
Summary
Abstract
Introduction
Protocol
Representative Results
Discussion
Acknowledgements
Materials
References
Neuroscience
Robuuste en zeer reproduceerbare generatie corticale hersenorganoïden voor het modelleren van neuronale senescentie in de hersenen in vitro
In deze studie bieden we een gedetailleerde techniek voor een eenvoudig maar robuust corticale organoïde kweeksysteem met behulp van standaard feedervrije hPSC-culturen. Dit is een snel, efficiënt en reproduceerbaar protocol voor het genereren van organoïden die aspecten van hersensenescentie in vitro modelleren.
Hersenorganoïden zijn driedimensionale modellen van het zich ontwikkelende menselijke brein en bieden een aantrekkelijk, geavanceerd platform voor ziektemodellering en grootschalige genomische en medicijnscreening. Vanwege de zelforganiserende aard van cellen in hersenorganoïden en het groeiende aantal beschikbare protocollen voor hun generatie, zijn problemen met heterogeniteit en variabiliteit tussen organoïden geïdentificeerd. In dit protocolartikel beschrijven we een robuust en repliceerbaar protocol dat deze problemen grotendeels overwint en binnen 1 maand corticale organoïden genereert uit neuro-ectodermale voorlopercellen, en dat langer dan 1 jaar kan worden gehandhaafd. Dit zeer reproduceerbare protocol kan gemakkelijk worden uitgevoerd in een standaard weefselkweekkamer en resulteert in organoïden met een rijke diversiteit aan celtypen die typisch worden aangetroffen in de zich ontwikkelende menselijke cortex. Ondanks hun vroege ontwikkelingssamenstelling, zullen neuronen en andere menselijke hersenceltypen de typische tekenen van senescentie in neuronale cellen beginnen te vertonen na langdurige in vitro cultuur, waardoor ze een waardevol en nuttig platform zijn voor het bestuderen van verouderingsgerelateerde neuronale processen. Dit protocol schetst ook een methode voor het detecteren van dergelijke senescente cellen in corticale hersenorganoïden met behulp van senescentie-geassocieerde bèta-galactosidase-kleuring.
Onze huidige kennis van het menselijk brein is grotendeels gebaseerd op diermodellen en post-mortem hersenspecimens. Stamcelbiologie is een snel voortschrijdend veld dat nieuwe inzichten biedt in de basisbiologie van de ontwikkeling van het menselijk brein en de pathologische oorzaken van menselijke hersenaandoeningen. Menselijke pluripotente stamcellen (hPSC's) zijn een waardevol hulpmiddel voor het modelleren van het menselijk brein via de generatie van organoïden, orgaanachtig driedimensionaal (3D) weefsel dat meestal de ontwikkelingstrajecten, cellulaire samenstelling en architectuur van het zich ontwikkelende menselijke brein samenvat. Hersenorganoïden zijn zelfgeassembleerd en samengesteld uit neurale stamcellen, gespecificeerde neurale voorlopers, volwassen neuronen en gliaceltypen. Organoïden bieden daarom een unieke kans om het vroege menselijke brein te bestuderen, dat vaak ontoegankelijk is voor directe experimenten, maar ook intrinsieke beperkingen heeft, zoals de afwezigheid van vasculatuur en een immuunsysteem.
Methodologieën om hersenorganoïden te genereren zijn op twee verschillende manieren nagestreefd: ongeleide en geleide differentiatie. Ongeleide hersenorganoïde methoden vertrouwen op de spontane intrinsieke differentiatiecapaciteiten van de stamcellen die weefselmorfogenese 1,2 aandrijven en zorgen voor de opkomst van een verscheidenheid aan cellijnidentiteiten, variërend van voorhersenen, middenhersenen en achterhersenen, tot choroïde plexus, netvlies en mesoderm. Daarentegen vereisen geleide hersenorganoïde methoden substantieel gebruik van externe factoren om hPSC's te sturen naar de gewenste patroonvorming van neuronale afstammingslijnen die één hersengebiedtype vertegenwoordigen, zoals mediale ganglionaire eminentie3, voorhersenen4, middenhersenen5, hypothalamus6, cerebellum7 en choroïde plexus8. Dit vermogen om verschillende hersengebieden met verschillende cellijningen te genereren, en het potentieel om deze naar believen te fuseren, maakt hersenorganoïden een uitstekend model voor het onderzoeken van de ontwikkeling van de menselijke hersenen en het ontcijferen van de onderliggende mechanismen van hersengerelateerde ziekten. Hoewel deze methoden voor het genereren van hersenorganoïden een doorbraak bieden in het modelleren van menselijke hersengebieden, blijven de variabiliteit en heterogeniteit tussen organoïden een significante beperking voor systematische en kwantitatieve studies, zoals geneesmiddelenscreening.
Het huidige protocol is gebaseerd op een methode die is ontwikkeld in ons recente artikel9 en omvat de selectieve differentiatie van hPSC-kolonies naar neuro-ectoderm (NEct) identiteit met dubbele SMAD-remmers (SB-431542 en LDN 193189), die vervolgens het vermogen hebben om zichzelf binnen 4 dagen te organiseren in 3D neuro-epitheelsferoïden onder invloed van FGF2-signalering. Deze neuro-epitheelsferoïden genereren op betrouwbare wijze homogene corticale organoïden met een in vivo-achtige cellulaire samenstelling binnen 4 weken na differentiatie. Het hier beschreven protocol is gebaseerd op onze eerdere bevindingen waaruit blijkt dat remming van dubbele SMAD (Suppressor of Mothers Against Decapentaplegic) signalering de differentiatie van hPSC's bevordert in de richting van rostrale neurale stamcellen afgeleid van neuro-ctodermale voorlopers10 door, onder andere, endodermale, mesodermale en trophectodermcel lotkeuze11 te remmen . Bovendien veroorzaakt de inbedding van de neuro-epitheliumsferoïden in de hESC-gekwalificeerde keldermembraanmatrix een aanzienlijke ontluiking van de neuro-epithelia, waardoor ventrikels met apicobasale polariteit worden gevormd. Grootschalige cultuur toonde reproduceerbaarheid en homogeniteit van corticale organoïden onafhankelijk van cellijnen, klonen of batches, en vertegenwoordigt dus een betrouwbaar en stabiel stamcelsysteem om de vroege menselijke corticale ontwikkeling in gezondheid en ziekte in vitro na te bootsen. We schetsen verder een protocol voor het detecteren van senescente neuronale celmarkers in hPSC's-afgeleide corticale hersenorganoïden die gedurende langere tijd zijn gekweekt.
