Reproducible dsRNA Microinjection and Oviposition Bioassay in Mosquitoes and House Flies
In This Article
Summary
This protocol describes a microinjection methodology that we have standardized and used for several years to deliver specific quantities of nucleic acids directly to the hemolymph of mosquitoes and house flies. This protocol results in minimal injection mortality and allows dose correlated measurements of fecundity.
Abstract
Synthetic dsRNAs, used to induce RNA interference, may have dose dependent phenotypic effects. These effects are difficult to define if the dsRNAs are delivered using a non-quantitative method. Accurate delivery of known quantities of nucleic acids or other chemicals is critical to measure the efficacy of the compound being tested and to allow reliable comparison between compounds.
Here we provide a reproducible, quantitative microinjection protocol that ensures accurate delivery of specific doses of dsRNA, reducing the mortality typically induced by injection injury. These modifications include the addition of Rhodamine B, a graduated injection needle, and an improved recovery method borrowed from Isoe and Collins. This method allows calculation of dose responses and facilitates comparisons between compounds. Versions of this method have been successfully used on three genera of mosquitoes as well as house flies to assess the reduction in fecundity resulting from gene silencing of ribosomal RNA transcripts.
This protocol provides strategies to reduce several challenges of small insect microinjection. Together, mechanical delivery of dsRNAs accompanied by visual verification, identification of effective locations for delivery, and inclusion of a post-injection recovery period ensure accurate dosing and low injury mortality. This protocol also describes an oviposition bioassay for uniform determination of effects on fecundity.
Introduction
Delivering small biomolecules, such as nucleic acids, to adult dipterans has proven to be challenging in both Culicidae and Muscidae. Oral uptake of dsRNAs has been reported to produce sterility (when targeting genes exclusively expressed in testes of adults) and mortality (when targeting SNF7 and a steroid receptor coactivator) in larval Aedes aegypti1,2. Mortality has also been observed when targeting HSP70 in larval Musca domestica3. Such phenotypic effects, however, have not resulted after feeding dsRNA to adult Ae. aegypti in sugar meals4,5.
Microinjection has been used to circumvent the midgut when introducing pathogens or nucleic acids, thereby inducing a systemic response5,6,7,8,9,10. Several microinjection techniques exist, involving equipment ranging from in-house produced apparatuses that require visual measurement of injection volume to microprocessor-controlled injectors that allow for automated volume delivery as low as 2.3 nL5,9,10,11. RNA interference (RNAi) triggers targeting ribosomal mRNAs, disrupt ovarian development in arthropods as diverse as the cattle tick Rhipicephalus microplus, the mosquitoes Aedes aegypti and Culex pipiens, and the house fly Musca domestica5,9,12,13. In these studies, monitoring the disruption of oviposition was essential for determining the efficacy of the inoculant as the phenotype may manifest as termination or reduction of progeny. This is a form of multigenerational lethal phenotype that is a critical and desired effect in many of the non-traditional biocontrol methods like Wolbachia-infected males introduction (sexual incompatibility) and RNAi induced sterility5,9,14. Tracking both mortality and fecundity is necessary for the characterizing and development of highly specific biorationals (naturally occurring pathogens and/or natural derivatives)15.
This protocol presents detailed microinjection techniques for both adult mosquitoes and house flies; a process that is often not well described in the literature. In addition, oviposition bioassay methodologies to adequately evaluate dsRNA effects on adult dipterans are described. These protocols were developed specifically for Aedes aegypti and Musca domestica but can be modified for other species.
Protocol
1. Insect Preparation
- Anesthetize mosquitoes by chilling them at 4 °C for several hours or by 2 min exposure to CO2 and then holding at 4 °C prior to injections. Cold anesthetize flies at 4 °C 2‒3 h prior to injections. Ensure mosquitoes and flies are mated if oviposition is a response variable.
