходят для содержания любого запахового материала, в этих исследованиях полотенца, которые служили постельным бельем в клетке в течение 2-3 недель для хорьков, естественного хищника мышей и крыс, помещаются в каждый лечебный чайный шарик. Каждое полотенце разрезается на квадраты размером 5 см х 5 см. Эта аликвотация также повторяется с идентичными контрольными полотенцами без запаха. Представление этих запахов без барьера (то есть чайного шарика) привело к тому, что мыши измельчали волокна в своих клетках, увеличивая физическую активность. Такое поведение не было столь заметным у крыс. Чайные шарики обеспечивают вентилируемую оболочку полотенца, давая полный доступ к запаху, оставаясь защищенными в течение всего экспериментального испытания. Эти чайные шарики могут быть продезинфицированы в соответствии с протоколами использования животных, приготовлены и введены непосредственно после операции, чтобы начать приучать животных к структуре вместе с контрольным стимулом. Мыши могут затем жить с дополнительным обогащением, уменьшая значимость острого представления стимула.
Привыкание к присутствию чайного шарика является лишь одним из аспектов привыкания, который имеет решающее значение для этого метода. Описанный протокол привыкания также состоит из повторного воздействия процедуры тестирования для нормализации испытательной среды (т.е. персонала, транспортировки и перемещения к месту тестирования, воздействия стимула). Это расширенное привыкание сводит к минимуму нюансированные реакции животных и фокусирует измерения на желаемых зависимых переменных (например, фармакологических или контекстуальных стимулах). Предыдущая оценка этого протокола определила четыре испытания в качестве минимального количества привычек, необходимых перед температурным тестированием в домашних клетках у крыс8. Если тестирование разделено длительными периодами (более 2-3 недель), животные должны быть повторно приучены. Для повторного привыкания достаточно как минимум одного-двух испытаний. Однако, если температурные тесты разделены более длительными приступами времени, может потребоваться повторение большего количества испытаний.
В постоянных усилиях по приучению мышей и крыс к процедуре тестирования в каждое экспериментальное испытание должен быть включен период акклиматизации перед представлением стимула. Это время акклиматизации имеет решающее значение для восстановления баланса температуры и активности после перемещения в место тестирования. Грызуны, как правило, имеют резкое повышение температуры из-за транслокации. Акклиматизация должна состоять минимум из 1 ч без взаимодействия со стороны экспериментатора в день тестирования перед любым добавлением фармакологического средства или контекстуальных стимулов. Это необходимо каждый день тестирования.
В описанных тестах температуры домашней клетки мыши имеют свободный диапазон своей домашней клетки, чтобы бродить в ответ на тестируемый стимул. Это может вызвать переменные сдвиги в активности, влияющие на точность показаний температуры и, следовательно, на анализ термогенных эффектов независимой переменной (например, фармакологического или контекстуального стимула). В знак признания потенциальных изменений температуры из-за уровня активности ниже включен протокол, описывающий использование температуры во время ходьбы на беговой дорожке. Опубликованная литература описывает успешное использование этой процедуры у крыс, и в настоящее время она используется смышами 8,10,14,15. Ходьба на беговой дорожке поддерживает постоянную скорость активности для испытуемого. Для этого исследования беговые дорожки строго используются для контроля уровня активности и, следовательно, устанавливаются на самую низкую доступную скорость на беговой дорожке, чтобы способствовать ходьбе для мышей и аналогично низкую настройку для крыс.
Следующая процедура описана для измерения температуры односторонней икроножной железы у мышей и проявления запаха хищника. Конструкция может быть использована в сочетании с фармакологическими агентами и переносится на крыс и другие группы скелетных мышц (т.е. квадрицепсы) у мышей. Для крыс транспондеры могут быть помещены в икроножную мышцу двусторонне и в коричневую жировую ткань. Из-за ограничений по размеру и расстоянию на мышь можно использовать только один транспондер. Незначительные изменения (например, удаление контекстуальных стимулов) могут быть сделаны для оценки термогенных реакций на фармакологические агенты.
Access restricted. Please log in or start a trial to view this content.
протокол
Эти методы могут быть применены как к моделям крыс, так и к мышам и были выполнены с институционального одобрения (Kent State University, IACUC Approval #359 и #340 CN 12-04). До введения в действие протокола животные должны содержаться в соответствии с Руководством по уходу за лабораторными животными и их использованию.
