
Summary
Abstract
Introduction
Protocol
Representative Results
Discussion
Acknowledgements
Materials
References
Biology
שחזור ואפיון מרוכבים של אקטין-מיקרוטובול עם דינמיקה ומכניקה מונעות מנוע ניתנות לכוונון
ERRATUM NOTICE
Important: There has been an erratum issued for this article. Read more …מאמר זה מציג פרוטוקולים להנדסה ולאפיון רשתות תלת-ממדיות מרוכבות הניתנות לכוונון של חוטי אקטין ומיקרו-טובולים המסתבכים. חומרים מרוכבים עוברים ארגון מחדש אקטיבי ותנועה בליסטית, מונעים על ידי מנועי מיוזין II וקינסין, ומכוונים על ידי הריכוזים היחסיים של אקטין, מיקרוטובולים, חלבונים מוטוריים וקרוסלינקרים פסיביים.
השלד המורכב, המורכב מרשתות אינטראקציה של חוטי אקטין גמישים למחצה ומיקרוטובולים קשיחים, בונה מחדש ומייצר כוחות באמצעות חלבונים מוטוריים כגון מיוזין II וקינסין כדי להניע תהליכים מרכזיים כגון הגירה, ציטוקינזיס, הידבקות ומכנוסנס. בעוד שאינטראקציות אקטין-מיקרוטובול הן המפתח לרבגוניות וליכולת ההסתגלות של השלד הציטוסקולרי, עדיין קיימת הבנה של יחסי הגומלין שלהם עם פעילות המיוזין והקינסין. עבודה זו מתארת כיצד להנדס רשתות תלת-ממדיות מרוכבות הניתנות לכוונון של חוטי אקטין ומיקרוטובולים המסתבכים שעוברים ארגון מחדש פעיל ותנועה בליסטית, המונעים על ידי מנועי מיוזין II וקינזין, ומכוונים על ידי הריכוזים היחסיים של אקטין, מיקרוטובולים, חלבונים מוטוריים והצלבות פסיביות. פרוטוקולים לתיוג פלואורסצנטי של מיקרוטובולים וחוטי אקטין כדי להמחיש בצורה היעילה ביותר ארגון מחדש ותנועה מרוכבת באמצעות הדמיה קונפוקלית מולטי-ספקטרלית מפורטים גם הם. לבסוף, מוצגות התוצאות של שיטות ניתוח נתונים שניתן להשתמש בהן כדי לאפיין באופן כמותי מבנה, דינמיקה ומכניקה שאינם שיווי משקל. יצירה מחדש וחקירה של פלטפורמה ביומימטית מתכווננת זו מספקת תובנה רבת ערך לגבי האופן שבו פעילות מוטורית מצומדת, מכניקה מרוכבת ודינמיקה של נימה יכולים להוביל לתהליכים תאיים רבים, ממיטוזה דרך קיטוב ועד לתחושת מכנו.
השלד הציטוסקולרי הוא רשת מרוכבת דינמית של ביופולימרים מתקשרים המספקת תמיכה מבנית ומכנית לתאים. מנועים מולקולריים קשורים וחלבונים קושרים בונים מחדש ומתאימים את השלד הציטוסקולרי כדי לאפשר לתאים לגדול, לשנות צורה, להתקשות, לנוע ואפילו לרפא את עצמם, מה שמאפשר אינספור תהליכים תאיים החל מהגירה וחלוקה וכלה במכנוסנס 1,2. מעבר לחשיבותו בביופיזיקה תאית, השלד הוא גם דוגמה מובהקת לחומר פעיל עם יישומים פוטנציאליים של חומרים, החל מריפוי פצעים ואספקת תרופות וכלה בסינון ורובוטיקה רכה 1,3,4,5,6,7,8,9.
שני המאפיינים המרכזיים המעניקים לשלד הציטוסקלטון את המגוון המבני והמכני הייחודי שלו ואת הרב-תכליתיות שלו הם: 1) האופי המורכב שלו, הכולל חוטי חלבון מרובים המקיימים אינטראקציה, כגון חוטי אקטין גמישים למחצה ומיקרוטובולים קשיחים, כמו גם חלבוני הקישור וההצלבה הקשורים אליהם 3,5,10; ו-2) יכולתו לבנות מחדש, לנוע, לגרס ולבצע עבודה באמצעות מנועים צורכי אנרגיה, כגון מיוסינים וקינסינים, תוך דחיפה ומשיכה של החלבונים החוטמים 1,7,11,12,13. בעוד שמורכבות אלגנטית זו מאפשרת לשלד הציטוסקולרי לתווך תהליכים מגוונים כמו תנועתיות תאים, ציטוקינזיס וריפוי פצעים 3,6,7,11, היא פוגעת ביכולתם של חוקרים לשחזר את מאפייני החתימה in vivo של השלד במערכת מבחנה משוחזרת.
מאמצי שחזור הגבול הנוכחיים מתמקדים במרוכבים של חוטי אקטין ומיקרוטובולים מסובכים ומקושרים 3,10,14,15,16,17, רשתות אקטומיוזין מחוללות כוח 2,8,18,19,20,21, ונמטיקה פעילה המונעת על ידי קינזין-מיקרוטובול אינטראקציות 22,23,24,25,26. הודגם כי מרוכבים מסוג אקטין-מיקרוטובול במצב יציב מציגים תכונות מכניות מתפתחות15,16,27, כגון ניידות נימה משופרת וקשיחות מוגברת בהשוואה למערכות חד-רכיביות 27. מחקרים על מערכות אקטומיוזין במבחנה דיווחו על מגוון רחב של תכונות מבניות ודינמיות התלויות בריכוזי אקטין, מיוזין וקרוסלינקרים 28,29,30,31. לדוגמה, עם הצלבה מספקת, רשתות אקטומיוסין עוברות התכווצות בקנה מידה גדול וגסות 2,28,30,32,33,34,35,36, בעוד שללא crosslinkers, רשתות מציגות זרימה מהירה, מערערת יציבות וקרע 19,29 . דווח כי נמטיקה פעילה מבוססת מיקרוטובול משוחזרת המשתמשת באשכולות של מנועי קינזין כדי להצליב ולמשוך צרורות מיקרוטובולים הפגינו זרימות טורבולנטיות ארוכות טווח, הרחבה, אבזם, שבירה וריפוי 12,22,23,24,25,37,38,39,40,41, 42,43,44,45,46,47.
לאחרונה, הוכח כי חומרים מרוכבים של אקטין-מיקרוטובול המונעים על ידי מיני-חוטים של מיוזין II מובילים להתכווצות מסודרת יותר ולשלמות הרשת בהשוואה לזרימה הלא מסודרת ולקרע ברשת שרשתות אקטומיוזין ללא crosslinkers מציגות 17,26,48. יתר על כן, השילוב של חוסן מרוכב וייצור כוח ממוטב כאשר אקטין ומיקרוטובולים נמצאים בריכוזים דומים. תכונות מתפתחות עיקריות באזור זה של מרחב הפורמולציה כוללות חוזק מכני משופר 26, תנועה מתואמת של אקטין ומיקרוטובולים26, התכווצות מתמשכת קבועה וארגון מחדש מזוקלי17.
כאן, פרוטוקולים מתוארים כדי להנדס ולכוונן מרוכבים מסתבכים ומקושרים של מיקרוטובולים וחוטי אקטין שנדחקים אל מחוץ לשיווי משקל על-ידי מיני-חוטים של מיוזין II וצבירי קינסין הפועלים על חוטי אקטין ומיקרוטובולים, בהתאמה (איור 1). ניתן לכוונן את הדינמיקה, המבנה והמכניקה של סוג זה של חומרים מרוכבים על ידי הריכוזים היחסיים של החוטים, המנועים וההצלבות כדי להציג מרחב פאזה עשיר של זרימה אדבקטיבית וטורבולנטית, כיווץ איזוטרופי, האצה, האטה, דה-ערבוב, התקשות, הרפיה וקריעה. המיקוד של עבודה זו הוא על הכנה וכוונון של סוג זה של מרוכבים cytoskeletal פעיל. עם זאת, כדי לסייע לחוקרים לבחון ולאפיין את החומרים המרוכבים הפעילים המתוארים, מפורטות גם שיטות הדמיה יעילות באמצעות מיקרוסקופיה קונפוקלית מולטי-ספקטרלית. לבסוף, מוצגות תוצאות של שיטות ניתוח חישוביות מרכזיות שניתן להשתמש בהן למדידת הדינמיקה, המבנה והמכניקה של החומרים המרוכבים. אנו מעודדים את החוקרים לאמץ שיטות אלה - הכוללות מיקרוסקופיה דינמית דיפרנציאלית (DDM), תיקון אוטומטי של תמונה מרחבית (SIA) ווולוצימטריה של תמונת חלקיקים (PIV) - מכיוון שהן עברו אופטימיזציה כדי לאפיין את הדינמיקה המורכבת ואת המגוון המבני של החומרים המרוכבים 17,26,49.