Na het platen van hPSC's met een zaaidichtheid van 20% -30%, worden de cellen gedurende 3 dagen behandeld met dubbele SMAD-remmers om hPSC-kolonies te differentiëren in de richting van neuro-ectodermale kolonies. Deze kolonies worden vervolgens voorzichtig opgetild met dispase en gezaaid in ultra-lage bevestiging 6-well platen aangevuld met FGF2. De drijvende 2D-kolonies organiseren zichzelf 's nachts in 3D neuro-elektro-intestinale sferoïden en worden gedurende 4 dagen in N2-medium gehouden, dagelijks aangevuld met FGF2. Zodra de sferoïden de neuro-epitheliale laag hebben vastgesteld, kunnen ze worden ingebed in de keldermembraanmatrix. Door routinematig een nieuw terminaal differentiatiemedium toe te voegen, zullen de onderzoekers progressieve expansie en ontluiking van neuro-epithelia in corticale organoïden waarnemen. Onderzoekers kunnen deze organoïden dissociëren om transcriptionele en proteomische profilering uit te voeren. Bovendien wordt brightfield imaging aanbevolen voor het bewaken van de kwaliteit van de corticale organoïden. Analyse kan worden uitgevoerd door fixatie, cryosectie en immunostaining. Beschrijvingen en methoden voor deze technieken zijn eerder beschreven12. Uiteindelijk stelt dit protocol onderzoekers in staat om snel en robuust homogene corticale hersenorganoïden te genereren voor het modelleren van het zich ontwikkelende menselijke corticale brein, met lage kosten en beperkte apparatuur, en voor het bestuderen van aspecten van cellulaire neuronale senescentie, zoals uiteengezet in dit artikel.
1. Corticale hersenorganoïden generatie
OPMERKING: Alle stappen in dit gedeelte van het protocol vinden plaats in een bioveiligheidskap van klasse 2, tenzij anders vermeld.
- Inductie van 2D neuro-ctodermale kolonies uit hPSC 2D-cultuur (dagen -1 tot 3)
- Vóór inductie, plaat de hPSC-kolonies op een hESC-gekwalificeerde keldermembraanmatrix in een 6-well plaat met een dichtheid van 20% -30%. Bereik deze dichtheid door hPSC-kolonies van één put van een 6-well plaat bij 60% confluency in drie putten van een 6-well plaat te passeren.
- Voor keldermembraanmatrixcoating verdunt u de keldermembraanmatrix in een verhouding van 1:50 in een gewoon basaal medium. Deponeer gelijkmatig 1 ml / put van een 6-well plaat, incubeer gedurende 1 uur bij kamertemperatuur (RT) en adem dan op.
- Houd hPSC-kolonies gedurende 1 dag in 2 ml van een serumvrij celkweekmedium voorafgaand aan differentiatie.
- Inspecteer op de dag van hPSC-differentiatie de hPSC-kolonies met behulp van brightfield-microscopie bij 4x tot 10x vergroting om gezonde kolonies te garanderen zonder detecteerbare differentiatie.
OPMERKING: Gezonde hPSC's vormen krappe randkolonies met cellen met een grote kern, een zeer klein cytoplasma en prominente nucleoli. Gedifferentieerde iPSC-kolonies zullen duidelijke morfologische verschillen vertonen met die van de hierboven beschreven hPSC-kolonies, met name rond de buitenranden van de kolonies of in het midden. - Voeg de in tabel 1 vermelde reagentia toe om het N2-medium te vormen dat nodig is voor differentiatie. Breng dit medium voor gebruik naar RT.
- Eenmaal bij RT, aspirateer het serumvrije celkweekmedium uit elke put van de 6-well plaat en vervang door 2 ml N2-medium voorzichtig toegevoegd door een serologische pipet van 5 ml.
- Voeg de dubbele SMAD-remmers, SB-431542 (10 μM) en LDN 193189 (100 nM) toe.
OPMERKING: De SMAD-remmers kunnen worden toegevoegd aan het N2-medium nadat het medium in elke put is geplaatst, of aan de vereiste hoeveelheid N2-medium voordat het serumvrije celkweekmedium wordt vervangen. Remmers kunnen gelijkmatig in het medium worden opgenomen door de plaat voorzichtig te draaien of de buis met het medium en de remmers 3-4 keer om te keren. - Voeg de komende 2 dagen dagelijks verse N2-media aangevuld met SB-431542 (10 μM) en LDN 193189 (100 nM) toe aan elke put.
OPMERKING: Vers N2-medium wordt toegevoegd om de langdurige blootstelling van cellen aan de DMSO te verminderen die wordt gebruikt om SB-431542 en LDN 193189 verbindingen op te lossen en cytotoxiciteit te voorkomen.
- Generatie van 3D neuro-ectodermale sferoïden uit geïnduceerde 2D neuro-ectodermale kolonies (dagen 3 tot 7)
- Til de geïnduceerde neuro-ectodermale kolonies op met behulp van dispase volgens stap 1.2.2-1.2.8.
- Verwijder eerst 2 ml N2-medium van de 6-wellsplaat en was 1x met HBSS om ervoor te zorgen dat al het N2-medium wordt verwijderd.
OPMERKING: N2-medium kan interfereren met de enzymactiviteit van dispase, waardoor adequate loslating van neuro-ectodermale kolonies uit de put wordt voorkomen. - Voeg 1 ml van 2,4 eenheid / ml dispase toe aan elke koloniebevattende put.
- Incubeer de put gedurende 20-25 min (maximaal 30 min) bij 37 °C. Controleer regelmatig op kolonieontleding.
OPMERKING: Kleine kolonies kunnen binnen 20 minuten losraken. Kolonies die na 30 minuten vast blijven zitten, moeten worden genegeerd. - Voeg na incubatie 1 ml N2-medium toe aan de put om de activiteit van het dispase-enzym te stoppen en breng de kolonies over in een buis van 15 ml met behulp van een P1000-pipetpunt met brede boring of een gemodificeerde P1000-pipetpunt die met steriele schaar is gesneden (waardoor het een P1000-tip met brede boring wordt).
- Laat de kolonieklompen met zwaartekracht naar de bodem van de buis zinken.
OPMERKING: Dit proces duurt ongeveer 1 minuut. - Zodra de klonten zijn gezonken, verwijdert u het supernatant voorzichtig met een standaard P1000 pipetpunt en vervangt u het door 1 ml vers N2-medium. Herhaal deze wasstap drie keer om ervoor te zorgen dat dispase volledig wordt verwijderd.
OPMERKING: Elke resterende dispase zal een uniforme vorming van neuro-ectodermale sferoïden voorkomen en celdood veroorzaken. - Resuspenseer na het wassen de celklonten in 3 ml N2-medium en breng over naar een put van een 6-wellsplaat en voeg 40 ng / ml bFGF toe.
OPMERKING: Als een groot aantal neuro-ectodermale kolonies werd losgemaakt, konden deze kolonies over twee of meer putten van een 6-wellsplaat worden geplaatst om sferoïdenfusie te voorkomen. Controleer 24 uur na het loslaten van de kolonies of de sferoïden zich hebben gevormd. - Houd de sferoïden in dezelfde media voor de komende 3-4 dagen, maar voeg dagelijks verse bFGF (40 ng / ml) toe aan elke put om neuro-ectodermale celproliferatie, zelforganisatorie te bevorderen en neuro-epithelia te induceren en uit te breiden.