1. Подготовка считывателя транспондера
ПРИМЕЧАНИЕ: Перед использованием необходимо установить считыватель транспондера; Следующие шаги включают только изменения настроек, необходимые для этого исследования. Эта часть протокола напрямую связана с портативными считывателями DAS-8027-IUS; другие модели считывателей должны следовать инструкциям, приведенным в руководстве, для достижения результатов программирования.
- Установите для параметра Звуковой сигнал значение ВЫКЛ.
- Включите устройство, нажав кнопку SCAN , и дождитесь появления подсветки на OLED-дисплее. Нажмите и удерживайте кнопку BACK/MENU , чтобы перейти на экран меню .
- С помощью кнопки NEXT/ENTER прокрутите параметры до operational setup. Здесь переключите стрелки вверх или вниз, чтобы повернуть ДА и открыть рабочее подменю.
- С помощью кнопки NEXT/ENTER прокрутите экран до пункта ЗВУКОВОЙ СИГНАЛ. Если по умолчанию установлено значение ВКЛ., переключите стрелки вверх или вниз и измените параметр на ВЫКЛ.
- Нажмите кнопку NEXT/ENTER , чтобы сохранить это изменение параметра.
- Установите для параметра Вибрация при чтении значение ВКЛ.
- Выполните шаги с 1.1 по 1.2 или выполните следующий шаг непосредственно после шага 1.4.
- С помощью кнопки NEXT/ENTER прокрутите до пункта ВИБРАЦИЯ ПРИ ЧТЕНИИ. Если по умолчанию установлено значение OFF, переключите стрелки вверх и вниз и измените настройку на ON , чтобы с помощью вибрации чувствовать, когда чтение завершено, независимо от возможности просмотра экрана.
2. Программные транспондеры
ПРИМЕЧАНИЕ: Каждый имплантированный транспондер должен быть сначала запрограммирован с идентификацией животного (идентификатор животного или идентификатор транспондера). Эта номенклатура может быть использована в качестве вторичной идентификации для испытуемого (например, четыре цифры для аббревиатуры штамма мыши, расположение транспондера и дополнительные три-четыре цифры для обозначения номера животного). Программирование может быть завершено за несколько дней до операции, сохраняя транспондеры стерильными до операции.
- Введите идентификационный код на транспондере.
- Приложите к головке считывателя катушку бустера — специальный аксессуар для модели DAS 8027-IUS, который помогает в процедуре программирования.
- Рукой в перчатке поместите транспондер (внутри аппликатора) в бустерную катушку.
- Включите устройство, нажав кнопку SCAN , и подождите, пока загорится OLED-дисплей. Нажмите и удерживайте кнопку BACK/MENU , чтобы перейти на экран меню .
- С помощью кнопки NEXT/ENTER прокрутите параметры до write transponder ID. Здесь переключите стрелки вверх или вниз, чтобы повернуть ДА.
- С помощью кнопки NEXT/ENTER переключите команду ВВЕДИТЕ ИДЕНТИФИКАЦИОННЫЙ КОД.
- Используйте клавиши со стрелками вверх и вниз для прокрутки цифр и букв. Нажимайте КЛАВИШУ NEXT/ENTER после каждого выбора символа, чтобы перейти к следующему символу.
- Когда идентификационный код будет завершен, нажмите SCAN , чтобы записать транспондер.
- Извлеките транспондер из катушки бустера и повторите по мере необходимости. Убедитесь, что транспондер считывает изменения температуры, нагревая закрытые транспондеры между руками в перчатках и измеряя с помощью сканера температуры.
ПРИМЕЧАНИЕ: Параметры AUTO MULTI WRITE и SEQUENTIAL COUNT могут быть установлены в положение ON , чтобы разрешить программирование нескольких или последовательных транспондеров во время сеанса. Каждый транспондер должен быть протестирован во время программирования.
3. Подготовьте «домашние шарики в клетке»
- Поместите 5 см х 5 см без запаха / контрольное полотенце в чайный шарик.
- Поместите эти домашние клетки в новые домашние клетки после операции, чтобы начать приучать животное к методу, в котором контекстуальные стимулы будут представлены во время тестирования. Заменяйте эти домашние шарики клетки каждые 2 недели.
4. Хирургия и послеоперационный уход
- Взвешивайте и записывайте предоперационную массу тела испытуемых. Используя индукционную камеру, обеспечьте животному анестезию (например, 2-5% изофлурана).
