
Summary
Abstract
Introduction
Protocol
Representative Results
Discussion
Acknowledgements
Materials
References
Biology
针对性斯尔纳的噬菌体介导的交付构造击倒基因表达
We describe a method to knock down gene expression in a growing population of E. coli cells using sequence-targeted sRNA expression cassettes delivered by an M13 phagemid vector.
的RNA介导的击倒被广泛用于控制基因表达。该技术多功能系列采用可与任何序列合成和设计,以配合针对任何沉默基因的短RNA(斯尔纳)的。因为的sRNA构建可以引入到许多细胞类型直接或使用各种载体,基因表达可以在活细胞中,而不费力遗传修饰被抑制。最常见的RNA拦截技术,RNA干扰(RNAi),利用了靶mRNA的内源RNA诱导的沉默复合物(RISC)介导的序列识别和切割的。因此这种技术的应用只限于RISC的表达生物体,主要是真核生物。近日,新一代生物技术RNA已经开发的替代机制,通过RNA调控基因表达,在细菌,因此成为可能RNA介导的基因击倒。在这里,我们介绍了一种沉默基因EXPRES锡永在E.大肠杆菌的功能类似于RNA干扰。在这个系统中的合成噬菌粒被设计来表达的sRNA,其可设计为靶向任何序列。表达构建体递送到大肠杆菌的群体的大肠杆菌细胞与非裂解M13噬菌体,在这之后能够稳定地复制作为质粒。反义识别与靶mRNA的沉默由HFQ蛋白质,内源性大肠杆菌介大肠杆菌 。该协议包括用于设计反义的sRNA,构建噬菌粒载体,包装噬菌粒到M13噬菌体,感染制备活细胞群体,并执行感染本身的方法。荧光蛋白mKate2和抗生素抗性基因氯霉素乙酰转移酶(CAT)定位到产生代表数据和量化击倒效力。
的RNA介导的基因击倒进行两个阶段。首先,将RNA分子引入到细胞系或研究的有机体。第二,内源性RNA结合蛋白促进RNA的目标识别和产生的消声效果。所有的RNA拦截技术受益于合成sRNAs的定制性质,可以容易地制造,以匹配感兴趣的特定目标。然而,RNA摄取和沉默的分子细节横跨模型系统广泛地变化,限制在哪里以及如何的RNA击倒可以应用。
在线虫,双链RNA(dsRNA)的分子可直接在介质或通过用双链RNA表达大肠杆菌的群体喂食蠕虫引入大肠杆菌细胞1,2-,在果蝇中 ,RNA干扰,可以通过用双链RNA 3显微注射的胚胎通过简单地添加的dsRNA至培养基4来实现,或在细胞系中实施。在哺乳动物细胞系,合成小干扰RNA(siRNA)可以通过电穿孔1,2,5递送到活细胞,封装在脂质体3,6,或从DNA质粒载体4,7-表达。一旦RNA种类到达胞质溶胶,RNAi途径依赖于RISC复合物处理的ds RNA,促进靶的反义识别,并催化翻译抑制,mRNA降解,或异染色质形成,这取决于主机上。
由于这些要求的,古典的RNAi只能在有效地吸收外源RNA并表达RISC或类RISC活性有机体进行。值得注意的是,这不包括模型细菌E.大肠杆菌 ,它缺乏RNAi途径。然而,最近在合成生物学的发展提供必要的工具来解决这两个问题,交付和沉默问题。
在这个协议中,斯尔纳构建在大肠杆菌表达来自大肠杆菌的DNA载体交付给李咏使用M13噬菌体/辅助系统细胞。噬菌粒是复制的噬菌体衍生的F1起源的质粒。辅助质粒,在此情况下M13KO,进行所有以产生病毒颗粒所要求的机械,但本身不是胜任复制和包装。当噬菌粒和辅助质粒被共转化,单独噬菌粒是在F1原点复制,包装和分泌的。该矢量噬菌体是那么能干感染现场E.通过F菌毛的大肠杆菌 。