השלבים המתוארים להלן מתמקדים בהכנת החומרים המרוכבים ובהדמיתם באמצעות מיקרוסקופיה קונפוקלית. פרוטוקולים המתארים ניתוח נתונים לאחר רכישה ומדידות פינצטה אופטיות ניתן למצוא בעבודות קודמות 17,26,48,50, ומסופקים על פי בקשה. כל החומרים מפורטים בטבלת החומרים המסופקת.
1. הכינו כיסויים מבודדים ושקופיות מיקרוסקופ למניעת ספיחה של חלבונים למשטחי התא
הערה: זהו תהליך של יומיים. ניתן להכין שקופיות מסולפות עד חודש לפני השימוש.
- הניחו את הכיסויים מס' 1 (24 מ"מ x 24 מ"מ) ואת מגלשות המיקרוסקופ (1 אינץ' x 3 אינץ') במדף ייעודי שיתאים למנקה הפלזמה. מניחים את המתלה במנקה פלזמה ופועלים במשך 20 דקות.
- העבירו את הכיסויים והמגלשות למתלה חדש המיועד לשימוש עם סילאן בלבד והניחו את המדף במיכל זכוכית לניקוי הכוסות כמתואר להלן.
- יש לטבול את הכיסויים ולהחליק ב-100% אצטון למשך שעה אחת. יש לטבול את הכיסויים ולהחליק ב-100% אתנול למשך 10 דקות.
- יש לטבול את הכיסויים והמגלשות במים שעברו דה-יוניזציה (DI) למשך 5 דקות. חזור על שלבי הניקוי פעמיים נוספות.
- טבלו את הכיסויים והשקופיות ב-0.1 M KOH שהוכן טרי למשך 15 דקות. יש לטבול את הכיסויים והשקופיות ב-DI טרי למשך 5 דקות. חזור על שלב זה פעמיים נוספות.
- כיסויים יבשים באוויר ומחליקים למשך 10 דקות. טפלו בכיסויים ובמגלשות מנוקים עם סילאן כדי לייצר משטחים הידרופוביים כמתואר להלן.
הערה: בצע את השלבים הבאים במכסה אדים.- יש לטבול את הכיסויים המיובשים ולהחליק ב-2% סילאן (מומס בטולואן) למשך 5 דקות. השתמשו במשפך כדי לשפוך את הסילאן בחזרה לבקבוק הייעודי שלו לשימוש חוזר עד חמש פעמים.
- יש לטבול את הכיסויים ולהחליק ב-100% אתנול למשך 5 דקות. החלף אתנול באתנול טרי. יש לטבול את הכיסויים והשקופיות למשך 5 דקות.
- יש לטבול את הכיסויים והשקופיות ב-DI טרי למשך 5 דקות. חזרו על שלב שטיפת האתנול וה-DI פעמיים נוספות באמצעות אתנול טרי ו-DI בכל פעם. כיסויים יבשים באוויר ומחליקים למשך 10 דקות.
2. הכנת אקטין-מיקרוטובול פעיל מרוכב מונע על ידי מיני-חוטים מיוזין
- הסר מיוזין לא פעיל באמצעות קשירת נימה של אקטין ובצע משיכה כלפי מטה באמצעות ultracentrifugation כמתואר להלן.
- פולימר אקטין לתוך חוטים. באמצעות מיקרופיפט מדויק וקצות פיפט סטריליים, שלב בצינור מיקרוצנטריפוגה: 1.87 μL של DI, 1.3 μL של 10x G-buffer, 1.3 μL של 10x F-buffer, 1.63 μL של 4 M KCl, 4.53 μL של אקטין (47.6 μM), ו 1.08 μL של 100 μM phalloidin.
הערה: כדי להבטיח פילמור מספיק, ריכוז האקטין ויחס הטוחנות אקטין:פאלוידין צריך להיות 18.4 מיקרומטר ו-2:1, בהתאמה. - מעבירים בעדינות את התמיסה למעלה ולמטה כדי לערבב ואז מניחים על קרח בחושך למשך ≥1 שעות. קירור אולטרה-צנטריפוגה ל-4°C. מוציאים מיוזין אליקוט מ-80 מעלות צלזיוס ומניחים על קרח.
הערה: השלם את שלב 2.2 בשלב זה בזמן שאקטין עובר פילמור. - לאחר ≥1 שעות של פילמור אקטין, יש להוסיף 1.3 μL של 10 mM ATP ו-2 μL של 19 μM מיוזין לאקטין הפולימרי.
הערה: יחס הטוחנות אקטין:מיוזין צריך להיות >5 כדי להבטיח הסרה מספקת של מנועי מיוזין לא פעילים (כלומר, ראשים מתים). - העבירו בעדינות את התמיסה למעלה ולמטה כדי לערבב. העבר לצינור כיתה אולטרה-צנטריפוגה.
צנטריפוגה ב-4 מעלות צלזיוס ו-121,968 x גרם למשך 30 דקות.
- פולימר אקטין לתוך חוטים. באמצעות מיקרופיפט מדויק וקצות פיפט סטריליים, שלב בצינור מיקרוצנטריפוגה: 1.87 μL של DI, 1.3 μL של 10x G-buffer, 1.3 μL של 10x F-buffer, 1.63 μL של 4 M KCl, 4.53 μL של אקטין (47.6 μM), ו 1.08 μL של 100 μM phalloidin.
- הכינו רשת מרוכבת משותפת של חוטי אקטין ומיקרוטובולים כמתואר להלן.
הערה: יש להתחיל 30 דקות לפני סיבוב המיוזין (שלב 2.1.4).- הגדר בלוק חום ל 37 מעלות צלזיוס. השתמש במיקרופיפט מדויק ובקצות פיפט סטריליים כדי להוסיף את הדברים הבאים לצינור מיקרוצנטריפוגה: 13.9 μL של PEM, 3 μL של 1% Tween20, 1.55 μL של 47.6 μM אקטין, 0.36 μL של 34.8 μM R-actin, 0.3 μL של 250 mM ATP, 0.87 μL של 100 μM phalloidin, 1.91 μL של 5-488-tubulin, 0.3 μL של 100 mM GTP, ו-0.75 μL של 200 μM טקסול, לנפח כולל של 23 μL.
הערה: הריכוזים של אקטין וטובולין המפורטים הם עבור מרוכב עם 2.9 μM אקטין ו 2.9 μM טובולין. ריכוז החלבון הכולל הוא c =c A + c T = 5.8 μM ושבר אקטין טוחן הוא c A/(c A + cT) = Φ A = 0.5. ראה שלב 2.5 כדי להתאים ערכים אלה. - צינורו בעדינות את התמיסה למעלה ולמטה כדי לערבב ולהניח על בלוק חום של 37 מעלות צלזיוס המוגן מפני אור למשך שעה אחת.
- הגדר בלוק חום ל 37 מעלות צלזיוס. השתמש במיקרופיפט מדויק ובקצות פיפט סטריליים כדי להוסיף את הדברים הבאים לצינור מיקרוצנטריפוגה: 13.9 μL של PEM, 3 μL של 1% Tween20, 1.55 μL של 47.6 μM אקטין, 0.36 μL של 34.8 μM R-actin, 0.3 μL של 250 mM ATP, 0.87 μL של 100 μM phalloidin, 1.91 μL של 5-488-tubulin, 0.3 μL של 100 mM GTP, ו-0.75 μL של 200 μM טקסול, לנפח כולל של 23 μL.
- הכן תאי דגימה לניסויי הדמיה קונפוקליים כמתואר להלן.
הערה: השלם את שלבים 2.1.4 ו- 2.2.2 במהלך תקופות המתנה.- הניחו שתי שקופיות מבודדות זו לצד זו על פלטה חמה (כבויה), הניחו שתי רצועות של סרט איטום תרמופלסטי על פני השקופיות ~ 3 מ"מ זו מזו, והניחו שני כיסויים מבודדים מעל סרט האיטום התרמופלסטי כדי ליצור תא דגימה.
- סובבו את הפלטה החמה בהגדרה נמוכה עד שהכיסויים ייקשרו היטב לשקופיות עם סרט איטום תרמופלסטי מומס (~1-2 דקות). לחץ כלפי מטה בלחץ אחיד כדי להבטיח הדבקה תוך שמירה על מרווח של ~ 100 מיקרומטר בין שני המשטחים.
- הסר את התאים וכבה את הפלטה החמה. תאי תוויות עם (+) ו- (-). התא (+) יהיה עבור הדגימה הפעילה (עם מיוזין) והתא (-) יהיה הבקרה (ללא מיוזין). ודא שכל תא יכול להכיל ≤10 μL של נוזל.
- הכן דוגמאות לתמונה כמתואר להלן.
הערה: חשוב להשלים שלב זה מיד לאחר השלמת שלבים 2.1 ו- 2.2.- הסר בזהירות את דגימת המיוזין-אקטין מהאולטרה-צנטריפוגה (שלב 2.1.4) ומיד העלה את 7.5 μL העליונים של הסופר-סנטרנט והעבר לצינור מיקרוצנטריפוגה חדש.
- הסר את דגימת האקטין-מיקרוטובול מבלוק החום וערבב בעדינות 1.5 μL של 10x D-גלוקוז, 1.5 μL של 10x GOC, ו-1.5 μL של 1 mM blebbistatin. חלק את התמיסה לשני אליקוטים של 13.7 μL ותווית כ- (+) ו- (-).