- Используя электрические кусачки, полностью побрейте заднюю конечность. Вводят анальгезию (например, 5 мг/кг кетопрофена, s.c.) в соответствии с институциональными рекомендациями.
ПРИМЕЧАНИЕ: Дополнительная анальгезия может потребоваться, если эта процедура сочетается с другими хирургическими методами. - Очистите область 70% спиртом (или коммерчески доступной стерильной спиртовой салфеткой) и промывкой повидоном-йодом (или коммерчески доступными стерильными, индивидуально обернутыми бетадиновыми тампонами), чередуя по крайней мере три раза, заканчивая повидоном-йодом.
- Верните животное в индукционную камеру и обезболите животное до хирургического уровня. Затем наденьте мышь в маску для лица для продолжения воздействия анестезии. Нанесите неомициновую офтальмологическую мазь на глаза животного, чтобы предотвратить сухость во время анестезии.
ПРИМЕЧАНИЕ: Процедура не должна начинаться до тех пор, пока у мыши не будет обнаружено признаков приема боли (т.е. рефлекса роговицы, реакции защемления хвоста, рефлекса защемления пальца ноги). - Используя только хирургические ножницы, сделайте неглубокий разрез через кожу на правой задней конечности.
- Двигаясь параллельно икроножной мышце, поместите острый край запрограммированного и неограниченного стерильного транспондера в разрез. Убедитесь, что зеленый плунжер обращен вверх и виден. Продолжайте проталкивать аппликатор транспондера в разрез до тех пор, пока отверстие аппликатора транспондера не перестанет быть видимым.
ПРИМЕЧАНИЕ: Не следует случайно нажимать зеленый плунжер на аппликатор транспондера на этапе 4.6. Преждевременный разряд транспондера приведет к неправильному размещению. - Поверните аппликатор на 180°, в результате чего зеленый поршень будет обращен вниз к конечности мыши, больше не виден экспериментатору. Вставьте аппликатор транспондера в конечное место. Оказавшись в идеальном месте, рядом или частично заключенном в икроножную мышцу, оттолкните зеленый поршень, позволяя давлению аппликатора направлять руку исследователя назад от мыши.
- С помощью щипцов скрепите вскрытую кожу и поместите раневой зажим со стерильным автоклипом или стерильным швом. При необходимости используйте рассасывающиеся швы перед стерильным автоклипом, чтобы закрыть слой фасции. С помощью транспондера-считывателя проверьте температуру мышц мыши.
- Извлеките мышь из анестезии и поместите ее в чистую домашнюю клетку, помещенную на водоциркуляционную грелку, установленную на низком уровне для восстановления. Убедитесь, что домашняя клетка включает чайный шарик с полотенцем без запаха, чтобы начать привыкание.
ПРИМЕЧАНИЕ: Мышь должна проснуться от операции в течение 15 минут. Пища может быть помещена на дно клетки для легкого доступа в дни восстановления. - Послеоперационный уход
- Записывайте вес и температуру мыши ежедневно с помощью транспондера-считывателя в течение как минимум 2 дней после операции или до тех пор, пока мыши не восстановят или не стабилизируют массу тела.
- Вводят ненаркотическую анальгезию (например, 5 мг/кг кетопрофена, s.c.) один раз в день мышам в течение не менее 2 дней после операции, с дополнительными дозами, предоставляемыми по мере необходимости.
ПРИМЕЧАНИЕ: Мыши и крысы должны полностью восстановиться в течение 5-8 дней после операции и могут пройти процедуры привыкания и тестирования.
5. Подготовка к тестированию - домашняя клетка
- Строительство стояков
ПРИМЕЧАНИЕ: Приведенный ниже шаг основан на отсеках с фильтром мыши 194 мм x 181 мм x 398 мм. Чтобы поместить большие клетки (например, домашнюю клетку для крыс), ширина должна быть отрегулирована.- Разрежьте трубу из ПВХ с помощью храпового резака ПВХ на восемь секций и соберите по рисунку 1С. Это даст открытую настольную структуру, которая может вместить примерно четыре клетки. Сделайте нужное количество стояков.
- Обстановка в номере
- Назначьте расположение каждой переходной платы в комнате тестирования. Отделите стояки, установленные для получения различных контекстуальных стимулов (т. Е. Запахов), как минимум на 2 м, чтобы избежать путаницы переменных.
