
Summary
Abstract
Introduction
Protocol
Representative Results
Discussion
Acknowledgements
Materials
References
Biology
표적 스르나의 파지 매개 배달에 유전자 발현을 허물고 구축
We describe a method to knock down gene expression in a growing population of E. coli cells using sequence-targeted sRNA expression cassettes delivered by an M13 phagemid vector.
RNA 매개 knockdowns 널리 유전자 발현을 제어하는 데 사용됩니다. 기술이 다용도 가족 어떤 순서로 합성 사일런 대상 모든 유전자를 보완하도록 설계 될 수 짧은 RNA (스르나)를 사용한다. 스르나 직접 여러 종류의 세포에 도입 또는 벡터의 다양한 사용하여 구축 될 수 있기 때문에, 유전자 발현은 유전자 조작 수고없이 살아있는 세포에서 억제 될 수있다. 가장 일반적인 RNA 녹다운 기술은 RNA 간섭 (RNAi), 표적 mRNA는 내인성 RNA 유도 복합체 (RISC)를 침묵 중재하는 시퀀스 인식 및 절단을 이용한다. 이 기술의 응용 프로그램은 따라서 RISC 발현 생물, 주로 진핵 생물로 제한됩니다. 최근 RNA의 생물 공학자의 새로운 세대는 RNA를 통해 유전자 발현을 제어하기위한 다른 메커니즘을 개발 등 세균 가능한 RNA 매개 유전자 knockdowns했다. 여기에서 우리는 유전자 EXPRES을 침묵하는 방법을 설명합니다E.에서 시온 대장균 기능의 RNAi는 비슷하게. 이 시스템에서 합성 파지 미드는 어떤 순서를 대상으로 설계 할 수있다 스르나를 표현하기 위해 설계되었습니다. 식 구조는 E.의 인구에 전달된다 이 플라스미드로 안정적으로 복제 할 수있는 후 비 용균 M13 파지와 대장균 세포. 안티센스 인식 대상의 mRNA의 사일런 싱은 E. 내인성 Hfq 단백질에 의해 매개되고 대장균. 이 프로토콜은, 안티센스 스르나를 설계 파지 미드 벡터를 구성, M13 박테리오파지에 파지 미드 포장, 감염 살아있는 세포 집단을 준비하고, 감염 자체를 수행하기위한 방법을 포함한다. 형광 단백질 mKate2 및 항생제 내성 유전자, 클로람페니콜 아세틸 트랜스퍼 라제 (CAT)의 대표 데이터를 생성하고, 최저 효과를 정량화하는 타겟팅.
RNA 매개 유전자 knockdowns은 두 단계로 진행합니다. 우선, RNA 분자는 세포주 또는 연구 유기체에 도입된다. 둘째, 내인성 RNA 결합 단백질이 RNA 표적 인식을 촉진하고 침묵 효과를 생산하고 있습니다. 모든 RNA 녹다운 기술은 쉽게 관심의 특정 목표에 맞게 제조 할 수 합성 sRNAs의 사용자 정의 특성, 혜택을 누릴 수 있습니다. 그러나 RNA 흡수와 침묵의 분자 세부 사항은 어디에 어떻게 RNA의 knockdowns을 적용 할 수 구속, 모델 시스템에 걸쳐 매우 다양합니다.
선충에서 이중 가닥 RNA는 (dsRNA를) 분자는 미디어 또는 dsRNA를 발현 E.의 인구 벌레에게 먹이를 직접 도입 할 수있다 콜라이 세포 1,2-. 초파리는 RNAi를 단순히 배지 4의 dsRNA의 dsRNA를 첨가하여 3 배아 마이크로 인젝션에 의해 달성 또는 세포주에서 구현 될 수있다. 포유 동물 세포 라인에서,합성 작은 간섭 RNA는 (siRNA의)는 리포좀 -3,6- 패키지로 전기 1,2,5-하여 살아있는 세포로 전달 또는 DNA 플라스미드 벡터 -4,7-로부터 발현 될 수있다. RNA의 종 세포질에 도달하면, RNAi의 경로는 dsRNA를 처리 대상의 안티센스 인식을 용이하게하고, 호스트에 따라 번역 억제, mRNA의 분해 또는 이질 염색질 형성을 촉진하기 위해 RISC 복합체에 의존한다.
때문에 이러한 요구 사항, 고전의 RNAi는 효율적으로 외래 RNA를지고 RISC 또는 RISC와 같은 활동을 표현하는 유기체에서 수행 할 수 있습니다. 특히,이 모델 박테리아 E. 제외 RNAi의 경로 부족 대장균. 그러나, 합성 생물학의 최근 발전은 배달 문제와 침묵의 문제를 모두 해결하기 위해 도구를 제공합니다.
이 프로토콜에서, 스르나 구조는 E.로 표현된다 하는 DNA 벡터에서 대장균은 리튬에 전달M13 파지 미드 / 헬퍼 시스템을 이용하여 세포를 ving. 파지 미드 복제의 파지 유래 F1 원점 어떤 플라스미드이다. 헬퍼 플라스미드는이 경우 M13KO에서 바이러스 입자를 생산하는데 필요한 모든 장치를 운반하지만, 복제 및 패키징 능력 그 자체이다. 파지 미드 및 헬퍼 플라스미드를 공동 - 형질 전환되는 경우, 단독 파지 미드는 F1 복제 원점 포장 분비된다. 벡터화 파지 미드 라이브 E.을 감염 후 능력이 은 F pilus를 통해 대장균.
이 시스템에서, 침묵 효과는 표적 - 결합 서열과 골격 서열과 함께 정의 스르나 카세트에 의해 제조된다. 타깃 결합 서열은 일반적으로 리보좀 결합 부위 (RBS)에서 mRNA를 표적으로 안티센스 24 염기쌍이다. 나 동료 (8)에 의해 개발 된 발판 순서는, MICC, E.에 내생 작은 규제 RNA로부터 추출 된 Hfq 바인딩 모티브를 포함 대장균. Hfq 단백질은 RNA-RNA의 bindin 자극g 및 mRNA의 분해, RNAi의에서 RISC와 비슷한이 시스템의 역할을 제공. 그림 1은 스르나 카세트 구조, 파지 미드 벡터화, 그리고 침묵기구와 파지 매개 스르나의 knockdowns을위한 완전한 방식을 보여줍니다.