NOTE: The mosquito anesthetization method depends upon the species and strain, e.g., Culex recover much quicker from cold anesthetization than Aedes aegypti and therefore CO2 knockdown is preferable. - On ice, carefully stage mosquitoes or flies by using soft forceps to lay them ventrally exposed on microscope slides ~0.5 cm apart. Standard slides can hold two rows of 6 mosquitoes each or 6–7 house flies. Leave a 1–1.5 cm space at the left or right end of the slide clear of mosquitoes or flies to aid slide handling.
- Keep slides of staged insects at 4 °C in large Petri dishes until ready for injection.
NOTE: Drilling a hole through the Petri dish lid will help prevent insects from being disturbed by air displacement when capping. Placing slides on a couple of glass capillaries secured to the dish bottom will ease removal from Petri dish.
2. Insect Injection
- Prepare dsRNA as described by Estep et al.5 and Sanscrainte et al.9. If free of contaminants, dsRNA can be quantified by using a spectrophotometer to measure the absorbance at 260 nm and calculating RNA concentration in μg/mL as A260 × dilution factor × 40.
- Set up dissecting microscope and microinjector over a chill table or shallow ice bucket to perform injections on the cold-anesthetized mosquitoes and flies (see step 1.1–1.3 and Figure 1).
- Pull glass capillaries to a fine tip with needle puller (settings: heater = 15 units, solenoid = 4 A).
NOTE: See Table of Materials for manufacturer’s details. - Place the capillary needle into a microinjector as per the manufacturer’s instructions and break needle tip by pinching with a pair of forceps to produce a sharp point.
NOTE: Suggested needle opening sizes are ~150 µm for mosquito and ~250 µm for housefly. - Set microinjector for desired delivery volume. Meniscus movement from ~50 nL aliquots is easily observable and recommended for mosquito microinjections. If available, enable fast injection settings. Rinse the needle by filling and expelling nuclease-free water 3 times.
- Prepare solutions for injection at a concentration such that 100‒150 nL will provide desired dose for mosquitoes and 500 nL will provide desired dose for house flies.
NOTE: Suggested initial doses: 1 µg for each mosquito and 5 µg for each housefly5,9. - Optionally for mosquitoes, add Rhodamine B (to aid in visualization) to solutions at a final concentration of 3.0 µg/mL.
NOTE: The dye will be clearly visible by its pink color after injection through the nearly clear cuticle on the ventral surface at the abdomen/thorax intersection. - Pipette 3–4 µL of solution to inject onto a clean surface (such as a piece of paraffin film) and draw into the glass needle without sucking in any air. Depress the inject button repeatedly until liquid begins to dispense from the needle and gently wipe droplets off with a delicate task wiper.
NOTE: While some models of microinjectors require backfilling the syringe, the injector referenced in the Table of Materials does not. - Using an ultra-fine point marker, draw hash marks ~1 mm apart starting from the liquid meniscus to the needle shank. This will aid in visualizing the meniscus movement and ensure that the solution has entered the mosquito during injection.
- Set the slide of mosquitoes or flies (as prepared in steps 1.2–1.3) under the needle on the chill table or ice (Figure 1). Ensure that field of view in the microscope is wide enough to see the solution meniscus in the needle.
- For mosquitoes, align the needle with the middle one-third of the mesokatepisternum (Figure 2A), and while bracing the mosquito against the needle with forceps placed on the opposite side, gently puncture the cuticle with the needle tip using the microinjector micrometer.
- For house flies, align the needle with the mesopleuron (Figure 2B) and, while bracing the fly against the needle, gently puncture the cuticle with the needle tip.
- Gently slide the needle into the mosquito or fly body until the tip has passed through the midline.
NOTE: Inserting the needle too shallow often results in the injected liquid beading out of the insect (see step 2.15). - While watching for movement of the liquid meniscus in the needle, depress the inject button until the desired amount of liquid has been injected. For mosquitoes, if set to 50.6 nL aliquots, press 2X for ~100 nL or 3X for ~150 nL. For house flies, depress the inject button until ~500 nL has been injected (i.e., with 69 nL aliquots press 7X for 483 nL).
- If the meniscus does not move, slowly slide the mosquito or fly off the needle while watching for meniscus movement. If a portion of the injected solution beads out of the cuticle upon needle removal or if the meniscus fails to move, discard the insect as this injection was not successful.