ПРИМЕЧАНИЕ: Каждая мышь должна иметь назначенное место для тестирования в испытательной комнате и на физических стояках настолько, насколько это возможно, чтобы избежать развития ассоциаций между различными местами и термогенными стимулами. - Используя магнитные полосы, прикрепите хирургические листы или халаты через стояки, создавая визуальный барьер между исследователем и испытуемыми. Установите этот барьер, чтобы свести к минимуму изменения температуры в результате активности мыши при просмотре экспериментаторов, движущихся к клетке или вокруг испытательной комнаты.
- (Необязательно) Разместите зеркала на поверхности под стояками, чтобы облегчить обзор дна клетки во время тестирования.
ПРИМЕЧАНИЕ: Стояки могут быть продезинфицированы через систему мойки клеток. Ткань или хирургические простыни должны быть выстираны перед привыканием и тестированием.
- Назначьте расположение каждой переходной платы в комнате тестирования. Отделите стояки, установленные для получения различных контекстуальных стимулов (т. Е. Запахов), как минимум на 2 м, чтобы избежать путаницы переменных.
- Приготовление чайного шарика
- Приготовьте чайные шарики с контролем и полотенцами PO (примерно 5 см х 5 см). Чтобы избежать перекрестного загрязнения, сначала приготовьте чайные шарики с контрольным запахом.
ПРИМЕЧАНИЕ: Полотенца с запахом хищника должны быть проверены на патогены перед использованием. Эти полотенца также должны содержаться, а материалы, которые взаимодействуют с ними, должны быть немедленно продезинфицированы (т. Е. Мытье клеток), предотвращая воздействие запаха на других животных.
- Приготовьте чайные шарики с контролем и полотенцами PO (примерно 5 см х 5 см). Чтобы избежать перекрестного загрязнения, сначала приготовьте чайные шарики с контрольным запахом.
6. Температурный тест - домашняя клетка
ПРИМЕЧАНИЕ: Животные должны быть приучены ко всей процедуре тестирования, исключая экспериментальные контекстуальные или фармакологические стимулы. Это должно быть выполнено как минимум в 4 раза перед тестированием.
- Перенесите животных в подготовленную испытательную комнату. Поместите животных в заранее назначенное место на стояке. Это место должно быть одинаковым во всех процедурах привыкания и тестирования.
- Извлеките «шарик домашней клетки» из домашней клетки мыши и снова накройте клетки тканью или хирургическим листом. Дайте мышам акклиматизироваться в испытательном пространстве в течение 1-2 ч.
- После завершения акклиматизации используйте сканер для измерения и записи базовой температуры каждого субъекта. Избегайте манипулирования тканевыми покрытиями во время измерений.
ПРИМЕЧАНИЕ: Здесь могут применяться фармакологические средства. Время ожидания после инъекции или приложения может быть добавлено по мере необходимости перед тестированием. Запись вторичного исходного уровня непосредственно перед тестированием рекомендуется после добавления фармакологического средства для мониторинга реакции на фармакологические стимулы. Если реакция запаха не проверяется, измерения температуры мышей могут начаться сразу после инъекции. Рандомизация должна использоваться при предоставлении любых стимулов. - Раскройте клетку и поместите чайный шарик (контрольный или PO) на пол домашней клетки. Замените крышку клетки и тканевое покрытие.
- Запустите секундомер. Измерьте температуру испытуемых в том же порядке размещения чайного шарика. Регистрируйте температуру и время часов измерений в соответствии с требуемыми временными точками.
- Когда эксперимент будет завершен, снимите лечебный шарик. Поместите мышей, получивших PO, в новую домашнюю клетку с оригинальным «домашним клеточным шаром». Верните «домашнюю клетку шара» в клетку мышей, получивших контрольный запах. Переместите мышей в место расположения корпуса.
ПРИМЕЧАНИЕ: Приведенная выше процедура может быть переведена на модели крыс в клетках соответствующего размера. Могут потребоваться корректировки измерений, предложенных на рисунке 1С , чтобы обеспечить лучший доступ к нижней части домашней клетки.