在这个系统中,沉默效应是由定制的sRNA盒结合的靶结合序列的支架序列产生。靶结合序列是反义的mRNA靶24个碱基对,通常是在核糖体结合位点(RBS)。该支架序列,由娜和他的同事开发出8,包含MICC,小调控RNA内源性大肠杆菌提取的HFQ结合基序大肠杆菌 。该HFQ蛋白刺激RNA的RNA结合蛋白g且mRNA降解,供应在该系统类似于RISC中的RNAi作用。 图1描绘了用于噬菌体介导的sRNA击倒,包括的sRNA盒结构,噬菌粒矢量,和消音机构的完整方案。
作为一种方法来调节在大肠杆菌中基因表达的大肠杆菌 ,斯尔纳沉默是简单,快速和灵活。有针对性的E.大肠杆菌是不是负担超出传播噬菌粒表达斯尔纳。这可能是在合成生物学或基础研究,其中较大的异源构建体的表达可以应变蜂窝资源9的上下文相关的。与新的目标噬菌粒可以用单个的PCR来制备和噬菌粒转化后收获一天。最后,几乎所有的表达可以有针对性。所述的sRNA调节盒(在标准的质粒)已经显示出对各种目标在代谢与典型的压制水平> 90%8工作。
10先前的工作扩展。首先,一个包装噬菌粒引入到大肠杆菌的分批培养大肠杆菌细胞并用于沉默荧光蛋白mKate2的表达。后续的荧光变化进行实时监测。第二,撞倒CAT基因中示出减少在琼脂平板上的表型氯霉素抗性。在这两种情况下,噬菌粒本身携带GFP标记,允许待测量独立的击倒效率感染率。
1.设计轴承斯尔纳沉默盒噬菌粒载体的构建
- 斯尔纳沉默盒8的从头设计
- 识别mRNA的完整序列使用DNA序列数据库被沉默。以产生靶序列,请注意编码序列的前24个碱基对,从位置+1至+24与起始密码子( 如,ATG)开始。
注意:沉默效率较低时的其他网站或mRNA的片段为目标8。 - 取靶序列的反向互补,以产生对的sRNA盒导向序列。请参阅表1为氯霉素乙酰转移酶(CAT)目标和指南序列的例子。
- 设计292 bp的完整的sRNA表达盒,安排PR启动子,指导序列,HFQ蛋白结合串联域和T1 / TE转录终止序列( 表2)。
- 添加所选择的附加的克隆位点,以方便的sRNA盒克隆到目标矢量。
- 通过商业基因合成或类似的方法来获得完整的斯尔纳磁带,它克隆与功能F1复制起点11的噬菌体载体。看到的最终噬菌粒载体的完整序列支持信息。
- 识别mRNA的完整序列使用DNA序列数据库被沉默。以产生靶序列,请注意编码序列的前24个碱基对,从位置+1至+24与起始密码子( 如,ATG)开始。
- 改变使用基于PCR的定点诱变12现有斯尔纳表达盒靶向序列
- 识别在现有的sRNA表达盒24 bp的指南序列。注:注释pAB.001质粒,在这项工作中使用,可作为补充的序列文件。
- 设计正向和反向与现有的sRNA盒侧翼新的24 bp的指南序列同源的短区域的引物。获得通过商业寡核苷酸合成引物。
注意:定点诱变引物设计是depic泰德于图2在表3中提供精确的引物模板的序列。 - E.准备的5毫升培养大肠杆菌携带模板斯尔纳表达噬菌粒。生长细胞过夜,在37℃振荡,在LB培养基中以适当的抗生素。
- 提取和使用DNA小量试剂盒或类似的方法12中的5毫升细菌培养净化模板斯尔纳表达噬菌粒。
- 准备使用的模板的sRNA表达噬菌粒和高保真聚合酶,一种具有前向和一个与反向引物( 表4)两个PCR反应。使用的PCR条件的建议通过聚合酶供应商( 表5)。增加模板浓度比标准反应10-50x更高占的事实,单引物反应不产生指数扩增。
- 结合上述两个PCR反应在一个离心管中。 Annea微升的产品通过加热到98℃的沸水浴中。放置微量离心管在水浴后,立即取出热源并允许浴超过1-2小时缓慢回到室温。
- 为了消除未突变模板斯尔纳表达噬粒,加入1微升。的DpnI限制性内切酶的混合物中,并孵育在37℃下1小时,或由制造商为完全消化所建议的时间。
注意:的DpnI消化只有甲基化靶位点,这是存在于主机的复制噬菌粒,但不PCR产物。 - 变换购买或准备13化学感受态大肠杆菌 1-5微升退火的PCR产物。通过在含有适当的抗生素的LB琼脂平板选择性电镀隔离转化菌株的单菌落。