- ערבבו 1.28 μL של הסופר-נאטנט משלב 2.4.1 עד (+) אליקוט. מערבבים ב-1.28 μL של DI לאליקוט (-). הזרימו באיטיות כל תמיסה לתא המתאים (שלב 2.3) באמצעות פעולה נימרית. היזהר לא להכניס בועות אוויר לערוץ.
- אטמו את שני הקצוות הפתוחים של כל תעלה באמצעות דבק אפוקסי או UV המתייבש במהירות. ודא שהדבק יבש לחלוטין לפני הנחתו על המיקרוסקופ. תמונה מיידית כמתואר בשלב 3.
הערה: לדבק UV יש יתרון מכיוון שהוא מרפא כמעט באופן מיידי עם חשיפה לקרינת UV. עם זאת, מכיוון ש-blebbistatin רגיש ל-UV, חשוב להאיר את הדבק באופן מקומי בלבד (בשולי תא הדגימה) באמצעות שרביט UV קטן כדי למנוע השבתה של ה-blebbistatin.
- אופציונלי: לשנות את ריכוזי החלבון כדי לכוונן את הדינמיקה והמבנה של החומרים המרוכבים.
הערה: השלבים הבאים מוצעים לשינויים בשלבים לעיל כדי לשנות את ריכוזי האקטין, המיקרוטובולים והמיוזין, אם תרצה בכך.- בצע את השלבים המתוארים לעיל למעט השינויים הבאים בשלבים 2.2.1 ו- 2.4.3.
- כדי לשנות את ריכוזי האקטין והמיקרוטובולים, ובכך להתאים את c ו- ΦA, להגדיל או להקטין את נפח האקטין, R-אקטין ו- 5-488-טובולין המשמשים בשלב 2.2.1, לפי הצורך26. כאשר משנים את ריכוז האטין, התאימו את ריכוזי הטוחנות R-אקטין ופלואידין באופן פרופורציונלי כדי לשמור על אותם יחסים טוחנות עם אקטין. התאם את עוצמת הקול של PEM כך שהנפח הסופי של התערובת יישאר 23 μL. כל שאר הנפחים והריכוזים של הרכיבים נשארים זהים.
- כדי לשנות את ריכוז המיוזין, התאם את נפח המיוזין שנוסף לאליקוט (+) בשלב 2.4.3 לפי הצורך. כוונן את עוצמת הקול של DI שנוספה ל-(-) aliquot בהתאם. התאם את עוצמת הקול של PEM בשלב 2.2.1 כדי לקחת בחשבון את העלייה או הירידה בעוצמת הקול של מיוזין (+) ו-DI (-), כדי להבטיח שהנפח הסופי של כל דגימה ((+) ו- (-)) הוא 14.98 μL.
3. הדמיה ואפיון של חומרים מרוכבים פעילים באמצעות מיקרוסקופיה קונפוקלית
- כדי לדמות חומרים מרוכבים של אקטומיוזין-מיקרוטובול שהוכנו בשלב 2, השתמש במיקרוסקופ קונפוקלי לסריקת לייזר (LSCM), או במיקרוסקופ דומה, עם מטרת טבילת שמן 60x 1.4 NA. כדי לדמיין בו-זמנית חוטי אקטין ומיקרוטובולים בתעלות פלואורסצנטיות נפרדות, השתמש בלייזר של 561 ננומטר עם מסנני עירור/פליטה של 565/591 ננומטר ובלייזר של 488 ננומטר עם מסנני עירור/פליטה של 488/525 ננומטר.
- מקם את תא הדגימה על המיקרוסקופ כך שערוץ הבקרה ממוקם ישירות מעל המטרה. וודאו שיש ממשק שמן בין המטרה לבין הכיסוי.
- השתמש בפקדי הבמה כדי למקד את קומפוזיט הבקרה, ולאחר מכן מצא את שני המשטחים של תא הדגימה. הזז את מיקום z למרכז תא הדגימה. בדוק את נוכחותן של רשתות נימה ברורות כפי שמוצג באיור 2.
- עדיין מדמיין את תא הבקרה, התאם את העוצמה של כל לייזר כדי לאפשר הדמיה בו זמנית של חוטי אקטין ומיקרוטובולים. לשמור על עוצמת הלייזר הנמוכה ביותר האפשרית כדי למנוע פוטו-הלבנה (שכיחה יותר בתעלת האטין) ולדמם דרך (בדרך כלל מהמיקרוטובולים לתעלת האטין).
- כדי לאפיין את דגימת הבקרה הלא פעילה, אסוף שלוש סדרות זמן (סרטונים) של תמונות בגודל 256 x 256 פיקסלים מרובעים (213 μm x 213 μm) בקצב של 2.65 ת/ש, ובסך הכל ≥1000 פריימים. אסוף כל סדרת זמן באזור אחר של תא הדגימה המופרד על ידי ≥500 מיקרומטר. ודא שיש תנועה מינימלית הניתנת לזיהוי וללא זרימה או ארגון מחדש.
- כבה את הלייזר בגודל 488 ננומטר והשתמש בפקדי הבמה כדי לעבור לתא (+).
- באמצעות לייזר 568 ננומטר, דמיינו את המיקרוטובולים בערוץ (+) כדי להבטיח היווצרות תקינה של הרשת (איור 2) וזהו את המרכז הצירי של תא הדגימה (שעשוי להיות שונה ממיקום z המרכזי של תא הבקרה).
- הפעל את הלייזר 488 ננומטר וחזור על שלב 3.5 לעיל עם השינויים הבאים. אסוף סדרות זמן למשך עד 45 דקות, ועצור את הרכישה כאשר הדגימה זזה מחוץ לשדה הראייה, נקרעת או מלבינים. הקלט 5-10 סדרות זמן ועקוב אחר הזמן שבו כל סדרת זמן מתחילה ביחס לתחילת סדרת הזמן הראשונה.
- נתחו נתונים באמצעות DDM, SIA ו-PIV כמתואר באיור 3, איור 4, איור 5, ובעבר 17,48,50,51.
הערה: לייזר 488 ננומטר מפעיל באופן מקומי את פעילות המיוזין ATPase על ידי ביטול ההפעלה של blebbistatin, כך שיש להפעיל אותו רק בתחילת רכישת הנתונים, כך ש- t = 0 נמצא בתחילת סדרת הזמן. פרמטרי רכישה אלה מותאמים לניתוח מיקרוסקופיה דינמית דיפרנציאלית (DDM) כפי שנעשה בעבר26.
4. הכנת חומרים מרוכבים פעילים של אקטין-מיקרוטובול המונעים על ידי מנועי קינזין
הערה: השלבים הבאים יוצרים מרוכבים של אקטין-מיקרוטובול המונעים מחוץ לשיווי משקל על-ידי מנועי קינזין או שילוב של קינזין ומיוזין50.
- הכן מנועי קינסין ומיוזין כמתואר להלן.
- אם אתה משלב מיוזין, בצע את שלב 2.1.
- כדי ליצור אשכולות מנוע קינסין הקושרים ומפעילים כוחות בין זוגות של מיקרוטובולים, השתמש במיקרופיפט ובקצות פיפט סטריליים כדי להוסיף את הדברים הבאים לצינור מיקרוצנטריפוגה סטרילי של 1.5 מ"ל: 1.16 μL PEM, 2.74 μL 8.87 μM דימרים קינסין, 7.29 μL 83.3 μM NeutrAvidin, 0.81 μL 2mM DTT . מערבבים בעדינות על ידי צינורית התמיסה למעלה ולמטה ודגירה מוגנת מפני אור (השתמש בצינור מיקרוצנטריפוגה שחור או עטוף בנייר כסף) במשך 30 דקות בטמפרטורה של 4 מעלות צלזיוס.
הערה: היחס הטוחן של דימרים קינסין ל-NA הוא 1:25.
- בצע את שלב 2.3 כדי להכין תאי דגימה וליצור שלושה תאים במקום שניים. בצע שלב זה במהלך דגירה קינזין (שלב 4.1.2) ואולטרה-צנטריפוגציה של מיוזין (שלב 4.1.1).
- הכינו רשת מרוכבת משותפת של חוטי אקטין ומיקרוטובולים.
- הגדר בלוק חום ל 37 מעלות צלזיוס. השתמש בקצוות מיקרופיפט ופיפטה סטריליים כדי להוסיף את הדברים הבאים לצינור מיקרוצנטריפוגה סטרילי של 1.5 מ"ל: 3.21 μL PEM, 4.5 μL של 1% Tween20, 2.18 μL של 47.6 μM אקטין, 3.46 μL של 5-R-tubulin, 4.5 μL של 100 mM ATP, 4.5 μL של 10 mM GTP, 1.13 μL של 200 μM טקסול, ו 1.57 μL של 20 μM 488-phalloidin. ודא שהנפח הכולל הוא 25 μL.
- העבירו בעדינות את התמיסה למעלה ולמטה כדי לערבב ולהניח על בלוק החום של 37°C המוגן מפני אור למשך שעה אחת. הסר את הצינור מבלוק החום והשתמש במיקרופיפטה כדי לערבב בעדינות 0.84 μL של 100 μM phalloidin. דגירה במשך 5-10 דקות בטמפרטורת החדר, מוגן מפני אור.
