
Summary
Abstract
Introduction
Protocol
Representative Results
Discussion
Acknowledgements
Materials
References
Neuroscience
インビトロでの脳ニューロン老化のモデリングのための皮質脳オルガノイドの堅牢で再現性の高い生成
本研究では、標準的なフィーダーフリーhPSC培養を用いた、シンプルでありながら堅牢な皮質オルガノイド培養システムのための詳細な技術を提供する。これは、 インビトロで脳老化の側面をモデル化するオルガノイドを生成するための迅速で効率的で再現可能なプロトコルです。
脳オルガノイドは、発達中のヒト脳の3次元モデルであり、疾患モデリング、大規模なゲノムおよび薬物スクリーニングのための説得力のある最先端のプラットフォームを提供します。脳オルガノイド中の細胞の自己組織化の性質と、それらの生成に利用可能なプロトコルの範囲が拡大しているため、オルガノイド間の不均一性および変動性の問題が同定されている。このプロトコルペーパーでは、これらの問題を大部分克服し、1ヶ月以内に神経外胚葉前駆細胞から皮質オルガノイドを生成し、1年以上維持することができる堅牢で複製可能なプロトコルについて説明します。この再現性の高いプロトコールは、標準的な組織培養室で容易に実施することができ、その結果、発達中のヒト皮質に典型的に見られる豊富な多様性を有するオルガノイドが得られる。初期の発達構成にもかかわらず、ニューロンおよび他のヒト脳細胞型は、長期間のイン ビトロ 培養後にニューロン細胞において老化の典型的な徴候を示し始め、老化関連ニューロンプロセスを研究するための貴重で有用なプラットフォームとなる。このプロトコルはまた、老化関連β−ガラクトシダーゼ染色を用いて皮質脳オルガノイド中のこのような老化細胞を検出するための方法を概説する。
人間の脳に関する私たちの現在の知識は、主に動物モデルと死後の脳標本に基づいています。幹細胞生物学は、人間の脳発達の基本的な生物学と人間の脳障害の病理学的要因に新しい洞察を提供する急速に進歩している分野です。ヒト多能性幹細胞(hPSC)は、オルガノイド、臓器様三次元(3D)組織の生成 を介して ヒト脳をモデル化するための非常に貴重なツールであり、典型的には発達軌道、細胞構成、および発達中のヒト脳のアーキテクチャを反復する。脳オルガノイドは自己組織化されており、神経幹細胞、特定の神経前駆細胞、成熟ニューロン、およびグリア細胞型で構成されています。したがって、オルガノイドは、初期のヒト脳を研究するユニークな機会を提供し、これはしばしば直接的な実験にはアクセスできないが、血管系や免疫系の欠如などの固有の制限も有する。
脳オルガノイドを生成するための方法論は、2つの異なる方法で追求されてきた:誘導されていない分化と誘導された分化。誘導されない脳オルガノイド法は、組織形態形成1,2を駆動し、前脳、中脳、および後脳から脈絡叢、網膜、および中胚葉に至るまでの様々な細胞系譜アイデンティティの出現を可能にする幹細胞の自発的内因性分化能力に依存する。対照的に、誘導脳オルガノイド法は、内側神経節の卓越性3、前脳4、中脳5、視床下部6、小脳7、および脈絡叢8などの1つの脳領域タイプを表すニューロン系統の所望のパターン化に向けてhPSCを駆動するために外的要因の実質的な使用を必要とする。異なる細胞系譜を持つ異なる脳領域を生成する能力と、これらを自由に融合させる可能性により、脳オルガノイドは、人間の脳の発達を調査し、脳関連疾患の根底にあるメカニズムを解読するための優れたモデルとなっています。脳オルガノイドを生成するためのこれらの方法は、ヒトの脳領域をモデル化する上で画期的なものを提供するが、オルガノイド間の変動性および不均一性は、薬物スクリーニングなどの系統的および定量的研究にとって依然として重要な制限である。
現在のプロトコルは、我々の最近の論文9 で開発された方法に基づいており、二重SMAD阻害剤(SB-431542およびLDN 193189)による神経外胚葉(NEct)同一性へのhPSCコロニーの選択的分化を含み、FGF2シグナル伝達の影響下で4日以内に3D神経上皮スフェロイドに自己組織化する能力を有する。これらの神経上皮スフェロイドは、分化から4週間以内 にin vivo様細胞組成を有する均質な皮質オルガノイドを確実に生成する。ここで説明するプロトコルは、二重SMAD(Suppressor of Mothers Against Decapentaplegic)シグナル伝達の阻害が、とりわけ内胚葉系、中胚葉系、および対流胚葉系細胞運命の選択を阻害することによって、神経外胚葉前駆細胞10 に由来する吻側神経幹細胞へのhPSCの分化を促進することを示す我々の以前の知見に基づいている11.さらに、hESC修飾基底膜マトリックスにおける神経上皮スフェロイドの埋め込みは、神経上皮の有意な出芽を誘発し、アピコ基底極性を有する心室を形成する。大規模培養は、細胞株、クローン、またはバッチに依存しない皮質オルガノイドの再現性と均質性を示し、したがって、 in vitroでの健康および疾患における初期のヒト皮質発達を模倣する信頼性が高く安定した幹細胞系を表す。我々はさらに、長期間培養されたhPSC由来皮質脳オルガノイド中の老化性ニューロン細胞マーカーを検出するためのプロトコルを概説する。
hPSCを20%〜30%の播種密度でメッキした後、細胞を二重SMAD阻害剤で3日間処理し、hPSCコロニーを神経外胚葉性コロニーに分化させる。次いで、これらのコロニーをディスパーゼで穏やかに持ち上げ、FGF2を添加した超低付着6ウェルプレートに播種する。浮遊2Dコロニーは、一晩で3D神経外胚葉性スフェロイドに自己組織化され、FGF2を毎日補充したN2培地中で4日間維持される。スフェロイドが神経上皮層を確立したら、基底膜マトリックスに埋め込むことができる。新鮮な終末分化培地を日常的に添加することにより、研究者らは皮質オルガノイドにおける神経上皮の進行性拡張および出芽を観察する。研究者は、転写プロファイリングおよびプロテオミクスプロファイリングを行うために、これらのオルガノイドを解離させたいと思うかもしれません。さらに、明視野イメージングは、皮質オルガノイドの品質を監視するために推奨される。分析は、固定、凍結切除、および免疫染色によって行うことができる。これらの技術に関する説明および方法は、以前に12に記載されてきた。最終的に、このプロトコルにより、研究者は、低コストで限られた機器で発達中のヒト皮質脳をモデル化し、細胞ニューロン老化の側面を研究するための均質な皮質脳オルガノイドを迅速かつ堅牢に生成することができます。