- 以验证正确的指导序列的掺入,筛选通过菌落PCR所得菌落。用200μl的枪头,合作从单个转化的菌落llect细胞的少量。马克和保留原来的殖民地核实后下游使用。
- 收集的细胞添加至50μl的无核酸酶水在一个离心管中。吹打向上和向下混合。
- 使用台式热循环或沸水浴,通过加热溶解细胞到95℃2分钟。
- PCR的扩增使用1μl的热裂解的细胞作为DNA模板噬菌粒区。在表6和7中提供的PCR条件和热循环协议。请参阅验证引物序列补充pAB.001序列文件。
- 测序的PCR产物以验证正确的指导序列的掺入。
- 接种大肠杆菌的5毫升培养大肠杆菌克隆携带序列验证斯尔纳表达噬菌粒。生长细胞过夜,在37℃振荡,在选择性LB培养基。
- 准备甘油股票的序列验证克隆。添加750微升过夜培养物,以250μl的60%的甘油在一螺丝帽冷冻管中。
- 在-80℃保存甘油下去。在过夜培养的剩余部分可被用作在步骤2中的sRNA表达噬菌粒的来源。
M13包装库存噬菌粒2.生产和收获
- E.准备的5毫升培养大肠杆菌携带斯尔纳表达噬菌粒。生长细胞过夜,在37℃振荡,在LB培养基中以适当的抗生素。注意:的sRNA表达噬菌粒可通过从头克隆如步骤1.1.5描述从现有噬菌粒来获得,或修改,并在步骤1.2.16收获。
- 同样准备E的5毫升培养大肠杆菌携带M13KO7辅助质粒。生长细胞过夜,在37℃振荡,在选择性LB培养基。
- 提取和纯化的斯尔纳expressioÑ 噬菌粒和用DNA提取试剂盒或类似的方法12辅助质粒。
- Cotransform购买或制备13化学感受态大肠杆菌与每个的sRNA表达噬菌粒和辅助质粒的1微升。通过这两个结构选择性抗生素的LB琼脂电镀选择cotransformants。
- 从在LB的共转化株与选择性抗生素的单菌落制备10ml的培养。在37℃孵育摇8-12小时或过夜。
- 离心培养3300 XG 10分钟。收集通过0.2微米的过滤器将上清液过滤。注意:在媒体泄漏的情况下,请用稀释的漂白剂(0.5%),该地区摧毁感染性噬菌体颗粒。
- 储存在4℃下将包装噬菌粒滤液。注:样品可以维持数天至数周而不丧失活性。
3. F +靶细胞的制备沉默
- 确定是否要针对沉默细胞表达歼14菌毛。如果F菌毛已经存在,继续步骤4。
注:E.常见的实验室菌株大肠杆菌被注解为F +或F'以指示在其基因组上或质粒在F菌毛的存在。 - E.获得的F +菌株大肠杆菌如TOP10F'。
注意:确保目标菌株携带一个独特的抗性标记,以从结合后的F-质粒供体分离。 - 要通过与F +菌株结合引进˚F菌毛,准备5毫升培养目标应变和F-质粒捐助14的都有。生长细胞过夜,在37℃振荡,在LB培养基中以适当的抗生素。
- 第二天,稀释两种菌株1:100在5ml选择性的LB,并继续培养,在37℃振荡。
- 通过测量选择确定细胞的生长相在使用台式分光光度计600纳米(OD 600)文化的iCal密度。培养得到的细胞约2小时,直到0.3的OD 600,指示数期生长15。
- 在微量离心管准备3偶联反应:0.5毫升F-质粒供体+0.5毫升靶菌株加入0.5ml的F-质粒供体+ 0.5毫升LB培养基培养基(阴性对照)和0.5ml靶应变+ 0.5毫升LB培养基培养基(阴性对照)。允许缀合,在37℃振荡进行2小时。
- 板100微升在选择性LB琼脂每个缀合反应与具体到F质粒的抗生素(通常四环素)和目标菌株。板阴性对照反应,以确认没有捐赠者或受体菌表达两种抗生素的抗性。
4.感染了沉默噬菌粒包装
- 接种F +靶细胞的单个菌落到LB中介的5毫升培养一个适当的抗生素。在37℃振荡孵育过夜。
- 第二天,稀释F +靶细胞1:100在5ml选择性LB 培养基,并继续培养,在37℃振荡。
- 通过使用台式分光光度计测定培养物的光密度在600nm(OD 600)测定细胞的生长阶段。培养得到的细胞约2小时,直到0.3的OD 600,指示数期生长15。注:在F菌毛和感染效率表达最高的是日志阶段。