방법 으로서는 E.에서 유전자 발현을 조절하는 대장균, 스르나의 침묵은 간단하고 신속하고 다양합니다. 대상 E. 대장균은 파지 미드를 전파하고 스르나을 표현 이상 부담하지 않습니다. 이 큰 이종 구조의 발현이 세포 자원 9 변형 할 수 합성 생물학 또는 기초 연구의 맥락에서 관련 될 수있다. 새로운 목표와 파지 미드는 단일 PCR로 제조하고 파지 미드 변환 후 하루를 수확 할 수있다. 마지막으로, 거의 모든 mRNA를 타겟팅 할 수 있습니다. 스르나 조절 카세트 (표준 플라스미드)> 90 % 8 전형적인 억제 수준 대사 대상의 다양한 작업을하는 것으로 나타났다.
10 카세트 사용하여 이전 작업에 확장합니다. 우선, 포장 파지 미드는 E.의 배치 배양에 도입 대장균 세포와 형광 단백질 mKate2의 발현을 침묵하는 데 사용됩니다. 이후 형광 변경 사항이 실시간으로 모니터링된다. 둘째, CAT 유전자를 쓰러 뜨린 것은 한천 플레이트에 표현형 클로람페니콜 저항을 줄이기 위해 표시됩니다. 두 경우 모두, 파지 미드 자체 감염률 독립적 최저 효율을 측정 할 수 있도록하는 GFP 마커를 운반한다.
스르나 음소거 카세트 베어링 1. 설계 및 건설 파지 미드 벡터의
- 스르나 음소거 카세트 (8)의 드 노보 디자인
- mRNA에의 완전한 시퀀스는 DNA 서열 데이터베이스를 사용하도록 침묵 식별한다. 표적 서열을 생성하기 위해, 개시 코돈 (즉, ATG)로 시작 위치로부터 +1 +24에, 코딩 서열의 처음 24 BP 참고.
참고 : 다른 사이트 또는 mRNA의 세그먼트가 8을 대상으로하는 경우 음소거 덜 효율적이다. - 스르나 카세트 용 GUIDE 시퀀스를 생성하기 위해 상기 표적 서열의 역 상보보십시오. 클로람페니콜 아세틸 트랜스퍼 라제 (CAT)에 대한 목표와 GUIDE 시퀀스의 예는 표 1을 참조하십시오.
- 292 bp의 전체 스르나 발현 카세트를 설계하기 위해 일련의 PR 프로모터, GUIDE 순서, Hfq 단백질 결합 도메인과 T1 / TE 전사 종결 서열 (표 2)를 배열합니다.
- 대상 벡터에 스르나 카세트의 복제를 용이하게하기 위해 선택의 추가 복제 사이트를 추가합니다.
- 상업 유전자 합성 또는 유사한 방법으로 전체 스르나 카세트를 구하여 기능 F1 복제 기원 (11)와 어떤 파지 미드 벡터로 복제. 최종 파지 미드 벡터의 전체 시퀀스 정보 지원을 참조하십시오.
- mRNA에의 완전한 시퀀스는 DNA 서열 데이터베이스를 사용하도록 침묵 식별한다. 표적 서열을 생성하기 위해, 개시 코돈 (즉, ATG)로 시작 위치로부터 +1 +24에, 코딩 서열의 처음 24 BP 참고.
- PCR 기반의 사이트 감독 돌연변이 유발 (12)를 사용하여 기존 스르나 식 카세트의 대상 시퀀스를 변경
- 기존 스르나 발현 카세트에서 24 bp의 가이드 순서를 확인합니다. 참고 :이 작업에 사용되는 주석 pAB.001 플라스미드는, 보충 시퀀스 파일로 사용할 수 있습니다.
- 디자인은 앞으로 새로운 24 bp의 가이드 시퀀스를 측면 기존 스르나 카세트에 동성 짧은 지역과 역방향 프라이머. 상업 올리고 뉴클레오티드 합성을 통해 프라이머를 얻습니다.
참고 : 부위 특이 적 변이에 대한 프라이머 디자인 depic입니다테드는 그림 2에 정확한 프라이머 템플릿 시퀀스는 표 3에 제공된다. - E.의 5 ml의 문화를 준비 대장균은 템플릿 스르나 발현 파지 미드를 들고. 적절한 항생제로 LB 배지에서 진탕 37 ° C에서 하룻밤 세포를 성장.
- 추출하여 DNA 미니 프렙 키트 또는 유사한 방법 (12)을 사용하여 5 ㎖ 세균 배양 물로부터 템플릿 스르나 발현 파지 미드를 정제.
- 템플릿 스르나 발현 파지 미드 높은 충실도 중합 효소, 전방 하나와 역방향 프라이머 (표 4) 하나를 사용하여 두 개의 PCR 반응을 준비합니다. 중합 효소 공급 업체 (표 5)에서 권장하는 PCR 조건을 사용합니다. 하나의 프라이머 반응이 기하 급수적으로 증폭을 생성하지 않는다는 사실을 고려하여 표준 반응보다 높은 10-50x하는 템플릿 농도를 증가시킨다.
- 미세 원심 관에 상기 두 PCR 반응을 결합한다. Annea리터 끓는 물을 욕조에 98 ° C까지 가열에 의한 제품. 바로 수조에 microcentrifuge 관을 배치 한 후, 열원을 제거하고, 욕을 서서히 1-2 시간에 걸쳐 실온으로 복귀 할 수있다.
- 1 μl를 추가합니다 unmutated 템플릿 스르나 발현 파지 미드를 제거합니다. 혼합물을 DpnI 제한 효소 1 시간 또는 완전 소화 제조자가 권장하는 시간 동안 37 ℃에서 배양한다.
참고 : DpnI는 PCR 제품 호스트 복제 파지 미드에 존재하지만 만 메틸화 대상 사이트를 소화. - 구입 또는 13 화학적으로 유능한 E. 준비 변환 어닐링 된 PCR 산물의 5 μL와 대장균. 적절한 항생제를 함유하는 LB 아가 플레이트에 선택적 도금에 의해 형질 전환 된 균주의 단일 콜로니를 분리.