1. 皮質脳オルガノイド生成
メモ:プロトコルのこのセクションのすべての手順は、特に明記されていない限り、クラス2バイオセーフティフードで行われます。
- hPSC 2D培養からの2D神経外胚葉性コロニーの誘導(-1~3日目)
- 誘導前に、hPSCコロニーを6ウェルプレートのhESC修飾基底膜マトリックス上に20%〜30%の密度でプレートします。この密度を達成するには、hPSCコロニーを6ウェルプレートの1ウェルから6ウェルプレートの3ウェルに60%コンフルエントで継代します。
- 基底膜マトリックスコーティングのために、基底膜マトリックスを平らな基礎培地中で1:50の比率で希釈する。6ウェルプレートに1mL/ウェルを均等に堆積させ、室温(RT)で1時間インキュベートした後、吸引する。
- 分化前にhPSCコロニーを無血清細胞培養培地2mL中で1日間維持する。
- hPSC分化の日に、4倍から10倍の倍率で明視野顕微鏡を用いてhPSCコロニーを検査し、検出可能な分化のない健康なコロニーを確実にする。
注:健康なhPSCは、大きな核、非常に小さな細胞質、および顕著な核小胞を有する細胞とタイトなエッジコロニーを形成する。分化したiPSCコロニーは、特にコロニーの外縁の周囲または中心において、上記に記載されたhPSCコロニーのそれと明確な形態学的相違を示すであろう。 - 表1に列挙した試薬を加えて、分化に必要なN2培地を構成する。使用前にこのメディアをRTに持参してください。
- RTに達したら、6ウェルプレートの各ウェルから無血清細胞培養培地を吸引し、5mL血清学的ピペットで穏やかに添加した2mLのN2培地と交換する。
- デュアルSMAD阻害剤SB-431542(10 μM)およびLDN 193189(100 nM)を追加します。
注:SMAD阻害剤は、培地が各ウェルに入れられた後、または無血清細胞培養培地を交換する前に必要量のN2培地に添加することができる。インヒビターは、プレートを穏やかに旋回させるか、培地およびインヒビターを含むチューブを3〜4回反転させることによって、培地に均一に組み込むことができる。 - SB-431542(10 μM)およびLDN 193189(100 nM)を添加した新鮮なN2培地を毎日各ウェルに添加し、その後2日間投与します。
注:SB-431542およびLDN 193189化合物を溶解し、細胞毒性を予防するために使用されるDMSOへの細胞の長期曝露を低減するために、新鮮なN2培地を添加する。
- 誘導された2D神経外胚葉性コロニーからの3D神経外胚葉性スフェロイドの生成(3~7日目)
- 誘導された神経外胚葉性コロニーを、工程1.2.2〜1.2.8に従ってディスパーゼを用いて持ち上げる。
- まず、6ウェルプレートから2mLのN2培地を取り出し、HBSSで1xを洗浄して、N2培地がすべて除去されたことを確認します。
注:N2培地は、ディスパーゼの酵素活性を妨げ、神経外胚葉性コロニーのウェルからの適切な剥離を妨げる可能性がある。 - 各コロニー含有ウェルに1 mLの2.4単位/mLディスパーゼを加える。
- ウェルを37°Cで20〜25分(最大30分)インキュベートする。 コロニーの剥離を定期的にチェックしてください。
注:小さなコロニーは20分以内に剥離することがあります。30分後に立ち往生したままのコロニーは無視する必要があります。 - インキュベーション後、ウェルに1 mLのN2培地を加えてディスパーゼ酵素の活性を停止し、広口径のP1000ピペットチップまたは滅菌はさみで切断した修正P1000ピペットチップを使用してコロニーを15 mLチューブに移します(広口径のP1000チップにします)。
- コロニーの塊が重力でチューブの底に沈むのを待ちます。
注: このプロセスには約 1 分かかります。 - 凝集塊が沈んだら、標準のP1000ピペットチップで上清を慎重に取り除き、1mLの新鮮なN2培地と交換します。この洗浄ステップを3回繰り返して、ディスパーゼを完全に除去します。
注:残りのディスパーゼは、神経外胚葉性スフェロイドの均一な形成を防ぎ、細胞死を誘導する。 - 洗浄後、細胞集塊を3 mLのN2培地に再懸濁し、6ウェルプレートの1ウェルに移し、40 ng/mLのbFGFを加えます。
注:多数の神経外胚葉性コロニーが剥離した場合、これらのコロニーを6ウェルプレートの2つ以上のウェルにまたがって播種し、スフェロイドの融合を防ぐことができます。コロニーの剥離から24時間後に、スフェロイドが形成されたかどうかを確認する。 - スフェロイドを次の3〜4日間同じ培地に維持しますが、毎日新鮮なbFGF(40ng/mL)を各ウェルに加え、神経外胚葉系細胞の増殖を促進し、自己組織化し、神経上皮を誘導および拡張します。
注:スフェロイドをより高い密度でメッキした場合、培地は黄色に変わる可能性があり、2日ごとに新鮮なbFGF(40ng/mL)と交換する必要があります。しかし、これは推奨されず、代わりに、この問題を避けるために、各ウェルでより少ない数の回転楕円体を維持する必要があります。スフェロイドは、神経上皮が明らかであれば、3日後に基底膜マトリックスに埋め込むことができる。神経上皮が明らかでない、または強く見えない場合は、回転楕円体を別の日維持し、もう一度確認してください。
- 皮質脳オルガノイドの分化と維持(8日目)
- 表2に列挙した試薬を用いて末端分化培地(DM)を調製した。このメディアをRTに持参してください。
- 氷上で100%hESC修飾基底膜マトリックスを解凍する。
- 10cmのシャーレに70%エタノールで滅菌したディンプル付きのパラフィルムを用意し、フードの下の実体顕微鏡の上に置きます。
メモ: 200 μL のピペットチップの空のトレイを使用して、ディンプルのグリッドを生成できます。 - 100 μL のピペットチップの端を切り取ります (ワイドボアにするため)。これは、ニューラルスフェロイドを壊さずに地下室行列に埋め込むために使用されます。
- 実体顕微鏡を使用して、6ウェルプレートから類似サイズの神経スフェロイド(500 μm)を選択し、100 μLの広口径ピペットチップを使用してパラフィルムディンプルに移し、ディンプルごとに単一の神経スフェロイドを配置します。
