
Summary
Abstract
Introduction
Protocol
Representative Results
Discussion
Acknowledgements
Materials
References
Neuroscience
Надежная и высоковоспроизводимая генерация органоидов кортикального мозга для моделирования старения нейронов головного мозга in vitro
В этом исследовании мы предоставляем подробную технику для простой, но надежной кортикальной органоидной системы культивирования с использованием стандартных культур hPSC без фидера. Это быстрый, эффективный и воспроизводимый протокол для генерации органоидов, которые моделируют аспекты старения мозга in vitro.
Органоиды мозга представляют собой трехмерные модели развивающегося человеческого мозга и обеспечивают убедительную, передовую платформу для моделирования заболеваний и крупномасштабного геномного и лекарственного скрининга. Из-за самоорганизующейся природы клеток в органоидах головного мозга и растущего диапазона доступных протоколов для их генерации были выявлены проблемы с гетерогенностью и изменчивостью между органоидами. В этой протокольной статье мы описываем надежный и воспроизводимый протокол, который в значительной степени преодолевает эти проблемы и генерирует кортикальные органоиды из нейроэктодермальных предшественников в течение 1 месяца, и который может поддерживаться более 1 года. Этот высоковоспроизводимый протокол может быть легко выполнен в стандартной комнате для культивирования тканей и приводит к органоидам с богатым разнообразием типов клеток, обычно встречающихся в развивающейся коре головного мозга человека. Несмотря на их ранний состав развития, нейроны и другие типы клеток головного мозга человека начнут проявлять типичные признаки старения в нейронных клетках после длительной культуры in vitro , что делает их ценной и полезной платформой для изучения нейронных процессов, связанных со старением. Этот протокол также описывает метод обнаружения таких стареющих клеток в корковых органоидах головного мозга с использованием окрашивания бета-галактозидазы, связанного со старением.
Наши текущие знания о человеческом мозге в значительной степени основаны на животных моделях и посмертных образцах мозга. Биология стволовых клеток является быстро развивающейся областью, которая дает новое понимание базовой биологии развития человеческого мозга и патологических факторов расстройств головного мозга человека. Человеческие плюрипотентные стволовые клетки (hPSCs) являются бесценным инструментом для моделирования человеческого мозга посредством генерации органоидов, органоподобной трехмерной (3D) ткани, которая обычно повторяет траектории развития, клеточный состав и архитектуру развивающегося человеческого мозга. Органоиды мозга самособираются и состоят из нервных стволовых клеток, определенных нейронных предшественников, зрелых нейронов и типов глиальных клеток. Органоиды, таким образом, предоставляют уникальную возможность для изучения раннего человеческого мозга, который часто недоступен для прямых экспериментов, но также имеет внутренние ограничения, такие как отсутствие сосудистой системы и иммунной системы.
Методологии генерации органоидов мозга использовались двумя различными способами: неуправляемая и управляемая дифференциация. Неуправляемые органоидные методы мозга основаны на спонтанных внутренних дифференцировальных способностях стволовых клеток, которые управляют тканевым морфогенезом 1,2 и позволяют возникать различные идентичности клеточных линий, начиная от переднего мозга, среднего мозга и заднего мозга до сосудистого сплетения, сетчатки и мезодермы. Напротив, управляемые органоидные методы мозга требуют существенного использования внешних факторов для стимулирования HPSC к желаемому паттерну нейронных линий, представляющих один тип области мозга, таких как медиальная ганглионная возвышение3,передний мозг 4, средний мозг5, гипоталамус6, мозжечок7 и сосудистое сплетение8. Эта способность генерировать различные области мозга с различными клеточными линиями и возможность их слияния по желанию делает органоиды мозга отличной моделью для исследования развития человеческого мозга и расшифровки основных механизмов заболеваний, связанных с мозгом. Хотя эти методы генерации органоидов мозга предлагают прорыв в моделировании областей человеческого мозга, изменчивость и неоднородность между органоидами остаются значительным ограничением для систематических и количественных исследований, таких как скрининг лекарств.
Текущий протокол основан на методе, разработанном в нашей недавней статье9 , и включает селективную дифференциацию колоний hPSC в сторону идентичности нейроэктодермы (NEct) с двойными ингибиторами SMAD (SB-431542 и LDN 193189), которые затем обладают способностью самоорганизовываться в течение 4 дней в 3D-сфероиды нейроэпителия под влиянием сигнализации FGF2. Эти сфероиды нейроэпителия надежно генерируют однородные кортикальные органоиды с клеточным составом in vivo в течение 4 недель после дифференцировки. Протокол, описанный здесь, построен на наших предыдущих выводах, показывающих, что ингибирование двойной передачи сигналов SMAD (Suppressor of Mothers Against Decapentaplegic) способствует дифференцировке hPSCs в сторону ростральных нервных стволовых клеток, полученных из нейроэктодермальных предшественников10 , путем, среди прочего, ингибирования эндодермальной, мезодермальной и трофэктодермальной судьбы клеток выбора11 . Кроме того, встраивание сфероидов нейроэпителия в матрицу базальной мембраны, сертифицированную hESC, вызывает значительное почкование нейроэпителии, образуя желудочки с апикобазальной полярностью. Крупномасштабная культура показала воспроизводимость и однородность корковых органоидов независимо от клеточных линий, клонов или партий и, таким образом, представляет собой надежную и стабильную систему стволовых клеток, имитирующую раннее развитие коры головного мозга человека в области здоровья и заболеваний in vitro. Далее мы описываем протокол обнаружения маркеров стареющих нейрональных клеток в органоидах коры головного мозга, полученных из hPSCs, которые культивировались в течение длительных периодов времени.
После покрытия hPSCs при плотности посева 20%-30% клетки обрабатывают двойными ингибиторами SMAD в течение 3 дней для дифференцировки колоний hPSC в сторону нейроэктодермальных колоний. Эти колонии затем осторожно поднимаются с помощью диспазы и высеваются в сверхнизкие прикрепленные 6-луночные пластины, дополненные FGF2. Плавающие 2D-колонии самоорганизуются в 3D-нейроэктодермальные сфероиды в течение ночи и выдерживаются в течение 4 дней в среде N2, ежедневно дополняемой FGF2. После того, как сфероиды установили нейроэпителиальный слой, они могут быть встроены в матрицу базальной мембраны. Регулярно добавляя свежую терминальную дифференцирующую среду, исследователи будут наблюдать прогрессирующее расширение и бутонизацию нейроэпителии в корковых органоидах. Исследователи могут пожелать диссоциировать эти органоиды для проведения транскрипционного и протеомного профилирования. Кроме того, визуализация Brightfield рекомендуется для мониторинга качества корковых органоидов. Анализ может быть выполнен путем фиксации, криосекции и иммуноокрашивания. Описания и методы для этих методов были ранее описаны12. В конечном счете, этот протокол позволяет исследователям быстро и надежно генерировать однородные органоиды кортикального мозга для моделирования развивающегося кортикального мозга человека с недорогим и ограниченным оборудованием, а также для изучения аспектов старения клеточных нейронов, как описано в этой статье.
