Количественная оценка глобальных посттрансляционных модификаций гистонов с помощью внутриядерной проточной цитометрии в изолированной микроглии головного мозга мышей
В этой статье
Резюме
В данной работе описан протокол количественной оценки глобальных модификаций гистонов с помощью внутриядерной проточной цитометрии в изолированной микроглии головного мозга. Работа также содержит протокол выделения микроглии, который использовался для сбора данных.
Аннотация
Контроль экспрессии генов происходит частично за счет модификаций в структуре хроматина, включая добавление и удаление посттрансляционных модификаций хвостов гистонов. Посттрансляционные модификации гистонов (HPTM) могут либо способствовать экспрессии генов, либо их подавлению. Например, ацетилирование остатков лизина гистонового хвоста нейтрализует положительный заряд и уменьшает взаимодействие между хвостом и отрицательно заряженной ДНК. Уменьшение взаимодействия гистонов с ДНК приводит к повышению доступности нижележащей ДНК, что позволяет увеличить доступ к транскрипционному фактору. Метка ацетилирования также служит сайтом узнавания бромдомен-содержащих активаторов транскрипции, что вместе приводит к усилению экспрессии генов. Гистоновые метки могут динамически регулироваться во время дифференцировки клеток и в ответ на различные клеточные среды и стимулы. Несмотря на то, что подходы к секвенированию нового поколения начали характеризовать геномные участки для отдельных модификаций гистонов, только одна модификация может быть исследована одновременно. Учитывая, что существуют сотни различных ВППТ, мы разработали высокопроизводительную количественную оценку глобальных ВППТ, которая может быть использована для скрининга модификаций гистонов перед проведением более обширных подходов к секвенированию генома. Этот протокол описывает метод на основе проточной цитометрии для обнаружения глобальных HPTM и может проводиться с использованием клеток в культуре или изолированных клеток из тканей in vivo . Мы приводим примеры данных изолированной микроглии головного мозга мышей, чтобы продемонстрировать чувствительность анализа для обнаружения глобальных сдвигов в HPTM в ответ на иммунный стимул, полученный из бактерий (липополисахарид). Этот протокол позволяет проводить быструю и количественную оценку HPTM и может быть применен к любому транскрипционному или эпигенетическому регулятору, который может быть обнаружен антителом.
Введение
Эпигенетика изучает механизмы, которые регулируют экспрессию генов, не изменяя лежащую в основе последовательность ДНК. Эпигенетическая регуляция экспрессии генов является динамической в клетках и может обеспечить быструю и скоординированную реакцию на различные стимулы окружающей среды. Динамическая регуляция происходит отчасти за счет изменений структуры хроматина на уровне нуклеосомы, состоящей из белков-гистонов (H2A, H2B, H3, H4), собранных в октамерное ядро, плотно обмотанное ДНК1. Взаимодействие между белками-гистонами и ДНК может контролировать доступ ДНК к механизмам транскрипции, которые, в конечном счете, могут контролировать экспрессию генов и другие аспекты биологии хроматина. Белки гистонов имеют неструктурированные хвосты, которые содержат положительно заряженные остатки, образующие электростатические взаимодействия с отрицательно заряженным каркасом ДНК. Эти взаимодействия приводят к плотной упаковке ДНК и снижению доступности ДНК. Ковалентные модификации хвостов гистонов, называемые посттрансляционными модификациями гистонов (HPTM), могут регулировать эти взаимодействия 3,4. Некоторые из наиболее хорошо охарактеризованных HPTM включают ацетилирование и метилирование хвоста гистонов, которые могут изменять сродство электростатических взаимодействий между хвостами гистонов и ДНК, что приводит к дифференциальному доступу к нижележащей ДНК и набору транскрипционных факторов, которые распознают эти HPTM в определенных участках. HPTM регулируются тремя важными классами ферментов, называемых считывателями, которые распознают, записывающими, которые откладывают, и ластиками, которые удаляют HPTM. Таким образом, рекрутирование или растворение считывающих, записывающих или ластиковых ферментов может, в конечном счете, изменить ландшафт HPTM и управлять структурой и функцией хроматина, делая их регуляцию и считывание необходимыми для понимания клеточной биологии и функции 3,4.
Клетки центральной нервной системы (ЦНС) эпигенетически гибкие, поскольку они изменяют свой транскриптом, чтобы адаптироваться к стимулам окружающей среды. Накопленные данные свидетельствуют о том, что изменения в эпигеноме, такие как метилирование ДНК, некодирующие РНК и HPTM, играют существенную роль в формировании памяти и синаптической функции5. Нарушение динамики HPTM путем манипулирования соответствующими считывателями, писателями или ластиками может блокировать или усиливать ассоциативное обучение и долговременное потенцирование 6,7,8. Микроглия, резидентная иммунная клетка ЦНС, быстро регулирует свой транскриптом в ответ на иммунную стимуляцию посредством динамических изменений в эпигеноме 9,10,11. Этот высокий уровень адаптации к локальной среде мозга затрудняет их изучение в изолированном контексте, поскольку исследования показали, что эпигеном и транскриптом микроглии изменяются уже через несколько часов в питательных средах после удаления из мозговой среды11. Кроме того, поскольку микроглия составляет только 10% клеток головного мозга, измерения, изучающие изменения на уровне всей ткани, не обладают чувствительностью и специфичностью12,13. В результате, микроглия должна быть быстро изолирована для изучения эпигенетических изменений, таких как уровни HPTM, ex vivo.
Методы, обычно используемые для исследования ВПЧ, включают секвенирование хроматина и иммунопреципитации (ChIP-seq) и расщепление под мишенями, а также секвенирование мечения (CUT&Tag-seq)4. Несмотря на то, что эти методы очень специфичны для конкретного ГПТМ и могут информировать о присутствии ВПЧТ в определенном геномном контексте, они могут исследовать только один из многих возможных ВПТТ в рамках одного эксперимента11,14 Поэтому, прежде чем приступать к таким экспериментам, которые требуют значительных затрат времени и денег, очень важно сузить список потенциально интересных ВПТМ для дальнейшего изучения, сначала изучив изменения в глобальных уровни HPTMs. Двумя основными подходами к изучению глобальных уровней HPTM являются иммуногистохимический и вестерн-блоттинг-анализ, но оба подхода являются только полуколичественными, малопроизводительными и требуют большого количества срезов тканей или изолированных клеток15,16. Таким образом, мы стремились разработать высокочувствительный количественный метод, который можно было бы использовать для быстрого изучения глобальных уровней HPTM на уровне отдельных клеток.
Представленный протокол позволяет быстро определять глобальные уровни HPTM с помощью внутриядерной проточной цитометрии. Предыдущие исследования раковых клеток обосновали важность изучения глобальных уровней с клинической точки зрения17,18. Кроме того, в исследованиях обычно используются глобальные уровни в качестве метода скрининга перед оценкой геномной локализации конкретных ВПТМ, представляющих интерес19,20. Что касается микроглии, то оценка глобальных уровней после выделения является сложной задачей из-за низкой клеточной продуктивности; Pan et al. представили глобальные уровни HPTM из изолированной микроглии, в которой микроглия трех животных была объединена для определения уровня белка с помощью вестерн-блоттинга19. Используя наш протокол, мы можем обнаруживать глобальные изменения с гораздо меньшим количеством клеток, что позволяет проводить скрининг нескольких меток для каждого животного и устраняет необходимость объединения образцов.
В данной статье мы опишем протокол для быстрого определения уровней HPTM с помощью количественной внутриядерной проточной цитометрии в изолированной микроглии. Несмотря на то, что для краткости мы уделяем особое внимание количественному определению HPTM, этот протокол можно использовать таким же образом для количественной оценки глобальных уровней ферментов считывателя, записи и ластика. Протокол состоит из двух частей: во-первых, метод выделения микроглии и, во-вторых, метод определения уровня HPTM на основе проточной цитометрии. Метод выделения позволяет получить клетки, которые можно использовать как для выделения РНК, так и для оценки уровня HPTM, что позволяет оценить экспрессию генов и уровни HPTM из одного и того же образца. Кроме того, метод оценки HPTM может быть использован на других типах клеток, как указано в протоколе.
протокол
Все протоколы ухода за животными были одобрены Комитетом по уходу за животными Университета Британской Колумбии в соответствии с рекомендациями Канадского совета по уходу за животными.
1. Пищеварение мозга для выделения микроглии

Рисунок 1: Простая блок-схема протокола. Мышей сначала транскардиально перфузируют HBSS, а мозг препарируют. Затем мозг диссоциирует посредством химического переваривания и механического разрушения, в результате чего образуется гомогенат одной клетки. Обогащенная иммунитетом фракция собирается с помощью прерывистого градиента плотности, после чего клетки окрашиваются на P2RY12. Окрашенные клетки либо 1) сортируются с помощью флуоресцентно-активированной клеточной сортировки (FACS) для проведения анализа РНК или последующего анализа белков и/или 2) фиксируются, пермеабилизируются и окрашиваются на внутриядерные белки. Уровень белка количественно определяют по медиане интенсивности флуоресценции в интересующем канале, определяемой методом проточной цитометрии. Коробки, окрашенные в синий цвет, являются частью шага протокола 1) Пищеварение мозга для выделения микроглии. Прямоугольники, окрашенные в красный цвет, являются частью шага протокола 2) Окрашивание внутриядерным потоком для анализа экспрессии белка. Создано с помощью BioRender.com. Пожалуйста, нажмите здесь, чтобы увидеть увеличенную версию этого рисунка.
{kind=link}
- Приготовление реагентов
ПРИМЕЧАНИЕ: Если вы планируете экстракцию для сбора как РНК, так и клеток для анализа HPTM, обратитесь к разделу 1.7.1 о модификациях, включающих ингибиторы транскрипции и трансляции. Тем не менее, это не требуется, если просто оценить сигнал белка, поскольку клетки в значительной степени находятся в состоянии покоя, когда их держат на льду.- Буфер флуоресцентно-активированной сортировки клеток (FACS) (20 мл на образец): растворите бычий сывороточный альбумин (BSA) в 1x сбалансированном солевом растворе Хэнкса (HBSS) для получения 2% раствора BSA. Растворите ЭДТА до конечной концентрации 1 мМ в 2% растворе БСА. Фильтр стерилизовать с помощью фильтра 0,2 мкм и хранить при температуре 4 °C до 1 недели перед использованием.
- Буфер для пищеварения (1 мл на образец): восстановите флакон папаина в HBSS до конечной концентрации 20 ед/мл в 1 мМ L-цистеина с 0,5 мМ ЭДТА. Активируйте при температуре 37 °C в течение минимум 10 минут или до тех пор, пока ткани не будут готовы к перевариванию. Непосредственно перед использованием добавьте ДНКазу I в активированный раствор папаина до конечной концентрации 200 Ед/мл. Приготовьте его в день эксперимента и не храните.
- Раствор изотонического градиента плотности (5,5 мл на образец): добавьте 10x HBSS в среду с холодным градиентом плотности до конечной концентрации 1x HBSS, в результате чего конечная плотность составит 1,117 г/мл. Вихрь перемешать не менее 30 с перед использованием. Положите на лед до использования.