Рисунок 1: Тестирование температуры транспондеров и домашней клетки. (А) Схема одностороннего размещения транспондеров для проверки температуры в икроножной мышце мыши. После программирования и размещения транспондер-считыватель (показан DAS-8027-IUS) может использоваться для измерения температуры. (B) Слева, фотография чайного шарика из нержавеющей стали с открытой сеткой и полотенца размером 5 см х 5 см. Справа, вложенный чайный шарик, используемый для хранения привыкания и запаха полотенец в домашней клетке тестирования. (C) Схема стояков, изготовленных из труб из ПВХ, для тестирования домашних клеток. (D) Рабочий процесс протокола тестирования домашней клетки. (E) Изображения объекта зоны тестирования домашней клетки. Слева четыре клетки для мышей на стояке. Магнитные полосы расположены на соседней стене, а магниты и хирургическая ткань находятся на столе. Справа, покрытые клетки для мышей на стояках. (A), (C) и (D) были созданы с Biorender.com. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
{kind=link}
7. Температурный тест - ходьба на беговой дорожке
- Назначьте каждому животному беговую дорожку в качестве назначенного места для привыкания и процедур тестирования.
- Подготовьте беговые дорожки к тестированию, убедившись, что шокеры функциональны.
ПРИМЕЧАНИЕ: Для ходьбы на беговой дорожке беговые дорожки должны быть установлены в самом низком доступном темпе, который способствует непрерывному движению, но не бегу как для привыкания, так и для тестирования. Для модульной закрытой метаболической беговой дорожки 1012M-2 это 5,2 м/мин для мышей и 7 м/мин для крыс. Этот темп, возможно, потребуется скорректировать на основе ожирения субъекта. Шокеры должны быть установлены на интенсивность и частоту повторения 5,0. - Привыкание
- Переместите мышей в комнату для тестирования. Дайте мышам 1-2 часа акклиматизироваться в комнате, перенесите в свои домашние клетки.
- После акклиматизации направьте мышей к открытию назначенной им беговой дорожки и закройте беговую дорожку. Запустите ремень, шокер и секундомер.
- Позвольте мышам ходить по беговым дорожкам в течение 15 минут, используя шоковый стимул в качестве мотивации для движения. Немедленно прекратите тест, если животное остается на активном шокере в течение длительного периода.
- После теста удалите мышей и верните их в домашние клетки.
- Очистите беговые дорожки, используя жидкое моющее средство и воду.
- Тестирование
- Переместите мышей в комнату для тестирования. Дайте мышам 1-2 часа акклиматизироваться в комнате передачи в их домашних клетках.
- Измерьте и запишите базовую температуру перед перемещением мыши на беговую дорожку.
ПРИМЕЧАНИЕ: Для испытаний, включая фармакологические средства, применяйте или вводите их здесь, следуя схеме, показанной на рисунке 2A. Время ожидания после инъекции может быть добавлено по мере необходимости, прежде чем мыши будут помещены на беговую дорожку. Рандомизация должна использоваться при предоставлении любых стимулов. - Поместите квадраты контроля размером 5 см х 5 см или полотенца PO в беговую дорожку, ближайшую к передней части беговой дорожки. Прикрепите полотенца к потолку беговой дорожки или под ней для удобного размещения и снятия.
- Направьте мышей на назначенную беговую дорожку. Включите ремень беговой дорожки и шокер.
- Запустите секундомер. Проведите измерения испытуемых в том же порядке, в котором мыши были установлены на беговых дорожках. Запишите температуру и время часов измерений в соответствии с желаемыми временными точками.
ПРИМЕЧАНИЕ: Температура может быть надежно измерена снаружи беговой дорожки, в то время как мышь находится внутри закрытой беговой дорожки во время ходьбы. Для крыс ограничения размера беговой дорожки и расстояния транспондера-считывателя могут потребовать от экспериментатора держать заднюю часть беговой дорожки открытой, чтобы вставить считыватель внутрь беговой дорожки, ближе к объекту. - Когда тест будет завершен, выключите шокеры и беговые дорожки; вернуть мышей в их домашние клетки. Переместите мышей в место расположения корпуса.
- Очистите беговые дорожки, используя жидкое моющее средство и воду, уделяя особое внимание удалению любых остаточных PO.
- Когда эксперименты будут завершены, усыпьте животных (например, используя вдыхание CO2 ) и визуально подтвердите местоположение транспондера.

Рисунок 2: Тестирование температуры с контролируемой активностью. (A) Рабочий процесс контролируемого температурного тестирования с помощью фармакологического агента с использованием беговой дорожки. (B) Изображения беговых дорожек. Слева, изображение полной настройки оборудования. Справа, более близкое изображение отдельных беговых дорожек и шокеров. (A) был создан с Biorender.com. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
{kind=link}
Access restricted. Please log in or start a trial to view this content.