OPMERKING: Als sferoïden met een hogere dichtheid zijn verguld, zullen de media waarschijnlijk geel worden en moeten ze om de 2 dagen worden vervangen door verse bFGF (40 ng / ml). Dit wordt echter niet aanbevolen en in plaats daarvan moet een lager aantal sferoïden in elke put worden gehandhaafd om dit probleem te voorkomen. Sferoïden kunnen na 3 dagen in de keldermembraanmatrix worden ingebed als neuro-epithelia duidelijk zijn. Als neuro-epithelia niet duidelijk zijn of er niet sterk uitzien, houd dan de sferoïden nog een dag aan en controleer opnieuw.
- Corticale hersenorganoïde differentiatie en onderhoud (dag 8)
- Bereid de terminaldifferentiatiemedia (DM) voor met behulp van de in tabel 2 vermelde reagentia. Breng deze media naar RT.
- Ontdooi 100% hESC-gekwalificeerde keldermembraanmatrix op ijs.
- Bereid een vel parafilm met kuiltjes gesteriliseerd met 70% ethanol in een petrischaaltje van 10 cm en plaats het op een stereomicroscoop onder een kap.
OPMERKING: Een lege lade met 200 μL pipetpunten kan worden gebruikt om een raster van kuiltjes te genereren. - Snijd het uiteinde af van een pipetpunt van 100 μL (om het breed te maken). Dit zal worden gebruikt voor het inbedden van neurale sferoïden in de keldermatrix zonder ze te breken.
- Selecteer met behulp van een stereomicroscoop neurale sferoïden van vergelijkbare grootte (500 μm) uit de 6-well plaat en breng ze over naar de parafilm-dimples met behulp van de 100 μL pipetpunt met brede boring, waarbij een enkele neurale sferoïde per kuiltje wordt geplaatst.
- Verwijder voorzichtig overtollige media, zodat er net genoeg overblijft om de sferoïden te bedekken.
OPMERKING: Dit is om de kwaliteit van de sferoïden te behouden voordat de keldermatrix wordt toegevoegd en om ervoor te zorgen dat ze niet uitdrogen. - Voeg voorzichtig 18 μL keldermatrix toe aan de sferoïde en plaats de sferoïde in het midden van de matrixdruppel. Gebruik het uiteinde van een pipetpunt van 10 μL om de sferoïden in de matrix te centreren.
OPMERKING: Probeer de keldermatrix zo snel mogelijk toe te voegen om te voorkomen dat de overtollige media rond de sferoïden uitdrogen. - Bedek de petrischaal van 10 cm en breng deze over in de couveuse en incubeer de parafilmschaal met keldermatrix ingebedde sferoïden bij 37 °C gedurende 25 minuten.
- Spoel na incubatie de ingebedde sferoïden af in een 24-well-plaat met lage bevestiging met behulp van een P1000-tip met 0,5 ml dm-media, waarbij u ervoor zorgt dat individuele ingebedde sferoïden elk in één put worden geplaatst.
OPMERKING: Als meer dan één ingebedde sferoïde in één put valt, gebruik dan een P1000-tip met brede boring om de andere sferoïde in een nieuwe put over te brengen. - Behoud de gedifferentieerde corticale hersenorganoïden in de DM-media gedurende langere tijd, waarbij mediaveranderingen elke 2 dagen optreden wanneer de organoïden groter en ouder worden.
OPMERKING: Tijdens de eerste week van differentiatie kan het medium om de 3 dagen worden vervangen.
2. Karakterisering van neuronale veroudering in corticale organoïden
- Proces corticale organoïden voor cryosecties:
OPMERKING: De stappen werden uitgevoerd in een bioveiligheidskap van klasse 2.- Bereid 2 ml buizen, elk gevuld met 1,5 ml 4% paraformaldehyde (PFA).
- Snijd het uiteinde van een P1000 pipetpunt (om er een brede boring van te maken) en breng elke organoïde voorzichtig over op een van de 2 ml-buizen die hierboven zijn voorbereid (één organoïde per buis).
OPMERKING: Om te voorkomen dat overtollige DM-media zich vermengen met de PFA, laat u de organoïde naar de opening van de P1000-pipetpunt zakken en laat u de punt net boven de bovenkant van de PFA in de buis rusten voordat u de organoïde eruit pipept. Dit zal de onderzoeker in staat stellen om alleen de organoïde en zeer minimale media over te brengen. - Laat het fixatieproces gedurende 1 uur plaatsvinden bij 4°C.
- Gebruik een ongesneden P1000 pipetpunt om overtollige PFA voorzichtig op te zuigen en voeg 1,5 ml koude 1x PBS toe.
- Breng de buizen over naar een orbitale shaker die op 70 rpm is ingesteld gedurende 10 minuten bij RT.
- Herhaal het wasproces met koude 1x PBS drie keer om ervoor te zorgen dat alle PFA grondig is verwijderd.
OPMERKING: Gooi PFA niet weg in gewone afvalcontainers; bereid hiervoor in plaats daarvan een specifieke container voor het verwijderen van chemisch afval voor, omdat PFA een gevaar vormt. - Dompel de organoïden onder in 1x PBS met 30% sucrose en incubeer bij 4 °C totdat alle organoïden naar de bodem van de buis zijn gezonken.
OPMERKING: De tijd die nodig is om de organoïden te laten zinken, is afhankelijk van de grootte / leeftijd van de organoïden. 3 maanden oude organoïden kunnen tot 5 uur duren. - Breng met behulp van een gesneden P1000-pipetpunt met brede boring voorzichtig drie tot vijf organoïden over in een montagemal met een montageoplossing gemaakt van 30% sucrose en 100% optimale snijtemperatuur (OCT) medium, in een verhouding van 3: 2.
- Gebruik een pipetpunt van 10 μL met behulp van een stereomicroscoop om organoïden in een rasterachtig patroon te oriënteren en te positioneren.
- Plaats de mal op droogijs om de sucrose OCT-oplossing te stollen voordat u verder gaat met cryo-sectioning (16-20 μm) met behulp van een cryostaat.
OPMERKING: Voor senescentie-geassocieerde bèta-galactosidase moeten alle weefsels worden verwerkt voor sectie zodra ze zijn gezonken. Voor immunofluorescentie kunnen weefsels worden verwerkt voor sectie de volgende dag. Alle dia's met secties moeten bij -20 °C worden bewaard vóór de daaropvolgende immunofluorescentie of bèta-galactosidase, zo niet onmiddellijk gekleurd.
- Proces voor analyse van senescentie in de corticale hersenorganoïden:
OPMERKING: De volgende stappen kunnen worden uitgevoerd op een gewone laboratoriumbank.- Breng de dia's over in een microscoopglaasje met een deksel en was het doorsneden organoïde weefsel drie keer met 1x PBS gedurende 10 minuten bij RT om overtollige montageoplossing te verwijderen.
