
Summary
Abstract
Introduction
Protocol
Representative Results
Discussion
Acknowledgements
Materials
References
Immunology and Infection
Analyse van somatische hypermutatie in de JH4-intron van Germinal Center B-cellen van Mouse Peyer's Patches
Hier wordt een test gepresenteerd om somatische hypermutatie binnen de immunoglobuline zware keten gen locus te kwantificeren met behulp van germinale centrum B cellen van muis Peyer's patches.
Binnen de kiemcentra van lymfoïde organen veranderen volwassen B-cellen hun uitgedrukte immunoglobuline (Ig) door niet-geploeterde mutaties in de variabele coderende exons van de Ig zware en lichte keten gen loci te introduceren. Dit proces van somatische hypermutatie (SHM) vereist het enzymactiveringsgeïnduceerde cytidinedeaminase (AID), dat deoxycytidines (C) omzet in deoxyuridines (U). Het verwerken van de aid-gegenereerde U:G mismatches in mutaties door de basis excisie en mismatch reparatie trajecten introduceert nieuwe Ig codeersequenties die een hogere affiniteit Ig kunnen produceren. Mutaties in AID- of DNA-reparatiegenen kunnen de soorten mutaties die in de Ig loci worden waargenomen, blokkeren of significant veranderen. We beschrijven een protocol om JH4-intronmutaties te kwantificeren dat gebruikmaakt van fluorescentie-geactiveerde celsortering (FACS), PCR en Sanger-sequencing. Hoewel deze test niet direct de rijping van de Ig-affiniteit meet, is deze indicatief voor mutaties in Ig-variabele coderingssequenties. Bovendien maken deze methoden gebruik van gemeenschappelijke moleculaire biologietechnieken die mutaties in Ig-sequenties van meerdere B-celklonen analyseren. Deze test is dus van onschatbare waarde in de studie van SHM- en Ig-diversificatie.
B-cellen, leden van het adaptieve immuunsysteem, herkennen en elimineren antigenen door antilichamen te produceren, ook bekend als immunoglobulinen (Ig). Elke Ig bestaat uit twee zware (IgH) en twee lichte (IgL) ketenpolypeptiden, die door disulfidebindingen bij elkaar worden gehouden om de karakteristieke "Y" vormstructuur van de Ig1te vormen. De N-termini van IgH en IgL omvatten het variabele (V) gebied van elk polypeptide en vormen samen de antigeenbindende plaats van de Ig, terwijl het constante gebied van IgH de effectorfunctie van de Ig geeft. Ontwikkeling van B-cellen in het beenmerg herschikt de V-coderende exonen van IgH en IgL in een proces dat bekend staat als V(D)J-recombinatie2,3,4. Transcriptie van de opnieuw samengevoegde V-exons, gekoppeld aan de respectieve constante regio-exonen, vormt het mRNA dat wordt vertaald in de Ig.
Rijpe B-cellen die een membraangebonden Ig uitdrukken, ook bekend als een B-celreceptor (BCR), circuleren naar secundaire lymfoïde organen, zoals de milt, lymfeklier of Peyer's patches, waar ze de omgeving onderzoeken op antigenen en interageren met andere cellen van het immuunsysteem1. Binnen de kiemcentra (GC) van secundaire lymfoïde organen worden B-cellen die antigeen herkennen via de BCR geactiveerd. Geholpen door folliculaire dendritische cellen en folliculaire helper T-cellen, kunnen geactiveerde B-cellen zich vervolgens verspreiden en differentiëren in plasma- en geheugencellen, die belangrijke effectoren zijn van een robuuste immuunrespons5,6,7,8,9. Bovendien kunnen deze geactiveerde B-cellen secundaire Ig-gendiversificatieprocessen ondergaan - klasseschakelaarrecombinatie (CSR) en somatische hypermutatie (SHM). Tijdens CSR wisselen B-cellen de standaard μ constante regio van het IgH-polypeptide uit met een ander constant gebied (γ, α, ε) door middel van een DNA-deletie-recombinatiereactie (Figuur 1). Dit maakt de uitdrukking van een andere constante exon en vertaling van een nieuwe Ig mogelijk. De B-cel schakelt over van het uitdrukken van IgM naar een ander isotype (IgG, IgA, IgE). CSR verandert de effectorfunctie van de Ig zonder de antigeenspecifiekheid10,11,12te wijzigen . Tijdens SHM muteren B-cellen echter de V-codeergebieden van IgH en IgL om de productie en selectie van Igs met een hogere affiniteit mogelijk te maken, waardoor een antigeen13,14,15 effectiever kan worden geëlimineerd ( figuur1). Belangrijk is dat zowel CSR als SHM afhankelijk zijn van de functie van één enzym: activeringsgeïnduceerd cytidinedeaminease (AID)16,17,18. Mensen en muizen met een tekort aan AID kunnen CSR of SHM niet voltooien en presenteren met verhoogde IgM serumtiters of Hyper-IgM17,19.
In MVO deaminates AID deoxycytidines (C) in de repetitieve schakelgebieden die voorafgaan aan elke constante codering exons, omzetten in deoxyuridines (U)20,21, waardoor een niet-overeenkomende basiskoppeling ontstaat tussen deoxyuridines en deoxyguanosines (U:G). Deze U:G mismatches worden omgezet in de dubbelstrengs DNA-breuken, die nodig zijn voor DNA-recombinatie, door ofwel de base excision repair (BER) of mismatch repair (MMR) pathway22,23,24,25. In SHM deaminateS AID C binnen de V-codering exons. Replicatie over de U:G mismatch genereert C:G naar T:A overgangsmutaties, terwijl verwijdering van de uracilbasis door het BER-eiwit, uracil DNA glycosylase (UNG), voorafgaand aan DNA-replicatie zowel overgangs- als transversiemutaties produceert16. Null-mutaties in UNG verhogen significant C:G naar T:A overgangsmutaties21,22. Net als MVO vereist SHM de complementaire rollen van MMR en BER. Tijdens SHM genereert MMR mutaties bij A:T basisparen. Het inactiveren van mutaties in MutS homologie 2 (MSH2) of DNA polymerase η (Polη) vermindert mutaties bij A:T-basen en samengestelde mutaties in MSH2 aanzienlijk en Polη schaft mutaties op A:T-basen21,30,31vrijwel af . In overeenstemming met de cruciale rol voor BER en MMR bij het omzetten van door AID gegenereerde U in overgangs - of transversiemutaties, geven muizen met een tekort aan zowel MSH2 als UNG (MSH2-/-UNG-/-) alleen C:G naar T:A overgangsmutaties weer die het gevolg zijn van replicatie over de U:G mismatch21.
De analyse van SHM in V-codeergebieden blijft ingewikkeld omdat het ontwikkelen van B-cellen een van de V(D)J-coderingseonen in de IgH- en IgL-loci 1,2,4kan combineren . Nauwkeurige analyse van deze uniek gehercombineerde en somatisch gemuteerde V-gebieden vereist de identificatie en isolatie van klonen van B-cellen of de Ig mRNA11,13. De JH4-intron, die 3' van de laatste J-coderende exon in de IgH-locus is, herbergt somatische mutaties als gevolg van de verspreiding van mutaties 3' van de V-promotor32,33,34 en wordt daarom vaak gebruikt als surrogaatmarker voor SHM in V-regio's31,35 ( figuur1). Om experimenteel te verduidelijken hoe specifieke genen of genetische mutaties SHM-patronen of -snelheden veranderen, kan de JH4-intron worden gesequenced uit Peyers patches (PP) germinale centrum B-cellen (GCBCs), die hoge percentages SHM36,37,38ondergaan . GCBCs kunnen gemakkelijk worden geïdentificeerd en geïsoleerd met fluorescerend geconjugeerde antilichamen tegen celoppervlakmarkers (B220+PNAHI)17,39.