- Transfer successfully injected mosquitoes in groups of 10-15 or house flies in groups of 5 to clear 3.5 oz holding cups. Cover with tulle or netting and recover at room temperature.
- After mosquitoes and flies have recovered (1 – 2 h), invert each holding cup over a cotton ball soaked with 10% sucrose solution.
NOTE: The inversion of the cup on top of the sucrose cotton aids in survival by providing easier access to a sugar meal10. - Rinse needle as in step 2.5 between injection solutions. When done injecting, discard used needles in an appropriate sharps container.
3. Aedes aegypti mortality and oviposition bioassay
- For three days after injection, continue to provide mosquitoes with 10% sucrose and record the number of visibly dead mosquitoes.
- Fill a ≤12-inch artificial membrane (such as collagen sausage casing) with fresh blood (i.e., bovine for Aedes aegypti) and heat to 60 °C in hot water bath.
- Briefly dry the membrane by rolling on a paper towel and lay across the tulle caps of the 3.5 oz clear holding cups. To enhance feeding, spike blood with 1 mM ATP after heating as a phagostimulant or by handle the warmed blood sausage with clean bare hands to leave human volatiles on the casing surface.
- After blood feeding, replace the cotton ball soaked with 10% sucrose on top of the holding cups and allow mosquitoes to rest for ~24 h before transferring to oviposition cups.
- Construct oviposition cups by filling clear 3.5 oz bioassay cups with ~30 mL deionized H2O and placing a piece of seed germination paper (~4 cm wide and 5 cm long with ridges running vertically) at the bottom of the cup and against the side (Figure 3A). Cover with tulle or netting and cut a small slit (~1 cm) in the tulle or netting cap.
- At 24 h post-blood feeding, cut a small slit (~1 cm) in the tulle cap of the holding cup and transfer only individual females that successfully fed — with a visible blood bolus in abdomen — by gentle mouth aspiration into oviposition cups (1 female/oviposition cup). Seal the small cut in the oviposition cap with a 10% sucrose saturated cotton ball.
- Hold cups at ~27 °C for 5‒7 days to allow mosquitoes to fully oviposit while tracking daily mortality. Count eggs under a dissecting scope.
4. Musca domestica mortality and oviposition bioassay
- For three days after injection, continue to provide the flies with 10% sucrose and record the number of visibly dead flies.
- Three days after injection, anesthetize flies by briefly placing a polyethylene tube dispensing CO2 over the holding cups until all flies are motionless at bottom of the cups.
- Transfer flies to a clean cage comprised of a 4 L plastic jar with a stockinette sleeve covering the opening (10 cm, Figure 3B). Provide flies with water and a mixture of granulated sucrose, powdered milk, and dried egg yolk in an 8:8:1 ratio (by volume) for four days, recording and removing dead flies daily.
- Prepare dry ingredients for fresh fly larval rearing medium by mixing 75% wheat bran with 25% pelleted livestock feed by weight. Add water to reach 62% moisture (until a drop of water can barely be squeezed out).
- Prepare a 2.5 cm diameter ball from a 1:1 mixture of the above fresh larval media and “used” fly larval medium (medium in which flies have previously pupated). Wrap the ball in a square of black cotton cloth, squeeze lightly until medium liquid seeps through, then use rubber bands to hold the cloth in place around the ball.
- Put the ball in a 60 mL cup and place it in the fly cage for 5 h (Figure 3B).
- Rinse eggs off the oviposition ball and ensure that the eggs are removed from under folds in the cloth. Shake eggs to disrupt clusters and transfer them to a graduated 20 mL centrifuge tube with a transfer pipet.
- Wait until the eggs settle and note the volume of the settled eggs in the graduated tube. Add sufficient water to bring the volume up to 20X the volume of settled eggs. Record the total volume of water plus eggs.
- While mixing the water and egg suspension with either a magnetic stir bar or vigorous stirring, use a 1 mL pipet tip (with the end of the tip cut off) to dispense 0.5 mL of the water and egg suspension onto a piece of pre-moistened black cloth in a series of lines. Count the eggs under a dissecting microscope.