- 添加M13-包装噬菌粒(来自步骤2.6)与靶细胞以1:1的体积比:100达到目标人口的近99%的感染。允许感染在37℃振荡30-60分钟进行。
- 根据所选择的方法测定了的sRNA沉默表型。
注意:对于荧光蛋白靶标,消音效果可以定量直接荧光10。交替,表型分析可用于观察基因敲除8的表型的影响。 - 准备甘油股票以下步骤1.2.14-1.2.16的斯尔纳表达噬菌粒主机。注意:噬菌粒将在宿主菌株无限期传播,并且可以与抗生素类似于传统的质粒被维持。
在液体介质mKate2荧光沉默
图1描绘了用于在这项工作中所描述的sRNA介导的击倒,包括的sRNA盒设计,噬菌粒矢量,和沉默机制的方案。以下协议1.2,质粒pAB.001的斯尔纳沉默盒改变目标mKate。该斯尔纳盒原料合成并克隆到噬菌粒Litmus28i_J23115-B0032-GFP,莫妮卡奥尔蒂斯和德鲁恩迪11的礼物。此噬菌粒携带GFP表达和卡那霉素抗性标记,允许被跟踪成功感染。制备包装噬菌股以下协议2。
E.的衍生物大肠杆菌 K12 MG1655背着一个组成型表达,染色体整合mKate2标记是由共轭的噬菌体感染准备用以下方案3细胞生长至对数中期,介绍以下协议4.噬菌粒感染后噬菌粒一个F质粒供体菌株,200微升的培养物转移到荧光板读数器和荧光24小时连续监测。
图3示出的sRNA介导的沉默对mKate2表达的影响。感染与抗mKate2噬菌粒的菌株没有表现出可检测的mKate2荧光超过本底。相比之下,这株没有表达GFP标记,表明噬菌体的成功吸收。未感染的对照细胞产生mKate2荧光但不GFP。附加控制,其中,反mKate2靶向域与一个序列靶向的CAT取代,对mKate2荧光没有影响。
氯霉素抗性的琼脂平板上沉默
内容“FO:保together.within页=”1“>以下协议1.1,制作一个斯尔纳沉默卡带目标CAT 大肠杆菌 K12 MG1655背着一个组成型表达,染色体整合CAT标志是为准备的衍生物。噬菌体感染通过与以下方案3.将细胞在F-质粒供体菌株偶联生长到对数中期和噬菌粒在37℃下引入以下协议4.温育1小时后,感染的细胞进行系列稀释,并在镀的氯霉素浓度。将板范围温育过夜,并抗性细胞中的每一个氯霉素浓度的比例通过计数菌落形成单位(CFU的)次日确定。图4示出的sRNA介导的沉默对氯霉素抗性的表型的影响。未感染的细胞,或感染噬菌粒靶向mKate2细胞,分别以抗氯霉素在所有测试浓度。相比之下,感染了针对噬菌粒细胞CAT在低浓度氯霉素减少显示出生存,近99%的较高浓度杀。添加氨苄青霉素,要选择只携带细菌噬菌体,减少氯霉素生存检测不到的水平。这与早期的工作一致表示从噬菌体感染的逃避逃跑沉默10共同路线。
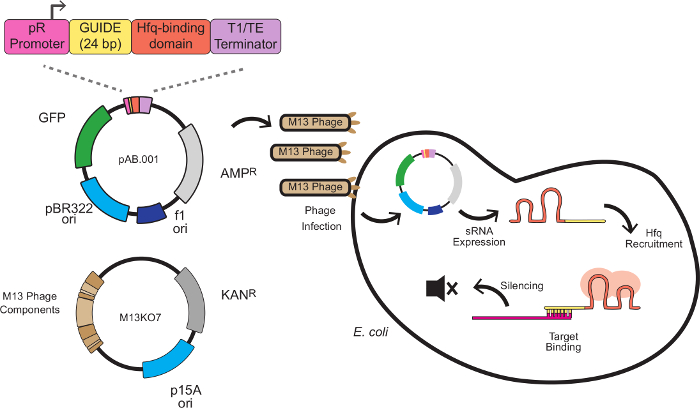
图1:在 大肠杆菌 的基因沉默 大肠杆菌 用的sRNA表达盒由M13噬菌体递送的的sRNA盒是由4个模块:所述PR启动子(来自λ噬菌体来源的组成型启动子),一个24bp的靶向结构域,HFQ结合域从MICC提取和转录终止子8 大肠杆菌大肠杆菌表达在F菌毛,其中斯尔纳表达开始。该斯尔纳然后招募HFQ蛋白(红色表示),并结合反义靶mRNA核糖体结合位点附近,导致翻译抑制和mRNA降解。 请点击此处查看该图的放大版本。
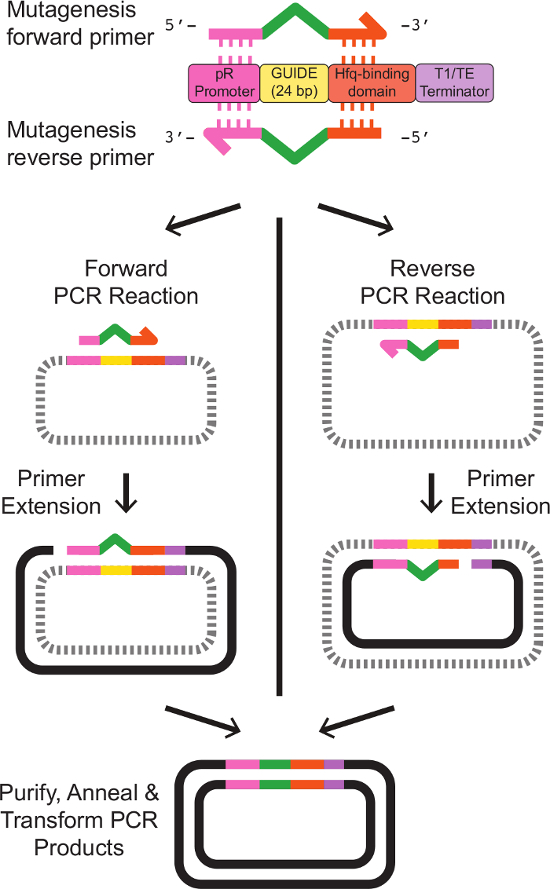
图2:引物的设计和的sRNA目标站点的定点诱变的引物被设计具有部分同源性的现有的sRNA盒。正向引物含有20bp的同源的5'端的PR启动子,接着表示新的指南序列24碱基,然后18碱基对同源的3'末端的HFQ结合结构域。反向引物是正向引物的确切反向互补,与现有的sRNA盒侧翼的新指南序列的反向互补同源性的区域。确切的引物序列在表3中给出。与正向和反向引物的独立的单引物的PCR反应产生具有所需修饰的序列的直链,单链DNA。退火正向和反向的反应产物,按照如在协议中所述的清理,结果在双链DNA质粒带有所需修改斯尔纳磁带。 请点击此处查看该图的放大版本。