Er wordt een gedetailleerd protocol gepresenteerd om JH4-intronmutaties in PP-GCBCs van muizen te karakteriseren met behulp van een combinatie van FACS (fluorescentie geactiveerde celsortering), PCR en Sanger-sequencing (figuur 2).
Alle gemuteerde muizen werden onderhouden op een C57BL/6 achtergrond. Leeftijdsgematchte (2-5 maanden oude) mannelijke en vrouwelijke muizen werden gebruikt voor alle experimenten. De veeteelt van en experimenten met muizen werden uitgevoerd volgens protocollen die zijn goedgekeurd door het City College of New York Institutional Animal Care and Use Committee.
1. Dissectie van Peyer's patches
- Euthanaseer de muis met 100% CO2 op 3 L/min gedurende 5 minuten gevolgd door cervicale dislocatie om de dood te bevestigen. Steriliseer dissectiegereedschappen (schaar, tang, fijne tang) en gehandschoende handen met 70% ethanol.
- Leg de muis op het dissectiekussen met de buik bloot. Spuit het lichaam van de muis royaal met 70% ethanol voordat u incisies maakt om het dissectiegebied te steriliseren.
- Maak een incisie in de huid over de buik en verwijder de huid uit de buik door tegelijkertijd aan beide zijden van de incisie naar het hoofd en de staart te trekken met behulp van een tang (of gesteriliseerde, gehandschoende handen).
- Speld de voor- en achterpoten van de muis vast.
- Snijd de buikholte met een schaar om de inwendige organen bloot te leggen.
- Zoek de dunne darm tussen de maag en het caecum ("J" vormige structuur in de buurt van de dikke darm). Verwijder de dunne darm door onder de maag en boven het caecum te snijden.
- Verwijder bindweefsel en vet dat de plooien van de dunne darm met elkaar verbindt.
OPMERKING: Vet heeft een kenmerkende witte kleur, in tegenstelling tot de roze kleur van de dunne darm. - Onderzoek het externe oppervlak van de dunne darm op de vlekken van de Peyer (PPs), die kleine (~ 1 mm) ovaalvormige structuren zijn die wit lijken onder een dunne laag doorschijnende epitheelcellen.
- Verwijder zorgvuldig alle zichtbare PP met een schaar.
OPMERKING: Een C57BL/6 wild-type (WT) muis kan 4-8 PPs opleveren, terwijl een AID-/- muis 6-10 PPs heeft. - Verzamel de PPs in een microcentrifugebuis van 1,5 ml met 1 ml FACS-buffer op ijs.
OPMERKING: De PP moet zinken, terwijl vet naar de oppervlakte drijft en kan worden verwijderd.
2. Celisolatie voor FACS
- Plaats een filter van 40 μm in een 6-puts schaal met 1 ml koude (4 °C) FACS buffer.
- Giet de PPs van de 1,5 ml buis op het filter.
- Was PPs met 1 ml koude FACS-buffer en zorg ervoor dat ze altijd in vloeistof en op ijs zitten.
- Gebruik het platte uiteinde van de zuiger van een spuit van 1 ml als stamper om de PPs op het filter te verpletteren totdat er alleen bindweefsel op het filter achter blijft.
- Was het filter en de zuiger met 1 ml koude FACS-buffer om de cellen in de 6-putschaal vrij te maken.
- Verzamel de ~4 ml cellen in koude FACS-buffer en filtreer ze door een 40 μm zeefdop FACS-buis.
- Was de zeefdop met 1 ml koude FACS-buffer.
- Pellet de cellen bij 600 x g bij 4 °C gedurende 5 minuten in een swingende emmercentrifuge.
- Decanteer de supernatant.
- Resuspendeer de cellen in 0,4 ml koude FACS-buffer.
- Verwijder 10 μL voor het tellen van cellen om de opbrengst te verifiëren (verwacht ~5 x 106 cellen/muis, zie figuur 3A)
- Filtreer de resterende cellen door een zeefdop van 40 μm in een FACS-buis en ga verder met kleuring voor FACS.
3. Kleuring GCBCs voor FACS
- Voeg 1 μL Fc blok (ongelabelde antimuis CD16/CD32) toe aan de 400 μL celsuspensie en zet de cellen 15 min op ijs.
- Voeg 2 ml koude FACS-buffer toe om de cellen te wassen.
- Pelletcellen bij 600 x g bij 4 °C gedurende 5 min en gooi het supernatant weg.
- Resuspend de cellen in 80 μL koude FACS buffer.
- Verwijder 10 μL cellen uit de WT PP voor elke kleuringsregelaar (4 in totaal, inclusief 3 enkele vlekregelaars en 1 onbesmet besturingselement). Laat 40 μL van de WT PP over voor de volgende stap. U kunt ook compensatieparels gebruiken voor de kleuringsregelaars.
- Beits elk van de experimentele monsters (bv. WT en AID-/-) in 500 μL koude FACS-buffer met 2,5 μL pindaagglutinine (PNA)-biotine gedurende 15 minuten op ijs.
- Voeg 2 ml koude FACS-buffer toe om de cellen te wassen.
- Pelletcellen bij 600 x g bij 4 °C gedurende 5 min en gooi het supernatant weg.
- Bevlek elk experimenteel monster met 500 μL cocktail in het donker, op ijs, gedurende 15 minuten (tabel 1). Zorg ervoor dat de cellen volledig zijn geresuspendeerd in de kleuringcocktail.
- Bereid enkele vlekbesturingselementen voor de compensatiematrix voor.
- Beits de cellen in 500 μL koude FACS-buffer met behulp van de in tabel 2gespecificeerde verdunningen .
- Incubeer de kleuringscontroles in het donker, op ijs, gedurende 15 minuten.
- Voeg 2 ml koude FACS-buffer toe aan alle buizen in stap 3.7 en 3.8, pellet de cellen en gooi het supernatant weg om ongebonden antilichamen of DAPI af te wassen.
- Resuspend de cellen in 500 μL koude FACS buffer en plaats op ijs.
- Verzamel B220+PNAHI met behulp van een celsorteerder uit elk gekleurd experimenteel monster. Figuur 3B toont de typische percentages B220+ PNAHI verkregen uit WT en AID-/- PPs. Figuur 3C toont de FACS gating strategie.
4. DNA-extractie uit GCBCs
- Pellet gesorteerde cellen bij 600 x g bij 4 °C gedurende 5 min en gooi het supernatant weg.
- Resuspendeer de cellen in 1 ml koude FACS-buffer en breng de cellen over in een microcentrifugebuis van 1,5 ml.
- Pellet de cellen bij 600 x g bij 4 °C gedurende 5 min en gooi het supernatant weg.
- Resuspendeer de cellen in 500 μL DNA-extractiebuffer en 5 μL van 20 mg/ml Proteïnase K.
- Incubeer 's nachts bij 56 °C.
- Precipitaat DNA met 500 μL isopropanol en 1 μL 20 mg/ml glycogeen. Meng de buis grondig door 5-6x om te keren.
- Incubeer bij kamertemperatuur gedurende 10 minuten.
- Centrifugeer gedurende 15 min in een microcentrifuge bij 25 °C bij 21.000 x g.
- Gooi het supernatant weg en behoud de pellet, die het neergeslagen DNA en glycogeen bevat.
- Was de DNA-pellet met 1 ml 70% ethanol.