- Repeat step 4.9 two more times. If one of the counts is a severe outlier (i.e., differs from the other two counts by >35%), make a fourth count and disregard the outlier.
- Calculate the mean number of eggs per sample and multiply this value by 2 to get the number of eggs/mL of the suspension. Multiply the eggs/mL value obtained by the volume of the egg suspension from step 4.8. This is the estimate for the total number of eggs laid.
Representative Results
Microinjection of dsRNA in mosquitoes
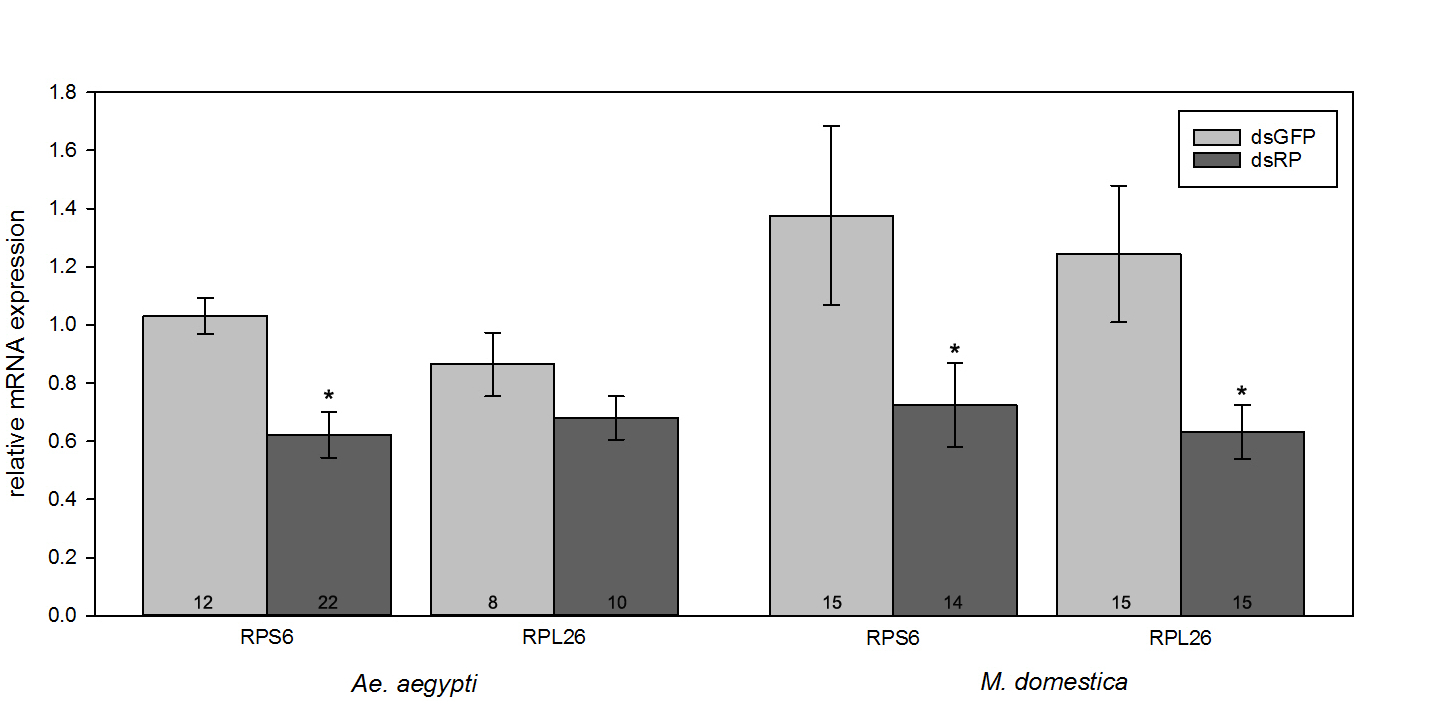
This microinjection method has been used in our laboratory to evaluate gene expression, mortality and oviposition response for over 80 dsRNA triggers across several mosquito genera (Aedes, Anopheles, Culex). Injecting females from the colony strain of Ae. aegypti (ORL1952) with 1 µg of dsRNA derived from the ribosomal transcripts RPS6 and RPL26 (see Estep et al.5 for dsRNA production methods and sequence data) showed significant reduction in relative expression (RE) across multiple oviposition cycles when compared to mosquitoes injected with a control dsGFP. Aedes aegypti RE measured following the 2nd gonotrophic cycle showed significant reduction of RPS6 expression (P < 0.05) in mosquitoes injected with dsRPS6 as determined by a Student's t-test between the dsGFP control of the same group (Figure 4)5.