- 余分なメディアをそっと取り除き、回転楕円体を覆うのに十分な量を残します。
注: これは、基底マトリックスを追加する前に回転楕円体の品質を維持し、それらが乾燥しないようにするためです。 - スフェロイドの上に18μLの基底マトリックスを静かに加え、スフェロイドをマトリックスドロップの中心内に配置します。10 μL のピペットチップの端を使用して、スフェロイドをマトリックスの中央に配置します。
注: スフェロイドの周囲の余分なメディアが乾燥しないように、できるだけ早く地下室マトリックスを追加してください。 - 10cmのシャーレを覆い、インキュベーターに移し、地下マトリックス埋め込みスフェロイドと共にパラフィルム皿を37°Cで25分間インキュベートする。
- インキュベーション後、0.5 mLのDM培地を含むP1000チップを使用して、埋め込みスフェロイドを低付着24ウェルプレートに洗い流し、個々の埋め込みスフェロイドがそれぞれ1つのウェルに配置されていることを確認します。
メモ: 複数の埋め込み回転楕円体が 1 つのウェルに落ちた場合は、穴の広い P1000 チップを使用して、もう一方の回転楕円体を新しいウェルに移します。 - 分化した皮質脳オルガノイドをDM培地に長期間維持し、オルガノイドが大きくて古くなると2日ごとに培地の変化が起こる。
注:分化の最初の1週間の間、培地は3日ごとに交換することができる。
2. 皮質オルガノイドにおける神経細胞老化の特性評価
- 凍結切片のためのプロセス皮質オルガノイド:
注:手順は、クラス2バイオセーフティフードで実行しました。- それぞれ1.5 mLの4%パラホルムアルデヒド(PFA)で満たされた2 mLチューブを用意する。
- P1000ピペットチップの端をカットし(広口径にするため)、各オルガノイドを上記で調製した2 mLチューブの1つ(チューブあたり1つのオルガノイド)に静かに移します。
メモ: 過剰な DM 媒体が PFA と混ざらないようにするには、オルガノイドを P1000 ピペットチップの開口部に向かって沈み込ませ、オルガノイドをピペッティングする前にチューブ内の PFA の上部のすぐ上にチップを載せます。これにより、研究者はオルガノイドと非常に最小限の媒体のみを転送することができます。 - 固定プロセスを4°Cで1時間行います。
- 未カットのP1000ピペットチップを使用して、余分なPFAを慎重に吸引し、1.5mLの冷たい1x PBSを加えます。
- チューブを70rpmに設定したオービタルシェーカーにRTで10分間移します。
- 冷たい1x PBSによる洗浄プロセスを3回繰り返して、すべてのPFAが完全に除去されていることを確認します。
注:PFAを通常の廃棄物容器に廃棄しないでください。代わりに、PFAは危険であるため、このために特定の化学廃棄物処理容器を準備してください。 - オルガノイドを30%スクロースを含む1x PBSに浸漬し、すべてのオルガノイドがチューブの底に沈むまで4°Cでインキュベートする。
注:オルガノイドが沈むのに必要な時間は、オルガノイドのサイズ/年齢によって異なります。生後3ヶ月のオルガノイドは最大5時間かかることがあります。 - カットされた広口径のP1000ピペットチップを使用して、30%スクロースと100%最適切断温度(OCT)培地からなる取り付け溶液を含む取り付け金型に、3:2の比率で3〜5個のオルガノイドを静かに移します。
- 実体顕微鏡の助けを借りて10 μLのピペットチップを使用して、オルガノイドを格子状のパターンで配向させ、配置します。
- 金型をドライアイスの上に置き、スクロース OCT 溶液を固化してから、クライオスタットを使用してクライオセクショニング (16 ~ 20 μm) を進めます。
注:老化関連β-ガラクトシダーゼの場合、沈んだらすべての組織を切片化のために処理する必要があります。免疫蛍光の場合、組織は翌日に切片化するために処理することができる。切片を含むすべてのスライドは、直ちに染色しない場合は、その後の免疫蛍光またはβ-ガラクトシダーゼの前に-20°Cで保存する必要があります。
- 皮質脳オルガノイドにおける老化の分析のためのプロセス:
メモ: 次の手順は、通常のラボ ベンチで実行できます。- スライドを蓋付きの顕微鏡スライド染色容器に移し、切片化したオルガノイド組織を1x PBSで3回、RTで10分間洗浄し、余分な取り付け溶液を除去します。
- これに続いて、洗浄した組織を新たに作製したβ-ガラクトシダーゼ染色液と共に37°Cで一晩インキュベートする。
注:β-ガラクトシダーゼ染色溶液は、リン酸緩衝液(10 mLのリン酸緩衝液:8.15 mLの1 M NaH2PO4、1.85 mLの1M Na2 HPO4)で調整されたpH = 6、100 mMのヘキサシアノ鉄酸カリウム(III)、100 mMのヘキサシアノ鉄酸カリウム(II)三水和物、5 MのNaCl、1 MのMgCl2、 20 ミリグラム/mL の X-Gal.CO2がβ-ガラクトシダーゼ染色溶液のpHを変化させるので、CO2を含む標準的な細胞培養インキュベーターの使用を避ける。 - 染色した組織を1x PBSでRTでそれぞれ10分間3回洗浄し、β-ガラクトシダーゼ溶液を除去した。
- 洗浄した組織をガラスの退色防止マウント剤でマウントし、顕微鏡で見る前に取り付け溶液をRTで30分間固化させます。
ここでは、研究者が培養後1〜3ヶ月以内に in vivo ヒト皮質脳領域を模倣する均質なhPSC由来皮質脳オルガノイドを生成することを可能にする堅牢なプロトコルについて説明した。hPSCコロニーは、まず分化培地中で培養して神経外胚葉性コロニーを生成し、次いで神経スフェロイドを形成するために使用することができる。これらのスフェロイドは、その後、基底膜マトリックスに埋め込まれ、長期間維持されて、ニューロンの老化をモデル化するために使用できるオルガノイドを生成する(プロトコルの概要については 、図1 を参照)。これらのオルガノイドを超コーティングされていない24ウェルプレートで培養すると、細胞ストレスを引き起こし、13週間の in vitro培養で 老化関連表現型を促進することは注目に値します。このプロトコールに由来するオルガノイドは、皮質プレート神経細胞の最適な増殖および分化のために、または気液界面で、攪拌バイオリアクター内で維持することもできる。