- Pellet het DNA in een microcentrifuge gedurende 10 min bij 25 °C bij 21.000 x g.
- Verwijder de 70% ethanol en droog de DNA-pellet 5-10 minuten aan de lucht.
OPMERKING: Vermijd overdrogen omdat het DNA mogelijk niet volledig rehydrateert.
- Resuspend het DNA in 30 μL TE buffer en incubeer 's nachts bij 56 °C.
5. JH4 intron sequentieversterking en analyse
- Kwantificeer DNA door de absorptie te meten op een golflengte van 260 nm (A260).
OPMERKING: De typische concentratie DNA die wordt teruggevonden uit gesorteerde B220+PNAHI GCBCs van een C57BL/6 muis is 20-40 ng/μL. - Voer de geneste PCR uit voor de JH4-intron (tabel 3, tabel 4). Normaliseer de totale hoeveelheid genomisch DNA die in de eerste PCR wordt gebruikt tot het minst geconcentreerde monster. (als het minst geconcentreerde monster bijvoorbeeld 5 ng/μL is, gebruikt u 58,75 ng DNA voor alle monsters in het maximale volume water (11,75 μL) in PCR-#1).
- Los het PCR-product gedurende 20 minuten op op een 1,5% agarosegel op 200 V. De verwachte amplicongrootte is 580 bp.
- Verwijder het amplicon uit de gel en extraheer het DNA met behulp van een gelextractiekit volgens de instructies van de fabrikant (zie aanvullende figuur 1).
- Ontleed het DNA met 30 μL water en kwantificeer de hoeveelheid DNA door de A260 te meten.
OPMERKING: De typische concentratie van het gezuiverde PCR-product is 3-10 ng/μL.
- Ontleed het DNA met 30 μL water en kwantificeer de hoeveelheid DNA door de A260 te meten.
- Bind het gezuiverde PCR-product in een plasmide met stompe uiteinden. Standaardiseer de totale hoeveelheid PCR-product die bij elke ligatiereactie wordt gebruikt (Tabel 5).
- Incubeer de ligatiereactie bij kamertemperatuur gedurende 5 minuten of 's nachts bij 16 °C.
- Transformeer elektrocompetente bacteriële cellen met 2 μL van de ligatiereactie.
- Elektroporaat bij 1,65 kV.
- Redding in 600 μL SOC-media gedurende 1 uur bij 37 °C in een schudincubator bij 225 tpm.
- Plaat 100 μL getransformeerde bacteriën op LB aangevuld met ampicilline (100 μg/ml) agarplaten en 's nachts incuberen bij 37 °C.
- Dien de plaat met bacteriële kolonies in voor Sanger-sequencing met behulp van de T7 forward primer. U kunt ook 's nachts culturen van elke bacteriële kolonie kweken en een plasmidezuivering uitvoeren.
- Herhaal indien nodig de PCR, ligatie en/of transformatie om de opbrengst van bacteriële kolonies te optimaliseren
OPMERKING: Van elke plaat moeten minimaal 30 kolonies worden geplukt.
- Herhaal indien nodig de PCR, ligatie en/of transformatie om de opbrengst van bacteriële kolonies te optimaliseren
- De reeksgegevens in de .txt-bestanden standaardiseren
- Verwijder de plasmidevolgorde.
- Zorg ervoor dat elke sequentie 5' tot 3' is georiënteerd volgens de JH4 intron referentiesequentie (NG_005838). Genereer indien nodig de omgekeerde aanvulling van elke reeks.
- Lijn de voor elke PCR verkregen sequenties uit tegen de JH4 intron referentiesequentie (NG_005838) met behulp van een Clustal Omega software (Figuur 4A).
- Verschillen met de referentiesequentie identificeren als mutaties
- Controleer of alle mutaties echte puntmutaties zijn door het elektroferogram van de Sanger-sequencing te onderzoeken. Herhaal indien nodig de volgorde. (figuur 4B,C).
- Unieke mutaties in de JH4-intron voor elk genotype tabuleren en kwantificeren (figuur 5).
- Tel sequenties met identieke mutaties slechts één keer
OPMERKING: Het is niet mogelijk om te bepalen of de identieke sequenties zijn gegenereerd tijdens de PCR of identieke SHM-gebeurtenissen in verschillende B-cellen. - Tel elk geval van WT-kiembaan JH4-intronsequenties (d.w.z. die zonder mutaties) als een unieke sequentie.
- Tel sequenties met identieke mutaties slechts één keer
Flow cytometrie
Volwassen B-cellen circuleren naar kiemcentra waar ze affiniteitsrijping, klonale expansie en differentiatie in plasma- of geheugencellen40,41,42,43,44ondergaan. Deze GCBCs zijn te herkennen aan talrijke celoppervlakmarkers, waaronder een hoge expressie van de CD45R/B220-receptor en binding van pindaagglutinine (PNA)45,46. Om geactiveerde GCBCs te isoleren, werden PP-cellen gekleurd met anti-B220-antilichamen geconjugeerd aan fycoerythrine (PE) en biotinylated-PNA, gevolgd door streptavidine geconjugeerd tot APC-eFluor780. Dode cellen werden geëlimineerd met behulp van de fluorescerende 4',6-Diamidino-2-Phenylindole (DAPI) kleurstof, die het nucleïnezuur van stervende of dode cellenbevlekt 47,48. De bevlekte cellen werden vervolgens geanalyseerd en gesorteerd via flowcytometrie. De PPs bestonden uit ~80% B220+ cellen49,50. WT PPs bevatten gemiddeld 4 x 106 cellen per muis (figuur 3A). Ongeveer 8% van de WT PP-cellen was B220+PNAHI, wat de helft is van het aantal waargenomen in AID-/ - ( Figuur3B). Zo werden 0,3-0,6 x 106 B220+PNAHI GCBCs verkregen na sortering, die voldoende waren om mutaties in de JH4 intron te analyseren.
JH4-sequentieanalyse
De JH4 intron werd versterkt door een geneste PCR met behulp van gangbare VHJ558 familieprimers (J558FR3Fw en VHJ558.2) gevolgd door JH4 intron over primers VHJ558.3 en VHJ558.435,37. Van de 105 unieke sequenties verkregen uit WT GCBCs werden in totaal 226 mutaties gevonden (Figuur 5A). Analyse van het GCBC-mutatiespectrum bij de WT-muizen toonde een reeks overgangen en transversies met een snelheid van 4 x 10-3 mutaties/bp, die werd berekend door het totale aantal gemuteerde basen te delen door het totale aantal basen dat32 , 36,37,38was gesequenced . Bovendien bevatte elk JH4 PCR-product van WT GCBCs 1-25 mutaties (figuur 5B), waarbij vaak meerdere mutaties werden aangetroffen op één sequentie33,36. Slechts twee mutaties werden geïdentificeerd in 113 AID-/- sequenties (Figuur 5A). AID-/- B-cellen vertoonden 1,66 x 10-5 mutaties/bp, wat significant lager was dan WT B-cellen (p <0,05)36 en is vergelijkbaar met het foutenpercentage van de high fidelity polymerase (5,3 x 10-7 sub/base/verdubbeling)51,52. Aid -/- B-cellen dienden dus als een nuttige negatieve controle voor deze test.