הערה: הוספת phalloidin בשלב זה, ולא בשלב 4.3.1, משפרת את הסימון הפלואורסצנטי של חוטי אקטין, שכן 488-phalloidin אינו צריך להתחרות עם phalloidin ללא תווית עבור אתרי קשירת אקטין.
- הכן חומרים מרוכבים פעילים להדמיה קונפוקלית.
- הוסף 1.13 μL של 200 μM blebbistatin, 1.35 μL של 10x Glu, ו- 1.35 μL של 10x GOC לתמיסה משלב 4.3.2 וערבב בעדינות על ידי pipetting למעלה ולמטה. חלק את התמיסה לשלושה אליקוטים של 10 μL ותייג כ- (K), (K+M) ו- (-).
- יש לערבב 2.54 μL של מיוזין משלב 2.1.4 ועד לאליקוט (K+M). ערבבו ב-2.54 μL PEM ל-(K) ו-(-) אליקוטים.
- השתמש במיקרו-פיפטה ובקצות פיפט סטריליים כדי להוסיף 2.5 μL של אשכולות קינסין משלב 4.1.2 ל-(K) ו-(K+M) אליקוטים. פיפט למעלה ולמטה כדי לערבב. ערבבו ב-2.5 μL PEM ל-(-) באותה טכניקה.
הערה: הריכוזים של אקטין וטובולין המפורטים הם עבור מרוכב עם 2.32 μM אקטין ו 3.48 μM טובולין. ריכוז החלבון הכולל הוא c = cA + c T = 5.8 μM ושבר אקטין טוחן הוא c A / (c A + cT) = Φ A = 0.4. ריכוזי קינסין ומיוזין הם 0.35 מיקרומטר ו-0.47 מיקרומטר, בהתאמה. עיין בשלב 2.5 לקבלת הנחיות כלליות להתאמת c A,C T, C ו- ΦA. - באמצעות מיקרופיפטה, מזרימים באיטיות כל תמיסה לתעלה המתאימה של תאי הדגימה המוכנים (שלב 4.2) באמצעות פעולה נימרית. דחפו מטה באיטיות ובעדינות רבה על הפיפט כדי לא להכניס בועות אוויר לערוץ.
- אטמו את שני הקצוות הפתוחים של כל תעלה באמצעות דבק אפוקסי המתייבש במהירות או בדבק הניתן לריפוי UV. ודא שהדבק יבש לחלוטין לפני הנחתו על המיקרוסקופ.
הערה: חשוב ששלב זה ייעשה במהירות כדי למזער את הזמן שבו הקינזין פועל מבלי להיות מנוטר. מסיבה זו, אפוקסי שמרפא תוך דקה אחת (ולא 5 או 10 דקות) מומלץ. דבק הניתן לריפוי UV הוא יתרון בהקשר זה מכיוון שהוא מרפא כמעט באופן מיידי עם חשיפה לקרינת UV.
- התמונה הכינה דוגמאות באופן מיידי, לאחר שלב 3, למעט השינויים החשובים הבאים. מכיוון שקינזין אינו נשלט על ידי הפעלת אור, הוא מתחיל לעבוד מיד לאחר שלב 4.4.3, לכן סמן הפעם כ- t = 0. כדי לדמות את הערוצים המורכבים קרוב ככל האפשר למצב הלא פעיל ההתחלתי (t = 0), דמיין תחילה את הערוצים (K) ו- (K+M) וציין את הזמן שחלף בין שלב 4.4.3 לתחילת רכישת הנתונים (שלב 3.8). בפועל, הזמן שחלף הוא ~ 5 דקות.
5. שילוב קרוסלינקרים פסיביים בחומרים מרוכבים אקטיביים
הערה: שלבים אלה מתארים כיצד להשתמש בתת-יחידות אקטין וטובולין שעברו ביוטינילציה וב-NeutrAvidin (NA) כדי לקשר באופן פסיבי בין אקטין לאקטין (A-A) או מיקרוטובולים למיקרוטובולים (M-M) בחומרים המרוכבים הפעילים המתוארים בשלב 4.
- הכינו קומפלקסים של A-A או M-M עם חלבונים שעברו ביוטינילציה (ביוטין-אקטין או ביוטין-טובולין), NA וביוטין ביחס של 2:2:1 ביוטין-אקטין/טובולין:ביוטין:NA. התחל תהליך זה לפני שלב 4 .
- עבור קרוסלינקרים של A-A, השתמש במיקרופיפט ובקצות פיפט סטריליים כדי להוסיף 2 μL של 11.6 μM ביוטין-אקטין, 1.39 μL של 8.33 μM NA, 2.27 μL של 1.02 μM ביוטין, ו-4.34 μL של PEM לצינור מיקרוצנטריפוגה. מערבבים בעדינות על ידי צנרת למעלה ולמטה.
- עבור קרוסלינקרים M-M, השתמש במיקרופיפט ובקצות פיפט סטריליים כדי להוסיף 1.86 μL של 4.55 μM ביוטין-טובולין, 1.11 μL של 8.33 μM NA, 1.82 μL של 1.02 μM ביוטין, ו-5.21 μL של PEM לצינור מיקרוצנטריפוגה. מערבבים בעדינות על ידי צנרת למעלה ולמטה.
- יש לעטוף את הצינוריות משלב 5.1.1 ו/או 5.1.2 בסרט איטום תרמופלסטי כדי ליצור אטימה אטומה למים. מניחים ברפסודת ציפה באמבט סוניקטור מבוקר טמפרטורה המוגדר ל-4 מעלות צלזיוס.
- סוניקט במשך 90 דקות ב-4 מעלות צלזיוס. בפועל עדיף לשים את הסוניקטור בחדר קר ולהוסיף חבילות קרח לאמבטיית הסוניקציה כדי לשמור על טמפרטורה נמוכה.
- כדי לשלב קומפלקסים של crosslinker בדגימות להדמיה, בצע את שלב 4.3, ושנה את שלב 4.3.1 כמתואר להלן עבור קישור צולב A-A (שלב 5.2.1) או M-M crosslinking (שלב 5.2.2).
- עבור קישור צולב A-A, שלב את הדברים הבאים בצינור מיקרוצנטריפוגה: 1.94 μL של PEM, 4.50 μL של 1% Tween20, 2.18 μL של 47.6 μM אקטין, 3.46 μL של 45.5 μM 5-R-tubulin, 1.13 μL של A-A crosslinkers (שלב 5.1.1), 4.50 μL של 100 mM ATP, 4.50 μL של 10 mM GTP, 1.13 μL של 200 μM טקסול, ו-1.57 μL של 20 μM 488-phalloidin. ודא שהנפח הכולל הוא 25 μL.
- עבור קישור צולב M-M, שלב את הדברים הבאים בצינור מיקרוצנטריפוגה: 1.97 μL של PEM, 4.50 μL של 1% Tween20, 2.18 μL של 47.6 μM אקטין, 3.76 μL של 45.5 μM 5-R-tubulin, 1.13 μL של דילול 1:4 של M-M crosslinkers (שלב 5.1.2), 4.50 μL של 100 mM ATP, 4.50 μL של 10 mM GTP, 1.13 μL של 200 μM טקסול, ו-1.57 μL של 20 μM 488-phalloidin. ודא שהנפח הכולל הוא 25 μL.
- בצע את השלבים 4.3.2-4.5 עם הריכוזים הספציפיים עבור יחס טוחנת crosslinker:actin של RA = 0.02 ויחס טוחנת crosslinker:tubulin של RT = 0.005. ערכי R A ו-R T אלה גורמים לאורכים דומים בין צולבים לאורך חוטי אקטין ומיקרוטובולים (d A 60 ננומטר ו-d MT 67 ננומטר), הנאמדים באמצעות d A =I monomer/2R A
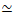 , כאשר מונומר I הוא אורכו שלמונומר אקטין,ו-d MT = I ring/26RT, כאשרטבעת I היא אורך של טבעת של 13 טובולינים 15, 17.
, כאשר מונומר I הוא אורכו שלמונומר אקטין,ו-d MT = I ring/26RT, כאשרטבעת I היא אורך של טבעת של 13 טובולינים 15, 17.