1. Генерация органоидов кортикального мозга
ПРИМЕЧАНИЕ: Все этапы, описанные в настоящем разделе протокола, будут осуществляться в вытяжке биобезопасности класса 2, если не указано иное.
- Индукция 2D нейроэктодермальных колоний из культуры hPSC 2D (дни от -1 до 3)
- Перед индукции пластинчатые колонии hPSC на матрице базальной мембраны, сертифицированной hESC, в 6-луночной пластине с плотностью 20%-30%. Достигайте этой плотности путем пропускания колоний hPSC из одной скважины 6-луночной плиты при 60% слиянии в три скважины из 6-луночной плиты.
- Для покрытия матрицы базальной мембраны разбавляют матрицу базальной мембраны в соотношении 1:50 в простой базальной среде. Равномерно нанесите 1 мл/лунку 6-луночной пластины, инкубируйте в течение 1 ч при комнатной температуре (RT), а затем аспирируйте.
- Поддерживать колонии hPSC в течение 1 дня в 2 мл безсыворочной питательной среды клеток перед дифференцировкой.
- В день дифференциации hPSC осмотрите колонии hPSC с помощью микроскопии с ярким полем с увеличением от 4x до 10x, чтобы обеспечить здоровые колонии без обнаруживаемой дифференциации.
ПРИМЕЧАНИЕ: Здоровые hPSCs будут образовывать плотные краевые колонии с клетками, которые имеют большое ядро, очень маленькую цитоплазму и выступающие ядрышки. Дифференцированные колонии iPSC будут демонстрировать явные морфологические различия с описанными выше колониями hPSC, особенно вокруг внешних краев колоний или в центре. - Добавьте реагенты, перечисленные в таблице 1 , чтобы составить среду N2, необходимую для дифференцировки. Перед использованием доведите эту среду до RT.
- После приема РТ аспирируйте безсыворочную среду клеточной культуры из каждой лунки 6-луночной пластины и замените 2 мл среды N2, осторожно добавленной серологической пипеткой 5 мл.
- Добавьте двойные ингибиторы SMAD, SB-431542 (10 мкМ) и LDN 193189 (100 нМ).
ПРИМЕЧАНИЕ: Ингибиторы SMAD могут быть добавлены в среду N2 после того, как среда была помещена в каждую лунку, или в необходимое количество среды N2 до замены среды клеточной культуры без сыворотки. Ингибиторы могут быть равномерно включены в среду путем осторожного закручивания пластины или инвертирования трубки, содержащей среду и ингибиторы, 3-4 раза. - Добавляйте свежие среды N2, дополненные SB-431542 (10 мкМ) и LDN 193189 (100 нМ) ежедневно в каждую лунку в течение следующих 2 дней.
ПРИМЕЧАНИЕ: Свежая среда N2 добавляется для уменьшения длительного воздействия на клетки DMSO, который используется для растворения соединений SB-431542 и LDN 193189 и предотвращения цитотоксичности.
- Генерация 3D нейроэктодермальных сфероидов из индуцированных 2D нейроэктодермальных колоний (дни с 3 по 7)
- Поднимите индуцированные нейроэктодермальные колонии с помощью диспазы, выполнив шаги 1.2.2-1.2.8.
- Во-первых, удалите 2 мл среды N2 из 6-луночной пластины и промыть 1x HBSS, чтобы убедиться, что вся среда N2 удалена.
ПРИМЕЧАНИЕ: Среда N2 может влиять на ферментную активность диспазы, препятствуя адекватному отслоению нейроэктодермальных колоний из колодца. - Добавьте 1 мл 2,4 единицы / мл диспазы к каждому колониесодержащему колодцу.
- Инкубировать лунку в течение 20-25 мин (максимум 30 мин) при 37 °C. Регулярно проверяйте отряд колонии.
ПРИМЕЧАНИЕ: Небольшие колонии могут отделяться в течение 20 минут. Любые колонии, которые остаются застрявшими через 30 минут, следует игнорировать. - После инкубации добавьте в скважину 1 мл среды N2, чтобы остановить активность фермента диспазы и перенесите колонии в трубку объемом 15 мл с использованием широкоствольного наконечника пипетки P1000 или модифицированного наконечника пипетки P1000, разрезанного стерильными ножницами (что делает его широкобурным наконечником P1000).
- Позвольте колониальным скоплениям опуститься на дно трубы под действием силы тяжести.
ПРИМЕЧАНИЕ: Этот процесс займет примерно 1 минуту. - После того, как комки утонули, осторожно удалите супернатант стандартным наконечником пипетки P1000 и замените его 1 мл свежей среды N2. Повторите этот шаг промывки три раза, чтобы обеспечить полное удаление диспазы.
ПРИМЕЧАНИЕ: Любая оставшаяся диспаза предотвратит равномерное образование нейроэктодермальных сфероидов и вызовет гибель клеток. - После промывки повторно суспендируют сгустки клеток в 3 мл среды N2 и переложат в одну лунку 6-луночной пластины и добавят 40 нг/мл bFGF.
ПРИМЕЧАНИЕ: Если бы большое количество нейроэктодермальных колоний было отделено, эти колонии могли бы быть покрыты двумя или более скважинами 6-луночной пластины, чтобы предотвратить слияние сфероидов. Через 24 ч после рассеивания отряда колоний проверьте, образовались ли сфероиды. - Поддерживайте сфероиды в одной и той же среде в течение следующих 3-4 дней, но добавляйте свежий bFGF (40 нг / мл) в каждую скважину ежедневно, чтобы способствовать нейроэктодермальной пролиферации клеток, самоорганизации, а также индуцировать и расширять нейроэпителию.