- 올바른 GUIDE 시퀀스의 통합을 확인하려면, 식민지 PCR에 의한 결과 식민지를 화면. 200 μL 피펫 팁을 사용하여, 공동단일 콜로니 형질 세포의 소량 llect. 마크와 검증 후 하류에 사용하기 위해 원래의 식민지를 유지합니다.
- 미세 원심 튜브에 뉴 클레아없는 물 50 μL에 수집 된 세포를 추가합니다. 로 pipetting 아래로 섞는다.
- 벤치 - 탑 열 순환기 또는 끓는 물 중탕을 사용하여 2 분 동안 95 ℃로 가열하여 세포를 용균.
- 하는 DNA를 주형으로 가열 용해 셀 1 μL를 사용하여 파지 미드 영역을 PCR 증폭은 -. PCR의 조건으로 열 순환기 프로토콜은 표 6 및 표 7에 제공된다. 검증 프라이머 서열에 대한 추가 pAB.001 시퀀스 파일을 참조하십시오.
- 올바른 GUIDE 서열의 혼입을 확인하기 위해 PCR 생성물 시퀀스.
- E.의 5 ml의 문화를 접종 순서 검증 스르나 발현 파지 미드를 운반하는 대장균 클론. 선택적 LB 배지에서 진탕 37 ° C에서 하룻밤 세포를 성장.
- 글리세롤 주식을 준비순서 검증 된 클론의. 스크류 캡 냉동 관에 60 % 글리세롤의 250 μL에 하룻밤 문화의 750 μl를 추가합니다.
- 무기한 -80 ° C에서 글리세롤 주식을 저장합니다. 밤새 배양의 나머지 단계 2에서 스르나 발현 파지 미드의 소스로 사용될 수있다.
M13 패키지 파지 미드 주식 2. 생산 및 수확
- E.의 5 ml의 문화를 준비 대장균은 스르나 발현 파지 미드를 운반. 적절한 항생제로 LB 배지에서 진탕 37 ° C에서 하룻밤 세포를 성장. 참고 : 스르나 발현 파지 미드 단계 1.1.5에 설명 된대로 드 노보 복제를 통해 얻은, 또는 수정 기존 파지 미드에서 단계 1.2.16에 수확 할 수있다.
- 마찬가지로 E.의 5 ml의 문화를 준비 대장균은 M13KO7 도우미 플라스미드를 들고. 선택적 LB 배지에서 진탕 37 ° C에서 하룻밤 세포를 성장.
- 추출 및 스르나의 expressio을 정화N 개의 파지 미드 및 DNA 추출 키트 또는 유사한 방법을 사용하여 12 헬퍼 플라스미드.
- Cotransform 구입 또는 1 μL 스르나 발현 파지 미드 및 헬퍼 플라스미드를 각각 13 화학적 유능한 대장균을 준비 하였다. 두 구조에 대한 선택적 항생제와 LB 한천에 도금 cotransformants을 위해 선택합니다.
- 선택적 항생제와 LB의 cotransformed 변형의 단일 식민지에서 10 ml의 문화를 준비합니다. 8 ~ 12 시간 또는 밤새 진탕 37 ° C에서 품어.
- 10 분 동안 3,300 XG에 문화를 원심 분리기. 0.2 μm의 필터를 통해 뜨는 및 필터를 수집합니다. 주의 : 미디어 누출의 경우, 감염성 파지 입자를 파괴 희석 표백제 (0.5 %)로 부분을 청소.
- 4 ° C에서 포장 된 파지 미드 여과 액을 저장합니다. 주의 : 샘플을 활성의 손실없이 주 일 동안 유지 될 수있다.
F + 대상 세포의 3. 준비 음소거에 대한
- 침묵의 대상이되는 세포가 F의 pilus (14)을 표현할 수 있는지 확인합니다. 은 F pilus이 이미 존재하는 경우, 4 단계로 진행합니다.
참고 : E.의 일반적인 실험실 균주를 대장균은 게놈 또는 플라스미드에 F의 pilus의 존재를 나타 내기 위해 + F 또는 F '로 주석된다. - E.의 F + 균주를 확보 대장균 등 TOP10F로 '.
참고 : 대상 변형 접합 후 F-플라스미드 기증자로부터 분리하기 위해 고유 저항 마커를 수행했는지 확인하십시오. - A ~ F + 균주와 결합하여 F의 pilus을 소개하고, 대상 균주 및 F-플라스미드 기증자 (14) 모두 5 ml의 문화를 준비합니다. 적절한 항생제로 LB 배지에서 진탕 37 ° C에서 하룻밤 세포를 성장.
- 다음날 두 균주 희석 한 100 선택적 LB 5 ㎖ 및 진탕하면서 37 ℃에서 배양을 계속한다.
- 선택 해제를 측정하여 세포의 성장 단계를 결정벤치 탑 분광 광도계를 사용하여 600 nm의 (OD 600)에서 문화의 iCal의 밀도. 문화 0.3 OD (600)까지 약 2 시간 동안 세포를 로그 성장 단계 (15)를 나타내는 달성된다.
- 마이크로 원심 튜브에 3 접합 반응을 준비 : 0.5 ml의 F-플라스미드 기증자 + 0.5 ml의 목표 변형, 0.5 ml의 F-플라스미드 기증자 + 0.5 ml의 LB 미디어 (음성 대조군) 0.5 ml의 대상 균주 + 0.5 ml의 LB 미디어 (음성 대조군). 공액가 진탕 37 ° C에서 2 시간 동안 진행 할 수 있습니다.
- 플레이트 F 플라스미드 특정 항생제 (전형적으로 테트라시 클린) 및 대상 균주 선택적 LB 한천 배지에 각각 접합 반응 100 ㎕. 도 기증자 또는받는 사람의 피로가 모두 항생제 저항을 표현하는지 확인하려면 음성 대조군 반응 플레이트.
음소거를위한 패키지 파지 미드 4. 감염
- LB의 메디칼의 5 ml의 문화에 F + 대상 세포의 단일 콜로니를 접종적절한 항생제와 함께. 진탕 37 ° C에서 밤새 품어.