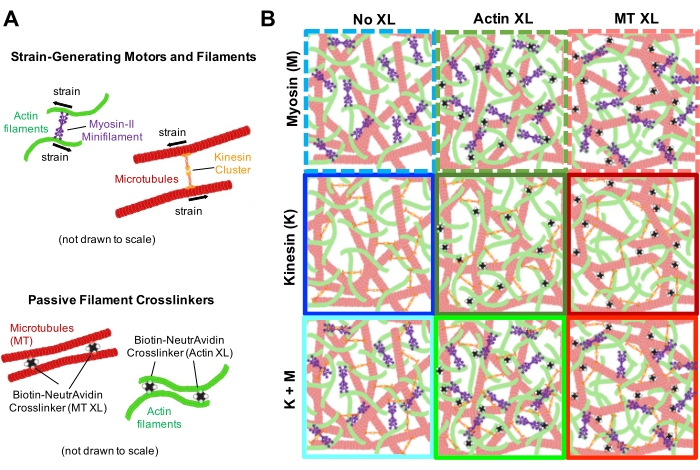
כדי לקבוע הכנה מוצלחת של חומרים מרוכבים פעילים (איור 1), וכדי לאפיין את הדינמיקה והמבנה שלהם, נעשה שימוש במיקרוסקופ פלואורסצנטי לסריקת לייזר עם לפחות שני ערוצים פלואורסצנטיים כדי לדמיין את חוטי האקטין והמיקרוטובולים בו-זמנית (איור 2 ואיור 6). כל חוטי האקטין והמיקרוטובולים בחומרים המרוכבים מסומנים בדלילות, במקום סימום בחוטים בהירים, כפי שנעשה לעתים קרובות במחקרים במבחנה. שיטה זו מבטיחה שהדינמיקה והמבנה הנמדדים מייצגים את המרוכב עצמו ולא את העוקבים הנוצרים בתנאים שונים מהחומרים המרוכבים. מסיבה זו, בדרך כלל לא ניתן לפתור חוטי אקטין בודדים ומיקרוטובולים, אלא תמונות מתארות את מבנה הרשת המזוקלית (איור 2 ואיור 6).
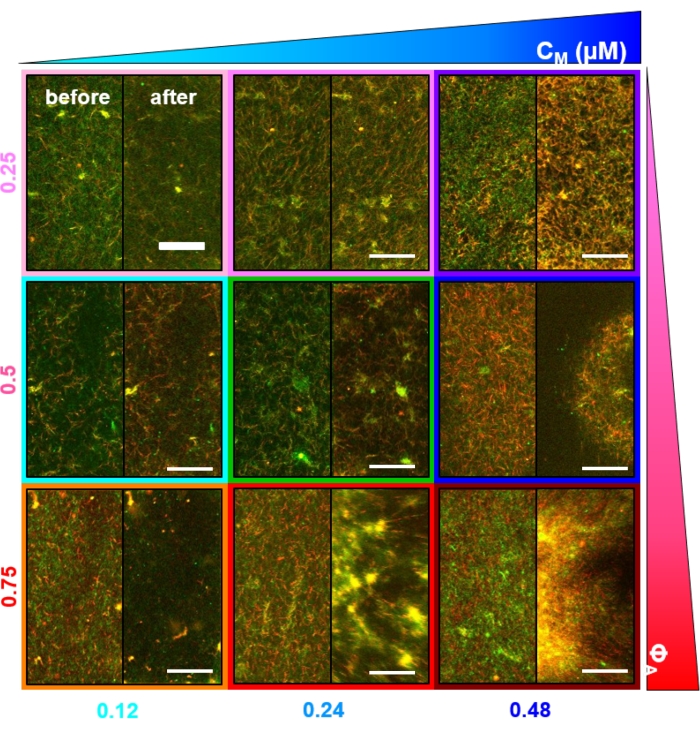
גישת תיוג זו עברה אופטימיזציה עבור ניתוחי התאמה אוטומטית של תמונה מרחבית (SIA) ומיקרוסקופיה דינמית דיפרנציאלית (DDM) הבוחנים את הדינמיקה והמבנה במרחב פורייה הדדי (איור 4, איור 5 ואיור 8)52,53,54,55. ניתן להשתמש בוולוצימטריה של תמונת חלקיקים (PIV) גם כדי לתאר ולאפיין שדות דינמיקה וזרימה (איור 3 ואיור 7), אך היא דורשת שילוב פיקסלים (רזולוציה מרחבית נמוכה יותר) והפרשי זמן השהיה גדולים יותר (רזולוציה זמנית נמוכה יותר) מאשר SIA ו-DDM כדי לסלק וקטורים שגויים הנובעים מרעש בתמונות הצפופות בעלות האות הנמוך. עם זאת, PIV מומלץ לבדיקה איכותית של שדות זרימה ולאימות תוצאות DDM (איור 4 ואיור 8)26,50.
אפיון מדגמי של הרשתות המתוארות באמצעות ניתוחים אלה (כלומר, DDM, SIA, PIV) מסופק כדי לסייע לחוקרים לאמץ ניתוחים דומים כדי למדוד ולאפיין את הדגימות שלהם. עם זאת, תיאורים מפורטים של טכניקות אלה הם מחוץ לתחום של עבודה זו. לתיאורים מפורטים של אופן ביצוע DDM במערכות אלה ודומות אחרות, כולל קוד Python ידידותי למשתמש, עיין בעבודות קודמות 17,26,49,50 וההפניות שם בפנים. לקבלת פרטים לגבי אופן ביצוע SIA ו- PIV במערכות המתוארות כאן, הקורא מופנה ליצירות קודמות 17,50.
יש לבצע מספר פקדים, המתוארים להלן, כדי להבטיח שהמרוכבים פועלים כצפוי. מרוכב ללא מיוזין או קינזין צריך להיראות סטטי בעיקרו עם תנודות תרמיות מינימליות או סחף. חוטי אקטין ומיקרוטובולים צריכים להיראות מסתבכים ומפוזרים בצורה הומוגנית, עם קשירה, צבירה או הפרדת פאזה מינימלית של אקטין ומיקרוטובולים לאורך שדה ראייה של ~200 מיקרומטר x 200 מיקרומטר (איור 2, משמאל)17. יש לצפות לתוצאה דומה עבור חומרים מרוכבים המכילים מיוזין אך אינם חשופים לאור של 488 ננומטר (כדי להשבית את הבלביסטטין).
לאחר שילוב של מיוזין וחשיפה לאור של 488 ננומטר, החומרים המרוכבים עוברים התכווצות שהיא במידה רבה איזוטרופית ודומה לאקטין ולמיקרוטובולים, כפי שניתן לראות בתמונות מיקרוסקופ שצולמו לפני ואחרי פעילות המיוזין (איור 2), כמו גם בשדות זרימת PIV מתאימים לזמנים משתנים במהלך הפעילות (איור 3). כדי לקבוע אם התנועה היא בליסטית, מפוזרת, תת-דיפוזיבית וכו', זמן ההפחתה האופייני τ(q) שנקבע מ-DDM מוערך כפונקציה של וקטור גל (כלומר, מרחב הדדי). ראה כפי שתואר בפירוט קודםלכן 17,26,49. איור 4 גם מדגים כיצד להשתמש ב-DDM כדי לאפיין את החומרים המרוכבים האלה. שינוי קנה מידה של חוק הכוח τ(q)~1/vq β, עם β = 1, מציין תנועה בליסטית עם מהירות v. לשם השוואה, β = 2 מייצג דינמיקה דיפוזיבית כאשר v הוא מקדם הדיפוזיה. כל החומרים המרוכבים הפעילים מפגינים קנה מידה בליסטי (איור 4A) עם מהירויות שמכוונות על-ידי ריכוזי האקטין והמיוזין (איור 4B), ושיכולות להשתנות בזמן במהלך הפעילות, בין אם הן מאיצות או מאטות (איור 4C,D).
ניתן לאפיין ארגון מחדש של רשתות ואשכולות, הנראים באיור 2 ובולטים יותר עבור ריכוזים גבוהים יותר של אקטין ומיוזין, באמצעות SIA, כפי שתואר באיור 5, ותואר קודם לכן 17,48,50. בקצרה, אורך מתאם ξ, שהוא מדד לגודל האופייני של תכונות בתמונה, ניתן לקבוע על ידי התאמת כל עקומת התאמה אוטומטית של עוצמה מרחבית g(r) לפונקציה מעריכית של מרחק r בין פיקסלים. פסגות g(r) גדולות יותר שנמשכות למרחקים ארוכים יותר מצביעות על תכונות מבניות גדולות יותר (כלומר, קשירה, אשכולות של החוטים הבודדים). כפי שניתן לראות באיור 5, עבור שברי אקטין וריכוזים גבוהים יותר של מיוזין, ארגון מחדש וצבירה משמעותיים משתקפים בעלייה ב-ξ לאורך זמן.
ניתן למדוד את התכונות הצמיגיות והתגובה המכנית הלא ליניארית של החומרים המרוכבים הפעילים גם באמצעות מיקרוריאולוגיה של פינצטה אופטית (OTM). עם זאת, פרוטוקולים ותוצאות מייצגות עבור ניסויים אלה הם מחוץ לתחום של עבודה זו. הקוראים המעוניינים מופנים לעבודות קודמות48,56 המתארות ביסודיות כיצד לבצע מדידות OTM ואת התוצאות הצפויות.
באמצעות אותה תוכנית של כלי ניסוי וניתוח שתוארה לעיל, החלק הבא מתאר כיצד הדינמיקה והמבנה משתנים כאשר מנועי קינזין ו-biotin-NA crosslinkers משולבים בחומרים המרוכבים (איור 6, איור 7 ואיור 8). איור 6 מראה תמונות קונפוקליות מייצגות של חומרים מרוכבים המונעים על-ידי קינזין בלבד (K) או קינזין ומיוזין (K+M), עם ובלי קישור צולב פסיבי (XL) של חוטי אקטין או מיקרוטובולים.