- Раствор с градиентом плотности 37% (4 мл на образец): Добавьте градиент изотонической плотности к 1x HBSS, чтобы получить конечную концентрацию 37% с конечной плотностью 1,043 г/мл. Добавьте 20 мкл фенольного красителя на каждый мл градиента плотности 37%, чтобы получить розовый раствор для визуализации во время наслоения. Вихрь не менее 30 с перед использованием. Положите на лед до использования.
- Раствор градиента плотности 70% (2 мл на образец): Добавьте изотонический градиент плотности к 1x HBSS, чтобы получить конечную концентрацию 70% с конечной плотностью 1,082 г/мл. Добавьте 5 мкл трипанового синего на каждый мл среды с плотностью 70%, чтобы получить синий раствор для визуализации во время наслоения. Вихрь не менее 30 с перед использованием. Положите на лед до использования.
- Перфузия и рассечение головного мозга
ПРИМЕЧАНИЕ: Протокол перфузии аналогичен протоколу Posel et al., который включает в себя видеоизображение торакотомии мыши, транскардиальной перфузии и удаления мозга21. Здесь мы используем взрослых самцов и самок мышей C57BL/6J (10-15 недель, 20-30 г), но этот протокол может быть использован для выполнения торакотомии для любой мыши. Все процедуры на животных должны быть одобрены комитетом по этике учреждения перед проведением экспериментов.- Анестезия мышей: Обезболивайте мышей 4% изофлураном в 100% кислороде до тех пор, пока они не перейдут плоскость хирургической анестезии, что может быть подтверждено защемлением пальца ноги или отсутствием рефлекса при сильном зажатии ноги мыши. Положите мышь на спину и плотно прижмите ее четыре лапы к хирургической доске для препарирования, расположенной под наклоном в пластиковом лотке, убедившись, что нос закреплен в носовом конусе изофторана. После переноса убедитесь, что животное все еще находится за пределами хирургической плоскости анестезии, прежде чем продолжить.
- Торакотамия мыши: захватите и приподнимите кожу живота с помощью щипцов и сделайте неглубокий разрез через кожу и брюшную стенку, чтобы обнажить ксиф, не повреждая нисходящую аорту или какие-либо нижележащие органы.
- Захватите ксифоз щипцами и сделайте боковые разрезы под грудной клеткой, чтобы обнажить диафрагму и печень. Сделайте аккуратные неглубокие надрезы через диафрагму по всей длине грудной клетки с помощью тонких ножниц и через грудную клетку с помощью тканевых ножниц и прижмите грудину к хирургическому посту возле головы мыши, чтобы обнажить сердце и легкие для транскардиальной перфузии.
- Транскардиальная перфузия: Подготовьте перистальтический перфузионный насос и прикрепите иглу 26,5G к одному концу трубки. Подготовьте трубку для процедуры, вставив один конец трубки во флакон с холодным 1x HBSS и включив насос, чтобы полностью заполнить трубку 1x HBSS.
- Удерживая сердце тупыми щипцами, введите кончик иглы 26,5G с прикрепленной перфузионной трубкой в левый желудочек сердца и сделайте небольшой разрез в правом предсердии. Включите перфузионную помпу, чтобы осторожно провести перфузию мыши со скоростью ~2-4 мл/мин не менее 15-20 мл холодного 1x HBSS.
ПРИМЕЧАНИЕ: Полная перфузия часто назначается, когда печень начинает очищать кровь и становится того же цвета, что и сердце. - Удаление мозга: обезглавьте мышь ножницами для рассечения тканей и сделайте разрез по средней линии кожи головы от шеи до носа. Отклейте кожные лоскуты в стороны, чтобы обнажить череп, и удалите лишнюю ткань и кости на каудальном конце черепа с помощью рассекающих ножниц.
- Осторожно просуньте одно лезвие ножниц под череп в большое затылочное отверстие острой стороной к кости и осторожно срежьте среднюю линию по направлению к носу. Сделайте боковые надрезы как у основания черепа, так и возле носа с помощью препарирующих ножниц. С помощью тонких щипцов прожмите череп от средней линии наружу, чтобы расколоть кусочки черепа и обнажить мозг. Аккуратно приподнимите мозг шпателем и положите на промокательную бумагу.
- Вскрытие мозга: Поместите мозг на лист промокательной бумаги, смоченный 1x HBSS, поверх закрытой чашки Петри, наполненной льдом. Удалите мозжечок и разделите полушария мозга пополам чистым лезвием бритвы.
- Удалите ствол мозга, полосатое тело и белое вещество из каждого полушария, сохранив при этом гиппокамп и кору головного мозга нетронутыми. Переложите полушария, содержащие изолированную ткань коры и гиппокампа, в пробирку объемом 15 мл с 5 мл холодного 1x HBSS и держите на льду.
ПРИМЕЧАНИЕ: Важно выполнить вскрытие как можно быстрее, чтобы ткань оставалась холодной, и между обезглавливанием и окончательным помещением рассеченной ткани в 1x HBSS на лед проходит не более 2 минут. При выделении микроглии от нескольких животных, мозг можно хранить на льду в 1x HBSS в течение ~1 ч, прежде чем приступить к обработке всей когорты животных для пищеварения и т. д.
- Пищеварение и гомогенизация мозга
- Механическая и химическая диссоциация: поместите ткань мозга каждой мыши и 1 мл пищеварительного буфера в отдельные чашки Петри на лед. Чистым лезвием скальпеля тщательно измельчите мозг на мелкие кусочки (<1 мм).
- Отрежьте кончик от пластиковой пипетки для переноса и осторожно переложите каждый из измельченных мозгов в отдельные лунки в пределах 24-луночной пластины на льду. Накройте тарелку прозрачной гибкой пленкой и инкубируйте на льду 30 минут.
ПРИМЕЧАНИЕ: При правильном измельчении мозговая ткань напоминает хорошо измельченный чеснок. - Гомогенизация Dounce: Переложите переваренный мозговой раствор из каждой лунки в отдельные стеклянные гомогенизаторы объемом 7 мл на льду, каждый из которых заполнен 5 мл холодного буфера FACS. Осторожно взбейте каждый мозг сыпучим пестиком (А) примерно 30-40 раз, пока не получите одноклеточную суспензию. После спринцевания пестиком А осторожно подпрыгните плотным пестиком (В) 3-4 раза, чтобы получить одноклеточную суспензию.
ПРИМЕЧАНИЕ: Не проталкивайте пестик более чем на 3/4 вниз, чтобы не раздавить ткань на дне гомогенизатора. Конечный раствор должен быть непрозрачным и молочным.
ПРИМЕЧАНИЕ: При переваривании нескольких мозгов в одном эксперименте засекайте время переноса мозга в буфер FACS так, чтобы каждый образец находился в буфере для переваривания только в течение 30 минут. Чрезмерное переваривание может привести к расщеплению поверхностных белков, снижая связывание антител и передачу сигналов.
- Получение иммунообогащенного фрагмента
- Установление градиента плотности: Перенесите гомогенат из каждого мозга в отдельные полипропиленовые пробирки по 15 мл и добавьте 2,125 мл изотонического градиента плотности и доведите до 8,5 мл с буфером FACS для каждой из них, чтобы получить окончательную концентрацию 25% градиента плотности. Осторожно переверните пробирки объемом 15 мл 20 раз, чтобы тщательно перемешать.
- Используя узкоградуйную трансферную пипетку, аккуратно нанесите 4 мл с градиентом плотности 37% на каждую пробирку, соблюдая осторожность для создания чистых слоев. Переключите пипетки и аккуратно подложите 2 мл с градиентом плотности 70% (Рисунок 2A). Переложите в центрифугу, охлажденную до 4 °C, и отжим при 500 x g в течение 20 мин при нулевой рампе торможения.
- Сбор фрагмента, обогащенного иммунитетом: Используя чистые пипетки для переноса, осторожно аспирируйте миелин из верхней части объема в пробирке объемом 15 мл с помощью чистой пипетки для переноса и выбросьте. Аккуратно соберите верхний фрагмент градиента плотности в чистую полипропиленовую пробирку объемом 15 мл с помощью трансферной пипетки.
- Осторожно соберите обогащенный иммунитетом фрагмент (1,5 мл сверху и 1,5 мл ниже в месте пересечения слоев градиента плотности 70% и 37%) в новую полипропиленовую пробирку объемом 15 мл (рис. 2B). Добавьте 10 мл буфера FACS к обогащенному иммунитетом образцу, чтобы разбавить среду градиента плотности, и осторожно переверните пробирку 20 раз, чтобы тщательно перемешать.
ПРИМЕЧАНИЕ: Поскольку клетки имеют тенденцию прилипать к стенкам пробирки, убедитесь, что вы собрали все клетки в образце на этапах сбора, медленно обводя пипетку вдоль стенок пробирки во время сбора жидкости. - Гранулируйте клетки в образце, обогащенном иммунитетом, центрифугируя пробирки объемом 15 мл в центрифуге с температурой 4 °C при 500 x g в течение 10 мин с нулевым тормозом на спуске. Сразу после окончания отжима осторожно удалите надосадочную жидкость, оставив примерно 300 мкл жидкости в пробирке объемом 15 мл, стараясь не повредить гранулу (которая может быть не видна).
- Соберите надосадочную жидкость в другую пробирку объемом 15 мл, чтобы убедиться, что клетки были гранулированы в отжиме (выбросьте эту фракцию после проверки количества клеток ресуспендированной гранулы). После ресуспендирования клеточной гранулы в объеме 300 мкл с помощью пипетки P1000 подсчитайте клетки гемацитометром, чтобы оценить общий выход клеток.

Рисунок 2: Получение иммунообогащенного фрагмента с помощью прерывистого градиента плотности. (А) Гомогенат мозга изготавливают до среды 25% плотности, подкладывают 4 мл среды плотности 37%, окрашенной в розовый цвет с помощью фенольного красителя и 2 мл среды 70% плотности, окрашенной в синий цвет с помощью трипанового синего. (B) После центрифугирования фракции разделились. Микроглия находится на границе раздела фрагментов сред плотностью 37% и 70%. Фрагмент миелина находится в верхней части пробирки объемом 15 мл и будет отброшен. Верхний фрагмент собирается в качестве резервной копии на случай, если спин не удался, и никакие клетки не будут восстановлены. Если это происходит, градиент можно повторить, используя эту дробь. Фракция, обогащенная иммунитетом, собирается ниже по течению. Нижняя фракция, содержащая эритроциты, остается в пробирке и выбрасывается. (C) Пример рисунка, изображающего полные слои. Создано с помощью BioRender.com. Пожалуйста, нажмите здесь, чтобы увидеть увеличенную версию этого рисунка.