- 다음 날, F + 대상 세포 1을 희석 : 100 선택적 LB 미디어 ml의 5 떨고와 37 ° C에서 문화를 계속합니다.
- 탁상용 분광 광도계를 이용하여 600 nm의 (OD 600)에서 배양액의 흡광도를 측정하여 세포의 성장 단계를 결정한다. 문화 0.3 OD (600)까지 약 2 시간 동안 세포를 로그 성장 단계 (15)를 나타내는 달성된다. 주 : F의 pilus 감염 효율의 발현 로그 단계에서 가장 높은 것입니다.
- 목표 인구의 약 99 %가 감염을 달성하는 100 : 1의 부피비로 표적 세포 (단계 2.6)을 패키지 M13 파지 미드를 추가한다. 감염이 30 ~ 60 분 동안 진탕 37 ° C에서 할 수있게합니다.
- 선택의 방법에 따라 스르나 - 침묵 표현형 분석.
참고 : 형광 단백질 대상의 경우, 침묵 효과를 정량화 할 수있다직접 형광 측정 (10)에 의해. 선택적으로, 표현형 분석은 유전자 녹다운 표현형 (8)의 결과를 관찰하기 위해 사용될 수있다. - 단계 1.2.14-1.2.16 다음 스르나 발현 파지 미드 호스트 글리세롤 주식을 준비합니다. 참고 : 파지 미드 무기한 호스트 변형에 전파하고 기존의 플라스미드에 항생제 비슷한으로 유지 될 수있다.
액체 미디어에서 mKate2 형광의 침묵
그림 1은 스르나 카세트 디자인, 파지 미드 벡터화, 그리고 침묵기구와이 작품에 설명 스르나 매개 knockdowns에 대한 방식을 보여줍니다. 프로토콜 1.2에 따라, 플라스미드 pAB.001의 스르나 침묵 카세트 mKate을 대상으로 변경되었다. 스르나 카세트 synthetized 및 파지 미드 Litmus28i_J23115-B0032-GFP, 모니카 오티즈와 드류 앤디 (11)의 선물에 클로닝 하였다. 이 파지 미드 성공적인 감염이 추적 될 수 있도록 GFP 발현 및 카나마이신 저항성 마커를 운반한다. 포장 파지 미드 주식은 프로토콜 2에 따라 제조 하였다.
E.의 유도체 구조적으로 표현, 염색체 통합 mKate2 마커를 운반하는 대장균 K12 MG1655은 접합에 의해 파지 감염을 제조 하였다 미드 로그 상 및 파지 미드 감염 후 프로토콜 4. 다음 도입 파지 미드로 성장시켰다 프로토콜 3. 세포를 후속하는 F 플라스미드 공여체 균주 200 ㎕의 배양 물은, 형광 플레이트 판독기로 전송하고, 형광은 24 시간 동안 연속적으로 모니터링 하였다.
그림 3은 mKate2 식에 스르나 매개 침묵의 효과를 보여줍니다. 항 mKate2의 파지 미드에 감염 균주 배경 mKate2 검출 가능한 형광 없었다. 대조적으로,이 균주는 파지 미드의 성공 흡수를 나타내는 GFP 마커를 표현했다. 감염되지 않은 대조군 세포는 형광 mKate2 아니라 GFP를 생산했다. 안티 mKate2 타겟팅 도메인 시퀀스 타겟팅 CAT로 치환되는 추가적인 제어는 mKate2 형광에 아무런 영향을 미치지 않았다.
한천 플레이트에 클로람페니콜 저항의 침묵
내용 : 1 "> 프로토콜 1.1에 따라, 스르나 침묵 카세트는 CAT를 대상으로 제작되었다"FO 유지-together.within 페이지는 = "는 구조적으로 표현, 염색체 통합 CAT 마커를 준비했다 운반하는 대장균 K12 MG1655의 유도체. 프로토콜 3. 셀에 후속하는 F-플라스미드 공여체 균주와 접합하여 파지 감염 미드 로그 상으로 성장하고, 파지 미드를 37 ° C에서 배양 한 시간 후에 프로토콜 4. 다음 도입 감염된 세포를 연속적으로 희석하고, 플레이 팅 된 클로람페니콜 농도. 플레이트 범위 밤새 배양하고, 각각의 농도에서 클로람페니콜 내성 세포의 비율 집락 형성 단위 (CFUs) 다음날 계수하여 결정 하였다.도 4는 클로람페니콜 내성 표현형 스르나 매개 된 침묵 화 효과를 나타낸다. 파지 미드는 mKate2 타겟팅 감염 감염되지 않은 세포 또는 세포에 내성이었다테스트 한 모든 농도에서 클로람페니콜. 대조적으로, 세포를 파지 미드 타겟팅 CAT 감염 클로람페니콜 낮은 농도에서 생존을 감소 보였고, 약 99 %보다 높은 농도에서 사망. 암피실린의 추가는 파지 미드를 들고 만 세균에 대한 선택은 발견 할 수없는 수준으로 클로람페니콜 생존을 감소시켰다. 이 파지 감염으로부터의 탈출을 보여주는 것은 10 침묵 탈출하는 일반적인 경로 인 이전 작업과 일치한다.
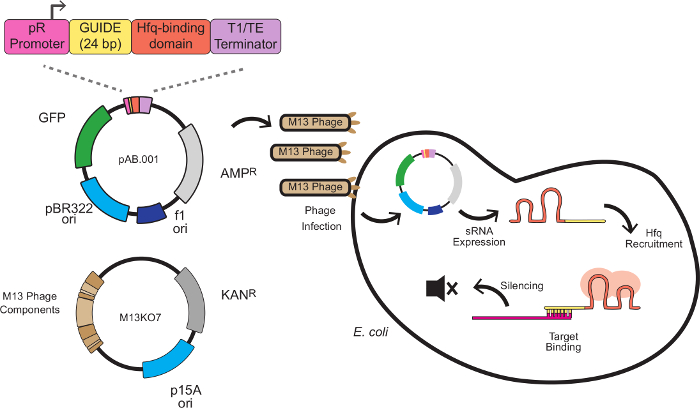
그림 1 : E.의 유전자 침묵 스르나 식 카세트와 대장균은 M13 파지에 의해 전달되는 스르나 카세트 4 모듈로 구성되어 있습니다. 홍보 프로모터 (박테리오파지 람다에서 파생 된 구성 적 프로모터), 24 bp의 대상 도메인, Hfq 바인딩 도메인은 MICC에서 추출 전사 터미네이터 (8) E.을 감염시킬 수 있습니다 대장균 스르나 발현이 시작 F의 pilus을 표현. 스르나는 (빨간색에 도시)에 Hfq 단백질을 모집하고 번역 억압과 mRNA의 분해 결과, 리보솜 결합 부위 근처 안티센스 mRNA의 목표를 결합한다. 이 그림의 더 큰 버전을 보려면 여기를 클릭하십시오.