Fecundity was assessed from individual females using the oviposition bioassay described here. After 3 days post-injection, mosquitoes were blood fed, transferred into individual oviposition containers, and allowed to lay to completion. Average clutch sizes for the first gonotrophic cycle were significantly reduced for both dsRPS6 treated Ae. aegypti (n = 102 females, 1.3 ± 0.8 (mean ± SE) eggs) and dsRPL26 treated (n = 86 females, 4.0 ± 1.1 eggs) when compared to dsGFP injections (n = 79 females, 49.3 ± 3.5 eggs, Figure 5A). Clutch assessment after a second gonotrophic cycle also showed significantly reduced oviposition for both dsRNA treated groups, but with reduced effect size. Group sizes of Ae. aegypti were variable and thus means separation was performed by Dunn's test5. Twenty-four-hour mortality was normally 3% or less.
A clear dose effect was observed when injecting dsRPS6 from 1 µg to 50 ng in female Ae. aegypti, and clutch size varied significantly when compared to 1 µg/female dsGFP injected controls in all but the 25 ng/female dsRPS6 dose (Figure 5B).
Microinjection of dsRNA in house flies
Colony Musca domestica (ORL normal) were injected with 5 µg of dsRNA constructs designed against the house fly RPS6 and RPL26 transcripts9. As was observed in Ae. aegypti, significant reduction of both specific transcript expression (RPS6 and RPL26) and reduction in clutch size was observed (Figure 4 and Figure 5A). Significant reduction in RE was determined by Student's t-test between the dsGFP control of the same group (P < 0.05) and the Holm-Sidak test was used to determine significance between clutch sizes for different M. domestica treatment groups. Ovarian dissections showed reduced oogenesis in dsRPS6 and dsRPL26 treatments, while dsGFP fed flies had normal vitellogenin deposition as rated on the Tyndale-Biscoe scale9.

Figure 1: Microinjection setup for delivering microvolumes to adult mosquitoes and house flies. Injections are performed on cold-anesthetized insects staged on microscope slides over a chill table. Please click here to view a larger version of this figure.
{kind=link}

Figure 2: Microinjection sites for Ae. aegypti and M. domestica. (A) Adult Ae. aegypti are injected in the middle one-third of the mesokatepisternum. (B) Adult M. domestica are injected in the mesopleuron. Arrows indicate sites of injection. Please click here to view a larger version of this figure.
{kind=link}

Figure 3: Oviposition bioassay setup. (A) Ae. aegypti oviposition assay cups with seed germination paper as an oviposition substrate, capped with tulle and 10% sucrose-soaked cotton. (B) Adult female M. domestica in an oviposition assay container with food (A), water (B), and larval media (C). Please click here to view a larger version of this figure.
{kind=link}

Figure 4: Relative expression of RPS6 in Ae. aegypti and M. domestica injected with species-specific dsRNAs. Aedes aegypti and M. domestica were injected with 1 and 5 µg respectively of either control dsRNA (dsGFP) or dsRNA designed against RPS6 or RPL26 transcripts from each insect. Significant reduction (P < 0.05) of RPS6 expression was observed in mosquitoes and flies injected with dsRPS6 and by flies injected with dsRPL26 (indicated by asterisks). Error bars represent mean ± SE and number at column base indicates number of individuals examined. This figure has been modified from Estep et al.5 and Sanscrainte et al.9. Please click here to view a larger version of this figure.