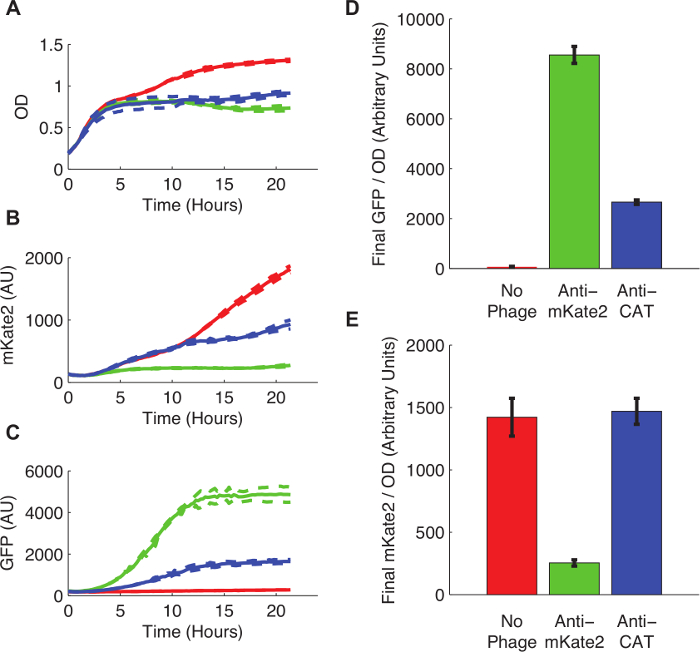
图3:K一个染色体整合mKate2荧光记者nockdown。E.大肠杆菌 MG1655 K12表达mKate2要么感染与抗mKate2噬菌粒(绿线和棒)左未处理(红色线和条),或感染了控制噬菌粒靶向CAT(蓝线和条)。 ( 一 )未经处理的E.大肠杆菌上升到更高的饱和密度,说明代谢成本噬菌体感染。虚线表示的3次重复的标准偏差。 (B)中的mKate2信号降低到接近背景水平在抗mKate2处理应变,而不是在对照菌株。 (C)GFP荧光,也由噬菌粒携带,只在噬菌粒处理的对照是可检测的。 (D,E)最终荧光读数生长24小时后标准化为外径。 GFP信号,指示噬菌体感染时,在未经处理的控制缺席,而且还大幅减少以下抗猫phagemid治疗。这可以指示噬菌粒的脱靶效应。所述mKate2信号减少了抗mKate2噬菌粒治疗相比未处理的对照。在CAT-针对性的控制噬菌显示对mKate2荧光没有影响。误差棒代表3次重复的标准差。 请点击此处查看该图的放大版本。
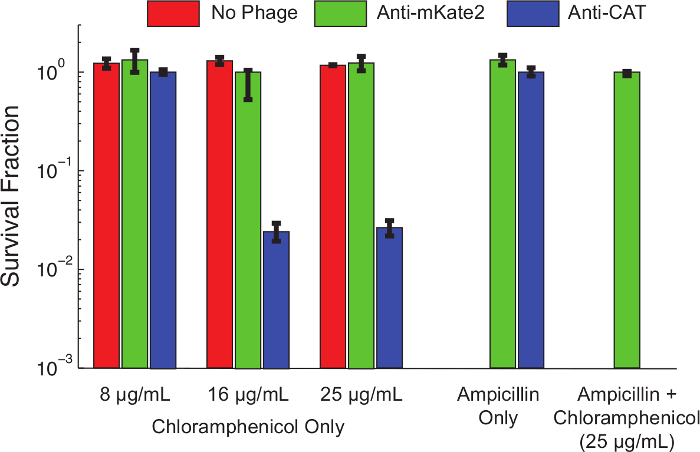
图4:CAT的击倒恢复氯霉素敏感性遗传抗性种群E.。 大肠杆菌 MG1655 K12表达染色体整合CAT基因不进行处理(红色条),用控制噬菌粒靶向mKate2(绿色柱)处理,或用表达抗CAT的sRNA(蓝色条)噬菌粒处理。 1小时后感染,生存能力上表示蚂蚁ibiotics被连续稀释和电镀评估。用抗CAT噬菌粒处理过的菌株通过在较高浓度氯霉素显著杀死(> 90%),而对照处理不受影响。加入氨苄青霉素到培养板正选择用于噬菌粒感染和消除未感染的细胞。在这些条件下,抗CAT处理后没有观察到氯霉素抗性菌落。这表明大多数幸存者代表感染的故障,而不是沉默的故障。误差棒代表3次重复的标准差。 请点击此处查看该图的放大版本。
| CAT靶序列 | 5' - ATGGAGAAAAAAATCACTGGATAT - 3' |
| CAT指导序列 | 5' - ATATCCAGTGATTTTTTTCTCCAT - 3' |
表1:示例TARGET和指导序列的CAT基因注意的反向互补关系。
| PR启动 | TAACACCGTGCGTGTTGACTATTTTACCTCTGGCGGTGATAATGGTTGC | ||||
| 指导序列 | ATATCCAGTGATTTTTTTCTCCAT | ||||
| HFQ结合域 | TTTCTGTTGGGCCATTGCATTGCCACTGATTTTCCAACATATAAAAAGACAAGCCCGAACAGTCGTCCGGGCTTTTTT TCTCGAG | ||||
| T1 / TE终结者 | CTCGAGCCAGGCATCAAATAAAACGAAAGGCTCAGTCGAAAGACTGGGCCTTTCGTTTTATCTGTTTTTGTCGGTGAA CGCTCTCTACTAGAGTCACACTGGCTCACCTTCGGGTGGGCCTTTCTGCGTTTATA | ||||
表2:所述的sRNA盒式序列组件每个序列写入5'-3'。完整的磁带是这4个元素的串联,以便与包括BP 292。
| 正向引物 | 5' - CTGGCGGTGATAATGGTTGC [GUIDE] TTTCTGTTGGGCCATTGC - 3' |
| 反向引物 | 5' - GCAATGGCCCAACAGAAA [TARGET] GCAACCATTATCACCGCCAG - 3' |