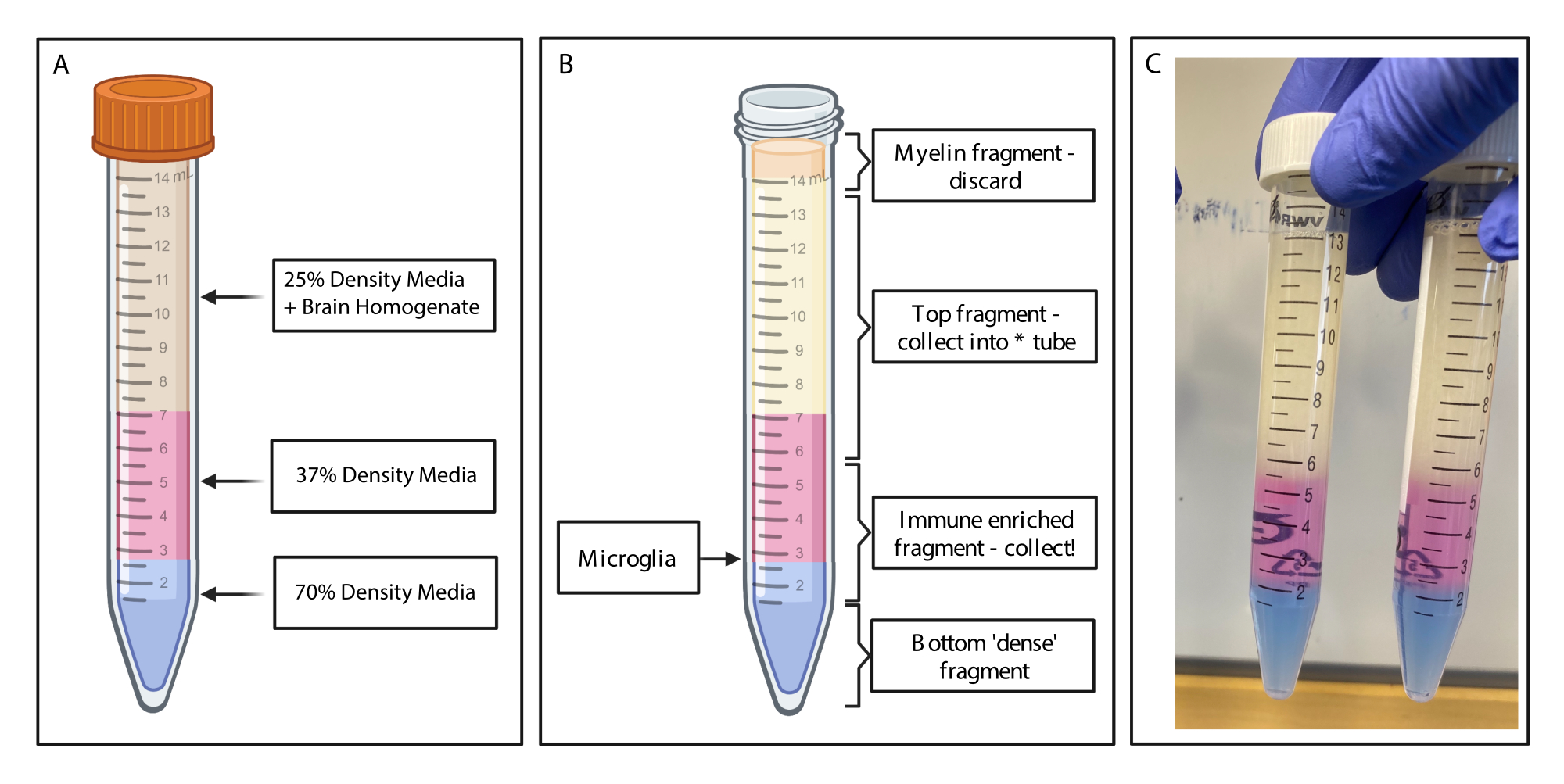{kind=link}
- Окрашивание внеклеточными антителами
- Блокировка: Переместите ячейки на пластину с круглым дном 96 лунок на льду и центрифуге при 500 x g с тормозом, чтобы гранулировать ячейки. Быстро удалите надосадочную жидкость из раковины, щелкнув пластиной, чтобы утилизировать надосадочную жидкость, оставив клеточную гранулу нетронутой на дне лунки.
- Ресуспендировать клетки в 50 мкл буфера FACS с антимышиным реагентом, блокирующим CD16/32 FC-рецепторы, с помощью пипетки P200 (конечная концентрация 10 мкг/мл, коэффициент разведения 1:50) для предотвращения неспецифического связывания антител с моноцитами или другими клетками, содержащими FcR. Выдерживать 10 мин на льду.
- Окрашивание антителами: Приготовьте соответствующий объем 2-кратного основного раствора, содержащего P2RY12-аллофикоцианин (APC; фактор разбавления 1:50, концентрация 4 мкг/мл для конечной луночной концентрации 1:100, концентрация 2 мкг/мл) и фиолетового мертвого красителя 525 (коэффициент разбавления 1:50 для конечной луночной концентрации 1:100). К клеточной суспензии (полученной после блокировки в разделе 1.5.1) добавляют 50 мкл окрашивающей мастер-смеси и инкубируют планшет в течение 30 мин в темноте на льду.
ПРИМЕЧАНИЕ: Для этого протокола мы представляем окрашивание клеток P2RY12. Во-первых, P2RY12 является гомеостатическим маркером микроглии, которая может быть подавлена в определенных контекстах заболевания. Например, мыши модели болезни Альцгеймера 5XFAD имеют пониженный уровень P2RY12, что может затруднитьих идентификацию. Альтернативные красители, которые можно использовать для изоляции, включают Tmem119, Cd11b и CD4523. Во-вторых, конъюгированный флуорохром APC может быть скорректирован в соответствии с желаемой панелью антител. Тем не менее, выбор яркого флуорохрома, такого как APC или PE, поможет обеспечить легкую различимость положительных и отрицательных популяций24. - После окрашивания добавьте 200 мкл буфера FACS непосредственно в каждую лунку для промывки клеток. Отжим при 500 x g при 4 °C, чтобы удалить надосадочную жидкость щелчком. Ресуспендируйте клетки в 200 мкл буфера FACS с помощью пипетки P200, отжим при 500 x g при 4 °C и отсоедините пластину для извлечения буфера из лунок.
- Подготовка регуляторов потока: Перед окрашиванием отделите необходимые объемы ячеек от каждого образца после блокировки на шаге 1.5.1 для требуемого контроля потока.
ПРИМЕЧАНИЕ: Для установки вентилей для каждого эксперимента необходимы элементы управления потоком. Регуляторы дебита могут быть взяты с дополнительного животного или с доли каждой из экспериментальных скважин. При разделении ячеек убедитесь, что на элемент управления назначено достаточное количество ячеек, так как для установки вентили с высокой степенью уверенности требуется 10 000-30 000 ячеек на контрольную группу.- Существует три соответствующих регулятора потока: отсутствие пятен, живой мертвец и контроль изотипа P2RY12. Для борьбы с пятнами не добавляйте антитела. В контроле изотипа P2RY12 обработайте клетки красителем жизнеспособности (1:100) и антителом контроля изотипа, конъюгированным с APC (1:100).
- Чтобы подготовить живой мертвый контроль, аликвотируйте ячейки в отдельную лунку и переместите половину объема ячейки в пробирку объемом 500 мкл. Поместите пробирку объемом 500 мкл в морозильную камеру при температуре -80 °C на 5 минут, а затем поместите в инкубатор при температуре 37 °C на 5 минут, чтобы убить клетки. Возвращают аликвоту мертвых клеток в контрольную скважину для живых мертвых клеток и окрашивают аминосвязывающим красителем жизнеспособности на фиолетовый 525 (коэффициент разбавления 1:100), чтобы пометить мертвые клетки.
ПРИМЕЧАНИЕ: Протокол написан для окрашивания пластин методом фликинга для удаления надосадочной жидкости. Однако для этого необходимо, чтобы надосадочная жидкость была удалена сразу после завершения вращения, и щелчок должен быть выполнен с достаточной силой, чтобы быстро удалить надосадочную жидкость, не повредив гранулу. В качестве альтернативы для окрашивания можно использовать пробирки объемом 1,5 мл, свободные от РНКазы/ДНКазы, со следующими модификациями: Пересадка клеток в пробирку для микроцентрифуги объемом 1,5 мл и гранулирование при 800 x g в течение 5 мин при 4 °C. Отсасывайте надосадочную жидкость с помощью пипеток. Совет: Для обеспечения скорости трансферная пипетка объемом 5 мл с наконечником P200 может быстро и точно аспирировать надосадочную жидкость. При аспирации проверьте наличие гранул. Если гранулы не видны, оставьте 50 мкл надосадочной жидкости и соответствующим образом скорректируйте расчеты. При вымывании антител добавляют дополнительные FACS, чтобы увеличить разведение антител (1000 мкл вместо 200 мкл), чтобы учесть неполное удаление надосадочной жидкости. В зависимости от цитометра используйте пробирки объемом 1,5 мл для сортировки, уменьшая количество необходимых расходных материалов.
- Сортировка FACS по микроглии
- Приготовление: Ресуспендировать каждую лунку в 200 мкл буфера FACS с помощью пипетки P200 и перенести в маркированные пробирки для сортировки потоком и добавить буфер FACS до 500 мкл до концентрации примерно 5 x 105 событий на мл. Хранить на льду в темноте до анализа. Подготовьте пробирки после сортировки, добавив 100 мкл буфера FACS в качестве подушки для клеток в пробирках без РНКазы объемом 1,5 мл.
- Настройки цитометра: Сортировка клеток на сортировщике клеток для проточной цитометрии, оснащенном соплом 100 мкм. Отсортируйте ячейки, используя давление 18-20 фунтов на квадратный дюйм.
- Стробирование: На цитометре затвор для размера клеток с использованием области бокового рассеяния (SSC) по сравнению с высотой прямого рассеяния (FSC) с использованием контроля отсутствия пятен, чтобы помочь различить мусор, поместите SSC-A на логарифмическую ось для визуализации популяции клеток и ворота для тщательного отбора клеток (вентиль S1; Рисунок 3). Чтобы удалить дублеты, постройте график FSC-H и FSC-W и плотно обойдите клеточную популяцию, удаляя любой мусор и дублеты (вентиль S2). Используя контроль изотипа P2RY12, исследуйте клетки в канале APC и установите вентиль для автофлуоресценции для определения клеток P2RY12+. Используя элементы управления «без окрашивания» и «живой мертвец», затвор для клеток, которые не флуоресцентны на фиолетовом 525 нм, как живые клетки.
- Сортировка: Постройте график фиолетового 525 нм по сравнению с APC и определите популяцию, которая является P2RY12+ и живет FMO (MG). Отсортируйте эти ячейки в помеченную пробирку для сортировки (рисунок 3). Окончательный процент сортировки составляет примерно 50% от общего числа событий, при этом большая часть общей потери событий приходится на мусор, удаленный в стробе S1 (~70% событий являются ячейками; Таблица 1).
- Выделение и анализ РНК
- Ингибиторы транскрипции и трансляции: При планировании экстракции РНК, чтобы исключить риск выделения транскриптомных сигнатур, включите ингибиторы трансляции и транскрипции в буферные этапы. Приготовьте коктейль ингибиторов, как описано Marsh et al., включая актиномицин D, анисомицин и триптолид25.
- Препарат ингибитора: Восстановите запасы ингибиторов и храните их следующим образом: Восстановите актиномицин D в диметилсульфоксиде (ДМСО) до 5 мг/мл и храните при -20 °C. Восстановите триптолид в ДМСО до 10 мМ и храните при -20 °C в защищенном от света месте. Анизомицин восстанавливают в ДМСО до 10 мг/мл и хранят при 4 °C, защищая от света. Храните все запасы ингибиторов не более 1 месяца после восстановления.