{kind=link}

Figure 5: Average clutch size of Ae. aegypti and M. domestica injected with species-specific dsRNAs. (A) Egg deposition was significantly reduced (P < 0.05) in both Ae. aegypti and M. domestica after injection of 1 and 5 µg respectively of either control dsRNA (dsGFP) or dsRNA designed against RPS6 or RPL26 transcripts from each insect. Asterisks indicate significant differences from the dsGFP control of the same group. Error bars represent mean ± SE and number at column base indicates number of individuals examined. This figure has been modified from Estep et al.5 and Sanscrainte et al.9. (B) Dose curve of Ae. aegypti injected with dsRPS6 from 50 ng/female to 1000 ng/female shows significant reductions in fecundity in comparison to dsGFP injected cohorts (50, 100, and 1000 ng/female). Error bars represent mean ± 95% CI from 36-81 individual organisms per dose. Points with letters represent significant difference between groups (P < 0.05) identified by ANOVA. This figure has been modified from Estep et al.5. Please click here to view a larger version of this figure.
{kind=link}
Discussion
Microinjection is a valuable laboratory technique to ensure delivery of dsRNA or other biorationals (i.e., pesticides, viruses, microsporidia). While many laboratories perform microinjection, the exact amount injected is often unclear due to technical limitations of the delivery system where delivered volume is not directly measured11. Delivered volume and concentration are critical parameters that allow calculation of standard toxicological measures such as EC50 or IC50 or to define minimal effective doses5,16. This is especially important in functional studies using RNAi in adult insects, where delivery by feeding does not always result in systemic exposure to the desired particles and the actual dose crossing the midgut is unclear5.
While the microinjection procedure can be initially challenging, rapid improvements in speed and delivery accuracy are achieved with practice and patience. Mastering the injection procedure itself is a time investment, but, once accomplished, the procedure produces repeatable results and injury mortality of <3%. The ratio of successful injections will increase as proficiency increases allowing injection of 300 mosquitoes or flies per day.
Many of the challenges of microinjection are due to the needle being used. Determining the proper capillary needle break point and tip angle is a challenging aspect of the procedure and requires some trial and error to determine the most effective opening size for a given species. The fine needle most useful for mosquito microinjection (~150 µm) is too small to quickly deliver the larger volume injected into flies while conversely, the larger needle opening that works on flies (~250 µm) causes extensive injury damage to the smaller mosquitoes and unacceptable injection mortality. A needle broken with a blunt tip is often difficult to get through the cuticle without tissue damage and increased mortality. Insect scales or tissue pieces can clog needles over the course of an experiment, so it is often helpful to have several needles pulled if replacement is required. We have found that it is easier to replace a difficult needle rather than spend time to try to clear a jammed or fouled needle.
Observing the injection solution entering the insect is critical so that unsuccessful injections can be removed (see steps 2.14-2.15). Rhodamine B addition to injection solutions provides a clear visual to ensure that cold-staged insects receive proper dosing and is especially helpful while practicing (see step 2.7).
The injection and oviposition bioassay methods presented here have successfully induced gene knockdown and measurable phenotypic effects in adult Ae. aegypti and M. domestica when using dsRNAs designed against species-specific ribosomal transcripts (Figure 4 and Figure 5A). Additionally, the ability of this method to accurately deliver microvolumes to individuals allows for dose response curves to be generated for small amounts of material (as low as 50 ng; Figure 5B). This is especially useful for methods such as screening siRNAs or dsRNAs, as producing large quantities of these molecules can be cost-prohibitive.
This method can be modified to inject biological agents (viruses, bacteria, microsporidia) or chemical pesticides, after ensuring that any delivery buffers are innocuous by injection. As delivery volume is limited by the size of the insect, producing concentrated testing solutions is of importance.