Результаты
Транспондеры были в одностороннем порядке имплантированы в правую икроножную мышцу десяти 4-6-месячных мышей дикого типа (WT), выведенных из штамма SF1-Cre (Tg(Nr5a1-cre)7Lowl/J, Strain #012462, C57BL/6J и FVB фонов; самка N = 5; самец N = 5). После выздоровления мышей приучали к процедуре температурного тестирования до...
Access restricted. Please log in or start a trial to view this content.
Обсуждение
Этот протокол температурного тестирования предоставляет поле возможность непосредственно измерить термогенез скелетных мышц. Это имеет решающее значение, поскольку исследования углубляются в идентификацию механизмов, лежащих в основе мышечного термогенеза33. Метод обе...
Access restricted. Please log in or start a trial to view this content.
Раскрытие информации
Авторы заявляют, что у них нет конфликта интересов.
Благодарности
Эта работа поддерживается R15 DK097644 и R15 DK108668. Мы благодарим д-ра Чайтанью К. Гавини и д-ра Меган Рич за предыдущие вклады и д-ра Стэнли Даннемиллера за обеспечение соблюдения нами институциональных руководящих принципов использования животных. Отдельное спасибо доктору Тиму Бартнессу за предоставление фундаментальных исследований, необходимых для построения этого метода и связанных с ним исследований. Рисунки 1A, C, D и 2A были созданы с использованием Biorender.com.
Access restricted. Please log in or start a trial to view this content.
Материалы
| Name | Company | Catalog Number | Comments |
| 1012M-2 Modular Enclosed Metabolic Treadmill for Mice, 2 Lanes w/ Shock | Columbus Instruments | ||
| 1012R-2 Modular Enclosed Metabolic Treadmill for Rats, 2 Lanes w/ Shock | Columbus Instruments | ||
| 1-1/4 in. Ratcheting PVC Cutter | BrassCraft | ||
| 1 mL Syringes | Fisher Scientific | BD 309659 | |
| Betadine Swabs | Fisher Scientific | 19-898-945 | |
| Booster Coil | BioMedic Data Systems | Transponder Accessory | |
| Electric Clippers | Andis | 40 Ultraedge Clipper Blade | |
| Flexible Mirror Sheets | Amazon | Self Adhesive Non Glass Mirror Tiles | |
| Forceps | Fisher Scientific | 89259-940 | |
| Heating Pad | |||
| Induction Chamber (isoflurane) | Kent Scientific | VetFlo-0730 | 3.0 L Low Cost Chambers for Traditional Vaporizers |
| Ketoprophen | Med-Vet Intl. | RXKETO-50 | |
| Magnetic Strips | Amazon | ||
| Magnets | Amazon | DIYMAG Magnetic Hooks 40lbs | |
| Needles | Med-Vet Intl. | 26400 | |
| Neomycin/Polymixin/Bacitracin with Hydrocortisone Ophthalmic Ointment, 3.5 g | Med-Vet Intl. | RXNPB-HC | |
| Oasis Absorbable Suture | Med-Vet Intl. | MV-H821-V | |
| Predator (Ferret) Odor Towels | Marshall BioResources | ||
| PVC pipe | |||
| Reflex Wound Clip Remover | CellPoint Scientific | ||
| Reflex Wound Clip, 7 mm (mouse) | CellPoint Scientific | ||
| Reflex Wound Clip, 9 mm (rat) | CellPoint Scientific | ||
| Srerile Autoclip, 7 mm (mouse) | CellPoint Scientific | Wound Clip Applier (mouse) | |
| Stainless Strainers Interval Seasonings Tea Infuser | Amazon | ||
| Sterile Autoclip, 9 mm (rat) | CellPoint Scientific | Wound Clip Applier (rat) | |
| Sterile Saline | Med-Vet Intl. | RX0.9NACL-10 | |
| Surgical Scissors | Fisher Scientific | 08-951-5 | |
| Surgical Sheets | |||
| Towels (Control/Habituation) | Amazon | 100% Cotton Towels, white | |
| Transponders | BioMedic Data Systems | Model: IPTT-300 | |
| Transponders Reader | BioMedic Data Systems | Model: DAS-8027-IUS/ DAS-7007R | |
| Versaclean | Fisher Scientific | 18-200-700 | liquid detergent |
| Webcol Alcohol Preps | Covidien | 22-246-073 | |
| Wedge pieces for PVC pipe |
Ссылки
- Periasamy, M., Herrera, J. L., Reis, F. C. G. Skeletal muscle thermogenesis and its role in whole body energy metabolism. Diabetes Metabolism Journal. 41 (5), 327-336 (2017).