ПРИМЕЧАНИЕ: Если сфероиды были покрыты более высокой плотностью, среда, вероятно, станет желтой и потребует замены каждые 2 дня на свежий bFGF (40 нг / мл). Однако это не рекомендуется, и вместо этого в каждой скважине следует поддерживать меньшее количество сфероидов, чтобы избежать этой проблемы. Сфероиды могут быть внедрены в матрицу базальной мембраны через 3 дня, если нейроэпителия очевидна. Если нейроэпителия не проявляется или не выглядит сильной, то сохраните сфероиды еще на один день и проверьте еще раз.
- Дифференциация и поддержание органоидов корков головного мозга (День 8)
- Подготовьте терминальную дифференцировочную среду (ДМ) с помощью реагентов, перечисленных в таблице 2. Принесите это СМИ на RT.
- Оттаивайте 100% hESC-сертифицированную матрицу базальной мембраны на льду.
- Подготовьте лист парапленки с ямочками, стерилизованный 70% этанолом, в чашке Петри 10 см и поместите его на стереомикроскоп под капюшоном.
ПРИМЕЧАНИЕ: Пустой лоток с наконечниками пипетки объемом 200 мкл можно использовать для создания сетки ямочек. - Отрежьте конец наконечника пипетки объемом 100 мкл (чтобы сделать его широкобурным). Это будет использоваться для встраивания нейронных сфероидов в подвальную матрицу без их разрушения.
- Используя стереомикроскоп, выберите нейронные сфероиды аналогичного размера (500 мкм) из 6-луночной пластины и перенесите их в парапленку-ямочки с помощью широкоствольного наконечника пипетки 100 мкл, поместив один нейронный сфероид на ямочку.
- Аккуратно удалите все лишние носители, оставив ровно столько, чтобы покрыть сфероиды.
ПРИМЕЧАНИЕ: Это делается для поддержания качества сфероидов перед добавлением подвальной матрицы и для обеспечения того, чтобы они не высыхали. - Осторожно добавьте 18 мкл фундаментной матрицы поверх сфероида, расположив сфероид в центре матричной капли. Используйте конец наконечника пипетки объемом 10 мкл, чтобы центрировать сфероиды в матрице.
ПРИМЕЧАНИЕ: Постарайтесь добавить подвальную матрицу как можно быстрее, чтобы избежать высыхания избыточной среды вокруг сфероидов. - Накройте чашку Петри 10 см и перенесите в инкубатор и инкубируйте чашку парапленки со встроенными сфероидами подвальной матрицы при 37 °C в течение 25 мин.
- После инкубации смойте встроенные сфероиды в 24-луночную пластину с низким креплением, используя наконечник P1000 с 0,5 мл среды DM, убедившись, что отдельные встроенные сфероиды размещены в одной скважине каждая.
ПРИМЕЧАНИЕ: Если в одну скважину попадает более одного встроенного сфероида, используйте широкопроходный наконечник P1000 для переноса другого сфероида в новую скважину. - Поддерживайте дифференцированные органоиды кортикального мозга в среде DM в течение длительных периодов времени, при этом изменения среды происходят каждые 2 дня, когда органоиды становятся больше и старше.
ПРИМЕЧАНИЕ: В течение первой недели дифференциации среда может меняться каждые 3 дня.
2. Характеристика старения нейронов в корковых органоидах
- Обработка кортикальных органоидов для криосекций:
ПРИМЕЧАНИЕ: Этапы выполнялись в вытяжке биобезопасности класса 2.- Подготовьте 2 мл пробирки, каждая из которых заполнена 1,5 мл 4% параформальдегида (PFA).
- Вырежьте конец наконечника пипетки P1000 (чтобы сделать его широкобурным) и аккуратно перенесите каждый органоид в одну из 2 мл пробирок, приготовленных выше (по одному органоиду на трубку).
ПРИМЕЧАНИЕ: Чтобы предотвратить смешивание избыточной среды DM с PFA, позвольте органоиду опуститься к открытию наконечника пипетки P1000 и положить наконечник чуть выше верхней части PFA в трубке, прежде чем пипетировать органоид. Это позволит исследователю перенести только органоид и очень минимальную среду. - Позвольте процессу фиксации происходить при 4°C в течение 1 ч.
- Используя неразрезанный наконечник пипетки P1000, осторожно аспирируйте избыток PFA и добавьте 1,5 мл холодного 1x PBS.
- Переведите трубки в орбитальный шейкер, установленный при 70 об/мин в течение 10 мин при RT.
- Повторите процесс стирки с холодным 1x PBS три раза, чтобы убедиться, что весь PFA был тщательно удален.
ПРИМЕЧАНИЕ: Не утилизируйте PFA в обычных контейнерах для отходов; вместо этого подготовьте для этого специальный контейнер для удаления химических отходов, поскольку PFA представляет опасность. - Поместите органоиды в 1x PBS, содержащий 30% сахарозы, и инкубируйте при 4 °C, пока все органоиды не опустятся на дно трубки.
ПРИМЕЧАНИЕ: Время, необходимое для того, чтобы органоиды опустились, зависит от размера/возраста органоидов. 3-месячным органоидам может потребоваться до 5 ч. - Используя разрезанный широкоствольный наконечник пипетки P1000, аккуратно переложите от трех до пяти органоидов в монтажную форму, содержащую монтажный раствор, изготовленный из 30% сахарозы и 100% оптимальной температуры резки (OCT) среды, в соотношении 3:2.
- Используйте наконечник пипетки объемом 10 мкл с помощью стереомикроскопа для ориентации и позиционирования органоидов в виде сетки.
- Поместите форму на сухой лед для затвердевания раствора сахарозы OCT, прежде чем приступить к криосекционированию (16-20 мкм) с использованием криостата.
ПРИМЕЧАНИЕ: Для бета-галактозидазы, связанной со старением, все ткани должны быть обработаны для разрезания после того, как они утонули. Для иммунофлуоресценции ткани могут быть обработаны для разрезания на следующий день. Все слайды, содержащие срезы, должны храниться при -20 °C до последующей иммунофлуоресценции или бета-галактозидазы, если они не окрашиваются немедленно.
- Процесс анализа старения в кортикальных органоидах головного мозга:
ПРИМЕЧАНИЕ: Следующие шаги могут быть выполнены на обычном лабораторном стенде.- Переложите слайды в контейнер для окрашивания предметных предметов микроскопа с крышкой и трижды промойте разрезанную органоидную ткань 1x PBS в течение 10 минут на RT, чтобы удалить любой лишний монтажный раствор.
- После этого инкубируют промытую ткань свежеприготовленным раствором для окрашивания бета-галактозидазы в течение ночи при 37 °C.