まず、hPSCコロニーを神経外胚葉分化の前に1日間培養する。これらのhPSCコロニーは、わずか20%〜30%のコンフルエント度で培養され、可能な限り最高の品質、すなわちコロニーを汚染する分化細胞のないタイトな平坦な単層であることが重要です(図2A、B)。hPSCコロニーの多能性は、NANOGなどのマーカーの発現によって確認されるべきである(図2C)。次いで、検証されたhPSCコロニーを、SB-431542およびLDN 193189を有するN2神経外胚葉性分化培地に曝露する。この培地での3日間の維持の後、hPSCコロニーは神経外胚葉性コロニーに分化し、もはやhPSCの同じタイトな平坦な単層形態を示さず(図2B)、むしろ、それらはより長い柱状細胞になる(図2B)。これらの細胞はまた、NANOGなどの多能性マーカーに対して陰性である(図2C)。
神経外胚葉性コロニーが異種で酵素的に剥離され、健常で首尾よく剥離したコロニーが自己組織化され、若い神経スフェロイドを形成することができるのはこの段階である(図2D、補足ビデオ1)。健康で清潔な神経外胚葉性コロニーのみが、ディスパーゼ活性に指定された時間枠内に剥離する。他のすべてのコロニーは、スフェロイドの品質が低くなるため、無視する必要があります。N2培地中のFGF2に毎日曝露されると、これらのスフェロイド(図2D、1日目)の神経幹細胞(SOX2+)は増殖し、かなりの数の神経ロゼットを形成する(図2D、4日目)。これらのロゼットは、ロゼットの中心内および回転楕円体の外縁に沿って位置する細胞においてタイトジャンクションおよび上皮マーカーZO1を発現し、回転楕円体の頂端 - 基底極性を示す(図2D、4日目)。回転楕円体のホールマウント3Dイメージングの方法は、13の前に説明されている。スフェロイドの毎日の検査は、スフェロイドのタイトで暗い外縁および明るい周辺部の形成を解明するはずであり、これは神経上皮層である。この層は、3〜4日後におよそ500μmの直径で十分に形成されるべきであり、その時点で回転楕円体を基底マトリックスに埋め込むことができる。この層が存在しないか、または弱く形成されているだけの場合、スフェロイドは前進するのに十分に発達していない。変化を観察するために別の日を待つことをお勧めしますが、これが観察されない場合は、これらの回転楕円体を無視してください。
培養3日後のスフェロイドの代表的な明視野像を 図2Dに見ることができる。他の隣接するスフェロイドと融合しておらず、半透明の組織を有し、神経ロゼット形成を示す、タイトな神経上皮層を有するスフェロイドは、地下マトリックスに埋め込まれるように選択される。一旦埋め込まれると、回転楕円体は急速に増殖し、出芽を開始する:コンパクトな組織の節が現れ、回転楕円体の本体から外側に広がる。これは、地下マトリックス中で1〜3週間の間に明らかであり、複数の細胞株にわたって観察することができる(図3A)。埋め込まれたスフェロイドの定量分析により、3つの異なる細胞株にわたって最大100%のスフェロイドに上皮細胞が存在することを確認し、このプロトコルから期待される均質性と再現性を確認しました(図3B)。 in vitro 分化中のオルガノイド直径の定量化は、hPSCの異なるラインにわたる再現性をさらに確認する(図3C)。出芽が起こらなければ、回転楕円体は適切に発達しておらず、廃棄されるべきである。スフェロイドがマトリックスに埋め込まれると、その発達が進み、スフェロイドはオルガノイドと呼ばれるようになりました。免疫蛍光染色はまた、神経前駆細胞(PAX6)ならびにCTIP2およびSATB2で染色された皮質層マーカーの存在を、明確な層状を有するオルガノイド中に確認する(図3D、E)。この層状化は、オルガノイド維持の異なる時点にわたって観察可能である(図3D、E)。組織の免疫組織化学の方法は、前に14に記載されてきた。
これらのオルガノイドの1つの可能な用途は、ニューロンの老化関連プロセスが脳にどのように影響するかを研究することである。これを調べるために、首尾よく生成されたオルガノイドは、老化関連β−ガラクトシダーゼおよびp21などの老化の標準的な分子バイオマーカーの切片化および染色のために、複数の異なる時点から採取される。 図4Aは 、基底膜マトリックスに包埋してから4および13週間後のオルガノイドの老化関連β−ガラクトシダーゼ染色の代表的な画像を示す。4週目から13週目の間に、老化関連β-ガラクトシダーゼの存在が著しく増加しており、生物老化の認識されたドライバーである細胞老化が培養においてこの時間にわたって起こったことが示唆される。13週目におけるオルガノイドの免疫蛍光染色は、成熟皮質ニューロンマーカー(CTIP2)で共標識された別の老化マーカーp21の存在を確認し、 図4Bに見ることができる。しかしながら、p21の存在は細胞周期停止のマーカーであり、それ自体は老化の決定的なマーカーではなく、p16およびSASP(老化関連分泌表現型)因子などの他の老化マーカーの検出は、細胞を老化として決定的に同定するために推奨されることに留意すべきである。
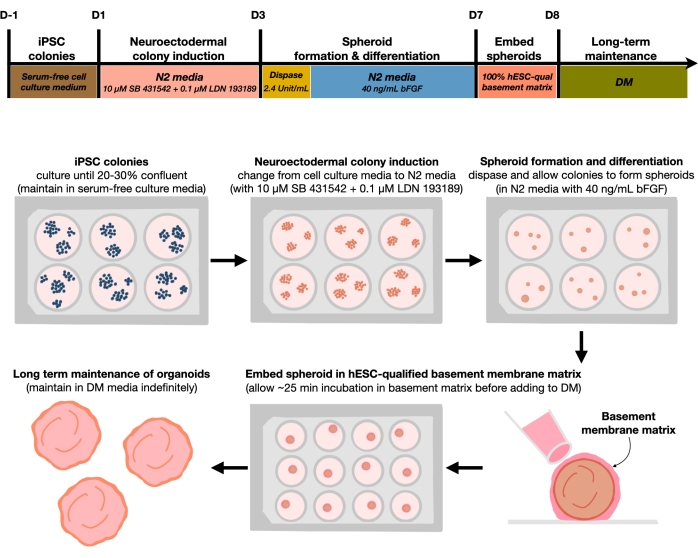
図1:再現可能な皮質脳オルガノイドを生成するための模式図。フィーダーフリー培地中に維持されたhPSCsからの皮質脳オルガノイドの生成のための実験手順の概略ワークフロー。このワークフローでは、2D hPSCをオルガノイド中の3Dパターン化された皮質プレートヒト組織に区別するために必要な6つのステップの概要を提供します。この図の拡大版を表示するには、ここをクリックしてください。
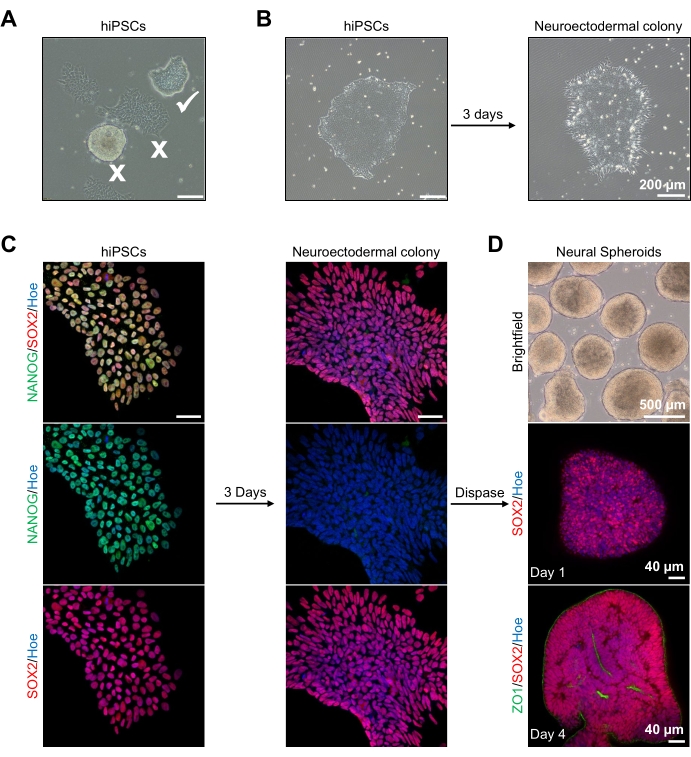
図2:神経外胚葉性コロニー-hPSCに由来する神経スフェロイドの生成 (A)最適(ホワイトダニ)および分化コロニー(白十字)を示すヒトPSCの代表的な画像。スケールバー:200μm、倍率4倍。(b)二重SMAD阻害剤治療の3日後のhPSCsに由来する神経外胚葉性コロニーの代表的な画像。スケールバー:200μm、倍率4倍。(c)ヒトPSCコロニーを神経外胚葉性コロニーに向けて分化させた。画像は、SOX2(赤色)、NANOG(緑色)でPSC(1日目)および神経外胚葉性(3日目)コロニーの染色を表し、全ての核はヘキスト33342(青色)で対比染色した。スケールバー:40μm、倍率100倍。(d)明視野下での インビトロ 培養における皮質脳スフェロイドの経時的な発達段階を示す画像、および1日目にSOX2(赤)で免疫染色されたホールマウント、および4日目にSOX2(赤)およびZO1(緑)で二重免疫染色した。明視野画像のスケールバーは500μm、倍率は4倍、下部画像のスケールバーは倍率40μm、倍率20倍である。 この図の拡大版を表示するには、ここをクリックしてください。