- Rowland, L. A., Bal, N. C., Periasamy, M. The role of skeletal-muscle-based thermogenic mechanisms in vertebrate endothermy. Biological Reviews of the Cambridge Philosophical Society. 90 (4), 1279-1297 (2015).
- Maurya, S. K., et al. Sarcolipin is a key determinant of the basal metabolic rate, and its overexpression enhances energy expenditure and resistance against diet-induced obesity. Journal of Biological Chemistry. 290 (17), 10840-10849 (2015).
- Grigg, G., et al. Whole-body endothermy: Ancient, homologous and widespread among the ancestors of mammals, birds and crocodylians. Biological Reviews of the Cambridge Philosophical Society. 97 (2), 766-801 (2022).
- Franck, J. P. C., Slight-Simcoe, E., Wegner, N. C. Endothermy in the smalleye opah (Lampris incognitus): A potential role for the uncoupling protein sarcolipin. Comparative Biochemistry and Physiology - Part A: Molecular & Integrative Physiology. 233, 48-52 (2019).
- Nowack, J., et al. Muscle nonshivering thermogenesis in a feral mammal. Scientific Reports. 9, 6378(2019).
- Oliver, S. R., Anderson, K. J., Hunstiger, M. M., Andrews, M. T. Turning down the heat: Down-regulation of sarcolipin in a hibernating mammal. Neuroscience Letters. 696, 13-19 (2019).
- Gorrell, E., et al. Skeletal muscle thermogenesis induction by exposure to predator odor. The Journal of Experimental Biology. 223, Pt 8 (2020).
- Gavini, C. K., et al. Leanness and heightened nonresting energy expenditure: Role of skeletal muscle activity thermogenesis. The American Journal of Physiology - Endocrinology and Metabolism. 306 (6), 635-647 (2014).
- Almundarij, T. I., Gavini, C. K., Novak, C. M. Suppressed sympathetic outflow to skeletal muscle, muscle thermogenesis, and activity energy expenditure with calorie restriction. Physiological Reports. 5 (4), 13171(2017).
- Harshaw, C., Lanzkowsky, J., Tran, A. D., Bradley, A. R., Jaime, M. Oxytocin and 'social hyperthermia': Interaction with beta3-adrenergic receptor-mediated thermogenesis and significance for the expression of social behavior in male and female mice. Hormones and Behavior. 131, 104981(2021).
- Caldwell, H. K. Oxytocin and vasopressin: Powerful regulators of social behavior. The Neuroscientist. 23 (5), 517-528 (2017).
- Harshaw, C., Leffel, J. K., Alberts, J. R. Oxytocin and the warm outer glow: Thermoregulatory deficits cause huddling abnormalities in oxytocin-deficient mouse pups. Hormones and Behavior. 98, 145-158 (2018).
- Gavini, C. K., Britton, S. L., Koch, L. G., Novak, C. M. Inherently lean rats have enhanced activity and skeletal muscle response to central melanocortin receptors. Obesity. 26 (5), 885-894 (2018).
- Gavini, C. K., Jones, W. C., Novak, C. M. Ventromedial hypothalamic melanocortin receptor activation: regulation of activity energy expenditure and skeletal muscle thermogenesis. The Journal of Physiology. 594 (18), 5285-5301 (2016).
- Zaretsky, D. V., Romanovsky, A. A., Zaretskaia, M. V., Molkov, Y. I. Tissue oxidative metabolism can increase the difference between local temperature and arterial blood temperature by up to 1.3(o)C: Implications for brain, brown adipose tissue, and muscle physiology. Temperature. 5 (1), 22-35 (2018).
- Yoo, Y., et al. Exercise activates compensatory thermoregulatory reaction in rats: A modeling study. Journal of Applied Physiology. 119 (12), 1400-1410 (2015).
- Langer, F., Fietz, J. Ways to measure body temperature in the field. Journal of Thermal Biology. 42, 46-51 (2014).
- Pence, S., et al. Central apolipoprotein A-IV stimulates thermogenesis in brown adipose tissue. International Journal of Molecular Sciences. 22 (3), 1221(2021).
- Li, D., et al. Homeostatic disturbance of thermoregulatory functions in rats with chronic fatigue. Journal of Neuroscience Research. 165, 45-50 (2021).
- Carlier, J., et al. Pharmacodynamic effects, pharmacokinetics, and metabolism of the synthetic cannabinoid AM-2201 in male rats. Journal of Pharmacology and Experimental Therapeutics. 367 (3), 543-550 (2018).