- Incubeer vervolgens het gewassen weefsel met vers gemaakte bèta-galactosidase-kleuringsoplossing 's nachts bij 37 °C.
OPMERKING: De bèta-galactosidasekleuringsoplossing is gemaakt van fosfaatbuffer (voor 10 ml fosfaatbuffer: 8,15 ml 1 M NaH2PO4, 1,85 ml 1M Na2HPO4) aangepaste pH = 6, 100 m M kaliumhexacyanoferraat (III), 100 mM kaliumhexacyanoferraat (II) trihydraat, 5 M NaCl, 1 M MgCl2, 20 mg/ml X-Gal. Vermijd het gebruik van een standaard celkweekincubator die CO2 bevat, omdat de CO2 de pH van de bèta-galactosidase-kleuringsoplossing zal veranderen. - Was de gekleurde weefsels met 1x PBS drie keer gedurende 10 minuten elk op RT om de bèta-galactosidase-oplossing te verwijderen.
- Monteer de gewassen weefsels met een glazen antivet mountant en laat de montageoplossing gedurende 30 minuten stollen bij RT voordat u het onder de microscoop bekijkt.
Hier hebben we een robuust protocol beschreven waarmee onderzoekers homogene hPSC-afgeleide corticale hersenorganoïden kunnen genereren die het in vivo menselijke corticale hersengebied nabootsen binnen 1-3 maanden van cultuur. hPSC-kolonies worden eerst gekweekt in differentiatiemedia om neuro-ectodermale kolonies te genereren, die vervolgens kunnen worden gebruikt om neurale sferoïden te vormen. Deze sferoïden worden vervolgens ingebed in een keldermembraanmatrix en gedurende langere tijd onderhouden om organoïden te produceren die kunnen worden gebruikt om neuronale veroudering te modelleren (zie figuur 1 voor een overzicht van het protocol). Het is vermeldenswaard dat het kweken van deze organoïden in ultra-ongecoate 24-well platen cellulaire stress veroorzaakt en senescentie-geassocieerde fenotypen bevordert gedurende 13 weken in vitro cultuur. Organoïden afgeleid van dit protocol kunnen ook worden onderhouden in geroerde bioreactoren voor optimale groei en differentiatie van corticale plaat neurale cellen of op de lucht-vloeistof interface.
Om te beginnen worden hPSC-kolonies gedurende 1 dag gekweekt voorafgaand aan neuro-ectodermale differentiatie. Het is van cruciaal belang dat deze hPSC-kolonies worden gekweekt tot slechts 20% -30% samenvloeiing en van de hoogst mogelijke kwaliteit zijn: een strakke platte monolaag zonder gedifferentieerde cellen die de kolonies besmetten (figuur 2A, B). De pluripotentie van de hPSC-kolonies moet worden bevestigd door de expressie van markers zoals NANOG (figuur 2C). De gevalideerde hPSC-kolonies worden vervolgens blootgesteld aan de N2 neuro-ectodermale differentiatiemedia met SB-431542 en LDN 193189. Na 3 dagen onderhoud in dit medium zouden de hPSC-kolonies moeten hebben gedifferentieerd tot neuro-ectodermale kolonies en niet langer dezelfde strakke platte monolaagmorfologie van de hPSC's vertonen (figuur 2B), maar zullen het langere kolomvormige cellen worden (figuur 2B). Deze cellen zullen ook negatief zijn voor pluripotentiemarkers zoals NANOG (figuur 2C).
Het is in dit stadium dat de neuro-ectodermale kolonies enzymatisch worden losgemaakt met dispase, en elke gezonde en met succes losgemaakte kolonie mag zichzelf organiseren en een jonge neurale sferoïde vormen (Figuur 2D, Aanvullende video 1). Alleen gezonde, schone neuro-ectodermale kolonies zullen loskomen binnen het tijdsbestek dat is gespecificeerd voor dispase-activiteit; alle andere kolonies moeten worden genegeerd, omdat ze zullen resulteren in een slechtere kwaliteit van sferoïden. Bij dagelijkse blootstelling aan FGF2 in de N2-media zullen de neurale stamcellen (SOX2+) in deze sferoïden (figuur 2D, dag 1) zich vermenigvuldigen en een aanzienlijk aantal neurale rozetten vormen (figuur 2D, dag 4). Deze rozetten zullen de tight junction en epitheliale marker ZO1 uitdrukken in cellen in het midden van de rozetten en langs de buitenrand van de sferoïde, wat de apicale-basale polariteit van de sferoïde aantoont (figuur 2D, dag 4). De methode voor wholemount 3D-beeldvorming van sferoïden is beschreven vóór13. Dagelijkse inspectie van de sferoïden moet de vorming van een strakke, donkere buitenrand en heldere periferie van de sferoïden, dit is de neuro-epitheliale laag, ophelderen. Deze laag moet na 3-4 dagen voldoende gevormd zijn met een geschatte diameter van 500 μm, waarna de sferoïden in de keldermatrix kunnen worden ingebed. Als deze laag niet of slechts zwak gevormd is, zijn de sferoïden niet voldoende ontwikkeld om verder te gaan. Het wordt aanbevolen om nog een dag te wachten om een verandering waar te nemen, maar als dit niet wordt waargenomen, negeer dan deze sferoïden.
Een representatief brightfield-beeld van de sferoïden na 3 dagen cultuur is te zien in figuur 2D. Sferoïden met een strakke neuro-epitheliale laag die niet zijn gefuseerd met andere naburige sferoïden, semi-transparant weefsel hebben en neurale rozetvorming vertonen, worden gekozen om te worden ingebed in de keldermatrix. Eenmaal ingebed, zal de sferoïde zich snel vermenigvuldigen en beginnen te ontluiken: knooppunten van compact weefsel zullen verschijnen en zich naar buiten uitbreiden vanuit het hoofdlichaam van de sferoïde. Dit is duidelijk tussen 1-3 weken in de keldermatrix en kan worden waargenomen over meerdere cellijnen (figuur 3A). Kwantitatieve analyse van de ingebedde sferoïden bevestigt de aanwezigheid van epitheelcellen in maximaal 100% van de sferoïden over drie verschillende cellijnen, wat de homogeniteit en reproduceerbaarheid bevestigt die van dit protocol wordt verwacht (figuur 3B). De kwantificering van de diameter van organoïden tijdens in vitro differentiatie bevestigt verder de reproduceerbaarheid over verschillende lijnen van hPSC (figuur 3C). Als ontluiking niet optreedt, ontwikkelen de sferoïden zich niet op de juiste manier en moeten ze worden weggegooid. Zodra de sferoïden in een matrix zijn ingebed, vordert hun ontwikkeling en worden de sferoïden nu organoïden genoemd. Immunofluorescentiekleuring bevestigt ook de aanwezigheid van neurale voorlopercellen (PAX6) en corticale laagmarkers gekleurd met CTIP2 en SATB2 in de organoïden met duidelijke gelaagdheid (figuur 3D, E). Deze gelaagdheid is waarneembaar over verschillende tijdstippen van organoïde onderhoud (figuur 3D, E). De methode van immunohistochemie van weefsels is vóór14 beschreven.