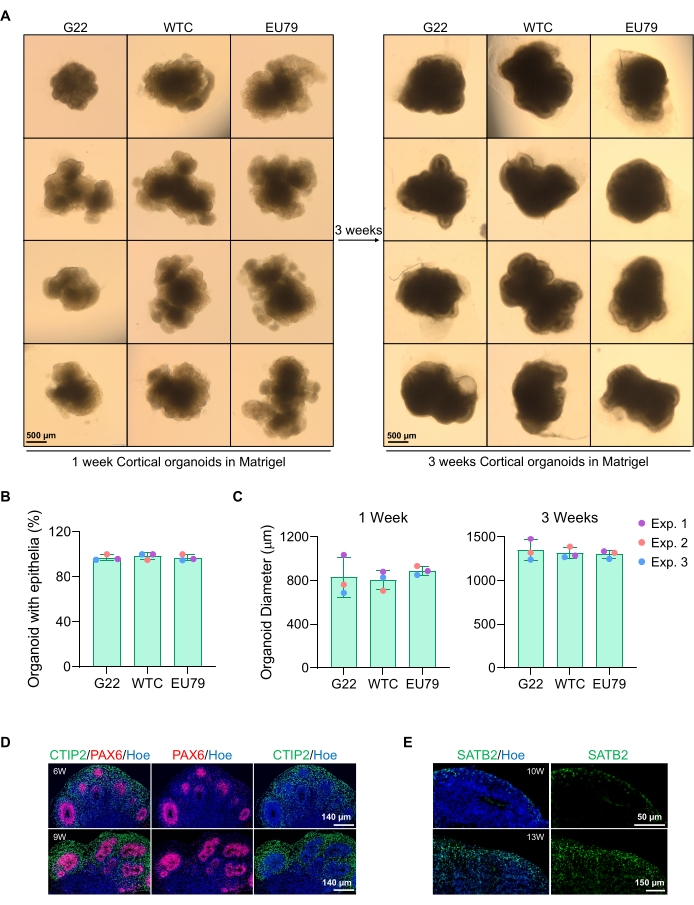
図3:異なるhPSCラインに由来する皮質脳オルガノイドの特性評価(A)インビトロで3週間にわたって培養されたG22、WTC、およびEU79ヒトiPSCラインに由来する皮質脳オルガノイドの代表的な画像。すべての画像のスケールバーは500μm、倍率2倍です。(B)異なるhPSC株(G22、WTC、およびEU79)における3週間のin vitro分化における皮質脳オルガノイドの生成の成功率。N = 3。データは平均±標準偏差として表示されます。(c)ヒト多能性幹細胞株(G22、WTC、およびEU79)の異なる株におけるインビトロ分化の1週目および3週目における皮質脳オルガノイドの成長(平均直径に基づく)を示す棒グラフ。N = 3。データは平均±標準偏差として表示されます。(d)心室帯PAX6(赤)および皮質プレートCTIP2(緑)について免疫染色されたG22 hPSCに由来する6週齢および9週齢の皮質脳オルガノイドの切片の代表的な画像。全ての切片をヘキスト33342(青色)で対比染色した。スケールバー= 140μm、倍率20倍。W は週です。(e)皮質層IV SATB2について免疫染色したWTC hPSCsに由来する10週齢および13週齢の皮質脳オルガノイドの切片の代表的な画像(緑色)。全ての切片をヘキスト33342(青色)で対比染色した。10週間画像スケールバー=50μm、倍率40倍。13週間画像スケールバー=150μm、倍率40倍。W は週です。この図の拡大版を表示するには、ここをクリックしてください。
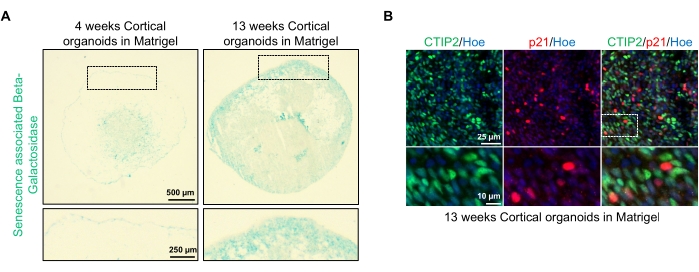
図4:hPSCsに由来する皮質脳オルガノイドにおける老化の特徴付け(A)インビトロで4週間および13週間培養し、SA-β-galで染色したWTC hPSCに由来するヒト皮質脳オルガノイドの切片の代表的な画像。スケールバー = 500 μm、ズーム画像のスケールバー = 250 μm、倍率4倍。点線のボックスは拡大画像を示します。(b)皮質ニューロンCTIP2(緑)およびp21(赤)について免疫染色されたヒトEU79 hPSCsに由来する13週齢の皮質脳オルガノイドの切片の代表的な画像。すべての切片をヘキスト33342(青)で対比染色した。スケールバー= 25μm、ズーム画像のスケールバー= 10μm、倍率40倍。この図の拡大版を表示するには、ここをクリックしてください。
| メディアコンポーネント | 濃度 |
| DMEM 栄養ミックス F12 10x 500 mL (DMEM/F-12) | |
| N2 サプリメント 5 mL (100x) | 1%で支持 |
| B 27 サプリメント 10 mL | 2%で補充 |
| MEM非必須アミノ酸溶液(100倍) | 1%で補充 |
| ペニシリン-ストレプトマイシン (10,000 U/mL) | 1%で補充 |
| 2-メルカプトエタノール 50mL(1000x) | 0.1%で補充 |
表1:N2ミディアムこの表は、N2培地を調製するために必要な試薬をリストしています。
| メディアコンポーネント | 濃度 |
| DMEM 栄養ミックス F12 10x 500 mL (DMEM/F-12) | DM培地は、DMEM/F12とニューロ基底培地の1:1の比率で作られています |
| 神経基底培地 | |
| N2 サプリメント 5 mL (100x) | 0.5%で維持 |
| B 27 サプリメント 10 mL | 1%で補充 |
| MEM非必須アミノ酸溶液(100倍) | 1%で補充 |
| グルタマックスサプリメント100倍 | 1%で補充 |
| ペニシリン-ストレプトマイシン (10,000 U/mL) | 1%で補充 |
| インスリン溶液ヒト組換え体 | 12.5 μL (50 mL の培地) |
| 2-メルカプトエタノール 50mL (1000x) | 17.5 μL (50 mL の培地) |
表2:分化培地(DM)。 表は、分化培地を調製するために必要な試薬を列挙する。
補足ビデオ 1.bFGFの治療下で誘導されたhNEct 2Dシート/コロニーの3Dへの変換のライブイメージング。hNEctの誘導コロニーを、上記で概説したようにディスパーゼでディッシュから穏やかに剥離し、低付着6ウェル培養プレートに移した。2D hNEctコロニーは、12時間以内に3D hNEctスフェロイドに変換された。シリアル画像は5分ごとにキャプチャされました。スケール バー = 100 μm。 このビデオをダウンロードするには、ここをクリックしてください。
薬物スクリーニングおよび疾患モデリングにおけるhPSC由来の脳オルガノイドの使用を可能にするためには、複製可能で信頼性の高いプロトコル15に従ってオルガノイドを製造することが極めて重要である。脳オルガノイドは、通常、hPSCに由来する胚様体から生成され、その後、組織拡張および神経分化を促進する細胞外マトリックスに埋め込まれる。ランカスターの1,16,17やベラスコ18のような、胚様体から始まり、発達中のオルガノイドが続くデフォルトの分化経路を可能にするプロトコルと比較すると、皮質脳オルガノイドの作成を胚様体ではなくヒトNEct細胞で開始すると、皮質脳オルガノイド形成の一貫性が向上することが見出された。これにより、薬物および表現型スクリーニングに必要なスケーリングも可能になります。ヒトNEct細胞は、かなりの量に拡大できるだけでなく、容易に凍結保存することができるので、このアプローチはまた、実験間の再現性を改善する。また、バイオリアクターおよび類似の技術の使用を採用した他のプロトコルと比較して、このプロトコルには特別な機器は必要なく、ラボ6に適しています。最後に、SATB2などの皮質層マーカーに陽性である成熟オルガノイドを生成するのに必要な時間は、ランカスター1およびバイオリアクタープロトコル6,19の両方と比較して短縮され、健康および疾患におけるヒト皮質発達の発達軌道を研究するのにより適している1,6,16。