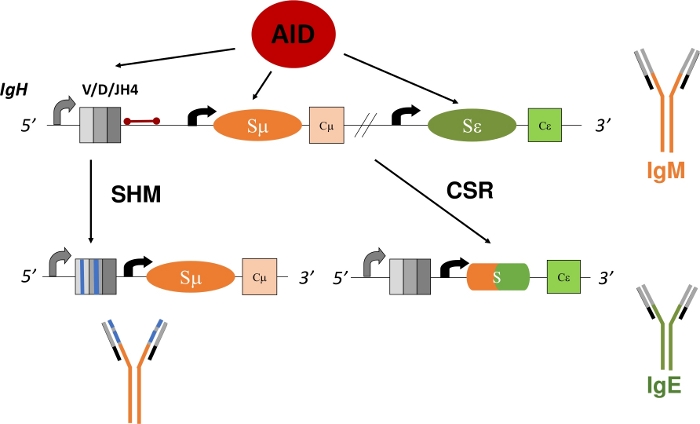
Figuur 1: Schematische weergave van de IgH-gen locus en de regio's waarop AID tijdens MVO en SHM is gericht. De rode balk geeft de JH4-intron van 580 bp aan die 3' VDJH4-herschikkingen is en wordt geanalyseerd in dit protocol. In MVO bevordert AID-afhankelijke deaminatie van intronische schakelgebieden (Sμ en Sε) DSB-vorming die deletie-recombinatie en de expressie van een nieuw antilichaamisotype (IgM tot IgE) mogelijk maakt. Tijdens SHM accumuleren V-regio's (grijze vakken) mutaties (blauwe lijnen) die kunnen leiden tot een hogere affiniteit Ig. Klik hier om een grotere versie van deze afbeelding te bekijken.
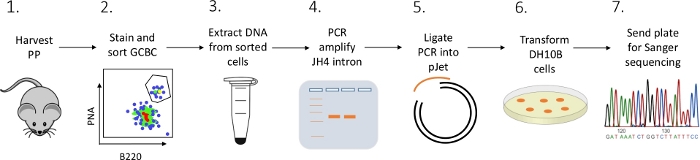
Figuur 2: Workflow om SHM van de JH4 intron te analyseren in GCBCs geïsoleerd van PPs. Klik hier om een grotere versie van deze afbeelding te bekijken.
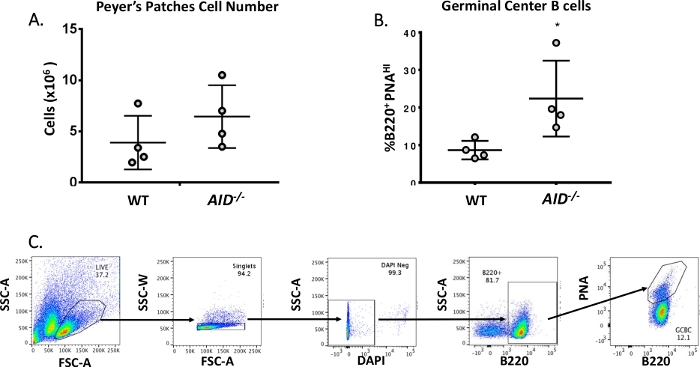
Figuur 3: Karakterisering van PP GCBCs. (A) Totaal aantal PP-cellen van WT- enAID-/- muizen (n = 4 per genotype). Foutbalken vertegenwoordigen standaardafwijking van het gemiddelde. (B) Percentage B220+PNAHI GCBCs verkregen uit PPs van WT en AID-/- muizen (n = 4 per genotype)36. Foutbalken vertegenwoordigen standaardafwijking van het gemiddelde, *p<0.05 met behulp van de t-toets van de student. (C) Representatieve FACS-percelen om B220+PNAHI GCBCs van PC's te sorteren. Klik hier om een grotere versie van deze afbeelding te bekijken.
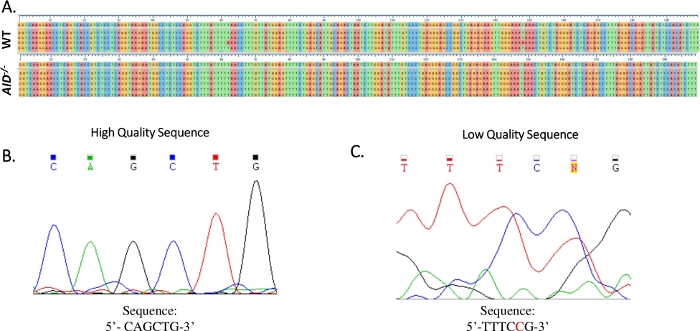
Figuur 4: Analyse van JH4 Sanger-sequentiegegevens. (A) Voorbeeldsequentie-uitlijningen van Sanger-sequentiegegevens van het JH4 PCR-product van WT (boven) en AID-/- (onder) GCBCs naar de referentie-genomische sequentie (NG_005838), de volgorde direct onder de genummerde vinkjes. Uitlijningen werden gegenereerd met Clustal Omega. (B) Elektroferogram van hoogwaardige Sanger-sequentiegegevens, die voor elke basis verschillende pieken vertoonden. (C) Elektroferogram van sequentiegegevens van lage kwaliteit, die dubbelzinnige pieken en niet-gespecificeerde basen (N) vertoonden. De nucleotide die in rood wordt weergegeven, moet handmatig worden geannoteerd in het sequentietekstbestand. Klik hier om een grotere versie van deze afbeelding te bekijken.
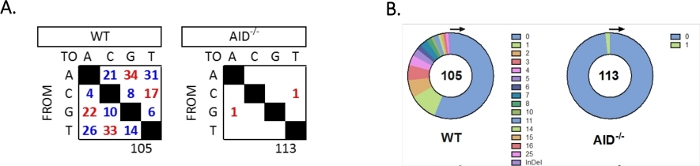
Figuur 5: Analyse van mutaties in het JH4-intron in WT en AID-/- GCBCs. (A) Het totale aantal overgangsmutaties (rood) en transversiemutaties (blauw) bij A-, C-, G- en T-basen voor elk genotype wordt samengevat in de tabellen. Het totale aantal geanalyseerde sequenties wordt onder de tabel aangegeven. (B) Het aantal mutaties per PCR-amplicon voor elk genotype wordt weergegeven in de cirkeldiagrammen. Dit cijfer is gewijzigd van Choi et al.36 Copyright 2020. De American Association of Immunologists, Inc. Klik hier om een grotere versie van dit cijfer te bekijken.
| KleuringCocktail voor GCBCs | Volume: 500 μL | ||
| Antilichaam of kleurstof | Fluorofoor | Verdunning | µl |
| B220 | Pe | 1000 | 0.5 |
| Streptavidin | APC-eFluor780 | 500 | 1 |
| DAPI | N/a | 500 | 1 |
Tabel 1: Kleuringcocktails voor GCBCs. Cocktail van de geïndiceerde antilichamen of kleurstof (cursief aangegeven) bij de gespecificeerde verdunningen werden gebruikt om PP-cellen in 500 μL te bevlekken voor flowcytometrie.
| Enkele vlekken voor compensatie | Volume: 500 μL | ||
| Antilichaam of kleurstof | Fluorofoor | Verdunning | µl |
| B220 | Pe | 1000 | 0.5 |
| B220 | APC-eFluor780 | 750 | 0.67 |
| DAPI | N/a | 500 | 1 |
Tabel 2: Enkele vlekregelaars voor compensatie. B220-antilichamen die aan de aangegeven fluoroforen waren geconjugeerd, werden gebruikt voor controles met één vlek om spectrale overlapping te compenseren.