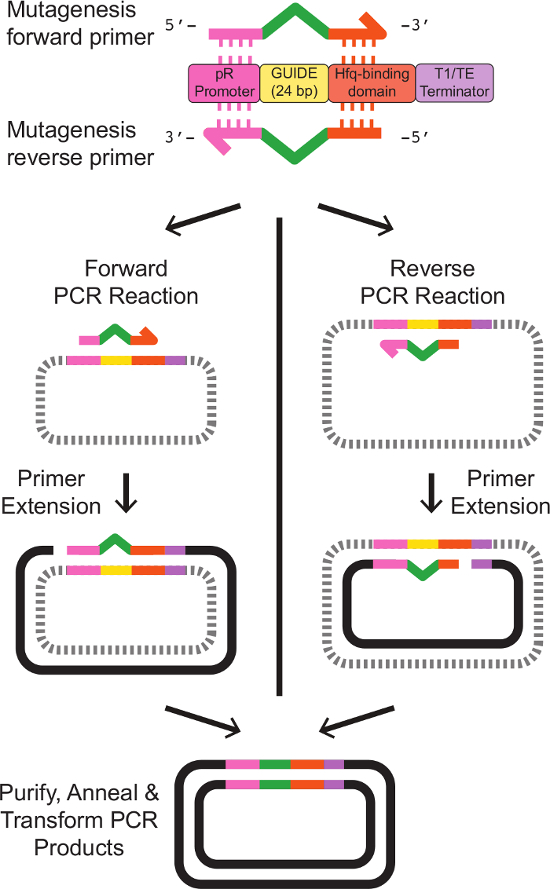
그림 2 :. 프라이머 디자인과 스르나 대상 사이트의 부위 특이 적 변이는 두 프라이머는 기존 스르나 카세트에 부분 동성으로 설계되었습니다. 순방향 프라이머는 다음 5 '말단의 PR 프로모터, 20 bp의 상동를 포함새로운 GUIDE 서열을 나타내는 24 염기쌍의 3 '말단에서 Hfq 결합 도메인을 다음 18 BP 상동. 역방향 프라이머는 새로운 GUIDE 서열의 역 상보 측부 기존 스르나 카세트에 상 동성 영역으로 순방향 프라이머 정반대 보수이다. 정확한 프라이머 서열은 표 3에 제시되어있다. 순방향 및 역방향 프라이머 별개의 단일 프라이머 PCR 반응은 원하는 변형 시퀀스의 선형 단일 가닥 DNA를 제조. 프로토콜에 설명 된대로 청소 다음, 앞으로 어닐링과 반응 생성물 역, 원하는 수정 스르나 카세트 베어링 이중 가닥 플라스미드 DNA의 결과. 이 그림의 더 큰 버전을 보려면 여기를 클릭하십시오.
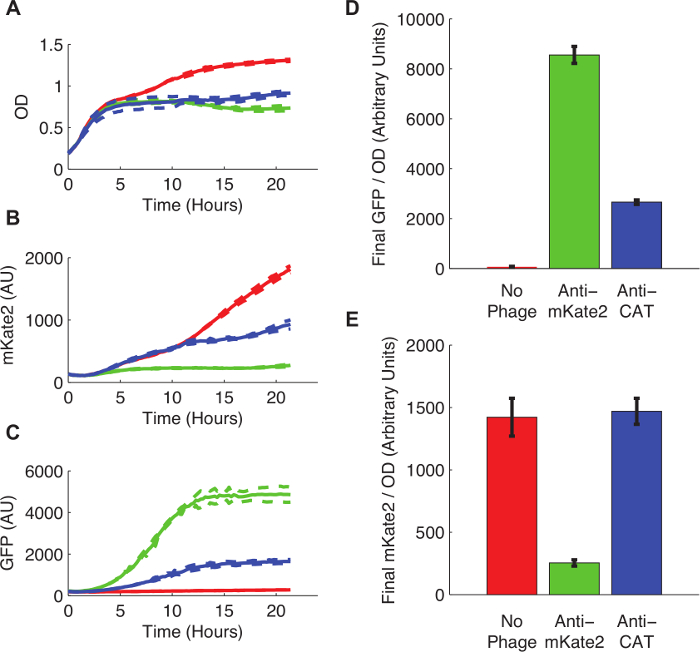
그림 3 : K염색체 통합 mKate2 형광 리포터의 nockdown. E. mKate2을 표현하는 대장균 MG1655 K12이었다 중 왼쪽 치료 (레드 라인과 바), 항 mKate2의 파지 미드 (녹색 선, 바)에 감염, 또는 제어 파지 미드 목표 CAT (블루 라인과 바)에 감염. (A) 비 처리 E. 콜라이 파지 감염에 대한 대사 비용을 나타내는 높은 포화 농도로 자랐다. 점선은 3 복제의 표준 편차를 나타냅니다. (B)이 mKate2 신호 방지 mKate2에 가까운 배경 수준으로 감소 하였다가 아니라 제어 균주, 균주를 처리 하였다. 또한, 파지 미드에 의해지지 (C) GFP 형광은 파지 미드 만 처리 제어에서 검출되었다. (D, E) 성장의 24 시간이 OD에 정상화 된 후 최종 형광 측정. GFP 신호는 파지 미드 감염을 나타내는 미처리 대조군 존재했지만, 실질적으로 방지 CAT 피 따라 감소hagemid 치료. 이 파지 미드의 오프 - 타겟 효과를 나타낼 수있다. mKate2 신호는 처리되지 않은 대조군과 비교 반 mKate2 파지 미드의 처리에 의해 감소되었다. 고양이 타겟 제어 파지 미드는 mKate2 형광에 영향을 보이지 않았다. 오차 막대가 3 복제의 표준 편차를 나타냅니다. 이 그림의 더 큰 버전을 보려면 여기를 클릭하십시오.