さらに、病因に寄与する脳内の老化細胞型の増加に関連する認知症などの加齢関連疾患の世界的なヘルスケアへの影響が継続的に拡大していることを考えると、脳の老化を改善することができる化合物を同定および試験する能力は大きな関心事である。hPSCは、リプログラミングプロセス20の間にエピジェネティックに若返ることが知られているにもかかわらず、我々は、長期間培養された皮質脳オルガノイドにおける老化細胞の堅牢な増加を見いだした。これは、脳からそのような老化細胞を排除する薬物(血清溶解薬)またはこのプロセスを遅らせる薬物(静水薬)のスクリーニングを可能にする有望な開発です21。ヒトNEct由来皮質脳オルガノイドはヒト由来であるため、このアプローチは、このような新規治療薬を市場に出すための伝統的な道を短縮する可能性が高い。
このプロトコルには 2 つの重要なステップがあります。1つ目は、分化時のhPSCコロニーのコンフルエンシーの正しいレベルである。hPSCコロニーは、生成されたNEctコロニーが隣接するコロニーと融合せず、個々のオルガノイドがクローン駆動であることを保証するために、最大30%コンフルエントでなければならない。第2の重要なステップは、NEctコロニーを持ち上げ、神経スフェロイドを産生するためのdispaseの正しい使用を含む。dispaseとのインキュベーションのタイミングは、生成される神経スフェロイドの最終的な品質にとって重要です。これは、ディスパーゼによるコロニーの過剰曝露が細胞22 に対して毒性であり、最終的に生成されたオルガノイドの品質に影響を及ぼすからである。このプロトコルの制限は、異なる部分で持ち上げられる初期コロニーのサイズに依存するため、神経回転楕円体のサイズを制御することが困難であるということです。ただし、この問題は、埋め込み段階に進むときに同様のサイズのニューラルスフェロイドを選択することで克服できます。
最後に、将来の用途は、その業界で一般的に使用されるロボット分析およびバイオ医薬品スクリーニングアプローチにおけるこれらの再現可能な皮質オルガノイドの使用にまで及ぶ可能性がある。これは、ヒトNEct細胞からの皮質脳オルガノイドの生成を容易に自動化することができ、これらのアプローチと互換性があることを示す私たちの研究室からの予備データによって支持されています。
著者らには開示するものは何もありません。
この研究は、医学研究未来基金加速研究、白質ジストロフィーの旗艦マッシモのミッション(EPCD000034)、医学研究未来基金幹細胞ミッション(APP2007653)によって支援されています。著者らは、 補足ビデオ1でデータを生成してくれた李淳龍博士(高麗大学)に感謝したい。
| Name | Company | Catalog Number | Comments |
| 16% Formaldehyde (W/V) Methanol-free | Thermo Fisher Scientific | 28908 | 4% of PFA are diluted in 1x PBS |
| 2-Mercaptoethanol 50 mL(1000x) | Life Technologies Australia (TFS) | 21985023 | Used in NM and DM media |
| B 27 Supplement 10 mL | Life Technologies Australia (TFS) | 17504044 | Used in NM and DM media |
| CKX53 microscope with SC50 camera | Olympus | ||
| Corning Costar 6 well cell culture plates | Sigma Aldrich Pty Ltd | CLS3516-50EA | |
| Dispase II powder | Thermo Fisher Scientific | 17105041 | Powder is dissolve in HBSS, filtered through 0.22 µm filter, aliquote at 10 mL and store at -20 °C |
| DMEM Nutrient Mix F12 10x 500 mL (DMEM/F-12) | Thermofisher | 11320082 | Used in NM and DM media |
| DMSO Dimethyl Sulfoxide | Sigma Aldrich Pty Ltd | D2650-100ML | |
| Dulbecco's Phosphate Buffered Saline | Sigma Aldrich Pty Ltd | D1408-500ML | |
| Falcon Matrigel hESC-qualified Matrix | In Vitro Technologies Pty Ltd | FAL354277 | Make aliquotes of 100 µL and stored at -20 °C |
| GlutaMAX Supplement 100x | Thermo Fisher Scientific | 35050061 | Used in NM and DM media |
| Hanks Balanced Salt Solution | Sigma Aldrich Pty Ltd | H8264 | |
| Human induced pluripotent stem cells (EU79) | In-house reporogrammed from skin fibroblast | ||
| Human induced pluripotent stem cells (G22) | Genea Biocells | Obtained from Genea Biocells (San Diego, United States) | |
| Human induced pluripotent stem cells (WTC) | Gift from Professor Bruce Conklin | ||