- Модификации буфера: Добавьте ингибиторы в четыре различных буфера в протокол следующим образом: При выполнении транскардиальной перфузии приготовьте HBSS с актиномицином D (5 мкг/мл, 1:1000 из запаса) и триптолидом (10 мкМ, 1:1000 из запаса). После перфузии мозг транспортируют в лабораторию в HBSS, содержащем актиномицин D (5 мкг/мл, 1:1000 из исходного материала), триптолид (10 мкМ, 1:1000 из исходного материала) и анисомицин (27,1 мкг/мл, 1:368,5 из исходного материала). Приготовьте буфер FACS с актиномицином D (5 мкг/мл, 1:1000 из запаса), триптолидом (10 мкМ, 1:1000 из запаса) и анизомицином (27,1 мкг/мл, 1:368,5 из запаса). Приготовьте буфер для пищеварения с актиномицином D (5 мкг/мл, 1:1000 от запаса), триптолидом (10 мкМ, 1:1000 от запаса) и анисомицином (27,1 мкг/мл, 1:368,5 от запаса). Приготовьте буфер для промывки после сортировки, содержащий HBSS, содержащий актиномицин D (5 мкг/мл, 1:1000 из запаса), триптолид (10 мкМ, 1:1000 из запаса) и анисомицин (27,1 мкг/мл, 1:368,5 из запаса).
ПРИМЕЧАНИЕ: При добавлении ингибиторов обязательно добавляйте их непосредственно перед использованием и защищайте все подготовленные буферы от света во время использования. Избегайте замораживания-оттаивания исходных растворов.
- Промывка после сортировки: Поскольку клетки были отсортированы по 1,5 мл пробирок без РНКазы в буфере FACS, что будет препятствовать выделению РНК, необходимо промыть клетки. Отжим клетки при 1000 x g при 4 °C в течение 5 мин и удалите надосадочную жидкость, оставив примерно 50 мкл жидкости.
- Добавьте 200 мкл 1x HBSS, содержащего актиномицин D (5 мкг/мл, 1:1000 из запаса), триптолид (10 мкМ, 1:1000 из запаса) и анисомицин (27,1 мкг/мл, 1:368,5 из запаса) и тщательно перемешать. Повторите отжим и удалите надосадочную жидкость, оставив 50 мкл жидкости (промывка 1). Добавьте 200 мкл буфера для промывки после сортировки, тщательно перемешайте, повторите отжим и удалите надосадочную жидкость, оставив 25 мкл жидкости (промывка 2).
- Экстракция РНК: Для выделения РНК из клеток микроглии используйте набор для выделения РНК с низким входом для получения высоких и стабильных выходов РНК и баллов RIN выше 9 (см. ниже и Таблицу материалов для рекомендаций по продукту). К клеточной грануле добавьте 350 мкл лизисного буфера из рекомендованного набора + β-меркаптоэтанол (1:100) и хорошо перемешайте.
ПРИМЕЧАНИЕ: При необходимости протокол может быть приостановлен на этом этапе. Образцы могут храниться в лизисном буфере при температуре -80 °C до экстракции РНК. При извлечении РНК после хранения разморозьте лизат на льду и следуйте инструкциям по выделению, описанным в наборе. - Переложите лизат в измельчитель ячеек на основе колонны (рекомендации по продукту см. в таблице материалов ) и центрифугируйте на максимальной скорости при 4 °C в течение 2 минут. Элюируют в воде без РНКазы объемом не менее 14 мкл и определяют соответствующую концентрацию. После этого РНК может быть использована для любого последующего применения.
- Ингибиторы транскрипции и трансляции: При планировании экстракции РНК, чтобы исключить риск выделения транскриптомных сигнатур, включите ингибиторы трансляции и транскрипции в буферные этапы. Приготовьте коктейль ингибиторов, как описано Marsh et al., включая актиномицин D, анисомицин и триптолид25.

Рисунок 3: Стратегия стробирования для сортировки потоком. События стробируются для размера ячейки на SSC-A и FSC-H (S1). Затем ячейки стробируются, чтобы быть синглетными по FSC-H и FSC-W (S2). Синглетные клетки сортируются как живые, если отрицательные на Comp-FL8-A::405-526-52 (фиолетовое мертвое пятно 525) и как P2RY12+, если положительные на Comp-FL32-A::640-671_30 (P2RY12-APC) с использованием контроля изотипа P2RY12. Клетки помечаются как MG и сортируются, если и живые, и P2RY12+. Пожалуйста, нажмите здесь, чтобы увидеть увеличенную версию этого рисунка.
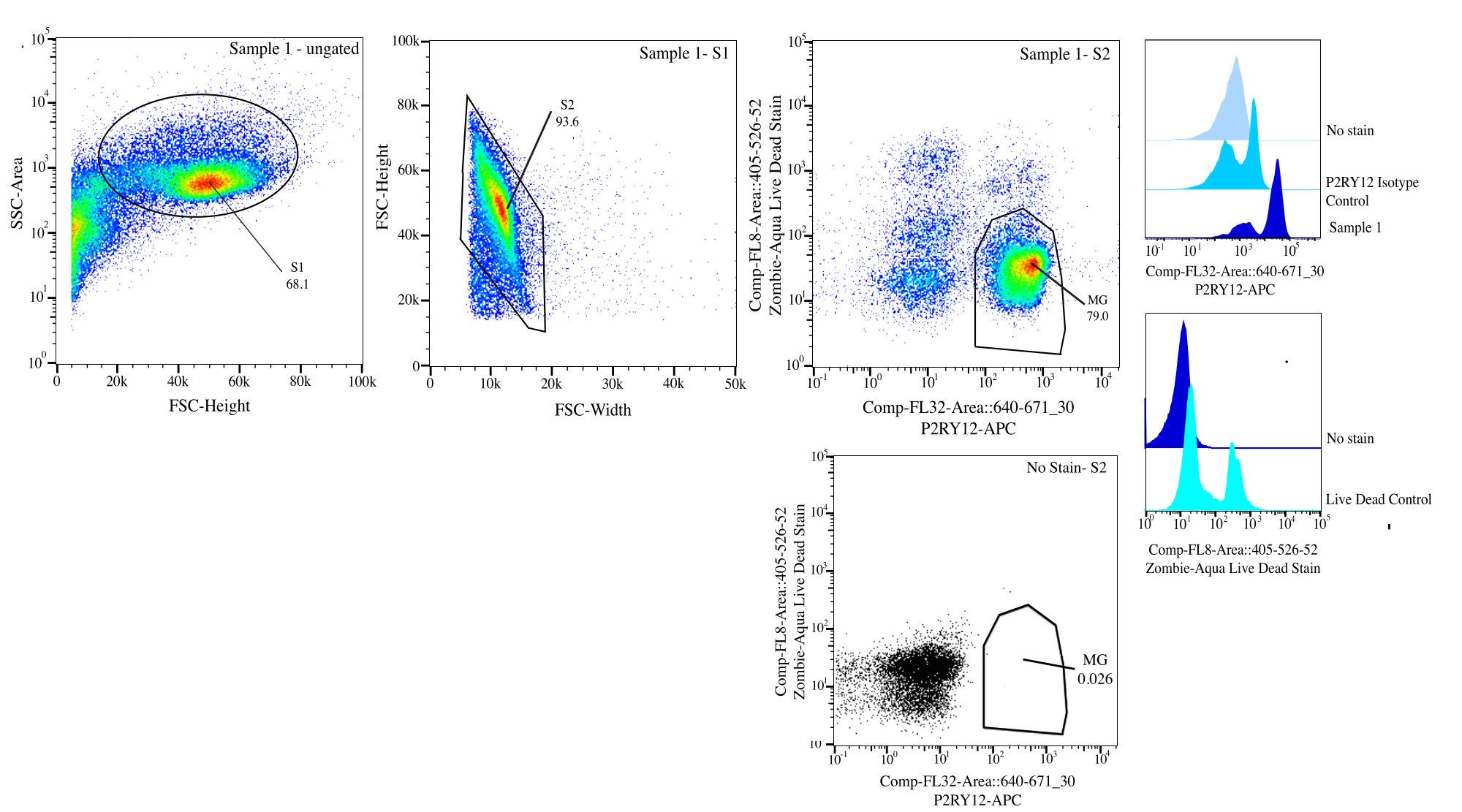{kind=link}
| ЗАКРЫТОЕ НАСЕЛЕНИЕ | Частота родительских операций | Частота Total | Считать |
| С1 | 68.10% | 68.10% | 162186 |
| С2>С1 | 93.59% | 63.70% | 151707 |
| P2Ry12+ (670+) > П2 > П1 | 83.05% | 52.90% | 125986 |
| Прямой эфир (525-) > S2 > S1 | 92.78% | 59.10% | 140752 |
| MG (P2RY12+ Live) >S2>S1 | 78.96% | 50.30% | 119794 |
Таблица 1: Пример таблицы происхождения с процентами стробирования и ожидаемыми номерами событий.
2. Окрашивание внутриядерным потоком для анализа экспрессии белков
ПРИМЕЧАНИЕ: На этом этапе могут быть начаты другие типы клеток, этот протокол протестирован на культивируемых клетках, включая клетки HEK293, клетки микроглии BV2 и микроглию, полученную из IPSC человека.
- Фиксация и окрашивание клеток
ПРИМЕЧАНИЕ: Для следующего протокола используйте набор для внутриклеточного окрашивания, оптимизированный для ядерного окрашивания. Видеть Содержание материалов для получения рекомендаций по продуктам.- Аликвотные внеклеточные окрашенные ячейки из секции 1.5.2 в 96-луночный планшет (5 х 104- 1 х 106 ячеек). Отжмите ячейки в течение 5 минут при 500 x g при 4 °C и щелкните, чтобы удалить буфер FACS.
ПРИМЕЧАНИЕ: Для получения данных с высокой достоверностью медианных уровней следует использовать не менее 10 000 ячеек на лунку. Несмотря на то, что рекомендуемого максимума не существует, лучше всего поддерживать постоянное количество клеток на протяжении всего эксперимента, чтобы убедиться в отсутствии значительного влияния различных коэффициентов вариации (CV). - Фиксация и пермеабилизация: Добавьте 200 мкл 1x фиксированного концентрата и осторожно перемешайте с пипеткой P200 для ресуспендирования клеток. Выдерживают в темноте 45-60 мин. Центрифужную пластину в течение 5 мин при 500 x g при комнатной температуре (RT) и прокрутите, чтобы удалить надосадочную жидкость.
ПРИМЕЧАНИЕ: При необходимости протокол может быть приостановлен на этом этапе. После удаления надосадочной жидкости повторно суспендируйте клетки в буфере длительного хранения для иммунных клеток (см. Таблицу материалов для рекомендаций по продукту). Образцы можно хранить при температуре 4 °C в течение 12-18 ч, защищая от света и накрывая прозрачной пленкой для защиты от буферного испарения. - Добавьте 200 мкл буфера для пермеабилизации 1x в каждую лунку и перемешайте пипетку с P200. Центрифужную пластину в течение 5 мин при 500 x g при RT и щелкните, чтобы удалить надосадочную жидкость. Повторите промывку буфера проницаемости в общей сложности 3 раза.
- Подготовка регуляторов потока: Разделение объема клеток из каждой пробы для требуемого контроля потока (достаточно 10 000-30 000 клеток на контрольную лунку).