表3:引物的设计改变现有引导元件的正向引物包括在过去的20碱基对的PR启动子,新的指导序列,和HFQ结合结构域的前18个碱基对。靶序列是指导序列的确切反向互补。反向引物是正向引物的确切反向互补。
表4:推荐条件的单引物诱变的PCR。
| 步 | 温度 | 时间 |
| 初始变性 | 98℃ | 30秒 |
| 30个循环 | 98℃ | 10秒 |
| 55°C | 30秒 | |
| 72℃ | 120秒 | |
| 最终延伸 | 72℃ | 300秒 |
| 存储 | 10℃ |
表5:推荐热循环协议的单引物诱变的PCR。
| 零件 | 卷 |
| 模板DNA | 1微升 |
| 10μM正向引物 | 0.5微升 |
| 10μM反向引物 | 0.5微升 |
| Taq酶2X预混 | 25微升 |
| 无核酸酶的水:- [R | 23微升 |
| 总成交量 | 50微升 |
表6:建议的条件为序列验证PCR。
| 步 | 温度 | 时间 |
| 初始变性 | 95℃ | 30秒 |
| 30个循环 | 95℃ | 30秒 |
| 55°C | 30秒 | |
| 68℃ | 30秒 | |
| 最终延伸 | 68℃ | 300秒 |
| 存储 | 10℃ |
表7:建议热循环仪专业版母育的序列验证PCR。
相比于不相关的对照本方法实现了mKate荧光水平减少80%。这是符合其他RNA敲除的方法,其中完整的沉默不遵守和50-90%的效率是典型的16,17。在表型水平,CAT定位击倒能够显著衰减氯霉素抗性,且消除它在某些条件下。
只有几个小时后感染( 图3B)后,击倒表型是在群体水平检测。这说明了基于噬菌体的输送的一个重要特征:高的击倒频率可以直接在分批培养无需事先遗传修饰而获得。不像使用质粒转化或基因组整合常规遗传修饰,噬菌体感染不要求一个人口从单个孤立菌落再生长。这使得噬菌体感染的效果是EXPLORED与复杂的空间动态11的人群,像生物膜18,或者在基因混合自然种群19预先存在的空间结构。
该方法的关键步骤是在高滴度的生产包装噬菌粒。与噬菌体颗粒的生产相关的代谢负担可能会导致在噬菌粒生产菌株的突变或质粒损失率高。建议在噬菌粒生产菌株从单个共转化菌落直接培养,不冷藏之前,噬菌体收获冷冻或子培养。引入噬菌粒和辅助质粒到大肠杆菌时,也可以观察到共转化的效率低同时大肠杆菌 。在这种情况下,更高的效率可以通过首先将所述辅助质粒,然后制备感受态细胞的轴承用于与噬菌粒随后转化辅助质粒而获得。
帕gemid感染或的sRNA表达还对靶细胞的可检测的代谢负担,并且可能导致一些表型扰动。例如,在mKate2荧光的减少观察到即使当细胞感染噬菌粒靶向CAT( 图3)。与M13感染是没有想到来触发E.全身应激反应大肠杆菌 20,但也可以间接地改变转录模式。另外,包括在噬菌粒的GFP或氨苄青霉素抗性标记可能会争夺资源的细胞,减少mKate2表达和9。最后,的sRNA盒本身可以全局通过滴定HFQ蛋白改变基因表达谱,或通过脱靶mRNA的沉默。脱靶效应在体内 RNA干扰21-23常见,但他们还没有被系统地研究了这个系统。
这种方法的一个限制是该感染艾菲效率低于100%,允许一些未感染的细菌中的人口持续。这项工作,并早期工作10的结果表明,未感染细胞代表最终人口的1-10%,并且负责大多数观察到的nonsilenced表型。各种路由M13-电阻是已知的,与菌毛表达24的最常见的突变损失。在这些限制的光,控制应该被用来确认高感染率和敲低效率。
在某些应用中另一个潜在的限制是污染辅助噬菌体偶尔传输。虽然M13K07包含突变的包装信号,它可以被包装成在低频噬菌体衣壳,并转移到受感染的人群中,导致胜任噬菌体生产和噬菌体超出了最初的感染事件25继续扩散的细胞。修改辅助噬菌体已被证明有效在以降低生产噬菌体26的成本降低了非特异性的包装,虽然有时。