שילוב קינזין בחומרים מרוכבים גורם בתחילה לדינמיקה דומה ולארגון מחדש של חומרים מרוכבים מונעי מיוזין, כפי שניתן לראות בשורה העליונה של איור 7 (Class 1). עם זאת, הדינמיקה עוברת בדרך כלל לזרימה אניזוטרופית בקנה מידה גדול (איור 7 שורה אמצעית, מחלקה 2), תאוצה והאטה (איור 7 שורה תחתונה, מחלקה 3). מאפיינים אלה משתלבים עם אשכולות וצבירה מזוקליים לאחר 5-30 דקות (איור 6 ואיור 8B). שדות זרימה שנוצרו על-ידי PIV ומפות צבע זמניות המוצגות באיור 7 מתארים דוגמאות של ארגון מחדש איזוטרופי (Class 1, לוח עליון), זרימה מכוונת (Class 2, לוחות אמצעיים) ותאוצה דו-כיוונית (Class 3, לוחות תחתונים).
מהירויות של אקטין ומיקרוטובולים בנקודות זמן משתנות במהלך פעילות, הנקבעות באמצעות התאמות לעקומות τ(q), ממחישות תאוצה ואחריה האטה (איור 8), התלויה בהצלבה. כפי שניתן לראות גם באיור 8, כאשר שני החלבונים המוטוריים משולבים, הדינמיקה למעשה איטית יותר מחומרים מרוכבים קינזין בלבד, ויש התחלה מאוחרת של זרימה מזוקלית. מיוזין תומך גם באינטרפרטציה הומוגנית יותר של רשתות אקטין ומיקרוטובול לאורך כל משך הפעילות, כמו גם פחות צבירה וארגון מחדש. ניתן לראות את ההשפעות האלה בתמונות באיור 6 , והן מכומתות על-ידי אורכי המתאם המשתנים בזמן המחושבים באמצעות SIA, שהם בדרך כלל קטנים יותר בנוכחות מיוזין (איור 8B).
איור 1. תכנון ואפיון של חומרים מרוכבים אקטיביים של אקטין-מיקרוטובול עם מנועים מחוללי כוח מרובים וקרוסלינקרים פסיביים. (A) מונומרים של אקטין ודימרים של טובולין עוברים פולימריזציה משותפת בריכוזים טוחנות c A ו-c T של 0.73-11.6 μM ושברים טוחנות של אקטין Φ A= c A/ (c A+ c T) = 0, 0.25, 0.5, 0.75 ו-1, ליצירת רשתות משותפות של חוטי אקטין (ירוק) ומיקרוטובולים (אדומים). הצלבה פסיבית מושגת באמצעות NA לקישור חוטי אקטין ביוטינילציה (Actin XL) או מיקרוטובולים (MT XL) ביחסים בין-כוכביים: חלבון טוחן של R A = 0.01-0.08 ו- RMT= 0.001-0.01 עבור אקטין ומיקרוטובולים, בהתאמה. צבירי מיני-חוטים של מיוזין-II (סגול) וקינזין (כתום), בריכוזים של c M= 0.12 - 0.48 μM ו- cK = 0.2 - 0.7 μM, דוחפים ומושכים את החוטים כדי להוציא את החומרים המרוכבים ממצב יציב. (B) סכמת מרחב ניסוח. מיני-חוטים של מיוזין II (M), אשכולות קינזין (K) או שני המנועים (K+M) משולבים בחומרים מרוכבים ללא קישורים צולבים פסיביים (No XL), קישורים צולבים של אקטין-אקטין (Actin XL) וקישורים צולבים של מיקרוטובול-מיקרוטובול (MT XL). כל הקריקטורות אינן נמשכות בקנה מידה. אנא לחץ כאן כדי להציג גרסה גדולה יותר של נתון זה.
איור 2. הדמיה קונפוקלית בשני צבעים של שלד ציטוסקלטון מונחה מיוזין עם ריכוזי מיוזין משתנים cM ושברי אקטין טוחנות ΦA. (A) תמונות מיקרוסקופיה קונפוקלית בשני צבעים (A) בגודל 256 x 128 פיקסלים מרובעים (212 x 106 μm2) מראות כיצד חומרים מרוכבים של חוטי אקטין (ירוק) ומיקרוטובולים (אדום) מאורגנים מחדש באמצעות פעילות מוטורית של מיוזין. אין מנועי קינסין או קרוסלינקרים פסיביים. בכל פאנל מוצגות תמונות שצולמו בהתחלה (משמאל, לפני) ובסוף (מימין, אחרי) של הפעלת מיוזין בת 45 דקות (באמצעות תאורה עם אור של 488 ננומטר כדי להשבית את blebbistatin). לוחות מסודרים על ידי הגדלת ריכוז טוחנת של מיוזין (cM), הולך משמאל לימין, והגדלת חלק טוחן של אקטין (ΦA), הולך מלמעלה למטה. הצבעים המתארים כל חלונית תואמים לקידוד הצבעים המשמש באיור 4 ובאיור 5. סרגלי קנה המידה הם 50 μM. כדי ללכוד בצורה הטובה ביותר דינמיקה ומבנה לניתוח, אנו משתמשים בקצבי פריימים של 1-5 ת/ש, ROIs עם צדדים של 50-250 מיקרומטר, ומשכי סדרות זמן של 5-45 דקות, בהתאם לקצב הכיווץ והסידור מחדש. לוחות שבהם התמונות לפני ואחרי נראות דומות מצביעים על ארגון מחדש מינימלי, כפי שניתן לראות בלוחות הוורודים, המג'נטה והציאן. אשכולות בקנה מידה קטן, המעידים על הטרוגניות מוגברת ונוכחות של תכונות נקביות בהירות, ניתן לראות בלוחות הכתומים, הירוקים והאדומים. התכווצות בקנה מידה גדול, הנתפסת כרשת מתכווצת באופן אחיד, ניכרת בלוחות הכחולים והסגולים. נתון זה שונה מהפניה17. אנא לחץ כאן כדי להציג גרסה גדולה יותר של נתון זה.
איור 3. וולוצימטריה של תמונת חלקיקים (PIV) מראה שפעילות אקטומיוזין מפעילה דינמיקה מתכווצת מתואמת של אקטין ומיקרוטובולים במרוכבים מסתבכים. שדות זרימה PIV עבור אקטין (שורה עליונה) ומיקרוטובולים (שורה תחתונה) במרוכב מונחה מיוזין עם (ΦA, cM) = (0.5, 0.24) בזמנים הולכים וגדלים במהלך סדרת זמן של 6 דקות. שדות זרימה נוצרו באמצעות תוסף פיג'י/ImageJ PIV עם זמן השהיה של 20 שניות ושילוב של 2 פיקסלים על 2 פיקסלים. גם אקטין וגם מיקרוטובולים מראים תנועה עקבית המכוונת לאזור המרכזי של שדה הראייה לאורך כל משך הסרט. סרגלי קנה המידה בכל התמונות הם 50 מיקרומטר. צבעי חצים שונים מתאימים למהירויות שונות כפי שמצוין בסולם הצבעים מימין לשדות וקטוריים. נתון זה שונה מהפניה26. אנא לחץ כאן כדי להציג גרסה גדולה יותר של נתון זה.
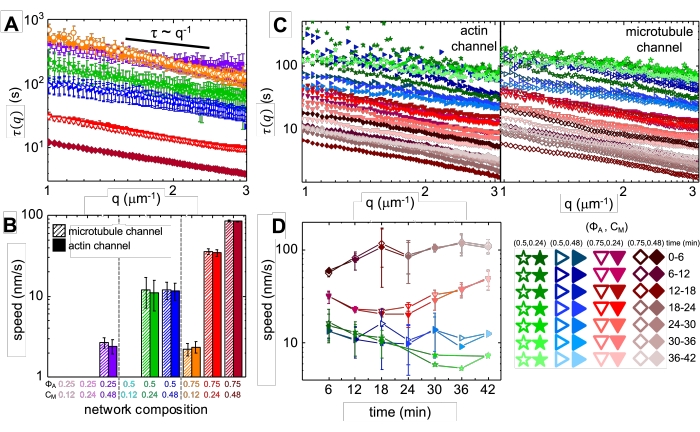
תרשים 4. מיקרוסקופיה דינמית דיפרנציאלית (DDM) הנפתרת בזמן מודדת את קצב וסוג התנועה של אקטין ומיקרוטובולים בחומרים מרוכבים פעילים. (A) DDM מבוצע בערוצי מיקרוטובול (סמלים עליונים ופתוחים) ואקטין (למטה, סמלים מלאים) של סדרות זמן כדי לקבוע זמני דעיכה אופייניים τ לעומת מספר גל q הן עבור אקטין (סמלים מלאים) והן עבור מיקרוטובולים (סמלים פתוחים) כפי שתואר קודם לכן17,26. כל העקומות עוקבות אחר קנה מידה τ ~ q-1, המציין תנועה בליסטית, עם מהירויות v הנקבעות באמצעות התאמות ל- τ(q) = (vq)-1. מהירויות גבוהות יותר מתאימות לערכי τ(q) קטנים יותר עבור כל q נתון. צבעים וצורות של סמלים תואמים לצירופים (Φ A, cM) המוצגים ב- B. (B) מהירויות כיווץ v נקבעות באמצעות התאמות לעקומות τ(q) המוצגות ב- A, אשר ממוצעות על פני כל זמני ההשהיה למשך כל סדרת זמן של 45 דקות. (C) DDM (trDDM) שנפתר בזמן מכמת את האופן שבו הדינמיקה משתנה לאורך זמן על ידי הערכת τ(q) עבור אקטין (סמלים מלאים, משמאל) ומיקרוטובולים (סמלים פתוחים, מימין) עבור מרווחים רצופים של 6 דקות (מסומנים בגוונים שונים של אותו צבע) במהלך זמן ההפעלה של 45 דקות. trDDM מבוצע עבור כל שילוב (ΦA, cM) (מסומן על ידי סמלים וצבעים שונים) כמתואר במקרא בפינה הימנית התחתונה. עקומות τ(q) המוצגות ב-C עוקבות אחר קנה מידה ומגמות דומות לאלה שב-A, אך גם מראות תלות בזמן עבור קומפוזיציות מסוימות (Φ A, cM), בעיקר עבור ΦA= 0.75. (D) מהירויות הכיווץ של חוטי אקטין (סמלים סגורים) ומיקרוטובולים (סמלים פתוחים) נקבעות מהתאמות לעקומות τ(q) מתאימות. קווי שגיאה בכל המגרשים מייצגים את שגיאת התקן של ערכים על פני שלושה עד חמישה שכפולים. נתון זה שונה מהפניה17. אנא לחץ כאן כדי להציג גרסה גדולה יותר של נתון זה.