| InSolution TGF-Β RI Kinase Inhibitor VI, SB431542 | Merck | US1616464-5MG | |
| Insulin Solution Human Recombinant | Sigma Aldrich Pty Ltd | I9278 | Used in NM and DM media |
| LDN193189 Dihydrochloride | Sigma Aldrich Pty Ltd | SML0559-5MG | Used during differentiation |
| MEM Non-Essential Amino Acids Solution (100x) | Thermo Fisher Scientific | 11140050 | Used in NM and DM media |
| mTeSR Plus | STEMCELL TECHNOLOGIES | 100-0276 | Used to maintain hiPSC colonies prior to differentiation with NM media |
| N2 Supplement 5 mL (100x) | Life Technologies Australia Pty Ltd | 17502048 | Used in NM and DM media |
| Neurobasal Medium | Thermo Fisher Scientific | 21103049 | Used in DM media |
| OCT Embedding Compound Sakura Clear (118 mL/Bottle) | Tissue Tek | 4583 | |
| Parafilm M Roll Size 4 in. x 125 Ft | Sigma Aldrich Pty Ltd | P7793 | |
| Penicillin-Streptomycin (10,000 U/mL) | Thermo Fisher Scientific | 15140122 | Used in NM and DM media |
| Potassium Hexacyanoferrate (II) Trihydrate | Sigma Aldrich Pty Ltd | CP1087 | |
| Potassium hexacyanoferrate(III) | Sigma Aldrich Pty Ltd | 455946 | |
| Prolong Glass Antifade Mountant | Life Technologies Australia (TFS) | P36980 | |
| Recombinant Human FGF basic | R&D Systems | 233-FB-01M | Aliquotes are made at 20 µg/mL and stored at -20 °C |
| SB431542 | Tocris | 1614 | Used during differentiation |
| Sucrose | Sigma Aldrich Pty Ltd | PHR1001-1G | 30% of sucrose are diluted in 1x PBS |
| Ultra-Low attachment multiwell plates , 24 well plate, polystyrene | Sigma Aldrich Pty Ltd | CLS3473-24EA | |
| X-GAL EA | Life Technologies Australia (TFS) | R0404 | Make aliquotes of 20 mg/mL and storde at -80 °C |
- Lancaster, M. A., Knoblich, J. A. Generation of cerebral organoids from human pluripotent stem cells. Nature Protocols. 9 (10), 2329-2340 (2014).
- Mansour, A. A., et al. An in vivo model of functional and vascularized human brain organoids. Nature Biotechnology. 36 (5), 432-441 (2018).
- Xiang, Y., et al. Fusion of regionally specified hPSC-derived organoids models human brain development and interneuron migration. Cell Stem Cell. 21 (3), 383-398 (2017).
- Bagley, J. A., Reumann, D., Bian, S., Lévi-Strauss, J., Knoblich, J. A. Fused cerebral organoids model interactions between brain regions. Nature Methods. 14 (7), 743 (2017).
- Kwak, T. H., et al. Generation of homogeneous midbrain organoids with in vivo-like cellular composition facilitates neurotoxin-based Parkinson's disease modeling. Stem Cells. 38 (6), 727-740 (2020).
- Qian, X., et al. Generation of human brain region-specific organoids using a miniaturized spinning bioreactor. Nature Protocols. 13 (3), 565-580 (2018).
- Muguruma, K., Nishiyama, A., Kawakami, H., Hashimoto, K., Sasai, Y. Self-organization of polarized cerebellar tissue in 3D culture of human pluripotent stem cells. Cell Reports. 10 (4), 537-550 (2015).