改造噬菌体已成为大肠杆菌的必备工具大肠杆菌合成生物学,允许快速交货的新基因,以不断增长的人口。最近的工作已经产生细胞间通信电路11或表达的转录因子来抑制抗生素耐药性的途径27。这里介绍的协议增加了工具,允许通过编程的RNA细菌生理控制不断增长的集合。 CRISPR-CAS核酸酶,突变以消除核酸酶活性时,已经示出在RNA的引导基因靶17,28到抑制转录。与此相反,的sRNA沉默工作在翻译水平,并且不需要外源蛋白的表达。下一代生物技术可联合转录和翻译控制与噬菌体介导的递送到复杂苯氧编程类型的实时性。
The authors have nothing to disclose.
这项工作的经费由基金会贝当古舒莱尔支持贝当古巴黎队iGEM大赛提供。我们感谢INSERM U1001科研单位,尚塔尔对于LOTTON技术援助和咨询。噬菌体Litmus28i_J23115-B0032-GFP是由莫妮卡·奥尔蒂斯和斯坦福大学的德鲁恩迪提供。
| Name | Company | Catalog Number | Comments |
| Plasmid Miniprep Kit | Qiagen | 27104 | |
| DpnI Enzyme | NEB | R0176S | |
| Phusion High Fidelity Polymerase | NEB | M0530S | |
| Taq 2x Master Mix | NEB | M0270L | |
| M13KO7 Helper Phage | NEB | N0315S | |
| DH5α Competent Cells | Life Technologies | 18265-017 | |
| TOP10F' Cells | Life Technologies | C3030-03 | |
| LB Broth | Sigma | L3022-250G | |
| Ampicillin | Sigma | A9393-5G | |
| Kanamycin | Sigma | 60615-5G | |
| Chloramphenicol | Sigma | C0378-5G | |
| Tetracycline | Sigma | 87128-25G |
- Ohkumo, T., Masutani, C., Eki, T., Hanaoka, F. Use of RNAi in C. elegans. RNAi. , 129-137 (2008).
- Fire, A., Xu, S., Montgomery, M. K., Kostas, S. A., Driver, S. E., Mello, C. C. Potent and specific genetic interference by double-stranded RNA in Caenorhabditis elegans. Nature. 391 (6669), 806-811 (1998).
- Iordanou, E., Chandran, R. R., Blackstone, N., Jiang, L. RNAi interference by dsRNA injection into Drosophila embryos. J Vis Exp. (50), e2477 (2011).
- Ramadan, N., Flockhart, I., Booker, M., Perrimon, N., Mathey-Prevot, B. Design and implementation of high-throughput RNAi screens in cultured Drosophila cells. Nat Protoc. 2 (9), 2245-2264 (2007).
- Tsong, T. Y. Electroporation of cell membranes. Biophys J. 60 (2), 297-306 (1991).