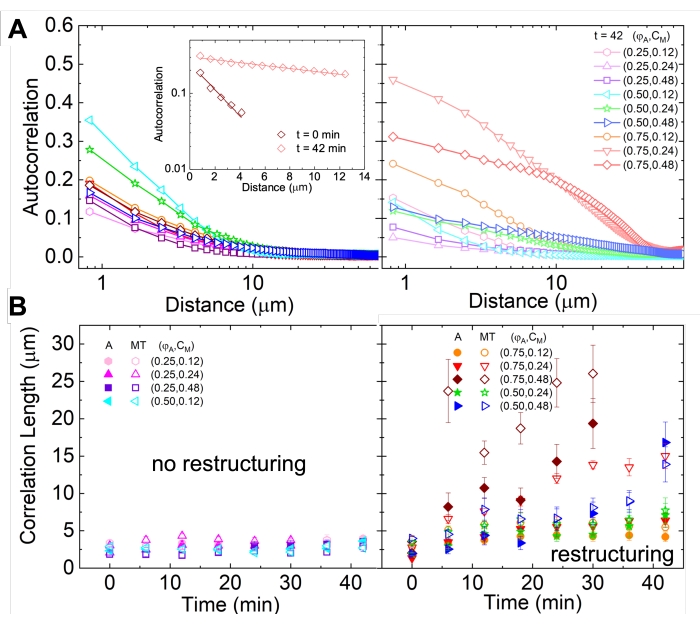
איור 5. ניתוח תיקון אוטומטי של תמונה מרחבית (SIA) מכמת את הארגון מחדש מונע המנוע של מרוכבים ציטוסקטליים פעילים. (A) התאמה אוטומטית g(r) עבור המיקרוטובולים בהתחלה (משמאל, t = 0 דקות, גוונים כהים) ובסוף (מימין, t = 42 דקות, גוונים בהירים) של הניסוי עבור (ΦA, cM) ניסוחים המפורטים במקרא. כניסה: התאמה לדוגמה של נתונים ל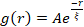 - בזמנים הראשוניים והסופיים עבור (ΦA, cM) = (0.75, 0.12). (B) אורכי מתאם ממוצעים ξ עבור אקטין (סמלים סגורים) ומיקרוטובולים (סמלים פתוחים) עבור כל אחד מהם (Φ A, cM) נקבעים באמצעות התאמות מעריכיות של כל עקומת g(r), כפי שמוצג בכניסה ב- A. הנתונים מחולקים לאלה המציגים ארגון מחדש מינימלי (משמאל) לעומת משמעותי (מימין). קווי שגיאה ב- A וב- B מייצגים את שגיאת התקן על-פני שלושה עד חמישה עותקים משוכפלים. נתון זה שונה מהפניה17. אנא לחץ כאן כדי להציג גרסה גדולה יותר של נתון זה.
- בזמנים הראשוניים והסופיים עבור (ΦA, cM) = (0.75, 0.12). (B) אורכי מתאם ממוצעים ξ עבור אקטין (סמלים סגורים) ומיקרוטובולים (סמלים פתוחים) עבור כל אחד מהם (Φ A, cM) נקבעים באמצעות התאמות מעריכיות של כל עקומת g(r), כפי שמוצג בכניסה ב- A. הנתונים מחולקים לאלה המציגים ארגון מחדש מינימלי (משמאל) לעומת משמעותי (מימין). קווי שגיאה ב- A וב- B מייצגים את שגיאת התקן על-פני שלושה עד חמישה עותקים משוכפלים. נתון זה שונה מהפניה17. אנא לחץ כאן כדי להציג גרסה גדולה יותר של נתון זה.
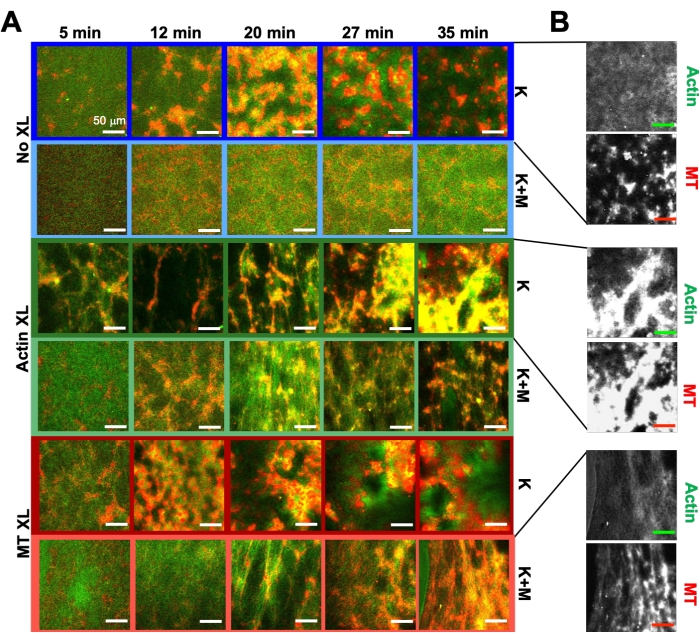
תרשים 6. שילוב מנועי קינזין וקרוסלינקרים פסיביים בחומרים מרוכבים אקטיביים כדי להגביר את יכולת התכנות ולהרחיב את מרחב הפאזה של הדינמיקה והמבנה. (A) תמונות קונפוקליות בשני צבעים של אקטין (ירוק) ומיקרוטובולים (אדום) בחומרים מרוכבים פעילים מראות ארגון מחדש תלוי ניסוח מורכב לאורך זמן (רשום במינימום). חמש התמונות בכל שורה מתאימות לחמש מסגרות של סדרת זמן של 2000 פריימים שנרכשו עבור קומפוזיט המונע על ידי קינזין (K, שורות 1, 3, 5) או קינזין ומיוזין (K+M, שורות 2, 4, 6), וכולל קישורים צולבים פסיביים (No XL, שורות 1, 2), קישורים צולבים של אקטין-אקטין (Actin XL, שורות 3, 4), או קישורים צולבים של מיקרוטובול-מיקרוטובול (MT XL, שורות 5, 6). סרגלי קנה המידה הם כולם 50 מיקרומטר. צבעי המיתאר תואמים לערכת הצבעים באיור 8. (B) תעלות פלואורסצנטיות נפרדות של אקטין ומיקרוטובול עבור מרוכבים של קינזין בלבד מראות מבנים מגוונים עם לוקליזציה משותפת של אקטין-MT והפרדת מיקרו-פאזות. התמונות המוצגות הן עבור חומרים מרוכבים עם c A = 2.32 μM, c T = 3.48 μM, c K = 0.35 μM, cM = 0.47 μM (שורות 2, 4, 6), R A = 0.02 (שורות 3, 4) ו- RMT = 0.005 (שורות 5, 6). כל החומרים המרוכבים מתחילים ברשתות אינטרפרטציה מפוזרות באופן אחיד של אקטין ומיקרוטובולים (עמודה 1). מרוכבים מונעי קינסין ללא crosslinkers (שורה 1) יוצרים אשכולות אמורפיים המחוברים באופן רופף שהם עשירים ב- MT. אקטין מתמקם במרכזים של אגרגטים אלה בתחילה, אך לאחר מכן נדחק אל מחוץ לאזורים העשירים ב-MT שממשיכים להתכווץ ולהתנתק זה מזה. קישור צולב של אקטין-אקטין (שורה 3) מעכב הפרדה זו בין אקטין-MT בקנה מידה זעיר, ובמקום זאת אגרגטים עשירים ב-MT מחוברים באמצעות גדילים ארוכים של אקטין. ההצלבה של אקטין מאפשרת גם קליטה איטית של אקטין לאזורים העשירים ב-MT, כך שהתרכובת הופכת לרשת מחוברת של אשכולות אקטין ו-MT מקומיים. קישור צולב מיקרוטובולי (שורה 5) מוביל לאשכולות אמורפיים של MTs המתלכדים עם הזמן, וכתוצאה מכך הפרדת פאזה בקנה מידה גדול יותר של אקטין ו- MTs. הוספת מיוזין (שורות 2, 4, 6) מפחיתה את הדה-ערבוב והארגון מחדש המונעים על ידי קינזין. ללא קישורים צולבים (שורה 2), מרוכבים מראים סידור מחדש מועט במהלך השעות. Crosslinking מגביר את הארגון מחדש ואת לוקליזציה משותפת של אקטין ומיקרוטובולים (שורות 4, 6). באופן ספציפי, כאשר מיקרוטובולים מקושרים (שורה 6), יש אינטרפרטציה משמעותית וארגון מחדש לרשתות דמויות רשת של סיבים. נתון זה שונה מהפניה50. אנא לחץ כאן כדי להציג גרסה גדולה יותר של נתון זה.