To adequately evaluate dsRNA efficacy, it is necessary to track oviposition as lethal phenotypes may only manifest in progeny. As presented here, holding female Ae. aegypti separately after blooding allows for individual clutch size determination and further testing on the same individuals can be done downstream of oviposition (i.e., gene expression studies, additional gonotrophic cycles, tissue specific knockdown) to correlate reproductive output with other measures such as gene expression. As M. domestica generally oviposit in a group response, inciting isolated gravid flies to lay is very labor intensive and not practical for large numbers of individuals17. Therefore, a method to determine mean clutch size is presented.
The microinjection method presented here provides consistent systemic delivery to mosquitoes and house flies. Coupled with mortality and oviposition tracking bioassays, these versatile tools can be used to assess the transcriptional and phenotypic effects of small RNA molecules, biological agents, and traditional pesticides where oral delivery is not viable.
Disclosures
All authors are employees or contractors of the United States Department of Agriculture or the Department of Defense. This manuscript was produced during the course of official duties and is by law in the public domain. The views expressed represent the opinion of the authors and do not represent an official position or endorsement by the US government. Mention of a commercial product is not a recommendation for use.
Acknowledgements
The authors thank Hanayo Arimoto (US Navy), Dana Johnson, Lucy Li, and Roxie White (USDA-ARS) for help with data acquisition and analysis. We also thank Drs. Pia Olafson (USDA-ARS) and Ke Wu (University of Florida) for critical review of the manuscript and Niklaus Hostettler (University of Florida) for filming the microscope footage. Funding was provided by the USDA, the WII-141C project of the Navy and Marine Corps Public Health Center, and the Deployed War Fighters Protection Program. The funders had no role in design or direction of the study or development of the manuscript.
Materials
| Name | Company | Catalog Number | Comments |
| needle pulling | |||
| vertical pipette puller | Kopf | 720 | settings: heater = 15 units, solenoid = 4 amps |
| glass capillaries, 3.5" long, ID = 0.530 mm ± 25 μm, OD 1.14 mm | World Precision Instruments | 504949 | capillaries for pulling glass needles |
| Name | Company | Catalog Number | Comments |
| microinjector station | |||
| Nanoliter 2010 | World Precision Instruments | NANOLITER2010 | microinjector |
| manual micromanipulator | World Precision Instruments | KITE-L | left hand KITE manipulator |
| magnetic stand | World Precision Instruments | M9 | holds micromanipulator |
| precision stereo zoom binocular microscope on boom stand | World Precision Instruments | PZMIII-BS | dissecting scope for microinjections |
| 1.5X objective | World Precision Instruments | 501377 | objective for microscope |
| light LED ring | World Precision Instruments | 504134 | light ring for injection microscope |
| laboratory chill table | BioQuip Products | 1431 | chill table for microinjections |
| microscope slides | Fisher Scientific | 12-544-1 | for staging insects while injections |
| Rhodamine B, 98+% | Acros Organics | AC296571000 | |
| Name | Company | Catalog Number | Comments |
| mosquito oviposition bioassays | |||
| large Petri dish | Fisher Scientific | FB0875714 | for holding staging slides |
| plastic cup - 3 1/2 oz. | Dart Container Corporation | TK35 | mosquito oviposition bioassay cups |
| matte tulle fabric | Joanne Fabrics | 1103068 | caps for oviposition bioassays |
| blood source | locally acquired | typically bovine or live chickens | |
| 1 x 30mm clear edible collagen casing | Butcher and Packer | 30D02-05 | |
| heavy weight seed germination paper | Anchor Paper Co | SD7615L | |
| oral aspirator with HEPA filter | John W. Hock Company | 612 | |
| Name | Company | Catalog Number | Comments |
| house fly oviposition bioassays | |||
| 4 L plastic food storage canister | Walmart | 555115143 | |
| 10-inch stockinette sleeve | Medonthego.com | FS15001H | |
| wheat bran | locally acquired | typically found at feed stores | |
| Calf Manna performance supplement | ValleyVetSupply.com | 16731 | pelleted livestock feed |
| dried egg yolk | BulkFoods.com | 40506 | |
| black cotton cloth | locally acquired | typically in craft supplies section | |
| 60 mL cup | Dart Container Corporation | P200-N |
References
- Whyard, S., et al. Silencing the buzz: a new approach to population suppression of mosquitoes by feeding larvae double-stranded RNAs. Parasites & Vectors. 8 (96), (2015).