- Kim, W. J., Chang, C. -. W., Lee, M., Kim, S. W. Efficient siRNA delivery using water soluble lipopolymer for anti-angiogenic gene therapy. J Control Release. 118 (3), 357-363 (2007).
- Shi, Y. Mammalian RNAi for the masses. Trends Genet. 19 (1), 9-12 (2003).
- Na, D., Yoo, S. M., Chung, H., Park, H., Park, J. H., Lee, S. Y. Metabolic engineering of Escherichia coli using synthetic small regulatory RNAs. Nat Biotechnol. 31 (2), 170-174 (2013).
- Ceroni, F., Algar, R., Stan, G. -. B., Ellis, T. Quantifying cellular capacity identifies gene expression designs with reduced burden. Nat Methods. 12 (5), 415-418 (2015).
- Libis, V. K., Bernheim, A. G., et al. Silencing of Antibiotic Resistance in E. coli with Engineered Phage Bearing Small Regulatory RNAs. ACS Synth Biol. 3 (12), 1003-1006 (2014).
- Ortiz, M. E., Endy, D. Engineered cell-cell communication via DNA messaging. J Biol Eng. 6 (1), 16 (2012).
- Edelheit, O., Hanukoglu, A., Hanukoglu, I. Simple and efficient site-directed mutagenesis using two single-primer reactions in parallel to generate mutants for protein structure-function studies. BMC Biotechnol. 9 (1), 61 (2009).
- Chung, C. T., Miller, R. H. Preparation and storage of competent Escherichia coli cells. Recombinant DNA Part I. , 621-627 (1993).
- Phornphisutthimas, S., Thamchaipenet, A., Panijpan, B. Conjugation in Escherichia coli. Biochem Mol Biol Educ. 35 (6), 440-445 (2007).
- Sezonov, G., Joseleau-Petit, D., D'Ari, R. Escherichia coli Physiology in Luria-Bertani Broth. J Bacteriol. 189 (23), 8746-8749 (2007).
- Mittal, V. Improving the efficiency of RNA interference in mammals. Nat Rev Genet. 5 (5), 355-365 (2004).
- Qi, L. S., Larson, M. H., et al. Repurposing CRISPR as an RNA-guided platform for sequence-specific control of gene expression. Cell. 152 (5), 1173-1183 (2013).
- Lu, T. K., Collins, J. J. Dispersing biofilms with engineered enzymatic bacteriophage. Proc Natl Acad Sci USA. 104 (27), 11197-11202 (2007).
- Yosef, I., Manor, M., Kiro, R., Qimron, U. Temperate and lytic bacteriophages programmed to sensitize and kill antibiotic-resistant bacteria. Proc Natl Acad Sci USA. 112 (23), 7267-7272 (2015).
- Karlsson, F., Malmborg-Hager, A. -. C., Albrekt, A. -. S., Borrebaeck, C. A. K. Genome-wide comparison of phage M13-infected vs. uninfected Escherichia coli. Can J Microbiol. 51 (1), 29-35 (2005).
- Senthil-Kumar, M., Mysore, K. S. Caveat of RNAi in plants: the off-target effect. Methods in molecular biology. 744, 13-25 (2011).
- Jackson, A. L., Linsley, P. S. Recognizing and avoiding siRNA off-target effects for target identification and therapeutic application. Nat Rev Drug Discov. 9 (1), 57-67 (2010).
- Cho, S. W., Kim, S., et al. Analysis of off-target effects of CRISPR/Cas-derived RNA-guided endonucleases and nickases. Genome Res. 24 (1), 132-141 (2014).
- Hagens, S., Blasi, U. Genetically modified filamentous phage as bactericidal agents: a pilot study. Lett Appl Microbiol. 37 (4), 318-323 (2003).
- Kasman, L. M., Kasman, A., Westwater, C., Dolan, J., Schmidt, M. G., Norris, J. S. Overcoming the phage replication threshold: a mathematical model with implications for phage therapy. J Virol. 76 (11), 5557-5564 (2002).
- Chasteen, L., Ayriss, J., Pavlik, P., Bradbury, A. R. M. Eliminating helper phage from phage display. Nucleic Acids Res. 34 (21), e145 (2006).
- Lu, T. K., Collins, J. J. Engineered bacteriophage targeting gene networks as adjuvants for antibiotic therapy. Proc Natl Acad Sci USA. 106 (12), 4629-4634 (2009).
- Bikard, D., Jiang, W., Samai, P., Hochschild, A., Zhang, F., Marraffini, L. A. Programmable repression and activation of bacterial gene expression using an engineered CRISPR-Cas system. Nucleic Acids Res. 41 (15), 7429-7437 (2013).
Tags
ABOUT JoVE
Copyright © 2024 MyJoVE Corporation. All rights reserved