- Чтобы подготовить контроль отсутствия пятен, зафиксируйте неокрашенные клетки из сортовых или аликвотных неокрашенных клеток в отдельную лунку, которая не будет получать антитела.
- Чтобы подготовить флуоресцентный минус один (FMO) контроль, аликвотные клетки для каждого из антител на панели, кроме одного в этом канале.
- Для соответствующих каналов включите антитела контроля изотипа в FMO для стробирования. Например, в панели, содержащей P2RY12-APC и H3K27Ac-AlexaFluor568 - должно быть два FMOS: (1) APC-FMO, который содержит только H3K27Ac-AlexaFluor568 и антитела к контролю изотипа P2RY12 и (2) 568-FMO, который содержит только P2RY12-APC и первичный контроль изотипа и 568 вторичный.
ПРИМЕЧАНИЕ: Этот протокол представлен для тестирования одного HPTM, однако могут быть созданы панели, содержащие много HPTM, конъюгированных с различными флуорофорами.
- Окрашивание первичными антителами: Добавьте 50 мкл 1x пермеабилизационного буфера с соответствующей концентрацией первичных антител в каждую лунку. Инкубируют в течение 30 мин при РТ в темноте. Промыть 2 раза 200 мкл 1x буфера проницаемости.
ПРИМЕЧАНИЕ: Концентрация антител, используемых для каждого HPTM, включена в таблицу материалов. Концентрацию определяют путем тестирования различных концентраций антител на культивируемых клетках, обработанных стимулятором, который может вызвать резкое увеличение, например, ингибитором HDAC для меток ацетилирования, и обеспечением того, чтобы как необработанные, так и обработанные клетки находились в пределах диапазона обнаружения (выше контроля изотипа и ниже максимального диапазона обнаружения цитометра). Оптимальная концентрация антител к HPTM должна иметь среднюю медиану интенсивности флуоресценции в канале флуорофора в диапазоне от 5 x 104 до 1 x 105. - Вторичное окрашивание антителами: Блокируйте 200 мкл 1x пермеабилизационного буфера с 2% нормальной ослиной сывороткой (NDS) в течение 10 мин при RT. Вращайте в течение 5 минут при 500 x g при RT и щелкайте, чтобы удалить надосадочную жидкость.
- Добавляют 50 мкл 1x пермеабилизационного буфера с 2% NDS и соответствующей концентрацией вторичных антител и инкубируют в течение 30 мин при RT в темноте. Добавьте 200 мкл 1-кратного буфера пермеабилизации в лунки для разбавления, центрифугируйте планшет в течение 5 минут при 500 x g при RT и щелкните, чтобы удалить надосадочную жидкость. Промывка ячеек 2x с 200 мкл 1x буфера пермеабилизации.
ПРИМЕЧАНИЕ: При необходимости приостановите протокол на этом этапе. Ресуспендировать клетки в 200 мкл буфера длительного хранения для иммунных клеток с помощью пипетки P200 (рекомендации см. в таблице материалов ) и хранить при 4 °C в течение 12-24 ч в защищенном от света месте. - Подготовка к проточной цитометрии: центрифужный планшет в течение 5 мин при 500 x g при RT и щелчок, чтобы удалить надосадочную жидкость. Ресуспендировать клетки в 200 мкл буфера FACS с помощью пипетки P200 для проточной цитометрии. Запечатать прозрачной пленкой для транспортировки к цитометру.
- Аликвотные внеклеточные окрашенные ячейки из секции 1.5.2 в 96-луночный планшет (5 х 104- 1 х 106 ячеек). Отжмите ячейки в течение 5 минут при 500 x g при 4 °C и щелкните, чтобы удалить буфер FACS.
- Проточная цитометрия
- Для анализа предлагаемой панели антител убедитесь, что цитометр оснащен как минимум четырьмя лазерами, включая фиолетовый (405 нм), синий (488 нм), желтый (561 нм) и красный (633 нм). Цитометр нуждается в фильтрах для обнаружения FITC (синий - 525 нм), KRO (фиолетовый - 525 нм), PE (желтый - 585 нм) и APC (красный - 660 нм). Добавьте дополнительные антитела в зависимости от выбранного цитометра.
- Калибровка и стандартизация: В начале каждого эксперимента запускайте радужные флуоресцентные шарики и регулируйте напряжение фотоумножителя (ФЭУ) до тех пор, пока пики шариков не будут сопоставимы с целевыми значениями, полученными для предыдущих экспериментов. Такой метод стандартизации позволяет учесть смещение оборудования с течением времени.
- Компенсация: После того, как напряжение и усиление ФЭУ были установлены для эксперимента, используйте компенсационные шарики, захваченные антителами, чтобы создать компенсационную матрицу для панели антител. Этот расчет гарантирует, что флуорофоры не вносят свой вклад в изменение сигнала в других каналах. Это становится все более необходимым при мультиплексировании нескольких антител.
- Определение размера: На точечной диаграмме постройте график SSC-A на логарифмическом графике и FSC-H на линейном. Удалите мусор и выберите размер ячейки с помощью элемента S1. Выберите синглетные ячейки на точечной диаграмме FSC-W vs FSC-H и вентиль как S2. (Рисунок 4).
- Установка затворов флуорофора: Используя соответствующий FMO для каждого канала флуорофора, установите вентили, чтобы определить, что является положительным сигналом в каждом канале, используя гистограммы с одним параметром (рисунок 4).
- Измерение образцов: Тщательно запишите образцы, используя установленную стратегию литника. Идентифицируют микроглию по сигналу P2RY12+, определяют экспрессию белка в соответствующих каналах только для микроглии.
- Анализ данных проточной цитометрии
- Установка стробов анализа: Используя описанные выше шаги для цитометра в пользовательском интерфейсе программного обеспечения для анализа, используйте те же вентили, которые использовались для записи для анализа.
- Получение значений MFI с помощью программного обеспечения для анализа проточной цитометрии (рекомендации см. в таблице материалов ): Повторите стратегию стробирования цитометра для анализа потока. С помощью функции добавления статистики выберите медиану для интересующей совокупности (например, 568+) на компенсированной высоте канала. Используя редактор таблиц, экспортируйте медианные значения интенсивности флуоресценции (MFI) для соответствующих каналов в электронную таблицу, чтобы продолжить статистический анализ (таблица 2).
ПРИМЕЧАНИЕ: Дополнительный файл S1 включает примеры данных мышей, которым вводили липополисахариды (ЛПС) и фосфатный солевой раствор (PBS), а также пример файла анализа со стратегией стробирования и значениями MFI. - Анализ значений MFI в зависимости от изменения складки белка: После получения значений MFI рассчитайте изменение складки MFI относительно контрольной или необработанной популяции (уравнение 1). Изменение складки MFI отражает изменение уровня белка в складке. Используя значения изменения свертки, оцените изменение экспрессии и рассчитайте статистическую значимость с помощью t-критерия или ANOVA.
 Уравнение 1
Уравнение 1

Рисунок 4: Стратегия стробирования для оценки MFI белка. События стробируются в первую очередь для размера ячейки SSC-A и FSC-H (S1). Затем ячейки стробируются для синглетов на FSC-H и FSC-W (S2). Затем синглетные клетки идентифицируют как микроглию по сигналу P2RY12-APC (APC+) с установлением затвора на основе флуоресценции в контроле APC-FMO, который содержит антитела к контролю изотипа. Затем ячейки стробируются для сигнала H3K27Ac-AlexaFluor568 на Comp-FL5-A::Y610-mCherry. Интенсивность флуоресценции 610+ клеток определяется как показатель экспрессии белка. Пожалуйста, нажмите здесь, чтобы увидеть увеличенную версию этого рисунка.
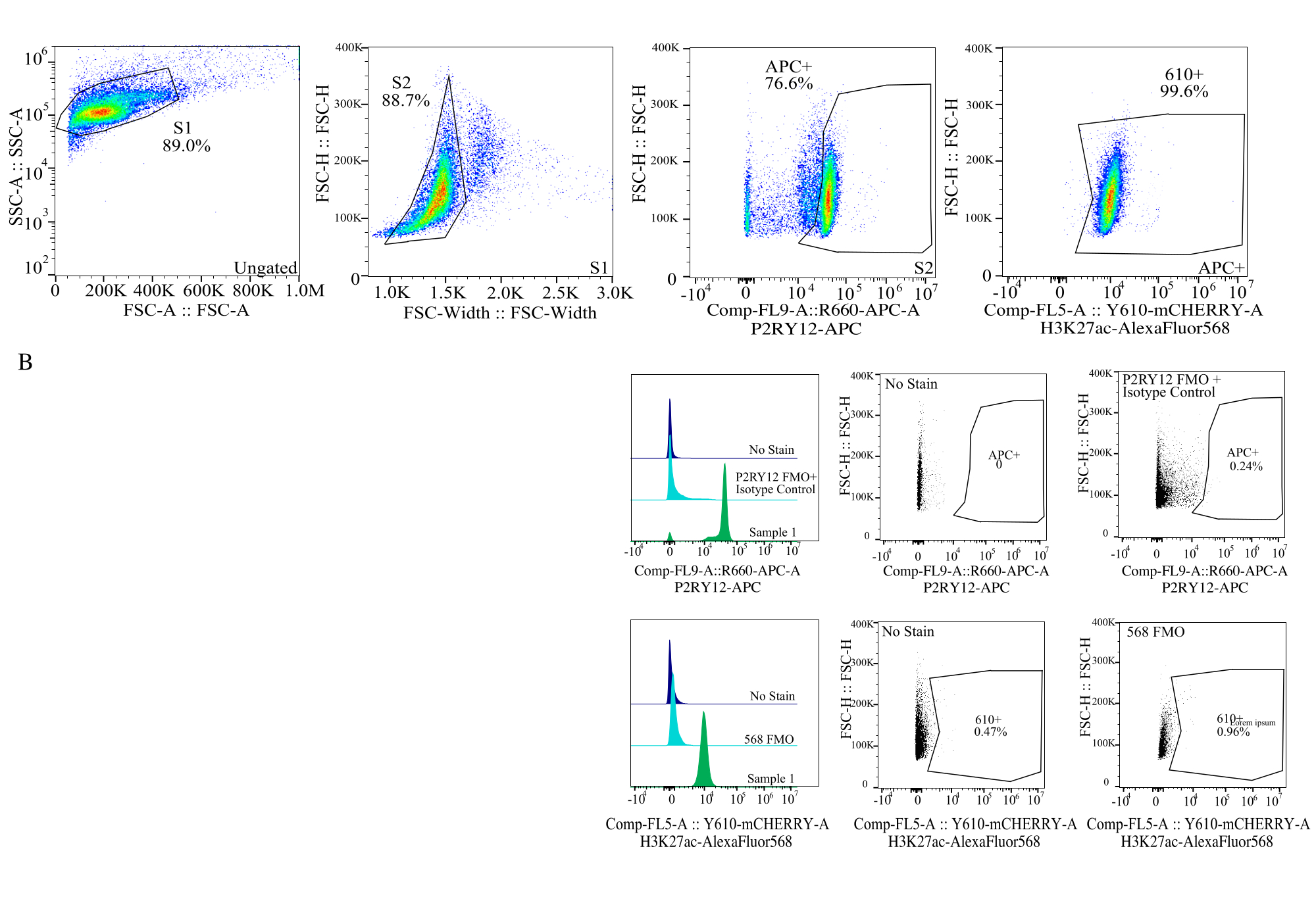{kind=link}
Результаты
Взрослых мышей транскардиально перфузивали и приносили в жертву для выделения микроглии. Микроглия была выделена на льду и окрашена живыми мертвыми антителами P2RY12-APC и фиолетового 525. Клетки, которые были определены как положительные на P2RY12 и отрицательные на живое мертвое пятно фиолетового 525, были отсортированы как живая микроглия. Средний выход микроглии из препарированного мозга мыши составил 1,28 x10,5 ± 0,05 (среднее ± стандартной ошибке среднего (SEM), N=100). Нет различий в выходе микроглии от самок (1,25 x 105 ± 0,09 [среднее ± SEM, N=46) и самцов (1,32 x 105 ± 0,07 [среднее ± SEM, N=54]) мышей (t(98)=0,6365, p=0,526). При выделении из определенных областей мозга средний выход микроглии из коры головного мозга мыши составляет 8,3 х 104 ± 0,08 (среднее ± СЭМ, N=15) и из гиппокампа мыши 4,1 х 104 ± 0,02 (среднее ± СЭМ, N=16). Как и ожидалось, существует существенная разница в выходе микроглии из каждой области мозга (F(2, 128)=25,25, P<0,0001). После выделения микроглии РНК выделяли из изолированных клеток с помощью набора для выделения РНК с низким входом. Показатель целостности РНК (RIN) постоянно был выше 9,0 (9,62 ± 0,05), а средний выход РНК на клетку составлял 0,25 ± 0,01 пг (среднее ± SEM, N=32; Дополнительный файл S2).