| PCR-#1 | ||||
| Reagens | Volume | Thermocycler Voorwaarden | ||
| 5x buffer | 4 μL | 1 | 95 °C | 3 min |
| 10 mM dNTP | 2 μL | 2 | 94 °C | 30 seconden |
| 10 μM J558FR3Fw | 1 μL | 3 | 55 °C | 30 seconden |
| 10 μM VHJ558,2 | 1 μL | 4 | 72 °C | 1:30 min |
| High Fidelity DNA polymerase | 0,25 μL | Cyclus 2-4 9x | ||
| Dna | x (standaardiseren tot minst geconcentreerd monster) | |||
| H2O | tot 20 μL | 5 | 72 °C | 5 min |
| Verdun PCR-product 1:5 in H2O voordat u doorgaat naar PCR-#2 | ||||
Tabel 3: Geneste PCR van de JH4 intron. PCR-componenten en thermocycleromstandigheden voor de eerste versterkingsreactie. Verdun het eerste PCR-product 1:5 met water en gebruik 1 μL van deze verdunning voor de tweede PCR.
| PCR-#2 | ||||
| Reagens | Volume | Thermocycler Voorwaarden #2 | ||
| 5x buffer | 4 μL | 1 | 94 °C | 3 min |
| 10 mM dNTP | 2 μL | 2 | 94 °C | 30 seconden |
| 10 μM VHJ558,3 | 1 μL | 3 | 55 °C | 30 seconden |
| 10 μM VHJ558,4 | 1 μL | 4 | 72 °C | 30 seconden |
| High Fidelity DNA polymerase | 0,25 μL | Cyclus 2-4 21x | ||
| Verdunde PCR#1 | 1 μL | |||
| H2O | tot 20 μL | 5 | 72 °C | 5 min |
Tabel 4: PCR-componenten en thermocycleromstandigheden voor de tweede PCR.
| Reagens | Volume |
| 2x buffer | 10 μL |
| Gezuiverde PCR | x (standaardiseren tot minst geconcentreerd monster) |
| Plasmide met stompe uiteinden | 1 μL |
| T4 DNA Ligase | 1 μL |
| H2O | tot 20 μL |
| Incubeer bij kamertemperatuur gedurende 5 minuten of 's nachts bij 16ºC | |
Tabel 5: Ligatiereactie. Componenten voor de ligatie van het gezuiverde JH4 intron PCR-product in de plasmide.
| FACS-buffer |
| Verwarm FBS gedurende een uur voor gebruik bij 56 °C. Supplement PBS, pH 7,4 (Gibco, #10010049) met 2,5% (v/v) van warmte-geactiveerde FBS. Bewaren bij 4°C. |
| DNA-extractiebuffer (100 mM Tris pH 8,0, 0,1 M EDTA, 0,5% (m/v) SDS) |
| Voeg 50 ml 1 M Tris pH 8,0, 100 ml 0,5 M EDTA en 12,5 ml 20% SDS toe. Voeg gedestilleerd water toe aan 500 ml. Bewaren bij kamertemperatuur. |
| TE Buffer (10 mM Tris pH 8,0, 1 mM EDTA) |
| Voeg 2,5 ml 1 M Tris pH 8,0 en 500 ml 0,5 M EDTA toe. Voeg gedestilleerd water toe aan 250 ml. Bewaren bij kamertemperatuur. |
Tabel 6: Bufferrecepten.
| Oligonucleotiden Lijst | ||
| J558FR3Fw | 5'-GCCTGACATCTGAGGACTCTGC-3' | |
| VHJ558.2 | 5'-CTGGACTTTCGGTTTGGTG-3' | |
| VHJ558.3 | 5'-GGTCAAGGAACCTCAGTCA-3' | |
| VHJ558.4 | 5'-TCTCTAGACAGCAACTAC-3' | |
Tabel 7: Oligonucleotiden gebruikt bij de test.
Aanvullende figuur 1: Representatieve agarose gel afbeelding na voltooiing van stap 5.4. Het JH4 intron geneste PCR-product werd opgelost op een 1,5% agarosegel en de amplicon van 580 bp werd verwijderd. WT PP geeft aan dat WT PP GCBC genomisch DNA werd gebruikt als sjabloon voor de eerste PCR en AID PP geeft aan dat AID-/- PP GCBC genomisch DNA werd gebruikt als sjabloon voor de eerste PCR. ɸ geeft de geen sjabloon PCR-besturingselement aan en - geeft aan dat er niets in de put van de agarose-gel is geladen. De laatste baan toont een 100 bp DNA ladder. Klik hier om deze afbeelding te downloaden.
Het karakteriseren van SHM binnen de IgH- en IgL V-coderingssequenties van een heterogene B-celpopulatie vormt een uitdaging, aangezien elke B-cel op unieke wijze V-coderingssegmenten reorganiseert tijdens V(D)J-recombinatie34. In dit artikel beschrijven we een methode om mutaties in het JH4-intron van GCBCs te identificeren. De JH4-intron, die zich 3' van het laatste J-coderingssegment in de IgH-locus bevindt, wordt gebruikt als surrogaat voor SHM van V-regio 's (Figuur 1)31,33,34,35. Om deze JH4 intron mutaties te catalogiseren en te beoordelen hoe specifieke genen de productie of het patroon van mutaties beïnvloeden, worden PP GCBCs specifiek geanalyseerd. Deze cellen accumuleren JH4 intron mutaties als gevolg van chronische stimulatie door intestinale microbiota53. Bovendien hebben de B220+PNAHI GCBCs van de PPs van niet-geïmmuniseerde muizen een mutatiespectra die zich verhoudt tot milt GCBCs van geïmmuniseerde dieren54,55. Mutaties in het JH4-intron kunnen echter niet worden gecorreleerd met Ig-affiniteitsrijping omdat deze mutaties niet-coderend zijn.
Om te bepalen of SHM de Ig-affiniteit verandert, moeten muizen intraperitoneaal worden geïmmuniseerd met een antigeen, zoals NP (4-hydroxy-3-nitrofenylacetyl) geconjugeerd aan CGG (kippen gammaglobuline) of KLH (sleutelgatlimpet hemocyanine)56. Vervolgens kan mRNA worden gezuiverd uit milt B220+PNAHI GCBCs om SHM te onderzoeken binnen VH186.2, de V-codering exon die het vaakst NP herkent en wordt gemuteerd na NP-CGG of NP-KLH immunisatie31,57,58,59,60. Mutatie van tryptofaan-33 naar een leucine in VH186.2 is gekarakteriseerd om de Ig-affiniteit te verhogen tot 10-voudig59,60 en is daarom een indicator dat SHM en klonale selectie een hoge affiniteit Ig hebben gegenereerd. Het meten van NP7- en NP20-specifieke serum-titers door ELISA en het berekenen van de Ig-specifieke NP7/NP20-ratio tijdens de immunisatie documenteert ook de Ig-affiniteitsrijping als gevolg van SHM van V-regio 's17,21,36. Beide assays kunnen worden gebruikt om SHM binnen de VH186.2 coderingssequenties te correleren met veranderingen in NP-specifieke Ig affiniteitsrijping.