Een mogelijke toepassing van deze organoïden is om te bestuderen hoe neuronale verouderingsgerelateerde processen de hersenen beïnvloeden. Om dit te onderzoeken, worden met succes gegenereerde organoïden geoogst uit meerdere verschillende tijdspunten voor sectie en kleuring voor standaard moleculaire biomarkers van senescentie zoals senescentie-geassocieerde bèta-galactosidase en p21. Figuur 4A toont een representatief beeld van senescentie-geassocieerde bèta-galactosidase kleuring van organoïden 4 en 13 weken na inbedding in de keldermembraanmatrix. Tussen week 4 en 13 is er een duidelijke toename van de aanwezigheid van senescentie-geassocieerde bèta-galactosidase, wat suggereert dat cellulaire senescentie, een erkende aanjager van organismale veroudering, in deze tijd in cultuur is opgetreden. Immunofluorescentiekleuring van organoïden in week 13 bevestigde de aanwezigheid van een andere senescentiemarker, p21, gelijktijdig gelabeld met de volwassen corticale neuronale marker (CTIP2) en kan worden gezien in figuur 4B. Er moet echter worden opgemerkt dat de aanwezigheid van p21 een marker is van celcyclusstilstand en op zichzelf geen definitieve marker van senescentie is, en detectie van andere markers van senescentie zoals p16 en SASP (senescentie-geassocieerd secretoir fenotype) factoren worden aanbevolen om cellen definitief als senescent te identificeren.
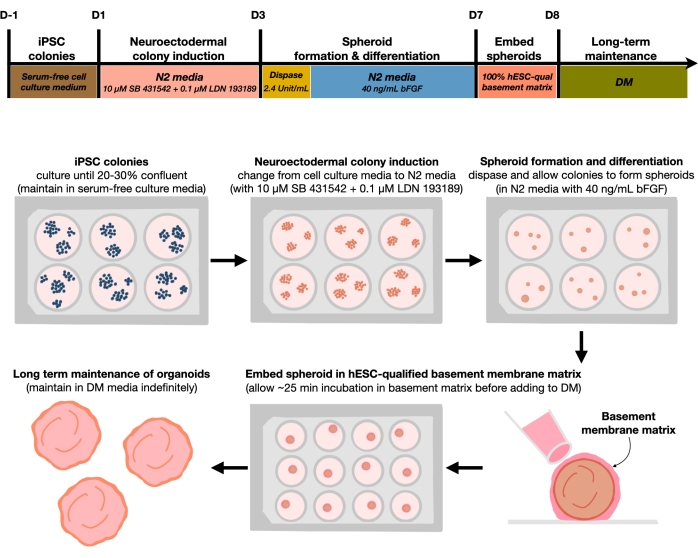
Figuur 1: Schematisch diagram voor het genereren van reproduceerbare corticale hersenorganoïden. Schematische workflow van de experimentele procedure voor de generatie van corticale hersenorganoïden uit hPSC's die in het feedervrije medium worden gehandhaafd. De workflow biedt een overzicht van zes stappen om de 2D hPSC's te differentiëren in 3D-patroon corticale plaat menselijke weefsels in organoïden. Klik hier om een grotere versie van deze figuur te bekijken.
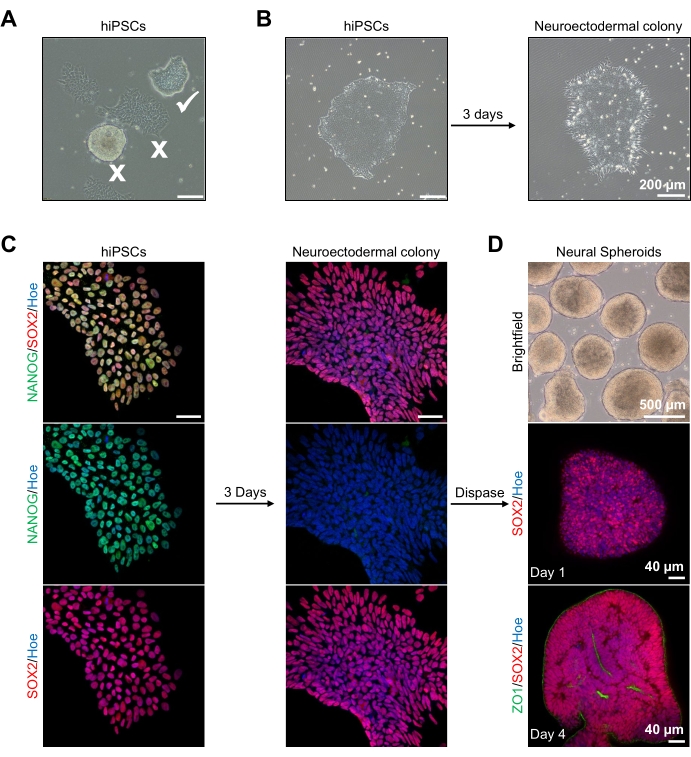
Figuur 2: Generatie van neurale sferoïden afgeleid van neuro-ectodermale kolonies-hPSC's. (A) Representatieve beelden van menselijke PSC met optimum (witte teek) en gedifferentieerde kolonies (wit kruis). Schaalbalk: 200 μm, 4x vergroting. (B) Representatief beeld van neuro-ectodermale kolonie afgeleid van hPSC's na 3 dagen van dubbele SMAD-remmerbehandelingen. Schaalbalk: 200 μm, 4x vergroting. (C) Menselijke PSC-kolonies werden gedifferentieerd naar neuro-ectodermale kolonies. Beelden tonen kleuring van PSC (op dag 1) en neuro-ectodermale (op dag 3) kolonies met SOX2 (Rood), NANOG (Groen), alle kernen werden tegengekleurd met Hoechst 33342 (blauw). Schaalbalk: 40 μm, 100x vergroting. (D) Afbeeldingen die de ontwikkelingsstadia van corticale hersensferoïden in de loop van de tijd in cultuur in vitro onder brightfield tonen, en wholemount immunostained met SOX2 (Rood) op dag 1, en dubbel immunostained met SOX2 (rood) en ZO1 (Groen) op dag 4. De schaalbalk van het brightfield-beeld is 500 μm, 4x vergroting, schaalbalken van de onderste afbeeldingen zijn 40 μm, 20x vergroting. Klik hier om een grotere versie van deze figuur te bekijken.