ПРИМЕЧАНИЕ: Раствор для окрашивания бета-галактозидазы изготовлен из фосфатного буфера (для 10 мл фосфатного буфера: 8,15 мл 1 М2PO4, 1,85 мл 1M Na2HPO4), скорректированного рН = 6, 100 мМ гексацианоферрата калия (III), 100 мМ тригидрата гексацианоферрата (II) калия, 5 М NaCl, 1 М MgCl2, 20 мг/мл X-Gal. Избегайте использования стандартного инкубатора клеточных культур, содержащего CO2 , поскольку CO2 изменяет рН раствора для окрашивания бета-галактозидазы. - Промыть окрашенные ткани 1x PBS три раза в течение 10 мин каждый на RT, чтобы удалить раствор бета-галактозидазы.
- Установите промытые салфетки со стеклянным антизатухающим креплением и дайте монтажному раствору затвердеть в течение 30 минут на RT перед просмотром под микроскопом.
Здесь мы описали надежный протокол, который позволяет исследователям генерировать однородные органоиды коры головного мозга, полученные из hPSC, которые имитируют in vivo кортикальную область мозга человека в течение 1-3 месяцев после культивирования. Колонии hPSC сначала культивируются в дифференцировочных средах для генерации нейроэктодермальных колоний, которые затем могут быть использованы для формирования нейронных сфероидов. Эти сфероиды впоследствии встраиваются в матрицу базальной мембраны и поддерживаются в течение длительных периодов времени для получения органоидов, которые могут быть использованы для моделирования старения нейронов (см. Рисунок 1 для краткого описания протокола). Стоит отметить, что культивирование этих органоидов в ультра-непокрытых 24-луночных пластинах вызывает клеточный стресс и способствует фенотипам, связанным со старением, в течение 13 недель культуры in vitro . Органоиды, полученные из этого протокола, также могут поддерживаться в перемешиваемых биореакторах для оптимального роста и дифференцировки нервных клеток кортикальной пластины или на границе раздела воздух-жидкость.
Для начала колонии hPSC культивируют за 1 день до нейроэктодермальной дифференцировки. Крайне важно, чтобы эти колонии hPSC культивировались только до 20-30% слияния и имели максимально возможное качество: плотный плоский монослой без дифференцированных клеток, загрязняющих колонии (рисунок 2A, B). Плюрипотентность колоний hPSC должна быть подтверждена экспрессией маркеров, таких как NANOG (рисунок 2C). Валидированные колонии hPSC затем подвергают воздействию нейроэктодермальной дифференцировочной среды N2 с SB-431542 и LDN 193189. После 3 дней поддержания в этой среде колонии hPSC должны дифференцироваться в нейроэктодермальные колонии и больше не показывать ту же плотную плоскую монослойную морфологию hPSCs (рисунок 2B), а скорее они станут более длинными столбцообразными клетками (рисунок 2B). Эти клетки также будут отрицательными для маркеров плюрипотентности, таких как NANOG (рисунок 2C).
Именно на этом этапе нейроэктодермальные колонии ферментативно отделяются диспазой, и каждой здоровой и успешно отсоединенной колонии разрешается самоорганизоваться и образовывать молодой нервный сфероид (рисунок 2D, дополнительное видео 1). Только здоровые, чистые нейроэктодермальные колонии будут отделяться в сроки, указанные для активности диспаза; все остальные колонии следует игнорировать, так как они приведут к ухудшению качества сфероида. При ежедневном воздействии FGF2 в среде N2 нервные стволовые клетки (SOX2+) в этих сфероидах (рисунок 2D, день 1) будут размножаться и образовывать значительное количество нейронных розеток (рисунок 2D, день 4). Эти розетки будут экспрессировать плотное соединение и эпителиальный маркер ZO1 в клетках, расположенных в центре розеток и вдоль внешнего края сфероида, демонстрируя апикально-базальную полярность сфероида (рисунок 2D, день 4). Метод цельной 3D-визуализации сфероидов был описан ранее13. Ежедневный осмотр сфероидов должен выявить образование плотного, темного наружного края и яркой периферии сфероидов, это является нейроэпителиальным слоем. Этот слой должен быть достаточно сформирован через 3-4 дня с приблизительным диаметром 500 мкм, после чего сфероиды могут быть внедрены в основную матрицу. Если этот слой отсутствует или только слабо сформирован, сфероиды недостаточно развиты, чтобы двигаться вперед. Рекомендуется подождать еще сутки, чтобы наблюдать какие-либо изменения, но если этого не наблюдается, не обращайте внимания на эти сфероиды.
Репрезентативное изображение сфероидов после 3 дней культивирования можно увидеть на рисунке 2D. Сфероиды с плотным нейроэпителиальным слоем, которые не слились с другими соседними сфероидами, имеют полупрозрачную ткань и демонстрируют образование нейронной розетки, выбираются для внедрения в фундаментную матрицу. После внедрения сфероид быстро размножится и начнет почковаться: появятся узлы компактной ткани, расширяющиеся наружу от основного тела сфероида. Это очевидно между 1-3 неделями в подвальной матрице и может наблюдаться через несколько клеточных линий (рисунок 3A). Количественный анализ встроенных сфероидов подтверждает наличие эпителиальных клеток в 100% сфероидов по трем различным клеточным линиям, подтверждая однородность и воспроизводимость, ожидаемые от этого протокола (рисунок 3B). Количественная оценка диаметра органоидов во время дифференцировки in vitro дополнительно подтверждает воспроизводимость через различные линии hPSC (рисунок 3C). Если почкование не происходит, сфероиды не развиваются должным образом и должны быть выброшены. Как только сфероиды были встроены в матрицу, их развитие прогрессирует, и сфероиды теперь называются органоидами. Иммунофлуоресцентное окрашивание также подтверждает наличие нейронных клеток-предшественников (PAX6), а также маркеров кортикального слоя, окрашенных CTIP2 и SATB2 в органоидах с прозрачным наслоением (рисунок 3D, E). Это наслоение наблюдается в разных временных точках поддержания органоидов (рисунок 3D, E). Метод иммуногистохимии тканей был описан ранее14.