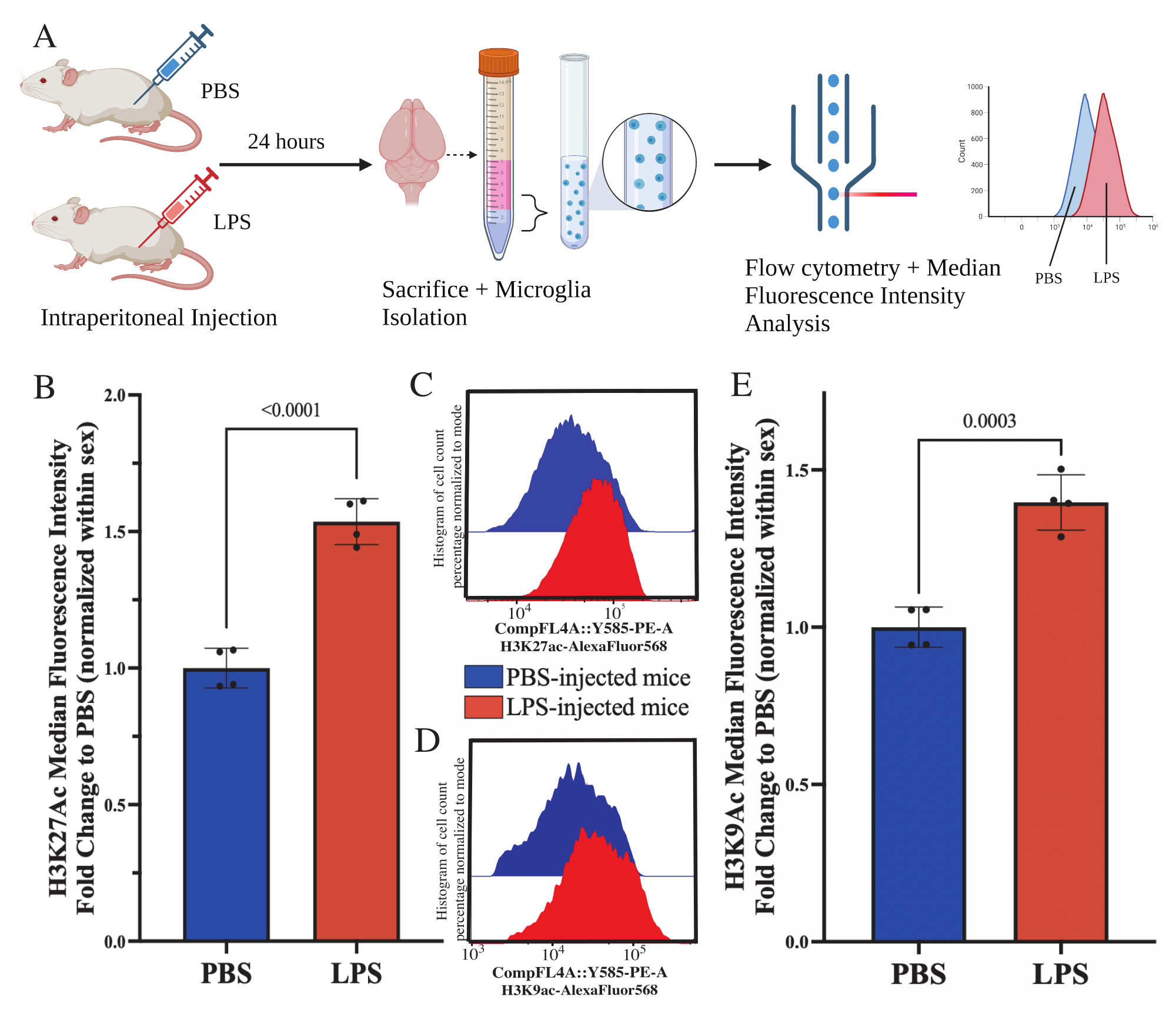
Взрослым мышам внутрибрюшинно вводили 1 мг/кг липополисахарида (ЛПС) за 24 ч до жертвоприношения. Мышей транскардиально перфузировали HBSS и выделяли микроглию из всего мозга по описанному протоколу (рис. 5А). Для каждого окрашивания было выделено 20 000-30 000 клеток на каждую панель антител. С помощью проточной цитометрии оценивали глобальные уровни ацетилирования гистон-3-лизина 27 (H3K27Ac) в изолированной микроглии. У самцов и самок мышей лечение ЛПС индуцировало увеличение H3K27Ac при нормализации MFI в пределах пола (t(6)=9,676, p<0,0001; Рисунок 5Б). При исследовании гистограмм окрашенных клеток популяции остаются нормально распределенными с аналогичными вариациями; однако клетки перешли к усиленной флуоресценции, что привело к увеличению MFI (рис. 5C). При исследовании H3K9Ac при том же лечении наблюдается аналогичное увеличение H3K9Ac (t(6)=7,299, p=0,0003; Рисунок 5D,E), однако кратное изменение ЛПС относительно ПБС сигнала H3K9Ac меньше, чем сигнала H3K27Ac.

Рисунок 5: Глобальные изменения ацетилирования гистонов в изолированной микроглии. (A) Мышам вводят внутрибрюшинно фосфатный буферный физиологический раствор (PBS) или 1 мг/кг липополисахарида (LPS) за 24 ч до жертвоприношения. Микроглии собирают из фракции, обогащенной иммунным иммунитетом, и фиксируют для проточной цитометрии и оценки глобальной посттрансляционной модификации гистонов. Медиана интенсивности флуоресценции оценивается как показатель экспрессии белка. Создано с помощью BioRender.com. (B) Глобальные уровни H3K27Ac повысились в ответ на лечение ЛПС. Изменение складки на PBS нормализовано в эксперименте и сексе. Непарный двусторонний t-критерий, t(6)=9,676, p<0,0001. Гистограмма изображает среднее значение ± SEM. N=8 животных; 2 на условие в 2 независимых экспериментах. (C) Пример гистограммы, показывающей сдвиг интенсивности флуоресценции H3K27Ac. На модальном рисунке изображены гистограммы мышей, которым вводили PBS, и мышей, которым вводили LPS. (D) Пример гистограммы, показывающей сдвиг интенсивности флуоресценции H3K9Ac. На модальном рисунке изображены гистограммы мышей, которым вводили PBS, и мышей, которым вводили LPS. (E) Глобальные уровни H3K9Ac повышались в ответ на лечение ЛПС. Изменение складки на PBS нормализовано в эксперименте и сексе. Непарный двусторонний t-критерий, t(6)=7,299, p=0,0003. Гистограмма изображает среднее значение ± SEM. N=8 животных; 2 на условие в 2 независимых экспериментах. Пожалуйста, нажмите здесь, чтобы увидеть увеличенную версию этого рисунка.
{kind=link}
Чтобы подтвердить, что описанный метод сопоставим с другими ранее использовавшимися методами количественной оценки глобальных модификаций гистонов, мы стремились использовать иммуноблот в качестве сравнительного инструмента. Тем не менее, выход изолированной микроглии слишком низок, чтобы его можно было оценить. Поэтому мы использовали культивируемые клетки BV2 для сравнения метода внутриклеточной проточной цитометрии с методом вестерн-блоттинга (WB). Клетки BV2 выращивали в полной среде (DMEMF12, 10% FBS, 1x пенициллин/стрептамицин и 1x L-глутамин) при 37 °C, 5% CO2. Клетки пассировали 0,25% трипсин-ЭДТА и покрывали при плотности 250 000 клеток/лунку и обрабатывали в восстановленных сывороточных средах (DMEM F12, 2% FBS, 1x пенициллин/стрептамицин и 1x L-глутамин) и давали восстановиться в течение 12 ч при 37 °C, 5% CO2. Клетки обрабатывали 25 нг/мл ЛПС в течение 24 ч до фиксации, как описано выше, или лизиса с помощью буфера для лизиса WB. Сигнал H3K27Ac осуществлялся обоими методами, при этом в качестве регулятора нагрузки для ББ использовался GAPDH. Для каждой группы был проведен анализ нормированной интенсивности флуоресцентного излучения в сравнении с контролем ФБС (рис. 6А). При изучении изменения нормализованного сигнала H3K27Ac по ББ наблюдалось увеличение состояния, обработанного ЛПС, в 1,527 раза по сравнению с контролемH2O, что было определено как значимое по непарному t-критерию (t=3,024, df=5; p=0,0293). При изучении изменения с помощью проточной цитометрии наблюдалось увеличение состояния, получавшего ЛПС, в 1,482 раза, которое было определено как значимое (t=7,843, df=10; p<0,0001). Используя 2-факторный ANOVA для сравнения методов, было определено значимое влияние лечения (F(1,15)=45,21,p<0,0001), но не метода (F(1,15)=0,05545, p=0,8697) или взаимодействия (F(1,15)=0,02785, p=0,8697). Кроме того, мы убедились в отсутствии изменений в уровнях гистонов H3 как при вестерн-блоттинге, так и при проточной цитометрии, поскольку 2-сторонняя ANOVA не выявила значимого эффекта лечения ЛПС (F(1,7)=0,02170, p=0,8870), метода (F(1,7)=0,01191, p=0,9162) или взаимодействия (F(1,7=0,01191, p=0,9162; Рисунок 6Б). Также показаны примеры блотов и сдвигов гистограммы для этих данных (рис. 6C,D).