Of geïmmuniseerde of niet-geïmmuniseerde dieren worden gebruikt om SHM van VH186.2 of de JH4-intron te analyseren, GCBCs moeten nauwkeurig worden geïdentificeerd. Wepresenteren een FACS-gebaseerde benadering om B220+PNAHI GCBCs te isoleren. Als alternatief kunnen Fas en niet-gesulfateerd α2-6-sialyl-LacNAc-antigeen, dat wordt herkend door het GL7-antilichaam61,62 , 63,64, ook worden gebruikt om GCBCs te isoleren, die worden geïdentificeerd als B220+Fas+GL7+65 of CD19+Fas . GL7-expressie weerspiegelt PNA nauw in geactiveerde GCBCs van de lymfeklieren64,65,66. Naast het gebruik van antilichaammarkers die specifiek zijn voor GCBCs, moeten kleurcocktails de excitatie van een fluorofoor en detectie van een biomarker maximaliseren en tegelijkertijd spectrale overlapping van fluorescentie-emissie minimaliseren. Antigenen die bij lage niveaus worden uitgedrukt , moeten worden gedetecteerd met een antilichaam dat is geconjugeerd tot een fluorofoor met een robuuste emissiefluorescentie67. Het aanbevolen kleuringsprotocol werd geoptimaliseerd voor analyse op een celsorteerder uitgerust met vier lasers (405nm, 488nm, 561nm, 633nm) en 12 filters; filterconfiguraties en laserbeschikbaarheid variëren echter tussen cytometers. Om het protocol te wijzigen op basis van de beschikbaarheid van reagentia en apparatuur, wordt de lezer verwezen naar aanvullende bronnen, online spectrumkijkers en gepubliceerde literatuur67,68,69,70,71,72,73. Het hierin beschreven multi-color kleuringsprotocol vereist compensatie van spectrale overlapping om ervoor te zorgen dat de gesorteerde celpopulaties GCBCs zijn in plaats van onnauwkeurige detectie van fluorescentie-emissie. B220 dient als een nuttige kleuringscontrole voor het beschreven FACS (tabel 1B) omdat PPs onderscheidende B220-negatieve en positieve populaties zullen hebben (figuur 3C), wat een passende compensatie van spectrale overlapping mogelijk maakt. De gatingstrategie in figuur 3C moet als richtsnoer worden gebruikt. De flow cytometrie plots kunnen variëren afhankelijk van de kleuringsomstandigheden en cytometer instellingen. Niettemin moet 4-10% van de levende cellen B220+PNAHI 35, 52 zijn.
Alle mutaties in het JH4-intron van PP GCBCs moeten worden gevalideerd om ervoor te zorgen dat de waargenomen mutaties echt een weerspiegeling zijn van SHM en geen artefact van PCR of sequencing. AID-/- B-cellen kunnen dienen als een nuttige negatieve controle bij het onderzoeken van het SHM-fenotype in andere gemuteerde muismodellen , omdat deze cellen SHM 17,19niet kunnen voltooien . De JH4 intron mutatiesnelheid in de van AID-/- GCBCs (1,66x10-5 mutaties/bp)20,21,36,37,38,50,74 is vergelijkbaar met het foutenpercentage van de high fidelity polymerase (5,3x10-7 sub/base/verdubbeling)51,52 die wordt gebruikt om het DNA te versterken. Als AID-/- muizen niet beschikbaar zijn, vergelijk dan het waargenomen mutatiepatroon en de frequentie met de gepubliceerde literatuur. Ig V-regio's accumuleren 10-3-10 -4 mutaties per basispaardeling, wat ongeveer 106keer hoger is dan de mutatiesnelheid van andere gen loci73,75. De resultaten kunnen variëren met de leeftijd van het dier76. Als alternatief kunnen B220+PNALO-cellen, die niet-GCBCs markeren, worden gebruikt als een negatieve controle bij afwezigheid van AID-/- muizen52. Als de mutatiefrequentie in WT GCBCs lager is dan verwacht, kan de WT-kiembaan JH4-intronische sequentie onevenredig worden weergegeven. Zorg er in dit geval voor dat GCBCs op de juiste manier zijn gekleurd en gesorteerd en dat PCR's vrij zijn van WT-kiembaan JH4-intronbesmetting. Bovendien moeten ruwe sequencinggegevens in elektroferogrammen grondig worden geanalyseerd om ervoor te zorgen dat mutaties in de sequentietekstgegevens geen artefacten van sequencingfouten zijn. Slechte Sanger-sequencingresultaten kunnen bijvoorbeeld de betrouwbaarheid van de sequentiegegevens verminderen (figuur 4). Deze kwaliteitscontrole van de Sanger-sequentiegegevens verhoogt de nauwkeurigheid en reproduceerbaarheid van de JH4-intronmutatieanalyse.
De auteurs hebben niets bekend te maken.
We danken Tasuku Honjo voor de AID-/- muizen. Dit werk werd ondersteund door The National Institute on Minority Health and Health Disparities (5G12MD007603), The National Cancer Institute (2U54CA132378) en The National Institute of General Medical Sciences (1SC1GM132035-01).
| Name | Company | Catalog Number | Comments |
| 0.2 ml PCR 8-tube FLEX-FREE strip, attached clear flat caps, mixed | USA Scientific | 1402-4708 | |
| Ampicillin sodium salt | Fisher | BP1760-5 | |
| APC-eFluor780 anti-CD45R/B220 | eBioscience | 47-0452-80 | clone RA3-6B2 |
| BD FACSAria II | BD | 643186 | four lasers (405nm, 488nm, 561nm, 633nm) and 12 filters (PacBlue (450/50), AmCyan (502LP; 530/30), SSC (488/10), FITC (502LP; 530/30), PerCP-Cy5.5 (655LP; 695/40), PE (585/15), PE-Texas Red (600LP; 610/20), PE-Cy5 (630LP; 670/14), PE-Cy7 (735LP; 780/60), APC (660/20), Alexa700 (710LP; 730/45), APC-Cy7 (755LP; 780/60)) |
| BD slip tip 1mL syringe | Fisher | 14-823-434 | sterile |
| Biotinylated peanut agglutinin (PNA) | Vector Labs | B-1075-5 | |
| C57BL/6J mice | Jackson Laboratories | 664 | |
| Corning Falcon test tube with cell strainer snap cap | Fisher | 08-771-23 | |
| DAPI (4',6-Diamidino-2-Phenylindole, dihydrochloride) | Fisher | D1306 | 0.5 mg/ml |
| dNTP | NEB | N0447L | 10 mM |
| ElectroMAX DH10B competent cells | Fisher | 18-290-015 | |
| Falcon cell strainer 40mm | Fisher | 08-771-1 | |
| Falcon round-bottom polystyrene tubes (FACS tubes) | Fisher | 14-959-5 | |
| Falcon round-bottom polystyrene tubes (capped) | Fisher | 149591A | |
| Fetal bovine serum | R&D Systems (Atlanta Biologicals) | S11150 | |
| Gibco phosphate buffered saline PBS pH 7.4 | Fisher | 10-010-049 | |
| Glycogen | Sigma | 10901393001 | |
| Lasergene Molecular Biology (MegAlign Pro) | DNA Star | version 15 | |
| PE anti-CD45R/B220 | BD | 553090 | clone RA3-6B2 |
| Proteinase K | Fisher | BP1700-100 | |
| Q5 High-Fidelity DNA Polymerase | NEB | M0491L | |
| QIAquick Gel Extraction Kit | Qiagen | 28706 | |
| Seal-Rite 1.5mL microcentrifuge tubes | USA Scientific | 1615-5500 | |
| Streptavidin APC-eFluor 780 Conjugate | eBioscience | 47-4317-82 | |
| T4 DNA ligase | NEB | M020L | |
| Thermo Scientific CloneJET PCR Cloning Kit | ThermoFisher | FERK1231 | |
| Tissue culture plate 6 well | Fisher | 08-772-1B | sterile |
| Unlabeled anti-mouse CD16/CD32 (Fc block), BD | Fisher | BDB553142 | Clone 2.4G2 |
- Murphy, K., Weaver, C. . Janeyway's Immunobiology. , (2016).
- Alt, F. W., et al. VDJ recombination. Immunology Today. 13 (8), 306-314 (1992).