Одним из возможных применений этих органоидов является изучение того, как процессы, связанные со старением нейронов, влияют на мозг. Чтобы исследовать это, успешно сгенерированные органоиды извлекают из нескольких различных временных точек для сечения и окрашивания для стандартных молекулярных биомаркеров старения, таких как бета-галактозидаза, связанная со старением, и р21. На фиг.4А показано репрезентативное изображение связанного со старением бета-галактозидазного окрашивания органоидов через 4 и 13 недель после встраивания в матрицу базальной мембраны. Между неделями 4 и 13 наблюдается заметное увеличение присутствия бета-галактозидазы, связанной со старением, что свидетельствует о том, что клеточное старение, признанный фактор старения организма, произошло за это время в культуре. Иммунофлуоресцентное окрашивание органоидов на 13-й неделе подтвердило наличие другого маркера старения, p21, меченого совместно со зрелым маркером кортикальных нейронов (CTIP2) и можно увидеть на рисунке 4B. Следует, однако, отметить, что наличие р21 является маркером остановки клеточного цикла и само по себе не является окончательным маркером старения, а обнаружение других маркеров старения, таких как p16 и SASP (связанный со старением секреторный фенотип), рекомендуется для окончательной идентификации клеток как стареющих.
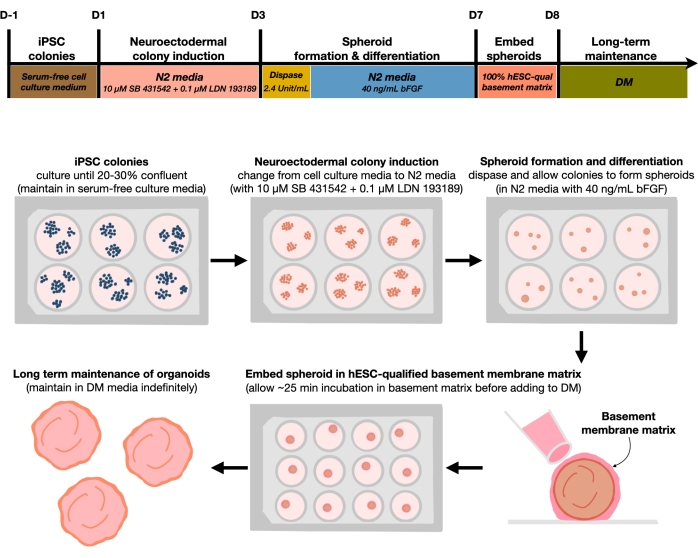
Рисунок 1: Принципиальная схема генерации воспроизводимых органоидов кортикального мозга. Схематический рабочий процесс экспериментальной процедуры генерации кортикальных органоидов головного мозга из гПСК, поддерживаемых в среде без фидера. Рабочий процесс предоставляет обзор шести шагов, связанных с дифференциацией 2D hPSCs в 3D-узорчатые корковые пластинки тканей человека в органоидах. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
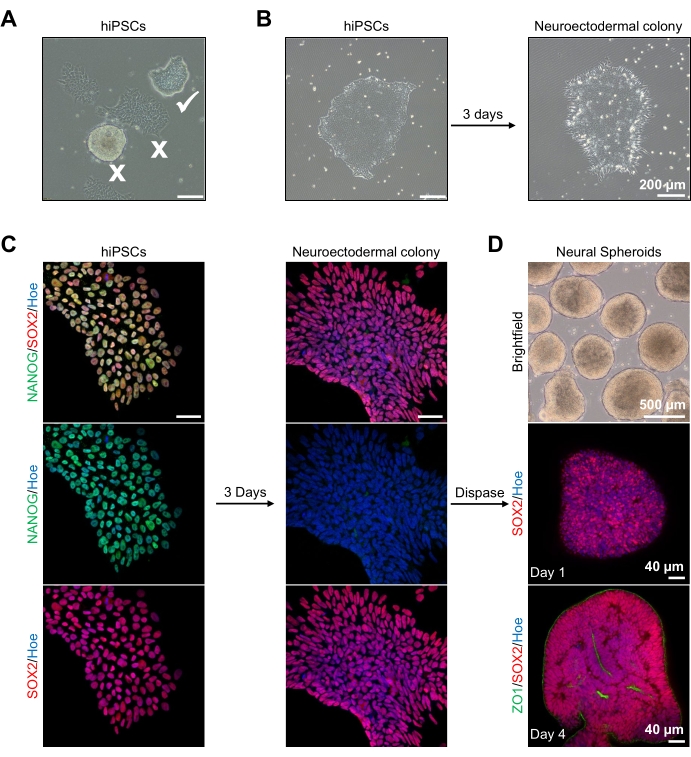
Рисунок 2: Генерация нейронных сфероидов, полученных из нейроэктодермальных колоний-hPSCs. (A) Репрезентативные изображения человеческого PSC, демонстрирующие оптимальные (белый клещ) и дифференцированные колонии (белый крест). Шкала: 200 мкм, 4-кратное увеличение. (B) Репрезентативное изображение нейроэктодермальной колонии, полученное из hPSCs после 3 дней лечения двойным ингибитором SMAD. Шкала: 200 мкм, 4-кратное увеличение. (C) Человеческие колонии PSC были дифференцированы в сторону нейроэктодермальных колоний. Изображения представляют собой окрашивание PSC (на 1-й день) и нейроэктодермальных (на 3-й день) колоний SOX2 (красный), NANOG (зеленый), все ядра были контрокрашены Hoechst 33342 (синий). Шкала: 40 мкм, 100-кратное увеличение. (D) Изображения, показывающие стадии развития кортикальных сфероидов головного мозга с течением времени в культуре in vitro под ярким полем и цельного иммуноразвитого SOX2 (красный) на 1-й день и двойной иммуноокрашенный SOX2 (красный) и ZO1 (зеленый) на 4-й день. Шкала изображения яркого поля составляет 500 мкм, 4-кратное увеличение, шкала нижних изображений - 40 мкм, 20-кратное увеличение. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
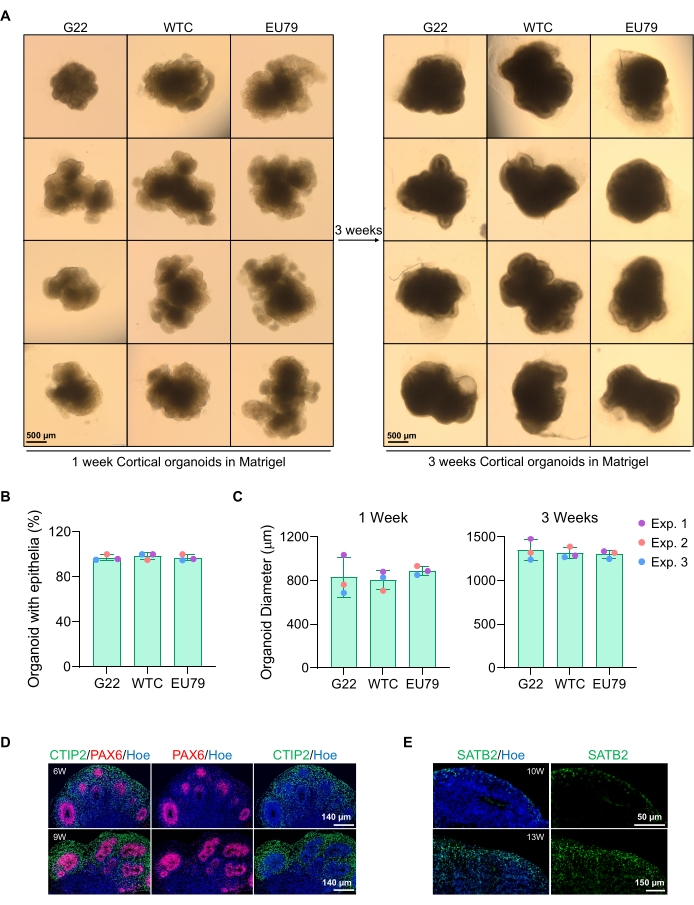