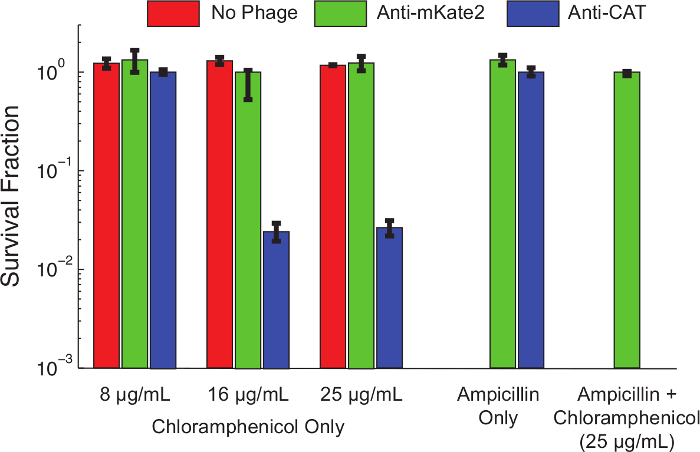
그림 4 : CAT의 최저은 유전자 저항하는 인구에 클로람페니콜 감도를 복원 E.합니다. 염색체 통합 CAT 유전자를 발현하는 대장균 MG1655 K12은 mKate2 (녹색 막대)를 대상으로 제어 파지 미드 치료, 치료 (빨간색 막대)를 왼쪽 또는 반 CAT 스르나 (파란색 막대)를 발현하는 파지 미드로 처리 하였다. 표시된 개미 1 감염의 시간, 생존 후ibiotics 시리얼 희석하고 도금에 의해 평가 하였다. 제어 처리가 영향을받지 동안 안티 CAT의 파지 미드로 처리 균주는 상당히 높은 농도에서 클로람페니콜에 의해 (> 90 %) 사망했다. 문화 판에 추가 암피실린은 긍정적으로 파지 미드 감염 선택하고 감염되지 않은 세포를 제거합니다. 이러한 조건에는 클로람페니콜 저항성 콜로니 항 CAT 처리 후에 관찰되지 않았다. 이것은 대부분의 생존자 감염 실패보다는 침묵의 실패를 나타내는 것을 나타낸다. 오차 막대가 3 복제의 표준 편차를 나타냅니다. 이 그림의 더 큰 버전을 보려면 여기를 클릭하십시오.
| CAT 표적 서열 | 5 '- ATGGAGAAAAAAATCACTGGATAT - 3' |
| CAT 가이드 순서 | 5 '- ATATCCAGTGATTTTTTTCTCCAT - 3' |
표 1. 고양이 유전자의 예 표적과 GUIDE 순서는 역 보완 관계를합니다.
| PR 프로모터 | TAACACCGTGCGTGTTGACTATTTTACCTCTGGCGGTGATAATGGTTGC | ||||
| GUIDE 순서 | ATATCCAGTGATTTTTTTCTCCAT | ||||
| Hfq 결합 도메인 | TTTCTGTTGGGCCATTGCATTGCCACTGATTTTCCAACATATAAAAAGACAAGCCCGAACAGTCGTCCGGGCTTTTTT TCTCGAG | ||||
| T1 / TE 종료 | CTCGAGCCAGGCATCAAATAAAACGAAAGGCTCAGTCGAAAGACTGGGCCTTTCGTTTTATCTGTTTTTGTCGGTGAA CGCTCTCTACTAGAGTCACACTGGCTCACCTTCGGGTGGGCCTTTCTGCGTTTATA | ||||
표 2 :. 스르나 카세트의 시퀀스 구성 요소 각각의 서열은 5'-3 '기록됩니다. 전체 카세트 위해 이들 4 요소의 연결하며 292 염기쌍을 포함한다.
| 앞으로 프라이머 | 5 '- CTGGCGGTGATAATGGTTGC [GUIDE] TTTCTGTTGGGCCATTGC - 3' |
| 역방향 프라이머 | 5 '- GCAATGGCCCAACAGAAA [TARGET] GCAACCATTATCACCGCCAG - 3' |
표 3 : 프라이머 디자인은 기존 GUIDE 요소를 변경할 수있는 정방향 프라이머는 PR 프로모터의 지난 20 bp의 새로운 GUIDE 순서 및 Hfq 결합 도메인의 첫 번째 18 염기쌍을 포함한다.. 표적 서열은 GUIDE 서열의 정반대 보완. 역방향 프라이머는 순방향 프라이머의 정확한 반대 보완.
표 4 : 단일 프라이머 변이원성 PCR에 대한 제안 조건.
| 단계 | 온도 | 시각 |
| 초기 변성 | 98 ° C | 30 초 |
| 30 회 | 98 ° C | 10 초 |
| 55 ° C | 30 초 | |
| 72 ° C | 120 초 | |
| 최종 확장 | 72 ° C | 300 초 |
| 저장 | 10 ° C |
표 5 : 단일 프라이머 변이원성 PCR을 위해 열 순환기 프로토콜 제안.
| 구성 요소 | 음량 |
| 템플릿 DNA | 1 μL |
| 10 μM 정방향 프라이머 | 0.5 μL |
| 10 μM 역방향 프라이머 | 0.5 μL |
| DNA 형성 촉매 2X 마스터 믹스 | 25 μL |
| 클레아없는 물 공급아르 자형 | 23 μL |
| 총 볼륨 | 50 μL |
표 6 : 순서 확인 PCR에 대한 제안 조건.
| 단계 | 온도 | 시각 |
| 초기 변성 | 95 ° C | 30 초 |
| 30 회 | 95 ° C | 30 초 |
| 55 ° C | 30 초 | |
| 68 ° C | 30 초 | |
| 최종 확장 | 68 ° C | 300 초 |
| 저장 | 10 ° C |
도표 7 : 열 순환기 프로 추천시퀀스 확인 PCR에 대한 로토콜.