- Schatz, D. G., Ji, Y. Recombination centres and the orchestration of V (D) J recombination. Nature Reviews Immunology. 11 (4), 251-263 (2011).
- Oettinger, M. A., Schatz, D. G., Gorka, C., Baltimore, D. RAG-1 and RAG-2, adjacent genes that synergistically activate V (D) J recombination. Science. 248 (4962), 1517-1523 (1990).
- Berek, C., Berger, A., Apel, M. Maturation of the immune response in germinal centers. Cell. 67 (6), 1121-1129 (1991).
- Linterman, M. A., et al. Foxp3+ follicular regulatory T cells control the germinal center response. Nature Medicine. 17 (8), 975 (2011).
- Shulman, Z., et al. T follicular helper cell dynamics in germinal centers. Science. 341 (6146), 673-677 (2013).
- Good-Jacobson, K. L., et al. PD-1 regulates germinal center B cell survival and the formation and affinity of long-lived plasma cells. Nature Immunology. 11 (6), 535 (2010).
- Kerfoot, S. M., et al. Germinal center B cell and T follicular helper cell development initiates in the interfollicular zone. Immunity. 34 (6), 947-960 (2011).
- Chaudhuri, J., Alt, F. W. Class-switch recombination: interplay of transcription, DNA deamination and DNA repair. Nature Reviews Immunology. 4 (7), 541-552 (2004).
- Alt, F. W., Zhang, Y., Meng, F. L., Guo, C., Schwer, B. Mechanisms of programmed DNA lesions and genomic instability in the immune system. Cell. 152 (3), 417-429 (2013).
- Xu, Z., Zan, H., Pone, E. J., Mai, T., Casali, P. Immunoglobulin class-switch DNA recombination: induction, targeting and beyond. Nature Reviews Immunology. 12 (7), 517-531 (2012).
- Di Noia, J. M., Neuberger, M. S. Molecular mechanisms of antibody somatic hypermutation. Annual Reviews of Biochemistry. 76, 1-22 (2007).
- Peled, J. U., et al. The biochemistry of somatic hypermutation. Annual Review of Immunology. 26, 481-511 (2008).
- Liu, M., Schatz, D. G. Balancing AID and DNA repair during somatic hypermutation. Trends in Immunology. 30 (4), 173-181 (2009).
- Methot, S., Di Noia, J. Molecular Mechanisms of Somatic Hypermutation and Class Switch Recombination. Advances in Immunology. 133, 37-87 (2017).
- Muramatsu, M., et al. Class switch recombination and hypermutation require activation-induced cytidine deaminase (AID), a potential RNA editing enzyme. Cell. 102 (5), 553-563 (2000).
- Petersen-Mahrt, S. K., Harris, R. S., Neuberger, M. S. AID mutates E. coli suggesting a DNA deamination mechanism for antibody diversification. Nature. 418 (6893), 99 (2002).
- Revy, P., et al. Activation-induced cytidine deaminase (AID) deficiency causes the autosomal recessive form of the Hyper-IgM syndrome (HIGM2). Cell. 102 (5), 565-575 (2000).
- Petersen-Mahrt, S. DNA deamination in immunity. Immunological Reviews. 203 (1), 80-97 (2005).
- Rada, C., Di Noia, J. M., Neuberger, M. S. Mismatch recognition and uracil excision provide complementary paths to both Ig switching and the A/T-focused phase of somatic mutation. Molecular Cell. 16 (2), 163-171 (2004).
- Rada, C., et al. Immunoglobulin isotype switching is inhibited and somatic hypermutation perturbed in UNG-deficient mice. Current Biology. 12 (20), 1748-1755 (2002).
- Schrader, C. E., Vardo, J., Stavnezer, J. Role for mismatch repair proteins Msh2, Mlh1, and Pms2 in immunoglobulin class switching shown by sequence analysis of recombination junctions. The Journal of Experimental Medicine. 195 (3), 367-373 (2002).
- Martin, A., et al. Msh2 ATPase activity is essential for somatic hypermutation at AT basepairs and for efficient class switch recombination. The Journal of Experimental Medicine. 198 (8), 1171-1178 (2003).
- Imai, K., et al. Human uracil-DNA glycosylase deficiency associated with profoundly impaired immunoglobulin class-switch recombination. Nature Immunology. 4 (10), 1023-1028 (2003).
- Masani, S., Han, L., Yu, K. Apurinic/apyrimidinic endonuclease 1 is the essential nuclease during immunoglobulin class switch recombination. Molecular and Cellular Biology. 33 (7), 1468-1473 (2013).
- Guikema, J. E., et al. APE1-and APE2-dependent DNA breaks in immunoglobulin class switch recombination. The Journal of Experimental Medicine. 204 (12), 3017-3026 (2007).
- Schrader, C. E., Guikema, J. E., Wu, X., Stavnezer, J. The roles of APE1, APE2, DNA polymerase β and mismatch repair in creating S region DNA breaks during antibody class switch. Philosophical Transactions of the Royal Society B: Biological Sciences. 364 (1517), 645-652 (2009).
- Roa, S., et al. MSH2/MSH6 complex promotes error-free repair of AID-induced dU: G mispairs as well as error-prone hypermutation of A: T sites. PLoS One. 5 (6), 11182 (2010).
- Delbos, F., Aoufouchi, S., Faili, A., Weill, J. C., Reynaud, C. A. DNA polymerase η is the sole contributor of A/T modifications during immunoglobulin gene hypermutation in the mouse. The Journal of Experimental Medicine. 204 (1), 17-23 (2007).
- Maul, R. W., Gearhart, P. J. AID and somatic hypermutation. Advances in Immunology. 105, 159-191 (2010).
- Shen, H. M., Tanaka, A., Bozek, G., Nicolae, D., Storb, U. Somatic hypermutation and class switch recombination in Msh6-/- Ung-/- double-knockout mice. The Journal of Immunology. 177 (8), 5386-5392 (2006).
- Cheng, H. L., et al. Integrity of the AID serine-38 phosphorylation site is critical for class switch recombination and somatic hypermutation in mice. Proceedings of the National Academy of Sciences. 106 (8), 2717-2722 (2009).
- Lebecque, S. G., Gearhart, P. J. Boundaries of somatic mutation in rearranged immunoglobulin genes: 5'boundary is near the promoter, and 3'boundary is approximately 1 kb from V (D) J gene. The Journal of Experimental Medicine. 172 (6), 1717-1727 (1990).
- Jolly, C. J., Klix, N., Neuberger, M. S. Rapid methods for the analysis of immunoglobulin gene hypermutation: application to transgenic and gene targeted mice. Nucleic Acids Research. 25 (10), 1913-1919 (1997).
- Choi, J. E., Matthews, A. J., Michel, G., Vuong, B. Q. AID Phosphorylation Regulates Mismatch Repair-Dependent Class Switch Recombination and Affinity Maturation. The Journal of Immunology. 204 (1), 13-22 (2020).
- McBride, K. M., et al. Regulation of class switch recombination and somatic mutation by AID phosphorylation. The Journal of Experimental Medicine. 205 (11), 2585-2594 (2008).
- Liu, M., et al. Two levels of protection for the B cell genome during somatic hypermutation. Nature. 451 (7180), 841-845 (2008).
- Ross, M., Birbeck, M., Wills, V., Forrester, J., Davis, A. Peanut lectin binding properties of germinal centers of mouse lymphoid tissues. Nature. 284, 364-366 (1980).
- Zhang, J., MacLennan, I. C., Liu, Y. J., Lane, P. J. Is rapid proliferation in B centroblasts linked to somatic mutation in memory B cell clones. Immunology Letters. 18 (4), 297-299 (1988).