Рисунок 3: Характеристика органоидов коркового мозга, полученных из различных линий hPSC. (A) Репрезентативные изображения кортикальных органоидов головного мозга, полученных из линий IPSC человека G22, WTC и EU79, культивируемых в течение 3 недель in vitro. Шкала всех изображений составляет 500 мкм, 2-кратное увеличение. (B) Процент успешной генерации органоидов кортикального мозга через 3 недели дифференцировки in vitro в различных линиях hPSC (G22, WTC и EU79). N = 3. Данные представлены в виде среднего ± стандартного отклонения. (C) Гистограммы, показывающие рост органоидов кортикального мозга (на основе среднего диаметра) на 1 и 3 неделях дифференцировки in vitro в различных линиях плюрипотентных стволовых клеток человека (G22, WTC и EU79). N = 3. Данные представлены в виде среднего ± стандартного отклонения. (D) Репрезентативные изображения участков 6-недельных и 9-недельных органоидов кортикального мозга, полученных из G22 hPSCs, иммуноокрашенных для зоны желудочков PAX6 (красный) и кортикальной пластины CTIP2 (зеленый). Все секции были покрыты Hoechst 33342 (синий). Шкала = 140 мкм, 20-кратное увеличение. W — неделя. (E) Репрезентативные изображения участков 10-недельных и 13-недельных органоидов кортикального мозга, полученных из HPSCs WTC, иммуноокрашенных для кортикального слоя IV SATB2 (зеленый). Все секции были покрыты Hoechst 33342 (синий). 10 недель шкала изображения = 50 мкм, 40-кратное увеличение. 13 недель изображение Шкала шкалы = 150 мкм, 40-кратное увеличение. W — неделя. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
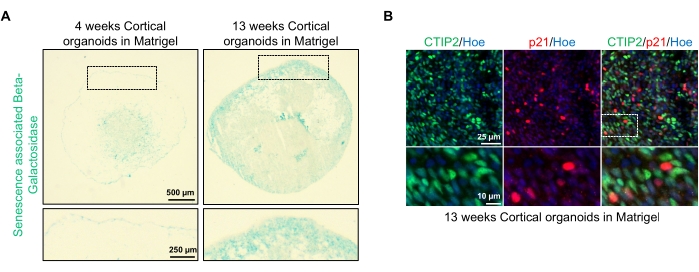
Рисунок 4: Характеристика старения в корковых органоидах головного мозга, полученных из hPSCs. (A) Репрезентативные изображения участков органоидов кортикального мозга человека, полученных из HPSCs WTC, культивируемых в течение 4 и 13 недель in vitro и окрашенных SA-β-gal. Шкала = 500 мкм, шкала увеличенных изображений = 250 мкм, 4-кратное увеличение. Пунктирное поле указывает на увеличенное изображение. (B) Репрезентативные изображения участков 13-недельных органоидов кортикального мозга, полученных из человеческих EU79 hPSCs, иммуноокрашенных для корковых нейронов CTIP2 (зеленый) и p21 (красный). Все секции были урезаны Hoechst 33342 (синий). Шкала = 25 мкм, шкала увеличенных изображений = 10 мкм, 40-кратное увеличение. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
| Компоненты мультимедиа | Концентрация |
| DMEM Питательная смесь F12 10x 500 мл (DMEM/F-12) | |
| Добавка N2 5 мл (100x) | Увеличено на 1% |
| B 27 Добавка 10 мл | Добавляется по 2% |
| MEM Раствор заменимых аминокислот (100x) | Добавляется в размере 1% |
| Пенициллин-стрептомицин (10 000 Ед/мл) | Добавляется в размере 1% |
| 2-Меркаптоэтанол 50 мл (1000x) | Добавляется в размере 0,1% |
Таблица 1: N2 Средний. В таблице перечислены реагенты, необходимые для приготовления среды N2.
| Компоненты мультимедиа | Концентрация |
| DMEM Питательная смесь F12 10x 500 мл (DMEM/F-12) | DM среда изготовлена с соотношением 1:1 DMEM/F12 и нейробазальных сред |
| Нейробазальная среда | |
| Добавка N2 5 мл (100x) | Наложено на 0,5% |
| B 27 Добавка 10 мл | Добавляется в размере 1% |
| MEM Раствор заменимых аминокислот (100x) | Добавляется в размере 1% |
| Добавка GlutaMAX 100x | Добавляется в размере 1% |
| Пенициллин-стрептомицин (10 000 Ед/мл) | Добавляется в размере 1% |
| Раствор инсулина человеческий рекомбинантный | 12,5 мкл на 50 мл среды |
| 2-Меркаптоэтанол 50 мл (1000x ) | 17,5 мкл на 50 мл среды |
Таблица 2: Дифференциационная среда (ДМ). В таблице перечислены реагенты, необходимые для приготовления дифференцировочной среды.
Дополнительное видео 1. Живая визуализация индуцированного hNEct 2D преобразования листа/колонии в 3D при обработке bFGF. Индуцированные колонии hNEct осторожно отделяли от тарелки с помощью диспазы, как описано выше, и переносили на 6-луночную культуральную пластину с низким прикреплением. Колонии 2D hNEct были преобразованы в сфероиды 3D hNEct в течение 12 ч. Серийные изображения снимались каждые 5 минут. Шкала = 100 мкм. Пожалуйста, нажмите здесь, чтобы загрузить это видео.