본 방법은, 불특정 대조군과 비교 mKate 형광 수준에서 80 % 감소를 달성했다. 이것은 완전한 침묵이 관찰되지 않고 50-90% 효율 16,17 전형적인 다른 RNA 넉다운 방법과 일치한다. 표현형 수준에서 CAT-대상 knockdowns 크게 클로람페니콜 저항을 약화, 일부 조건에서 그것을 제거 할 수 있었다.
최저 표현형은 몇 시간 동안 감염 후 (도 3b) 후에 집단 수준에서 검출되었다. 이 파지 기반 전달의 중요한 특징을 보여 높은 최저 주파수가 이전 유전자 변형없이 일괄 문화를 직접 얻을 수있다. 플라스미드 변환 또는 게놈 통합을 사용하여 기존의 유전자 변형과는 달리, 파지 감염 인구가 하나의 고립 된 식민지에서 다시 재배 할 필요가 없습니다. 이 파지 감염의 효과 EXPLOR 수 있도록18, 또는 유 전적으로 혼합 된 자연 인구 19 생물막 같은 기존의 공간 구조와 복잡한 공간 역학 (11)와 인구 에디션.
이 방법에서 중요한 단계 높은 역가의 포장 파지 미드의 생산이다. 파지 입자 생산과 관련된 대사 부담은 파지 미드 생산 균주의 돌연변이 나 플라스미드 손실의 높은 속도로 이어질 수 있습니다. 파지 미드 생산 균주는 단일 cotransformed 식민지에서 직접 배양 및 냉장하지, 냉동 또는 파지 수확하기 전에 서브 배양하는 것이 좋습니다. E.에 파지 미드 및 도우미 플라스미드를 도입 할 때 공동 변환의 낮은 효율을 관찰 할 수있다 동시에 대장균. 이 경우, 더 높은 효율성은, 제 헬퍼 플라스미드를 변형 파지 미드 후속 변환을위한 헬퍼 플라스미드 베어링 적격 세포를 제조함으로써 얻을 수있다.
PHAgemid 감염 스르나 발현은 표적 세포에 대한 검출 가능한 대사성 부담을 부과 일부 표현형 교란을 초래할 수있다. 세포는 파지 미드 타겟팅 CAT (도 3)에 감염되었을 때, 예를 들어, 형광 mKate2의 저하도 관찰되었다. M13 감염은 E.의 전신 스트레스 반응을 유발하는 것으로 생각되지 않는다 대장균 20 만 간접적으로 전사 패턴을 바꿀 수 있습니다. 대안 적으로, 파지 미드에 포함 된 GFP 또는 암피실린 내성 마커 mKate2 식 9과 성장을 감소 셀룰러 리소스를 위해 경쟁 할 수있다. 마지막으로, 스르나 카세트 자체가 세계적으로 Hfq 단백질을 적정에 의해 유전자 발현 프로파일을 변경, 또는 오프 대상 mRNA의 입을 통해 할 수있다. 오프 대상 효과는 생체 RNAi의 21 ~ 23을 대상으로 일반적인, 그러나 그들은 체계적으로이 시스템에 대한 조사가 아직.
이 방법의 한 가지 제한은 감염 effi입니다ciency 일부 비 감염된 박테리아 집단에서 유지 할 수 있도록 100 % 미만이다. 이 작품 이전 작업 (10)의 결과는 비감염 세포가 최종 인구 1-10 %를 차지 제안하고, 관찰 nonsilenced 표현형의 대부분에 대한 책임이 있습니다. M13 저항에 다양한 경로는 pilus 식 (24)의 가장 보편적 인 돌연변이 손실 알려져있다. 이러한 제한의 관점에서, 컨트롤은 높은 감염 속도와 최저 효율을 확인하기 위해 사용되어야한다.
일부 응용 프로그램에 대한 또 다른 잠재적 인 제한은 헬퍼 파지를 오염 가끔 전송합니다. M13K07가 돌연변이 된 패키징 신호를 포함하지만,이 파지 생산 및 초기 감염 이벤트 (25)를 넘어 파지의 지속적인 보급 용 적격 세포 결과 저주파수 파지 캡시드로 패키징 감염된 인구에 전송 될 수있다. 헬퍼 파지에 대한 수정 효과 입증감소 된 파지 생산 (26)의 비용으로 비록 때때로, 비특이적 포장을 줄이는.
엔지니어링 박테리오파지는 E.위한 필수적인 도구가되었습니다 인구 증가에 새로운 유전자의 빠른 전송을 허용하는 대장균 합성 생물학. 최근의 연구는 세포 간 통신 회로 (11)를 생산 또는 항생제 내성을 억제하는 전사 인자를 표현 (27) 진학했다. 여기에 제시된 프로토콜은 프로그램 RNA를 통해 세균의 생리를 제어 할 수있는 도구의 성장 컬렉션에 추가합니다. 클레아 제 활성을 제거하기 위해 변이 할 때 CRISPR-카스의 뉴 클레아는, RNA 유도 유전자 목표 17,28에서 전사를 억제하는 것으로 나타났다. 반면, 스르나의 침묵은 번역 수준에서 작동 및 외래 단백질의 발현을 필요로하지 않습니다. 차세대 생명 공학 복잡한 pheno을 프로그램 파지 매개 배달 전사 및 번역 제어를 결합 할 수 있습니다실시간 유형.
The authors have nothing to disclose.
이 작품에 대한 자금은 파리 Bettencourt iGEM 팀의 지원에 파운데이션 부문 Bettencourt SCHUELLER에 의해 제공되었다. 우리는 기술 지원과 조언을 INSERM의 U1001 연구 장치와 샹탈 Lotton 감사합니다. 파지 미드 Litmus28i_J23115-B0032-GFP는 모니카 오티즈와 스탠포드의 드류 앤디에 의해 제공되었다.
| Name | Company | Catalog Number | Comments |
| Plasmid Miniprep Kit | Qiagen | 27104 | |
| DpnI Enzyme | NEB | R0176S | |
| Phusion High Fidelity Polymerase | NEB | M0530S | |
| Taq 2x Master Mix | NEB | M0270L | |
| M13KO7 Helper Phage | NEB | N0315S | |
| DH5α Competent Cells | Life Technologies | 18265-017 | |
| TOP10F' Cells | Life Technologies | C3030-03 | |
| LB Broth | Sigma | L3022-250G | |
| Ampicillin | Sigma | A9393-5G | |
| Kanamycin | Sigma | 60615-5G | |
| Chloramphenicol | Sigma | C0378-5G | |
| Tetracycline | Sigma | 87128-25G |
- Ohkumo, T., Masutani, C., Eki, T., Hanaoka, F. Use of RNAi in C. elegans. RNAi. , 129-137 (2008).