- Shaker, M. R., Cooper-White, J., Wolvetang, E. J. Self-organizing 3D human choroid plexus-ventricle-cortical organoids. BioRxiv. , (2020).
- Shaker, M. R., Aguado, J., Chaggar, H. K., Wolvetang, E. J. Klotho inhibits neuronal senescence in human brain organoids. npj Aging and Mechanisms of Disease. 7 (1), 1-12 (2021).
- Shaker, M. R., et al. Anteroposterior Wnt-RA gradient defines adhesion and migration properties of neural progenitors in developing spinal cord. Stem Cell Reports. 15 (4), 898-911 (2020).
- Chambers, S. M., et al. Highly efficient neural conversion of human ES and iPS cells by dual inhibition of SMAD signaling. Nature Biotechnology. 27 (3), 275-280 (2009).
- Shaker, M. R., et al. Rapid and efficient generation of myelinating human oligodendrocytes in organoids. Frontiers in Cellular Neuroscience. 15, 631548 (2021).
- Lee, J. -. H., Shaker, M. R., Lee, E., Lee, B., Sun, W. NeuroCore formation during differentiation of neurospheres of mouse embryonic neural stem cells. Stem Cell Research. 34, 101691 (2020).
- Shaker, M. R., et al. Spatiotemporal contribution of neuromesodermal progenitor-derived neural cells in the elongation of developing mouse spinal cord. Life Sciences. 282, 119393 (2021).
- Shaker, M. R., et al. Neural epidermal growth factor-like like protein 2 Is expressed in human oligodendroglial cell types. Frontiers in Cell and Developmental Biology. 10, 803061 (2022).
- Lancaster, M. A., et al. Cerebral organoids model human brain development and microcephaly. Nature. 501 (7467), 373-379 (2013).
- Giandomenico, S. L., Sutcliffe, M., Lancaster, M. A. Generation and long-term culture of advanced cerebral organoids for studying later stages of neural development. Nature Protocols. 16 (2), 579-602 (2021).
- Velasco, S., et al. Individual brain organoids reproducibly form cell diversity of the human cerebral cortex. Nature. 570 (7762), 523-527 (2019).
- Qian, X., et al. Brain-region-specific organoids using mini-bioreactors for modeling ZIKV exposure. Cell. 165 (5), 1238-1254 (2016).
- Hunter, Z. L., Leeson, H. C., Shaker, M. R., Wolvetang, E. J., Vadlamudi, L. Human induced pluripotent stem cells generated from epilepsy patients for use as in vitro models for drug screening. Stem Cell Research. 60, 102673 (2022).
- Kaur, A., Macip, S., Stover, C. M. An appraisal on the value of using nutraceutical based senolytics and senostatics in aging. Frontiers in Cell and Developmental Biology. 8, 218 (2020).
- Wang, F., et al. Safety and efficacy of dispase and plasmin in pharmacologic vitreolysis. Investigative Ophthalmology & Visual Science. 45 (9), 3286-3290 (2004).
Tags
ABOUT JoVE
Copyright © 2024 MyJoVE Corporation. All rights reserved