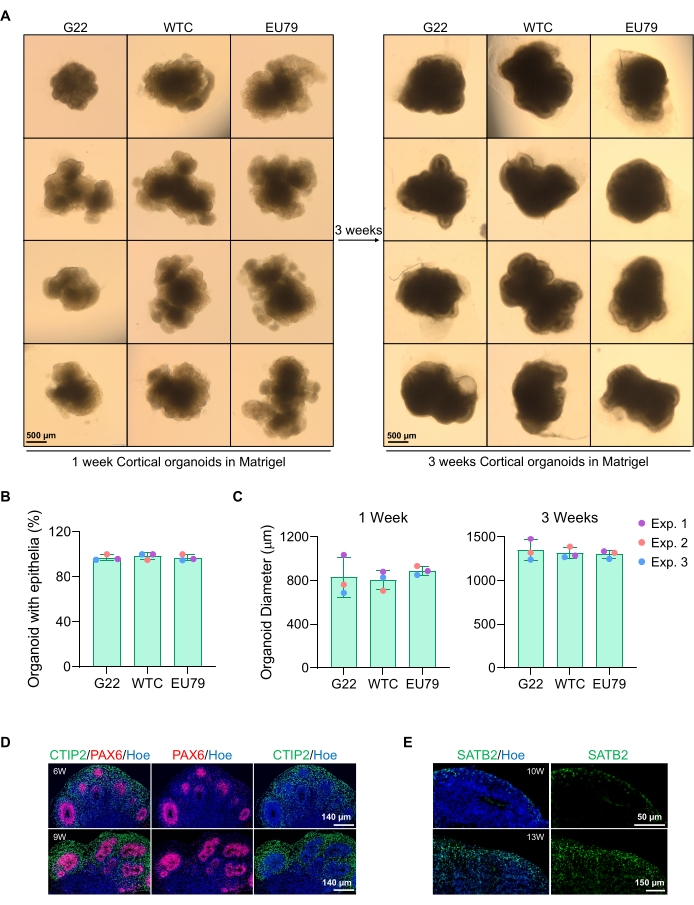
Figuur 3: Karakterisering van corticale hersenorganoïden afgeleid van verschillende hPSC-lijnen. (A) Representatieve beelden van corticale hersenorganoïden afgeleid van G22-, WTC- en EU79-menselijke iPSC-lijnen gekweekt gedurende 3 weken in vitro. De schaalbalk van alle afbeeldingen is 500 μm, 2x vergroting. (B) Percentages van de succesvolle generatie corticale hersenorganoïden na 3 weken in vitro differentiatie in verschillende hPSC-lijnen (G22, WTC en EU79). N = 3. De gegevens worden gepresenteerd als gemiddelde ± standaarddeviatie. (C) Staafdiagrammen die de groei van corticale hersenorganoïden (gebaseerd op gemiddelde diameter) tonen in week 1 en 3 van in vitro differentiatie in verschillende lijnen van menselijke pluripotente stamcellijnen (G22, WTC en EU79). N = 3. De gegevens worden gepresenteerd als gemiddelde ± standaarddeviatie. (D) Representatieve beelden van secties van 6 weken en 9 weken oude corticale hersenorganoïden afgeleid van G22 hPSC's, immunostained voor ventrikelzone PAX6 (rood) en corticale plaat CTIP2 (groen). Alle secties waren voorzien van Hoechst 33342 (blauw). Schaalbalk = 140 μm, 20x vergroting. W is week. (E) Representatieve beelden van secties van 10 weken en 13 weken oude corticale hersenorganoïden afgeleid van WTC hPSC's, immunoskleurd voor corticale laag IV SATB2 (groen). Alle secties waren voorzien van Hoechst 33342 (blauw). 10 weken beeld Schaalbalk = 50 μm, 40x vergroting. 13 weken beeld Schaalbalk = 150 μm, 40x vergroting. W is week. Klik hier om een grotere versie van deze figuur te bekijken.
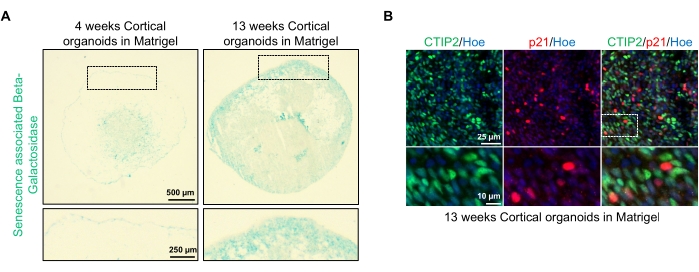
Figuur 4: Karakterisering van senescentie in corticale hersenorganoïden afgeleid van hPSC's. (A) Representatieve beelden van secties van menselijke corticale hersenorganoïden afgeleid van WTC hPSC's gekweekt gedurende 4 en 13 weken in vitro en gekleurd met SA-β-gal. Schaalbalk = 500 μm, schaalbalk van ingezoomde beelden = 250 μm, 4x vergroting. Het gestippelde vak geeft een vergrote afbeelding aan. (B) Representatieve beelden van secties van 13 weken oude corticale hersenorganoïden afgeleid van menselijke EU79 hPSC's, immunos gekleurd voor corticale neuronen CTIP2 (groen) en p21 (rood). Alle secties waren voorzien van Hoechst 33342 (blauw). Schaalbalk = 25 μm, schaalbalk van ingezoomde beelden = 10 μm, 40x vergroting. Klik hier om een grotere versie van deze figuur te bekijken.
| Mediacomponenten | Concentratie |
| DMEM Voedingsmix F12 10x 500 ml (DMEM/F-12) | |
| N2 Supplement 5 ml (100x) | Aangevuld met 1% |
| B 27 Supplement 10 ml | Aangevuld met 2% |
| MEM niet-essentiële aminozuren oplossing (100x) | Aangevuld met 1% |
| Penicilline-Streptomycine (10.000 U / ml) | Aangevuld met 1% |
| 2-Mercaptoethanol 50 ml (1000x) | Aangevuld met 0,1% |
Tabel 1: N2 Medium. De tabel bevat de reagentia die nodig zijn om het N2-medium te bereiden.
| Mediacomponenten | Concentratie |
| DMEM Voedingsmix F12 10x 500 ml (DMEM/F-12) | DM media is gemaakt met 1:1 verhouding van DMEM/F12 en Neurobasale media |
| Neurobasaal medium | |
| N2 Supplement 5 ml (100x) | Aangevuld met 0,5% |
| B 27 Supplement 10 ml | Aangevuld met 1% |
| MEM niet-essentiële aminozuren oplossing (100x) | Aangevuld met 1% |
| GlutaMAX Supplement 100x | Aangevuld met 1% |
| Penicilline-Streptomycine (10.000 U / ml) | Aangevuld met 1% |
| Insuline oplossing Human Recombinant | 12,5 μL voor 50 ml media |
| 2-Mercaptoethanol 50 ml (1000x) | 17,5 μL voor 50 ml media |
Tabel 2: Differentiatiemedium (DM). De tabel geeft een overzicht van de reagentia die nodig zijn om het differentiatiemedium te bereiden.