- Nieuwenhuis, P., Opstelten, D. Functional anatomy of germinal centers. American Journal of Anatomy. 170 (3), 421-435 (1984).
- Lau, A. W., Brink, R. Selection in the germinal center. Current Opinion in Immunology. 63, 29-34 (2020).
- Victora, G. D., Nussenzweig, M. C. Germinal centers. Annual Review of Immunology. 30, 429-457 (2012).
- Mesin, L., Ersching, J., Victora, G. D. Germinal center B cell dynamics. Immunity. 45 (3), 471-482 (2016).
- Reichert, R. A., Gallatin, W. M., Weissman, I. L., Butcher, E. C. Germinal center B cells lack homing receptors necessary for normal lymphocyte recirculation. The Journal of Experimental Medicine. 157 (3), 813-827 (1983).
- Rose, M., Birbeck, M., Wills, V., Forrester, J., Davis, A. Peanut lectin binding properties of germinal centers of mouse lymphoid tissues. Nature. 284, 364-366 (1980).
- Hamada, S., Fujita, S. DAPI staining improved for quantitative cytofluorometry. Histochemistry. 79 (2), 219-226 (1983).
- Otto, F. DAPI staining of fixed cells for high-resolution flow cytometry of nuclear DNA. Methods in Cell Biology. 33, 105-110 (1990).
- Butcher, E., et al. Surface phenotype of Peyer's patch germinal center cells: implications for the role of germinal centers in B cell differentiation. The Journal of Immunology. 129 (6), 2698-2707 (1982).
- Rogerson, B. J., Harris, D. P., Swain, S. L., Burgess, D. O. Germinal center B cells in Peyer's patches of aged mice exhibit a normal activation phenotype and highly mutated IgM genes. Mechanisms of Ageing and Development. 124 (2), 155-165 (2003).
- Potapov, V., Ong, J. L. Examining sources of error in PCR by single-molecule sequencing. PloS One. 12 (1), 0169774 (2017).
- Gonzalez-Fernandez, A., Milstein, C. Analysis of somatic hypermutation in mouse Peyer's patches using immunoglobulin kappa light-chain transgenes. Proceedings of the National Academy of Sciences. 90 (21), 9862-9866 (1993).
- Reboldi, A., Cyster, J. G. Peyer's patches: organizing B-cell responses at the intestinal frontier. Immunological Reviews. 271 (1), 230-245 (2016).
- Betz, A. G., Rada, C., Pannell, R., Milstein, C., Neuberger, M. S. Passenger transgenes reveal intrinsic specificity of the antibody hypermutation mechanism: clustering, polarity, and specific hot spots. Proceedings of the National Academy of Sciences. 90 (6), 2385-2388 (1993).
- Rada, C., Gupta, S. K., Gherardi, E., Milstein, C. Mutation and selection during the secondary response to 2-phenyloxazolone. Proceedings of the National Academy of Sciences. 88 (13), 5508-5512 (1991).
- Heise, N., Klein, U. Somatic Hypermutation and Affinity Maturation Analysis Using the 4-Hydroxy-3-Nitrophenyl-Acetyl (NP) System. Methods in Molecular Biology. 1623, 191-208 (2017).
- Smith, F., Cumano, A., Licht, A., Pecht, I., Rajewsky, K. Low affinity of kappa chain bearing (4-hydroxy-3-nitrophenyl) acetyl (NP)-specific antibodies in the primary antibody repertoire of C57BL/6 mice may explain lambda chain dominance in primary anti-NP responses. Molecular Immunology. 22 (10), 1209-1216 (1985).
- Takahashi, Y., Dutta, P. R., Cerasoli, D. M., Kelsoe, G. In situ studies of the primary immune response to (4-hydroxy-3-nitrophenyl) acetyl. V. Affinity maturation develops in two stages of clonal selection. The Journal of Experimental Medicine. 187 (6), 885-895 (1998).
- Allen, D., Simon, T., Sablitzky, F., Rajewsky, K., Cumano, A. Antibody engineering for the analysis of affinity maturation of an anti-hapten response. The EMBO Journal. 7 (7), 1995-2001 (1988).
- Cumano, A., Rajewsky, K. Clonal recruitment and somatic mutation in the generation of immunological memory to the hapten NP. The EMBO Journal. 5 (10), 2459-2468 (1986).
- Smith, K., Nossal, G., Tarlinton, D. M. FAS is highly expressed in the germinal center but is not required for regulation of the B-cell response to antigen. Proceedings of the National Academy of Sciences. 92 (25), 11628-11632 (1995).
- Hao, Z., et al. Fas receptor expression in germinal-center B cells is essential for T and B lymphocyte homeostasis. Immunity. 29 (4), 615-627 (2008).
- Cervenak, L., Magyar, A., Boja, R., László, G. Differential expression of GL7 activation antigen on bone marrow B cell subpopulations and peripheral B cells. Immunology Letters. 78 (2), 89-96 (2001).
- Naito, Y., et al. Germinal center marker GL7 probes activation-dependent repression of N-glycolylneuraminic acid, a sialic acid species involved in the negative modulation of B-cell activation. Molecular and Cellular Biology. 27 (8), 3008-3022 (2007).
- Olson, W. J., et al. Orphan Nuclear Receptor NR2F6 Suppresses T Follicular Helper Cell Accumulation through Regulation of IL-21. Cell Reports. 28 (11), 2878-2891 (2019).
- Dorsett, Y., et al. MicroRNA-155 suppresses activation-induced cytidine deaminase-mediated Myc-Igh translocation. Immunity. 28 (5), 630-638 (2008).
- Goetz, C., Hammerbeck, C., Bonnevier, J. . Flow Cytometry Basics for the Non-Expert. , (2018).
- Hawley, T. S., Herbert, D. J., Eaker, S. S., Hawley, R. G. . Flow Cytometry Protocols. , (2004).
- Costa, E., et al. A new automated flow cytometry data analysis approach for the diagnostic screening of neoplastic B-cell disorders in peripheral blood samples with absolute lymphocytosis. Leukemia. 20 (7), 1221-1230 (2006).
- McKinnon, K. M. Flow cytometry: An overview. Current Protocols in Immunology. 120 (1), 1-11 (2018).
- McKinnon, K. M. Multiparameter Conventional Flow Cytometry. Methods in Molecular Biology. , 139-150 (2018).
- Lucchesi, S., et al. Computational Analysis of Multiparametric Flow Cytometric Data to Dissect B Cell Subsets in Vaccine Studies. Cytometry Part A. 97, 259-267 (2019).
- Longerich, S., Tanaka, A., Bozek, G., Nicolae, D., Storb, U. The very 5' end and the constant region of Ig genes are spared from somatic mutation because AID does not access these regions. The Journal of Experimental Medicine. 202 (10), 1443-1454 (2005).
- Retter, I., et al. Sequence and characterization of the Ig heavy chain constant and partial variable region of the mouse strain 129S1. The Journal of Immunology. 179 (4), 2419-2427 (2007).
- Shen, H. M., Peters, A., Baron, B., Zhu, X., Storb, U. Mutation of BCL-6 gene in normal B cells by the process of somatic hypermutation of Ig genes. Science. 280 (5370), 1750-1752 (1998).
- Richter, K., et al. Altered pattern of immunoglobulin hypermutation in mice deficient in Slip-GC protein. Journal of Biological Chemistry. 287 (38), 31856-31865 (2012).
ABOUT JoVE
Copyright © 2024 MyJoVE Corporation. All rights reserved