Чтобы обеспечить возможность использования органоидов головного мозга, полученных из hPSC, в скрининге лекарств и моделировании заболеваний, крайне важно создавать органоиды в соответствии с воспроизводимым и надежным протоколом15. Органоиды головного мозга обычно генерируются из эмбриональных тел, полученных из hPSCs, которые затем встраиваются во внеклеточный матрикс, который способствует расширению тканей и нейронной дифференцировке. По сравнению с такими протоколами, как Lancaster's 1,16,17 и Velasco 18, которые начинаются с эмбриоидных тел и позволяют развивающимся органоидам следовать пути дифференцировки по умолчанию, мы обнаружили, что начало создания органоидов кортикального мозга с клетками NEct человека, а не с эмбриональными телами, улучшает консистенцию кортикального органоидного образования головного мозга. Это, следовательно, также позволяет масштабироваться, необходимый для лекарственного и фенотипического скрининга. Поскольку клетки NEct человека могут быть не только расширены до значительных количеств, но также могут быть легко криоконсервированы, этот подход также улучшает воспроизводимость между экспериментами. Следует также отметить, что по сравнению с другими протоколами, которые приняли использование биореакторов и аналогичных технологий, для этого протокола не требуется специализированное оборудование, что делает его пригодным для любой лаборатории6. Наконец, время, необходимое для генерации зрелых органоидов, которые являются положительными для маркеров коркового слоя, таких как SATB2, сокращается по сравнению с протоколами Ланкастера1 и биореактора 6,19, что делает его более подходящим для изучения траектории развития коры головного мозга человека в области здоровья и заболеваний 1,6,16.
Кроме того, учитывая постоянно растущее глобальное воздействие на здравоохранение связанных со старением заболеваний, таких как деменция, которые связаны с увеличением типов стареющих клеток в мозге, что способствует патогенезу, способность идентифицировать и тестировать соединения, которые могут улучшить старение мозга, представляет огромный интерес. Несмотря на то, что hPSCs, как известно, эпигенетически омолаживаются во время процесса перепрограммирования20, мы находим устойчивое увеличение стареющих клеток в органоидах кортикального мозга, культивируемых в течение длительных периодов времени. Это многообещающая разработка, которая теперь позволяет проводить скрининг препаратов, которые устраняют такие стареющие клетки из мозга (сенолитики) или замедляют этот процесс (сеностатики)21. Поскольку корковые органоиды головного мозга, полученные из NEct человека, имеют человеческое происхождение, этот подход, вероятно, сократит традиционный путь к продаже таких новых терапевтических средств.
В этом протоколе есть два важных шага. Во-первых, это правильный уровень слияния колоний hPSC в момент дифференциации. Колонии hPSC должны быть не более 30% сливающимися, чтобы гарантировать, что генерируемые колонии NEct не сливаются с соседними колониями и что отдельные органоиды управляются клонально. Второй критический шаг включает в себя правильное использование диспаза для подъема колоний NEct и производства нейронных сфероидов. Время инкубации с диспазой имеет решающее значение для конечного качества генерируемых нейронных сфероидов. Это связано с тем, что чрезмерное воздействие колоний с диспазой токсично для клеток22 и в конечном итоге влияет на качество генерируемых органоидов. Ограничение этого протокола заключается в том, что трудно контролировать размер нейронных сфероидов, потому что он зависит от размера начальных колоний, которые поднимаются с диспазой. Однако эту проблему можно преодолеть, выбрав нейронные сфероиды, которые имеют одинаковый размер при переходе к этапу встраивания.
Наконец, будущие приложения могут распространяться на использование этих воспроизводимых кортикальных органоидов в роботизированном анализе и биофармацевтических подходах к скринингу, обычно используемых в этой отрасли. Это подтверждается предварительными данными нашей лаборатории, указывающими на то, что генерация органоидов коркового мозга из клеток NEct человека может быть легко автоматизирована, что делает ее совместимой с этими подходами.
Авторам нечего раскрывать.
Эта работа поддерживается Фондом ускоренных исследований будущего медицинских исследований, флагманской миссией Лейкодистрофии Massimo (EPCD000034), Фондом медицинских исследований будущего - Миссией стволовых клеток (APP2007653). Авторы хотели бы поблагодарить д-ра Чжу Хён Ли (Корейский университет) за создание данных в дополнительном видео 1.
| Name | Company | Catalog Number | Comments |
| 16% Formaldehyde (W/V) Methanol-free | Thermo Fisher Scientific | 28908 | 4% of PFA are diluted in 1x PBS |
| 2-Mercaptoethanol 50 mL(1000x) | Life Technologies Australia (TFS) | 21985023 | Used in NM and DM media |
| B 27 Supplement 10 mL | Life Technologies Australia (TFS) | 17504044 | Used in NM and DM media |
| CKX53 microscope with SC50 camera | Olympus | ||
| Corning Costar 6 well cell culture plates | Sigma Aldrich Pty Ltd | CLS3516-50EA | |
| Dispase II powder | Thermo Fisher Scientific | 17105041 | Powder is dissolve in HBSS, filtered through 0.22 µm filter, aliquote at 10 mL and store at -20 °C |
| DMEM Nutrient Mix F12 10x 500 mL (DMEM/F-12) | Thermofisher | 11320082 | Used in NM and DM media |
| DMSO Dimethyl Sulfoxide | Sigma Aldrich Pty Ltd | D2650-100ML | |
| Dulbecco's Phosphate Buffered Saline | Sigma Aldrich Pty Ltd | D1408-500ML | |
| Falcon Matrigel hESC-qualified Matrix | In Vitro Technologies Pty Ltd | FAL354277 | Make aliquotes of 100 µL and stored at -20 °C |
| GlutaMAX Supplement 100x | Thermo Fisher Scientific | 35050061 | Used in NM and DM media |
| Hanks Balanced Salt Solution | Sigma Aldrich Pty Ltd | H8264 | |