Aanvullende video 1. Live beeldvorming van geïnduceerde hNEct 2D sheet/kolonie conversie naar 3D onder de behandeling van bFGF. Geïnduceerde kolonies hNEct werden voorzichtig losgemaakt van de schaal met dispase zoals hierboven beschreven en overgebracht naar een 6-well kweekplaat met lage bevestiging. 2D hNEct kolonies werden binnen 12 uur omgezet in 3D hNEct sferoïden. Seriebeelden werden elke 5 minuten vastgelegd. Schaalbalk = 100 μm. Klik hier om deze video te downloaden.
Om het gebruik van hPSC-afgeleide hersenorganoïden bij medicijnscreening en ziektemodellering mogelijk te maken, is het cruciaal om organoïden te maken volgens een repliceerbaar en betrouwbaar protocol15. Hersenorganoïden worden vaak gegenereerd uit embryoïde lichamen afgeleid van hPSC's, die vervolgens worden ingebed in een extracellulaire matrix die weefselexpansie en neurale differentiatie bevordert. In vergelijking met protocollen zoals Lancaster's1,16,17 en Velasco18, die beginnen bij embryoïde lichamen en het mogelijk maken dat een standaard differentiatieroute wordt gevolgd door de zich ontwikkelende organoïden, hebben we ontdekt dat het beginnen met corticale hersenorganoïde creatie met menselijke NEct-cellen in plaats van met embryoïde lichamen de consistentie van corticale hersenorganoïde vorming verbetert. Dit maakt dus ook de schaalvergroting mogelijk die nodig is voor geneesmiddelen- en fenotypische screening. Omdat menselijke NEct-cellen niet alleen in aanzienlijke hoeveelheden kunnen worden uitgebreid, maar ook gemakkelijk kunnen worden gecryopreserveerd, verbetert deze aanpak ook de repliceerbaarheid tussen experimenten. Er moet ook worden opgemerkt dat, in vergelijking met andere protocollen die het gebruik van bioreactoren en soortgelijke technologieën hebben aangenomen, er geen gespecialiseerde apparatuur nodig is voor dit protocol, waardoor het geschikt is voor elk laboratorium6. Ten slotte wordt de tijd die nodig is om volwassen organoïden te genereren die positief zijn voor corticale laagmarkers zoals SATB2 verminderd in vergelijking met zowel Lancaster1 als bioreactorprotocollen 6,19, waardoor het meer geschikt is voor het bestuderen van het ontwikkelingstraject van menselijke corticale ontwikkeling in gezondheid en ziekten 1,6,16.
Bovendien, gezien de voortdurend groeiende wereldwijde gezondheidszorgimpact van verouderingsgerelateerde ziekten zoals dementie, die geassocieerd zijn met een toename van senescente celtypen in de hersenen die bijdragen aan pathogenese, is het vermogen om verbindingen te identificeren en te testen die hersenveroudering kunnen verbeteren van enorm belang. Ondanks dat bekend is dat hPSC's epigenetisch verjongd zijn tijdens het herprogrammeringsproces20, vinden we robuuste toenames van senescente cellen in corticale hersenorganoïden die gedurende langere tijd worden gekweekt. Dit is een veelbelovende ontwikkeling die nu de screening mogelijk maakt van geneesmiddelen die dergelijke senescente cellen uit de hersenen elimineren (senolytica) of die dit proces vertragen (senostatica)21. Aangezien menselijke NEct-afgeleide corticale hersenorganoïden van menselijke oorsprong zijn, zal deze benadering waarschijnlijk het traditionele pad verkorten om dergelijke nieuwe therapieën op de markt te brengen.
Er zijn twee kritieke stappen in dit protocol. De eerste is het juiste niveau van samenvloeiing van de hPSC-kolonies op het moment van differentiatie. hPSC-kolonies moeten maximaal 30% samenvloeiend zijn om ervoor te zorgen dat gegenereerde NEct-kolonies niet fuseren met naburige kolonies en dat individuele organoïden clonaal worden aangedreven. De tweede kritieke stap omvat het juiste gebruik van dispase om de NEct-kolonies op te tillen en de neurale sferoïden te produceren. De timing van incubatie met dispase is van cruciaal belang voor de uiteindelijke kwaliteit van de gegenereerde neurale sferoïden. Dit komt omdat overmatige blootstelling van kolonies met dispase giftig is voor de cellen22 en uiteindelijk de kwaliteit van gegenereerde organoïden beïnvloedt. De beperking van dit protocol is dat het moeilijk is om de grootte van de neurale sferoïden te controleren, omdat het afhankelijk is van de grootte van de initiële kolonies die met dispase worden opgeheven. Dit probleem kan echter worden opgelost door neurale sferoïden te selecteren die van vergelijkbare grootte zijn wanneer u doorgaat naar de insluitfase.
Ten slotte kunnen toekomstige toepassingen zich uitstrekken tot het gebruik van deze reproduceerbare corticale organoïden in robotanalyse en biofarmaceutische screeningbenaderingen die doorgaans in die industrie worden gebruikt. Dit wordt ondersteund door voorlopige gegevens van ons laboratorium die aangeven dat de generatie van corticale hersenorganoïden uit menselijke NEct-cellen gemakkelijk kan worden geautomatiseerd, waardoor het compatibel is met deze benaderingen.
De auteurs hebben niets te onthullen.
Dit werk wordt ondersteund door het Medical Research Future Fund-Accelerated Research, Leukodystrophy vlaggenschip Massimo's Mission (EPCD000034), Medical Research Future Fund-Stem Cell Mission (APP2007653). Auteurs willen Dr. Ju-Hyun Lee (Korea University) bedanken voor het genereren van gegevens in Aanvullende Video 1.