- Pato, A. M., Romero, D. M., Sosa Holt, C. S., Nemirovsky, S. I., Wolansky, M. J. Use of subcutaneous transponders to monitor body temperature in laboratory rats. Journal of Pharmacological and Toxicological Methods. 114, 107145(2022).
- Almeida, D. L., et al. Lean in one way, in obesity another: Effects of moderate exercise in brown adipose tissue of early overfed male Wistar rats. International Journal of Obesity. 46 (1), 137-143 (2022).
- Brito, M. N., Brito, N. A., Baro, D. J., Song, C. K., Bartness, T. J. Differential activation of the sympathetic innervation of adipose tissues by melanocortin receptor stimulation. Endocrinology. 148 (11), 5339-5347 (2007).
- Vaughan, C. H., Shrestha, Y. B., Bartness, T. J. Characterization of a novel melanocortin receptor-containing node in the SNS outflow circuitry to brown adipose tissue involved in thermogenesis. Brain Research. 1411, 17-27 (2011).
- Kort, W. J., Hekking-Weijma, J. M., TenKate, M. T., Sorm, V., VanStrik, R. A microchip implant system as a method to determine body temperature of terminally ill rats and mice. Laboratory Animals. 32 (3), 260-269 (1998).
- Mei, J., et al. Body temperature measurement in mice during acute illness: Implantable temperature transponder versus surface infrared thermometry. Scientific Reports. 8, 3526(2018).
- Warn, P. A., et al. Infrared body temperature measurement of mice as an early predictor of death in experimental fungal infections. Laboratory Animals. 37 (2), 126-131 (2003).
- Hargreaves, K., Dubner, R., Brown, F., Flores, C., Joris, J. A new and sensitive method for measuring thermal nociception in cutaneous hyperalgesia. Pain. 32 (1), 77-88 (1988).
- Fiebig, K., Jourdan, T., Kock, M. H., Merle, R., Thone-Reineke, C. Evaluation of infrared thermography for temperature measurement in adult male NMRI nude mice. Journal of the American Association for Laboratory Animal Science. 57 (6), 715-724 (2018).
- Franco, N. H., Geros, A., Oliveira, L., Olsson, I. A. S., Aguiar, P. ThermoLabAnimal - A high-throughput analysis software for non-invasive thermal assessment of laboratory mice. Physiology & Behavior. 207, 113-121 (2019).
- Koganti, S. R., et al. Disruption of KATP channel expression in skeletal muscle by targeted oligonucleotide delivery promotes activity-linked thermogenesis. Molecular Therapy. 23 (4), 707-716 (2015).
- Bal, N. C., Periasamy, M. Uncoupling of sarcoendoplasmic reticulum calcium ATPase pump activity by sarcolipin as the basis for muscle non-shivering thermogenesis. Philosophical Transactions of the Royal Society B. 375 (1793), 20190135(2020).
- Hicks, C., et al. Body temperature and cardiac changes induced by peripherally administered oxytocin, vasopressin and the non-peptide oxytocin receptor agonist WAY 267,464: a biotelemetry study in rats. British Journal of Pharmacology. 171 (11), 2868-2887 (2014).
- Kasahara, Y., et al. Oxytocin receptor in the hypothalamus is sufficient to rescue normal thermoregulatory function in male oxytocin receptor knockout mice. Endocrinology. 154 (11), 4305-4315 (2013).
- Kasahara, Y., et al. Role of the oxytocin receptor expressed in the rostral medullary raphe in thermoregulation during cold conditions. Frontiers in Endocrinology. 6, 180(2015).
- Yuan, J., Zhang, R., Wu, R., Gu, Y., Lu, Y. The effects of oxytocin to rectify metabolic dysfunction in obese mice are associated with increased thermogenesis. Molecular and Cellular Endocrinology. 514, 110903(2020).
- Scholl, J. L., Afzal, A., Fox, L. C., Watt, M. J., Forster, G. L. Sex differences in anxiety-like behaviors in rats. Physiology & Behavior. 211, 112670(2019).
Access restricted. Please log in or start a trial to view this content.
Перепечатки и разрешения
Запросить разрешение на использование текста или рисунков этого JoVE статьи
Запросить разрешениеСмотреть дополнительные статьи
This article has been published
Video Coming Soon
Авторские права © 2025 MyJoVE Corporation. Все права защищены