- Fire, A., Xu, S., Montgomery, M. K., Kostas, S. A., Driver, S. E., Mello, C. C. Potent and specific genetic interference by double-stranded RNA in Caenorhabditis elegans. Nature. 391 (6669), 806-811 (1998).
- Iordanou, E., Chandran, R. R., Blackstone, N., Jiang, L. RNAi interference by dsRNA injection into Drosophila embryos. J Vis Exp. (50), e2477 (2011).
- Ramadan, N., Flockhart, I., Booker, M., Perrimon, N., Mathey-Prevot, B. Design and implementation of high-throughput RNAi screens in cultured Drosophila cells. Nat Protoc. 2 (9), 2245-2264 (2007).
- Tsong, T. Y. Electroporation of cell membranes. Biophys J. 60 (2), 297-306 (1991).
- Kim, W. J., Chang, C. -. W., Lee, M., Kim, S. W. Efficient siRNA delivery using water soluble lipopolymer for anti-angiogenic gene therapy. J Control Release. 118 (3), 357-363 (2007).
- Shi, Y. Mammalian RNAi for the masses. Trends Genet. 19 (1), 9-12 (2003).
- Na, D., Yoo, S. M., Chung, H., Park, H., Park, J. H., Lee, S. Y. Metabolic engineering of Escherichia coli using synthetic small regulatory RNAs. Nat Biotechnol. 31 (2), 170-174 (2013).
- Ceroni, F., Algar, R., Stan, G. -. B., Ellis, T. Quantifying cellular capacity identifies gene expression designs with reduced burden. Nat Methods. 12 (5), 415-418 (2015).
- Libis, V. K., Bernheim, A. G., et al. Silencing of Antibiotic Resistance in E. coli with Engineered Phage Bearing Small Regulatory RNAs. ACS Synth Biol. 3 (12), 1003-1006 (2014).
- Ortiz, M. E., Endy, D. Engineered cell-cell communication via DNA messaging. J Biol Eng. 6 (1), 16 (2012).
- Edelheit, O., Hanukoglu, A., Hanukoglu, I. Simple and efficient site-directed mutagenesis using two single-primer reactions in parallel to generate mutants for protein structure-function studies. BMC Biotechnol. 9 (1), 61 (2009).
- Chung, C. T., Miller, R. H. Preparation and storage of competent Escherichia coli cells. Recombinant DNA Part I. , 621-627 (1993).
- Phornphisutthimas, S., Thamchaipenet, A., Panijpan, B. Conjugation in Escherichia coli. Biochem Mol Biol Educ. 35 (6), 440-445 (2007).
- Sezonov, G., Joseleau-Petit, D., D'Ari, R. Escherichia coli Physiology in Luria-Bertani Broth. J Bacteriol. 189 (23), 8746-8749 (2007).
- Mittal, V. Improving the efficiency of RNA interference in mammals. Nat Rev Genet. 5 (5), 355-365 (2004).
- Qi, L. S., Larson, M. H., et al. Repurposing CRISPR as an RNA-guided platform for sequence-specific control of gene expression. Cell. 152 (5), 1173-1183 (2013).
- Lu, T. K., Collins, J. J. Dispersing biofilms with engineered enzymatic bacteriophage. Proc Natl Acad Sci USA. 104 (27), 11197-11202 (2007).
- Yosef, I., Manor, M., Kiro, R., Qimron, U. Temperate and lytic bacteriophages programmed to sensitize and kill antibiotic-resistant bacteria. Proc Natl Acad Sci USA. 112 (23), 7267-7272 (2015).
- Karlsson, F., Malmborg-Hager, A. -. C., Albrekt, A. -. S., Borrebaeck, C. A. K. Genome-wide comparison of phage M13-infected vs. uninfected Escherichia coli. Can J Microbiol. 51 (1), 29-35 (2005).
- Senthil-Kumar, M., Mysore, K. S. Caveat of RNAi in plants: the off-target effect. Methods in molecular biology. 744, 13-25 (2011).
- Jackson, A. L., Linsley, P. S. Recognizing and avoiding siRNA off-target effects for target identification and therapeutic application. Nat Rev Drug Discov. 9 (1), 57-67 (2010).
- Cho, S. W., Kim, S., et al. Analysis of off-target effects of CRISPR/Cas-derived RNA-guided endonucleases and nickases. Genome Res. 24 (1), 132-141 (2014).
- Hagens, S., Blasi, U. Genetically modified filamentous phage as bactericidal agents: a pilot study. Lett Appl Microbiol. 37 (4), 318-323 (2003).
- Kasman, L. M., Kasman, A., Westwater, C., Dolan, J., Schmidt, M. G., Norris, J. S. Overcoming the phage replication threshold: a mathematical model with implications for phage therapy. J Virol. 76 (11), 5557-5564 (2002).
- Chasteen, L., Ayriss, J., Pavlik, P., Bradbury, A. R. M. Eliminating helper phage from phage display. Nucleic Acids Res. 34 (21), e145 (2006).
- Lu, T. K., Collins, J. J. Engineered bacteriophage targeting gene networks as adjuvants for antibiotic therapy. Proc Natl Acad Sci USA. 106 (12), 4629-4634 (2009).
- Bikard, D., Jiang, W., Samai, P., Hochschild, A., Zhang, F., Marraffini, L. A. Programmable repression and activation of bacterial gene expression using an engineered CRISPR-Cas system. Nucleic Acids Res. 41 (15), 7429-7437 (2013).
ABOUT JoVE
Copyright © 2024 MyJoVE Corporation. All rights reserved