| Human induced pluripotent stem cells (EU79) | In-house reporogrammed from skin fibroblast | ||
| Human induced pluripotent stem cells (G22) | Genea Biocells | Obtained from Genea Biocells (San Diego, United States) | |
| Human induced pluripotent stem cells (WTC) | Gift from Professor Bruce Conklin | ||
| InSolution TGF-Β RI Kinase Inhibitor VI, SB431542 | Merck | US1616464-5MG | |
| Insulin Solution Human Recombinant | Sigma Aldrich Pty Ltd | I9278 | Used in NM and DM media |
| LDN193189 Dihydrochloride | Sigma Aldrich Pty Ltd | SML0559-5MG | Used during differentiation |
| MEM Non-Essential Amino Acids Solution (100x) | Thermo Fisher Scientific | 11140050 | Used in NM and DM media |
| mTeSR Plus | STEMCELL TECHNOLOGIES | 100-0276 | Used to maintain hiPSC colonies prior to differentiation with NM media |
| N2 Supplement 5 mL (100x) | Life Technologies Australia Pty Ltd | 17502048 | Used in NM and DM media |
| Neurobasal Medium | Thermo Fisher Scientific | 21103049 | Used in DM media |
| OCT Embedding Compound Sakura Clear (118 mL/Bottle) | Tissue Tek | 4583 | |
| Parafilm M Roll Size 4 in. x 125 Ft | Sigma Aldrich Pty Ltd | P7793 | |
| Penicillin-Streptomycin (10,000 U/mL) | Thermo Fisher Scientific | 15140122 | Used in NM and DM media |
| Potassium Hexacyanoferrate (II) Trihydrate | Sigma Aldrich Pty Ltd | CP1087 | |
| Potassium hexacyanoferrate(III) | Sigma Aldrich Pty Ltd | 455946 | |
| Prolong Glass Antifade Mountant | Life Technologies Australia (TFS) | P36980 | |
| Recombinant Human FGF basic | R&D Systems | 233-FB-01M | Aliquotes are made at 20 µg/mL and stored at -20 °C |
| SB431542 | Tocris | 1614 | Used during differentiation |
| Sucrose | Sigma Aldrich Pty Ltd | PHR1001-1G | 30% of sucrose are diluted in 1x PBS |
| Ultra-Low attachment multiwell plates , 24 well plate, polystyrene | Sigma Aldrich Pty Ltd | CLS3473-24EA | |
| X-GAL EA | Life Technologies Australia (TFS) | R0404 | Make aliquotes of 20 mg/mL and storde at -80 °C |
- Lancaster, M. A., Knoblich, J. A. Generation of cerebral organoids from human pluripotent stem cells. Nature Protocols. 9 (10), 2329-2340 (2014).
- Mansour, A. A., et al. An in vivo model of functional and vascularized human brain organoids. Nature Biotechnology. 36 (5), 432-441 (2018).
- Xiang, Y., et al. Fusion of regionally specified hPSC-derived organoids models human brain development and interneuron migration. Cell Stem Cell. 21 (3), 383-398 (2017).
- Bagley, J. A., Reumann, D., Bian, S., Lévi-Strauss, J., Knoblich, J. A. Fused cerebral organoids model interactions between brain regions. Nature Methods. 14 (7), 743 (2017).
- Kwak, T. H., et al. Generation of homogeneous midbrain organoids with in vivo-like cellular composition facilitates neurotoxin-based Parkinson's disease modeling. Stem Cells. 38 (6), 727-740 (2020).
- Qian, X., et al. Generation of human brain region-specific organoids using a miniaturized spinning bioreactor. Nature Protocols. 13 (3), 565-580 (2018).
- Muguruma, K., Nishiyama, A., Kawakami, H., Hashimoto, K., Sasai, Y. Self-organization of polarized cerebellar tissue in 3D culture of human pluripotent stem cells. Cell Reports. 10 (4), 537-550 (2015).
- Shaker, M. R., Cooper-White, J., Wolvetang, E. J. Self-organizing 3D human choroid plexus-ventricle-cortical organoids. BioRxiv. , (2020).
- Shaker, M. R., Aguado, J., Chaggar, H. K., Wolvetang, E. J. Klotho inhibits neuronal senescence in human brain organoids. npj Aging and Mechanisms of Disease. 7 (1), 1-12 (2021).
- Shaker, M. R., et al. Anteroposterior Wnt-RA gradient defines adhesion and migration properties of neural progenitors in developing spinal cord. Stem Cell Reports. 15 (4), 898-911 (2020).
- Chambers, S. M., et al. Highly efficient neural conversion of human ES and iPS cells by dual inhibition of SMAD signaling. Nature Biotechnology. 27 (3), 275-280 (2009).
- Shaker, M. R., et al. Rapid and efficient generation of myelinating human oligodendrocytes in organoids. Frontiers in Cellular Neuroscience. 15, 631548 (2021).
- Lee, J. -. H., Shaker, M. R., Lee, E., Lee, B., Sun, W. NeuroCore formation during differentiation of neurospheres of mouse embryonic neural stem cells. Stem Cell Research. 34, 101691 (2020).
- Shaker, M. R., et al. Spatiotemporal contribution of neuromesodermal progenitor-derived neural cells in the elongation of developing mouse spinal cord. Life Sciences. 282, 119393 (2021).
- Shaker, M. R., et al. Neural epidermal growth factor-like like protein 2 Is expressed in human oligodendroglial cell types. Frontiers in Cell and Developmental Biology. 10, 803061 (2022).
- Lancaster, M. A., et al. Cerebral organoids model human brain development and microcephaly. Nature. 501 (7467), 373-379 (2013).
- Giandomenico, S. L., Sutcliffe, M., Lancaster, M. A. Generation and long-term culture of advanced cerebral organoids for studying later stages of neural development. Nature Protocols. 16 (2), 579-602 (2021).
- Velasco, S., et al. Individual brain organoids reproducibly form cell diversity of the human cerebral cortex. Nature. 570 (7762), 523-527 (2019).
- Qian, X., et al. Brain-region-specific organoids using mini-bioreactors for modeling ZIKV exposure. Cell. 165 (5), 1238-1254 (2016).
- Hunter, Z. L., Leeson, H. C., Shaker, M. R., Wolvetang, E. J., Vadlamudi, L. Human induced pluripotent stem cells generated from epilepsy patients for use as in vitro models for drug screening. Stem Cell Research. 60, 102673 (2022).
- Kaur, A., Macip, S., Stover, C. M. An appraisal on the value of using nutraceutical based senolytics and senostatics in aging. Frontiers in Cell and Developmental Biology. 8, 218 (2020).
- Wang, F., et al. Safety and efficacy of dispase and plasmin in pharmacologic vitreolysis. Investigative Ophthalmology & Visual Science. 45 (9), 3286-3290 (2004).
Tags
ABOUT JoVE
Copyright © 2024 MyJoVE Corporation. All rights reserved