- Das, S., Debnath, N., Cui, Y., Unrine, J., Palli, S. R. Chitosan, carbon quantum dot, and silica nanoparticle mediated dsRNA delivery for gene silencing in Aedes aegypti: a comparative analysis. ACS Applied Materials & Interfaces. 7 (35), 19530-19535 (2015).
- Tang, T., et al. Stress-induced HSP70 from Musca domestica plays a functionally significant role in the immune system. Journal of Insect Physiology. 58 (9), 1226-1234 (2012).
- Coy, M. R., et al. Gene silencing in adult Aedes aegypti mosquitoes through oral delivery of double-stranded RNA. Journal of Applied Entomology. 136 (10), 741-748 (2012).
- Estep, A. S., Sanscrainte, N. D., Becnel, J. J. DsRNA-mediated targeting of ribosomal transcripts RPS6 and RPL26 induces long-lasting and significant reductions in fecundity of the vector Aedes aegypti. Journal of Insect Physiology. 90, 17-26 (2016).
- Wen, D., et al. N-glycosylation of Viral E Protein Is the Determinant for Vector Midgut Invasion by Flaviviruses. mBio. 9 (1), (2018).
- Liu, Y., Cheng, G., Colpitts, T. Techniques for Experimental Infection of Mosquitoes with West Nile Virus. Methods in Molecular Biology. 1435, 151-163 (2016).
- Puglise, J. M., Estep, A. S., Becnel, J. J. Expression profiles and RNAi silencing of Inhibitor of Apoptosis transcripts in Aedes, Anopheles, and Culex mosquitoes (Diptera: Culicidae). Journal of Medical Entomology. 53 (2), 304-314 (2015).
- Sanscrainte, N. D., et al. Reduction in Musca domestica fecundity by dsRNA-mediated gene knockdown. PloS one. 13 (1), e0187353 (2018).
- Isoe, J., Collins, J., Badgandi, H., Day, W. A., Miesfeld, R. L. Defects in coatomer protein I (COPI) transport cause blood feeding-induced mortality in Yellow Fever mosquitoes. Proceedings of the National Academy of Sciences. 108 (24), E211-E217 (2011).
- Drake, L. L., Price, D. P., Aguirre, S. E., Hansen, I. A. RNAi-mediated gene knockdown and in vivo diuresis assay in adult female Aedes aegypti mosquitoes. Journal of Visualized Experiments: JoVE. (65), (2012).
- Kurscheid, S., et al. Evidence of a tick RNAi pathway by comparative genomics and reverse genetics screen of targets with known loss-of-function phenotypes in Drosophila. BMC Molecular Biology. 10 (1), 20 (2009).
- Kim, M., Sim, C., Denlinger, D. L. RNA interference directed against ribosomal protein S3a suggests a link between this gene and arrested ovarian development during adult diapause in Culex pipiens. Insect molecular biology. 19 (1), 27-33 (2010).
- Atyame, C. M., et al. Comparison of irradiation and Wolbachia based approaches for sterile-male strategies targeting Aedes albopictus. PLoS one. 11 (1), e0146834 (2016).
- Becnel, J. J., Floore, T. G. Introduction to"Biorational Control of Mosquitoes.". Journal of the American Mosquito Control Association. 23, 1-2 (2007).
- Bolognesi, R., et al. Characterizing the mechanism of action of double-stranded RNA activity against western corn rootworm (Diabrotica virgifera virgifera LeConte). PloS one. 7 (10), e47534 (2012).
- Jiang, Y., et al. Semiochemicals from ovaries of gravid females attract ovipositing female houseflies, Musca domestica. Journal of Insect Physiology. 48 (10), 945-950 (2002).
Reprints and Permissions
Request permission to reuse the text or figures of this JoVE article
Request PermissionThis article has been published
Video Coming Soon
ABOUT JoVE
Copyright © 2025 MyJoVE Corporation. All rights reserved