| Name | Company | Catalog Number | Comments |
| 16% Formaldehyde (W/V) Methanol-free | Thermo Fisher Scientific | 28908 | 4% of PFA are diluted in 1x PBS |
| 2-Mercaptoethanol 50 mL(1000x) | Life Technologies Australia (TFS) | 21985023 | Used in NM and DM media |
| B 27 Supplement 10 mL | Life Technologies Australia (TFS) | 17504044 | Used in NM and DM media |
| CKX53 microscope with SC50 camera | Olympus | ||
| Corning Costar 6 well cell culture plates | Sigma Aldrich Pty Ltd | CLS3516-50EA | |
| Dispase II powder | Thermo Fisher Scientific | 17105041 | Powder is dissolve in HBSS, filtered through 0.22 µm filter, aliquote at 10 mL and store at -20 °C |
| DMEM Nutrient Mix F12 10x 500 mL (DMEM/F-12) | Thermofisher | 11320082 | Used in NM and DM media |
| DMSO Dimethyl Sulfoxide | Sigma Aldrich Pty Ltd | D2650-100ML | |
| Dulbecco's Phosphate Buffered Saline | Sigma Aldrich Pty Ltd | D1408-500ML | |
| Falcon Matrigel hESC-qualified Matrix | In Vitro Technologies Pty Ltd | FAL354277 | Make aliquotes of 100 µL and stored at -20 °C |
| GlutaMAX Supplement 100x | Thermo Fisher Scientific | 35050061 | Used in NM and DM media |
| Hanks Balanced Salt Solution | Sigma Aldrich Pty Ltd | H8264 | |
| Human induced pluripotent stem cells (EU79) | In-house reporogrammed from skin fibroblast | ||
| Human induced pluripotent stem cells (G22) | Genea Biocells | Obtained from Genea Biocells (San Diego, United States) | |
| Human induced pluripotent stem cells (WTC) | Gift from Professor Bruce Conklin | ||
| InSolution TGF-Β RI Kinase Inhibitor VI, SB431542 | Merck | US1616464-5MG | |
| Insulin Solution Human Recombinant | Sigma Aldrich Pty Ltd | I9278 | Used in NM and DM media |
| LDN193189 Dihydrochloride | Sigma Aldrich Pty Ltd | SML0559-5MG | Used during differentiation |
| MEM Non-Essential Amino Acids Solution (100x) | Thermo Fisher Scientific | 11140050 | Used in NM and DM media |
| mTeSR Plus | STEMCELL TECHNOLOGIES | 100-0276 | Used to maintain hiPSC colonies prior to differentiation with NM media |
| N2 Supplement 5 mL (100x) | Life Technologies Australia Pty Ltd | 17502048 | Used in NM and DM media |
| Neurobasal Medium | Thermo Fisher Scientific | 21103049 | Used in DM media |
| OCT Embedding Compound Sakura Clear (118 mL/Bottle) | Tissue Tek | 4583 | |
| Parafilm M Roll Size 4 in. x 125 Ft | Sigma Aldrich Pty Ltd | P7793 | |
| Penicillin-Streptomycin (10,000 U/mL) | Thermo Fisher Scientific | 15140122 | Used in NM and DM media |
| Potassium Hexacyanoferrate (II) Trihydrate | Sigma Aldrich Pty Ltd | CP1087 | |
| Potassium hexacyanoferrate(III) | Sigma Aldrich Pty Ltd | 455946 | |
| Prolong Glass Antifade Mountant | Life Technologies Australia (TFS) | P36980 | |
| Recombinant Human FGF basic | R&D Systems | 233-FB-01M | Aliquotes are made at 20 µg/mL and stored at -20 °C |
| SB431542 | Tocris | 1614 | Used during differentiation |
| Sucrose | Sigma Aldrich Pty Ltd | PHR1001-1G | 30% of sucrose are diluted in 1x PBS |
| Ultra-Low attachment multiwell plates , 24 well plate, polystyrene | Sigma Aldrich Pty Ltd | CLS3473-24EA | |
| X-GAL EA | Life Technologies Australia (TFS) | R0404 | Make aliquotes of 20 mg/mL and storde at -80 °C |
- Lancaster, M. A., Knoblich, J. A. Generation of cerebral organoids from human pluripotent stem cells. Nature Protocols. 9 (10), 2329-2340 (2014).
- Mansour, A. A., et al. An in vivo model of functional and vascularized human brain organoids. Nature Biotechnology. 36 (5), 432-441 (2018).
- Xiang, Y., et al. Fusion of regionally specified hPSC-derived organoids models human brain development and interneuron migration. Cell Stem Cell. 21 (3), 383-398 (2017).
- Bagley, J. A., Reumann, D., Bian, S., Lévi-Strauss, J., Knoblich, J. A. Fused cerebral organoids model interactions between brain regions. Nature Methods. 14 (7), 743 (2017).
- Kwak, T. H., et al. Generation of homogeneous midbrain organoids with in vivo-like cellular composition facilitates neurotoxin-based Parkinson's disease modeling. Stem Cells. 38 (6), 727-740 (2020).
- Qian, X., et al. Generation of human brain region-specific organoids using a miniaturized spinning bioreactor. Nature Protocols. 13 (3), 565-580 (2018).
- Muguruma, K., Nishiyama, A., Kawakami, H., Hashimoto, K., Sasai, Y. Self-organization of polarized cerebellar tissue in 3D culture of human pluripotent stem cells. Cell Reports. 10 (4), 537-550 (2015).
- Shaker, M. R., Cooper-White, J., Wolvetang, E. J. Self-organizing 3D human choroid plexus-ventricle-cortical organoids. BioRxiv. , (2020).
- Shaker, M. R., Aguado, J., Chaggar, H. K., Wolvetang, E. J. Klotho inhibits neuronal senescence in human brain organoids. npj Aging and Mechanisms of Disease. 7 (1), 1-12 (2021).
- Shaker, M. R., et al. Anteroposterior Wnt-RA gradient defines adhesion and migration properties of neural progenitors in developing spinal cord. Stem Cell Reports. 15 (4), 898-911 (2020).
- Chambers, S. M., et al. Highly efficient neural conversion of human ES and iPS cells by dual inhibition of SMAD signaling. Nature Biotechnology. 27 (3), 275-280 (2009).
- Shaker, M. R., et al. Rapid and efficient generation of myelinating human oligodendrocytes in organoids. Frontiers in Cellular Neuroscience. 15, 631548 (2021).
- Lee, J. -. H., Shaker, M. R., Lee, E., Lee, B., Sun, W. NeuroCore formation during differentiation of neurospheres of mouse embryonic neural stem cells. Stem Cell Research. 34, 101691 (2020).
- Shaker, M. R., et al. Spatiotemporal contribution of neuromesodermal progenitor-derived neural cells in the elongation of developing mouse spinal cord. Life Sciences. 282, 119393 (2021).
- Shaker, M. R., et al. Neural epidermal growth factor-like like protein 2 Is expressed in human oligodendroglial cell types. Frontiers in Cell and Developmental Biology. 10, 803061 (2022).
- Lancaster, M. A., et al. Cerebral organoids model human brain development and microcephaly. Nature. 501 (7467), 373-379 (2013).
- Giandomenico, S. L., Sutcliffe, M., Lancaster, M. A. Generation and long-term culture of advanced cerebral organoids for studying later stages of neural development. Nature Protocols. 16 (2), 579-602 (2021).
- Velasco, S., et al. Individual brain organoids reproducibly form cell diversity of the human cerebral cortex. Nature. 570 (7762), 523-527 (2019).
- Qian, X., et al. Brain-region-specific organoids using mini-bioreactors for modeling ZIKV exposure. Cell. 165 (5), 1238-1254 (2016).
- Hunter, Z. L., Leeson, H. C., Shaker, M. R., Wolvetang, E. J., Vadlamudi, L. Human induced pluripotent stem cells generated from epilepsy patients for use as in vitro models for drug screening. Stem Cell Research. 60, 102673 (2022).
- Kaur, A., Macip, S., Stover, C. M. An appraisal on the value of using nutraceutical based senolytics and senostatics in aging. Frontiers in Cell and Developmental Biology. 8, 218 (2020).
- Wang, F., et al. Safety and efficacy of dispase and plasmin in pharmacologic vitreolysis. Investigative Ophthalmology & Visual Science. 45 (9), 3286-3290 (2004).
ABOUT JoVE
Copyright © 2024 MyJoVE Corporation. All rights reserved