Рисунок 6: Сравнение методов количественной оценки глобального изменения модификации гистонов между проточной цитометрией и вестерн-блоттингом. (A) Клетки BV2 обрабатывают 25 нг/мл липополисахаридом (ЛПС) или H2O в течение 24 ч перед анализом. Интенсивность флуоресценции H3K27Ac изображается как изменение складки в контроле транспортного средства, фосфатно-солевого буфера (PBS), как для проточной цитометрии, так и для вестерн-блоттинга. 2-сторонняя ANOVA выявила значимый эффект лечения ЛПС (F(1,15)=45,21, p<0,0001), но не метода (F(1,15)=0,05545, p=0,8697) или взаимодействия (F(1,15)=0,02785, p=0,8697). Поправка Тьюки для проверки множественных гипотез была применена к остаткам. * представляет 0,0332, ** представляет 0,0021. (B) Интенсивность флуоресценции для гистона H3 изображается как изменение складки PBS как для проточной цитометрии, так и для вестерн-блоттинга. Двухсторонний ANOVA не выявил значимого эффекта лечения ЛПС (F(1,7)=0,02170, p=0,8870) или метода (F(1,7)=0,01191, p=0,9162) или взаимодействия (F(1,7=0,01191, p=0,9162). (C) Показаны примеры блотов и (D) сдвигов проточной цитометрии. Размер гистограммы нормируется в процентах в зависимости от количества клеток, присутствующих в режиме интенсивности флуоресценции. Гистограмма изображает среднее значение SEM. n=2 независимых эксперимента, по 2 на каждое условие на эксперимент. Пожалуйста, нажмите здесь, чтобы увидеть увеличенную версию этого рисунка.
{kind=link}
В совокупности эти результаты показывают, что этот метод может быть использован для количественной оценки глобальных уровней HPTM в изолированной микроглии. Кроме того, было показано, что этот метод сопоставим с предыдущими методами, но требует гораздо меньших затрат на клетки. Кроме того, несмотря на то, что данная методика не показана, при надлежащей компенсации она может быть использована с несколькими антителами на одной и той же панели для оценки различных HPTM.
Дополнительный файл S1: Примеры файлов анализа. Этот файл содержит файл анализа wsp и 7 файлов fcs, включая файлы без пятен, P2RY12FMO, 568FMO, двух животных, обработанных PBS, и двух животных, обработанных LPS, окрашенных H3K27Ac. Цель этого файла — продемонстрировать анализ и построение эксперимента, которые могут показать, как выглядит успешный эксперимент. Пожалуйста, нажмите здесь, чтобы скачать этот файл.
Дополнительный файл S2: Данные изоляции. Прилагаемый файл содержит соответствующие данные после сортировки микроглии, которые содержат выход микроглии и РНК из описанного протокола. Пожалуйста, нажмите здесь, чтобы скачать этот файл.
| ЗАКРЫТОЕ НАСЕЛЕНИЕ | Частота родительских операций | Частота Total | Считать |
| С1 | 89.00% | 89.00% | 25672 |
| С2>С1 | 88.73% | 78.97% | 22779 |
| APC+ > S2 > S1 | 76.61% | 60.50% | 17452 |
| 610+ > APC+ > S2 > S1 | 99.56% | 60.24% | 17376 |
Таблица 2: Пример диаграммы родословной показывает процентное соотношение и количество событий, необходимых для точного обнаружения белка.
Обсуждение
Представленный протокол позволяет количественно оценить глобальные уровни HPTM с помощью проточной цитометрии. Несмотря на то, что этот протокол представляет собой новый метод, в предыдущих исследованиях проводилась количественная оценка белков с использованием аналогичногоподхода. Предыдущие методы, используемые для оценки глобальных уровней ВПЧМ, включали иммуногистохимию и вестерн-блот 16,17,19,20. Представленный метод, основанный на проточной цитометрии, является легко поддающимся количественной оценке, в то время как вестерн-блот и иммуногистохимия являются полуколичественными и имеют меньшую пропускную способность. Вестерн-блот основан на лизисе клеток и, таким образом, требует как нормализации белка, так и белка, контролирующего нагрузку, который, как предполагается, не изменяется в экспериментальном условии27. Иммуногистохимия является полуколичественной и очень низкопроизводительной, так как трудно количественно оценить количество белка без исследования на уровне одной клетки16. Аналогичным образом, для изолированной микроглии использование метода проточной цитометрии имеет преимущество из-за ограниченного выхода, поскольку вестерн-блот требует гораздо большего количества белка19. Требования к низкому количеству клеток позволяют запускать несколько панелей окрашивания от одного и того же животного.
Однако, как и в случае с любым другим методом, у этого метода есть ограничения, включая стоимость и доступность антител, поскольку не все антитела хорошо работают в условиях проточной цитометрии. Кроме того, по сравнению с иммуноблоттингом, концентрация необходимых антител значительно выше. В то время как мультиплексирование позволяет использовать несколько антител на одной и той же панели клеток, клетки не могут быть удалены от антител после анализа, что ограничивает использование клеток до одного на каждый вид антител. Это отличается от иммуноблоттинга, при котором один и тот же блот может использоваться многократно. Однако, в зависимости от наличия антител и количества каналов обнаружения на цитометре, можно будет исследовать до десятка меток одновременно.
Текущий метод охватывает только глобальные уровни экспрессии белка, а не конкретное местоположение генома, и изменения в глобальных уровнях могут не отражать изменения в отдельных геномных локусах. Аналогичным образом, отсутствие изменений на глобальных уровнях может не означать, что геномные локусы не претерпевают изменений, просто глобальные изменения в сумме не приводят к отсутствию различий между методами лечения. Таким образом, этот метод предназначен для использования в качестве скрининга для выявления HPTM, представляющих интерес для геномного анализа. Кроме того, этот метод не позволяет проводить сравнение между различными белковыми марками, за исключением случаев, когда они оцениваются как изменение свертки по сравнению с контролем. Таким образом, это ограничено по сравнению со стандартным методом, основанным на кривых, таким как ИФА для определения белка.
Представленный протокол предлагает стратегию выделения живой микроглии мозга. Этот протокол основан на экспрессии белка P2RY12 для выделения микроглии. Тем не менее, P2RY12 является гомеостатическим маркером микроглии и может быть подавлен в моделях заболеваний, таких как 5XFAD22. Таким образом, при использовании модельного животного обязательно выбирайте другие маркерные белки, такие как TMEM119, CD11b или CD45, чтобы помочь в выделении микроглии23. Точно так же мы представляем этот протокол как изоляцию от гиппокампа и/или коры головного мозга. Этот протокол будет работать для изоляции микроглии от других областей мозга, включая области белого вещества, однако для получения достаточного количества микроглии может потребоваться несколько животных в зависимости от размера интересующих областей.
Представленный протокол может надежно изолировать живую микроглию головного мозга, но есть несколько этапов, описанных ниже, на стадии изоляции, которые могут снизить выход клеток при неправильном выполнении.
Перфузии по этому протоколу приводят к более высокому проценту микроглии в обогащенном иммунитетом фрагменте, что сокращает время нахождения в сортировщике. Однако перфузия не требуется, и при необходимости могут быть использованы другие методы эвтаназии.
Во время выделения микроглии миелин должен быть полностью удален. Проточные цитометры полагаются на то, что клетки могут перемещаться по узким трубкам в быстром темпе. Из-за своей вязкости и склонности к слипанию миелин вызывает проблемы с цитометрами, часто вызывая засорение, которое может как повредить оборудование, так и разрушить образец, что резко снижает выход продукта. Будьте осторожны, чтобы удалить весь миелин во время сбора обогащенных иммунитетом фрагментов, чтобы избежать проблем в дальнейшем.
Окрашивание пластин в сравнении с окрашиванием в пробирках: В этом протоколе мы описали два варианта окрашивания ячеек либо в пробирках объемом 1,5 мл, либо в 96-луночном планшете. Сценарий использования каждого из них зависит от эксперимента; Однако, как правило, окрашивание в пробирку имеет меньший риск повлиять на выход продукта, чем окрашивание в пластину, так как при неправильном окрашивании существует риск потери клеток. Окрашивание пластин происходит намного быстрее, так как аспирация надосадочной жидкости для каждой пробирки занимает много времени. Перед фиксацией (для сортировки и т.д.) используйте окрашивание пробирок, чтобы максимизировать выход и снизить риск потери. Тем не менее, при анализе HPTM после того, как клетки зафиксированы для внутриядерного окрашивания, гранула более стабильна, и снижается риск потери при выщелкивании.
Установление прерывистого градиента плотности: При установлении слоев необходимо правильно расположить слои для получения фракции, обогащенной иммунитетом. Если слои нарушены или перемешаны и выглядят мутными, клетки не будут сортироваться в нужном месте, и будет трудно получить обогащенную иммунитетом клеточную фракцию. В этом случае отжим со средой плотности, чтобы удалить миелин, а затем соберите все оставшиеся фракции, разбавьте 3 мл буфера FACS до 1 мл среды плотности и хорошо перемешайте (для этого потребуется несколько пробирок). Отжим при 500 x g в течение 10 минут с включенным тормозом 0. Выбросьте надосадочную жидкость, оставив только ~300 мкл раствора. Соберите всю пробу и покрасьте. Это приведет к уменьшению процента сортировки и большему количеству времени, проведенного за цитометром, но выход все равно может быть сопоставимым.
При использовании метода изоляции полезно иметь возможность собирать клетки для РНК и для оценки HPTM из одного и того же мозга мыши. В этой ситуации, после сортировки живой микроглии, клетки можно разделить, чтобы выделить часть для оценки РНК (минимальное входное количество клеток для получения достойного выхода РНК составляет 75 000 клеток) и часть для дальнейшего анализа методом проточной цитометрии (минимум 10 000 клеток на лунку для хорошего определения MFI). В этом случае требуется сортировка проточным цитометром. Однако, если вы только планируете использовать клетки для анализа HPTM, сортировка не требуется, и иммунная фракция может быть окрашена антителами P2RY12 и HPTM. Стробирование на цитометре может быть настроено на микроглию P2RY12+, как это было бы сделано для сортировки потока, чтобы анализировать только сигнал HPTM в микроглии. Исключение сортировки позволяет сделать протокол более быстрым и экономичным. Кроме того, при оценке HPTM из культивируемых клеток достаточно начать с протокола окрашивания, и антитела к клеточным маркерам не требуются, как показано на рисунке 6. Протокол оценки HPTM может быть использован для многих типов клеток, включая культивируемые, первичные клетки и клетки, полученные из IPSC.
Наконец, несмотря на то, что мы представили только два потенциальных применения микроглии после выделения, существует множество других, включая эпигенетические методы, такие как ChIP, CUT&Tag и CUT&RUN. В случае геномных эпигенетических методов, где представляет интерес характеристика изменений в конкретных локусах, следует выбирать специфические ингибиторы для записи и стирания хроматиновых меток11 , адаптированные к экспериментам, чтобы гарантировать, что любые профилированные эпигенетические модификации микроглии не являются техническими артефактами от каких-либо этапов процедуры выделения, таких как ферментативное расщепление. При оценке изменений глобальных уровней эпигенетических маркеров, например, с помощью количественной проточной цитометрии, не ожидается, что любые изменения, вызванные процедурой, будут настолько большими, чтобы их можно было обнаружить на глобальном уровне.
В целом, рассмотренные методы представляют собой новый одноклеточный метод для количественной оценки глобальных уровней модификаций гистонов и других эпигенетических изменений с помощью проточной цитометрии. Показано, что данный метод достаточно чувствителен для выявления глобальных изменений маркера-энхансера H3K27ac в микроглии в ответ на ЛПС in vivo. Это согласуется с предыдущим ChIP-секвенированием H3K27ac после стимуляции ЛПС, демонстрирующим резкое ремоделирование энхансеров, реагирующих на ЛПС28. Применение этого метода позволит исследовать глобальные эпигенетические изменения в различных типах клеток мозга в процессе развития и заболевания.
Раскрытие информации
Авторам нечего раскрывать.
Благодарности
Спасибо Yanyang Bai за помощь с иммуноблоттингом на рисунке 5. Эта работа была поддержана Канадскими институтами исследований в области здравоохранения [CRC-RS 950-232402 to AC]; Совет по естественным наукам и инженерным исследованиям Канады [RGPIN-2019-04450, DGECR-2019-00069 в AC]; Благотворительный фонд Шотландского обряда [21103 в AC] и Фонд Brain Canada [AWD-023132 в AC]; Стипендия для выпускников Университета Британской Колумбии для аборигенов (6481 в MT); Стипендия для выпускников Британской Колумбии (6768 для MT); Премия Canadian Open Neuroscience Platform Student Scholar Award (10901 для JK); Четырехлетняя докторская стипендия Университета Британской Колумбии (6569 для JK). Спонсоры не играли никакой роли в планировании исследования, сборе и анализе данных, принятии решения о публикации или подготовке рукописи.
Материалы
| Name | Company | Catalog Number | Comments |
| 0.5M EDTA | Invitrogen | AM9260G | |
| 15 mL Falcon Centrifuge Tubes, Polypropylene, Sterile | Falcon | 352196 | |
| 24-well Clear Not Treated Plates | Costar | 3738 | |
| 2-Mercaptoethanol | Gibco | 21985023 | |
| 96 Well Clear Polystyrene Microplate, clear round bottom, non treated surface | Corning | 3788 | |
| Acetyl Histone 3 K9 (C5B11) | Cell Signalling Technology | 9649S | Dilution: 1:100 |
| Acetyl Histone H4 K8 (2594) | Cell Signalling Technology | 2594S | Dilution: 1:100 |
| Acetyl-Histone H3 K27 (D5E4) | Cell Signalling Technology | 8173S | Dilution: 1:100 |
| Acetyl-Histone H3 Lys27 (MA523516) | Invitrogen | MA523516 | Dilution: 1:100 |
| Actinomycin D | New England Biolabs | 15021S | |
| Anisomycin | New England Biolabs | 2222S | |
| Anti-Histone H3 (tri methyl K4) | Abcam | ab213224 | Dilution: 1:100 |
| Anti-Lactyl-Histone H4 (Lys 12) Rabbit mAb | PTM Biolabs | PTM-1411RM | Dilution: 1:250 |
| Anti-L-Lactyllysine Rabbit pAb | PRM Biolabs | PTM-1401RM | Dilution: 1:250 |
| Apc anti-P2RY12 Antibody, Clone: S16007D | BioLegend | 848006 | |
| BSA | Tocris | 5217 | |
| Cyto-Last Buffer | BioLegend | 422501 | |
| dimethylsulfoxide, sterile | Cell Signalling Technology | 12611S | |
| DNAse I | STEMCELL Technologies | 07900 | |
| Donkey Anti Mouse AlexaFluor488 | Jackson ImmunoResearch | 715-546-150 | Dilution: 1:500 |
| Donkey Anti Rabbit AlexaFluor488 | ABclonal | AS035 | Dilution: 1:500 |
| Donkey Anti Rabbit AlexaFluor568 | Invitrogen | A10042 | Dilution: 1:500 |
| Donkey Anti Rabbit Brilliant Violet 421 | BioLegend | 406410 | Dilution: 1:500 |
| Fisherbrand Disposable Graduated Transfer Pipettes | Fisherbrand | 13-711-9AM | |
| Fisherbrand Disposable PES Filter Unit, 250mL | Fisherbrand | FB12566502 | |
| H3K18ac Polyclonal Antibody | Invitrogen | 720095 | Dilution: 1:100 |
| HBSS (10X), no calcium, no magnesium, no phenol red | Gibco | 14185052 | |
| HBSS, no calcium, no magnesium, no phenol red | Gibco | 14175103 | |
| Histone 3 Trimethyl K27 (ab6002) | Abcam | ab6002 | Dilution: 1:100 |
| KONTES Dounce Tissue Grinders 125mm 7mL | VWR | 885300-0007 | |
| Lactyl-Histone H3 (Lys 18) Rabbit mAb | PTM BIolabs | PTM-1406RM | Dilution: 1:250 |
| Lipopolysacharide | Sigma-Aldrich | L5418 | |
| Normal Donkey Serum | Jackson ImmunoResearch | 017-000-121 | |
| OneComp eBeads Compensation Beads | Invitrogen | 01-1111-41 | |
| PDS Kit, Papain Vial - Worthington Biochemical | Cedarlane | LK003178 | |
| Percoll | Sigma-Aldrich | GE17-0891-02 | |
| Phenol Red | VWR | RC57004 | |
| QIAshredder | Qiagen | 79656 | |
| Rainbow Fluorescent Particles, 1 peak (3.0-3.4 uM - Mid Range Intensity | BioLegend | 422905 | |
| RNase-free Microfuge Tubes, 1.5 mL | Invitrogen | AM12400 | |
| Rneasy Plus Micro Kit | Qiagen | 74034 | |
| Round Bottom Polypropylene Tubes with Caps, 5 mL | Corning | 352063 | |
| Triptolide | New England Biolabs | 97539 | |
| True Nuclear Transcription Factor Buffer Set | BioLegend | 424401 | |
| TruStain FcX PLUS (anti-mouse CD16/32) Antibody | BioLegend | 156604 | |
| Trypan Blue | VWR | 97063-702 | |
| Zombie Aqua Fixable Viability Kit | BioLegend | 423102 |
Ссылки
- Miller, J. L., Grant, P. A. The Role of DNA Methylation and Histone Modifications in Transcriptional Regulation in Humans. Epigenetics: Development and Disease. 61, 289-317 (2013).
- Kouzarides, T. Chromatin Modifications and Their Function. Cell. 128 (4), 693-705 (2007).
- Bannister, A. J., Kouzarides, T. Regulation of chromatin by histone modifications. Cell Research. 21 (3), 381-395 (2011).
- Barski, A., et al. High-resolution profiling of histone methylations in the human genome. Cell. 129 (4), 823-837 (2007).
- Vogel Ciernia, A., LaSalle, J. The landscape of DNA methylation amid a perfect storm of autism aetiologies. Nature Reviews. Neuroscience. 17 (7), 411-423 (2016).
- Keiser, A. A., et al. Systemic HDAC3 inhibition ameliorates impairments in synaptic plasticity caused by simulated galactic cosmic radiation exposure in male mice. Neurobiology of Learning and Memory. 178, 107367 (2021).
- McQuown, S. C., et al. HDAC3 is a critical negative regulator of long-term memory formation. The Journal of Neuroscience: The Official Journal of the Society for Neuroscience. 31 (2), 764-774 (2011).
- Barrett, R. M., et al. Hippocampal Focal Knockout of CBP Affects Specific Histone Modifications, Long-Term Potentiation, and Long-Term Memory. Neuropsychopharmacology. 36 (8), 1545-1556 (2011).
- Datta, M., et al. Histone Deacetylases 1 and 2 Regulate Microglia Function during Development, Homeostasis, and Neurodegeneration in a Context-Dependent Manner. Immunity. 48 (3), 514.e6-529.e6 (2018).
- Belhocine, S., et al. Context-dependent transcriptional regulation of microglial proliferation. Glia. 70 (3), 572-589 (2022).
- Gosselin, D., et al. An environment-dependent transcriptional network specifies human microglia identity. Science (New York, N.Y.). 356 (6344), eaal3222 (2017).
- Kettenmann, H., Hanisch, U. -. K., Noda, M., Verkhratsky, A. Physiology of Microglia. Physiological Reviews. 91 (2), 461-553 (2011).
- Sullivan, O., Ciernia, A. V. Work hard, play hard: how sexually differentiated microglia work to shape social play and reproductive behavior. Frontiers in Behavioral Neuroscience. 16, 989011 (2022).
- Das, P. M., Ramachandran, K., vanWert, J., Singal, R. Chromatin immunoprecipitation assay. BioTechniques. 37 (6), 961-969 (2004).
- Mahmood, T., Yang, P. C. Western blot: technique, theory, and trouble shooting. North American Journal of Medical Sciences. 4 (9), 429-434 (2012).
- Crowe, A., Yue, W. Semi-quantitative Determination of Protein Expression Using Immunohistochemistry Staining and Analysis: An Integrated Protocol. BIO-PROTOCOL. 9 (24), (2019).
- Seligson, D. B., et al. Global histone modification patterns predict risk of prostate cancer recurrence. Nature. 435 (7046), 1262-1266 (2005).
- Liu, B., et al. Global Histone Modification Patterns as Prognostic Markers to Classify Glioma Patients. Cancer Epidemiology, Biomarkers & Prevention. 19 (11), 2888-2896 (2010).
- Pan, R. Y., et al. Positive feedback regulation of microglial glucose metabolism by histone H4 lysine 12 lactylation in Alzheimer's disease. Cell Metabolism. 34 (4), 634.e6-648.e6 (2022).
- Zhang, D., et al. Metabolic regulation of gene expression by histone lactylation. Nature. 574 (7779), 575-580 (2019).
- Pösel, C., Möller, K., Boltze, J., Wagner, D. C., Weise, G. Isolation and Flow Cytometric Analysis of Immune Cells from the Ischemic Mouse Brain. Journal of Visualized Experiments. (108), 53658 (2016).
- Oblak, A. L., et al. Comprehensive Evaluation of the 5XFAD Mouse Model for Preclinical Testing Applications: A MODEL-AD Study. Frontiers in Aging Neuroscience. 13, 713726 (2021).
- Bohlen, C. J., Bennett, F. C., Bennett, M. L. Isolation and Culture of Microglia. Current Protocols in Immunology. 125 (1), e70 (2019).
- McKinnon, K. M. Multiparameter Conventional Flow Cytometry. Flow Cytometry Protocols. 1678, 139-150 (2018).
- Marsh, S. E., et al. Dissection of artifactual and confounding glial signatures by single-cell sequencing of mouse and human brain. Nature Neuroscience. 25 (3), 306-316 (2022).
- Wang, L., Gaigalas, A. K., Marti, G., Abbasi, F., Hoffman, R. A. Toward quantitative fluorescence measurements with multicolor flow cytometry. Cytometry Part A. 73A (4), 279-288 (2008).
- Rumbaugh, G., Miller, C. A. Epigenetic changes in the brain: measuring global histone modifications. Methods in Molecular Biology (Clifton, N.J). 670, 263-274 (2011).
- Xavier, A. M., et al. Systematic delineation of signaling and epigenomic mechanisms underlying microglia inflammatory activity in acute and chronic brain pathologies. BioRvix. , (2022).
Перепечатки и разрешения
Запросить разрешение на использование текста или рисунков этого JoVE статьи
Запросить разрешениеСмотреть дополнительные статьи
This article has been published
Video Coming Soon
Авторские права © 2025 MyJoVE Corporation. Все права защищены