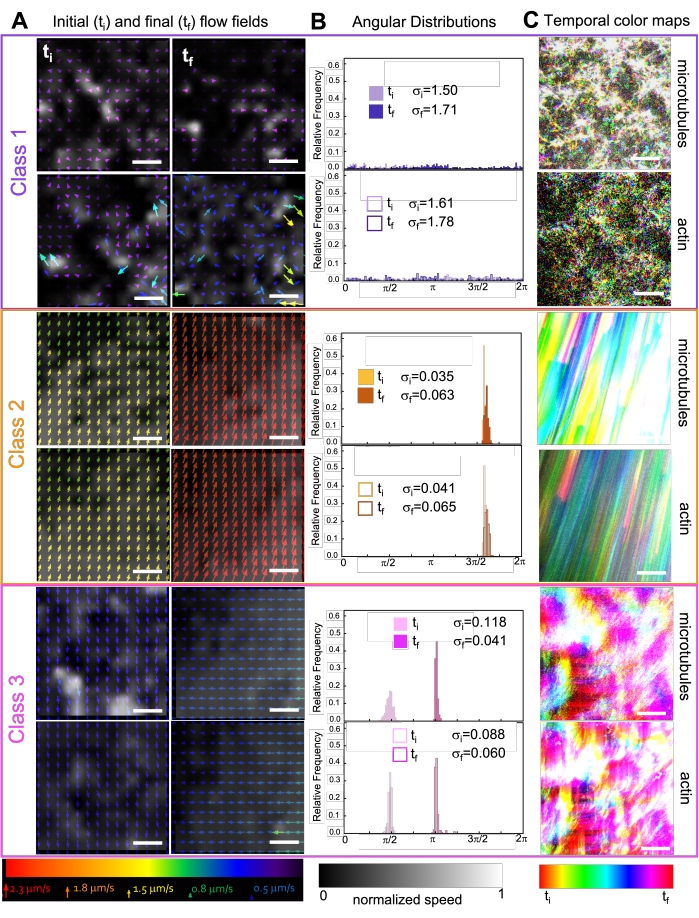
תרשים 7. PIV מראה כי מרוכבים פעילים מציגים שלושה סוגים של שדות זרימה מובחנים מרחבית. (A) שדות זרימה PIV עבור המסגרות הראשונות (t i) והאחרונות (tf) של שלוש סדרות זמן מייצגות, המציגות את המחלקות הדינמיות השונות שחומרים מרוכבים המוצגים באיור 6 מציגים. שדות זרימה PIV עבור מיקרוטובולים (למעלה) ואקטין (למטה) עבור מחלקה 1 (למעלה, סגול), מחלקה 2 (אמצע, כתום) ומחלקה 3 (למטה, מגנטה) לדוגמה, עם צבעי החצים המתאימים לסולם המהירות האוניברסלי בתחתית, ומפת הצבעים בגווני אפור המציגה את התפלגות המהירות המרחבית, מנורמלת בנפרד עבור כל שדה זרימה בהתאם לקנה המידה המוצג בתחתית. סרגלי קנה המידה הם כולם 50 μM. (B) התפלגויות זוויתיות של וקטורי מהירות מ-A (ביחידות של רדיאנים) עם סטיות תקן ראשוניות וסופיות מפורטות σi ו-σ-f. (C) מפות צבע זמניות של סרטוני הווידאו שנותחו ב-A וב-B מציגות את המיקום של פריים למסגרת של כל פיקסל ביחס לנקודת ההתחלה שלו. מפות מחלקה 1 מציגות תנועה אקראית בקנה מידה קטן; מפות מחלקה 2 מתארות תנועה חד-כיוונית מהירה עם שונות מרחבית או זמנית מינימלית; מפות מחלקה 3 מציגות תכונות של מחלקה 1 ו-2. נתון זה שונה מהפניה50. אנא לחץ כאן כדי להציג גרסה גדולה יותר של נתון זה.
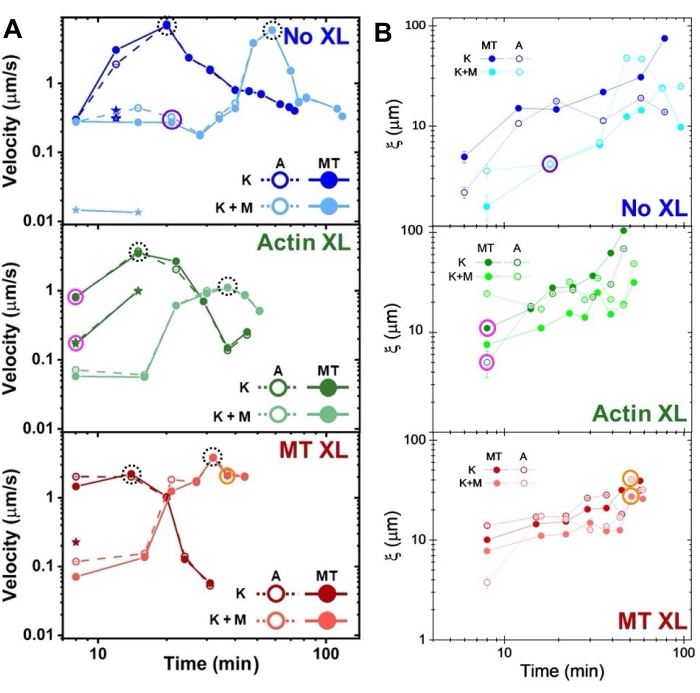
תרשים 8. DDM ו-SIA מודדים את הדינמיקה והמבנה המשתנים בזמן של מרוכבים דו-מנועיים מסוג אקטין-מיקרוטובול. (A) מהירויות עבור חומרים מרוכבים המתוארות באיור 6 ובאיור 7, הנמדדות באמצעות DDM, מראות תאוצה והאטה של חומרים מרוכבים, המתוכנתים על-ידי פעילות קרוסלינקינג ומיוזין. מהירויות של מיקרוטובולים (MT, מעגלים סגורים) ואקטין (A, עיגולים פתוחים) משורטטות כפונקציה של זמן פעילות במרוכבים ללא crosslinking (למעלה, כחול), actin crosslinking (אמצע, ירוק), microtubule crosslinking (למטה, אדום), ללא מיוזין (K, גוונים כהים יותר), ועם מיוזין (K+M, גוונים בהירים יותר). במקרים מסוג 3, בעלי שתי מהירויות, המהירות האיטית יותר מסומנת על ידי כוכב. נקודות נתונים המוקפות בעיגולים שחורים מקווקווים מתאימות למהירות מרבית vמקסימוםעבור כל ניסוח. קווי שגיאה (הקטנים מכדי לראותם) הם שגיאת התקן מעל ההתאמות של חוק הכוח של τ(q) המתאים. (B) אורכי מתאם מבני ξ, שנקבע באמצעות SIA, לעומת זמן פעילות, עבור אותה קבוצה של סדרות זמן המוערכות ב-A. כל נקודת נתונים היא ממוצע של אורכי המתאם שנקבעו עבור המסגרת הראשונה והאחרונה של סדרת הזמן המתאימה. באופן כללי, ξ עולה בזמן הן עבור אקטין והן עבור מיקרוטובולים בכל מערכות החומרים המרוכבים, ולחומרים מרוכבים המונעים אך ורק על ידי קינזין יש אורכי מתאם גדולים יותר מאלה שבהם מיוזין קיים גם כן. נקודות נתונים ב-A וב-B המתאימות לשלוש סדרות הזמן שנותחו באיור 7 מוקפות בעיגול בצבע המחלקה המתאים (1 = סגול, 2 = כתום, 3 = מגנטה). נתון זה שונה מהפניה50. אנא לחץ כאן כדי להציג גרסה גדולה יותר של נתון זה.
התקדמות מרכזית של המערכת המשוחזרת שתוארה לעיל היא המודולריות והכוונון שלה, ולכן אנו מעודדים את המשתמשים לשנות את ריכוזי החלבונים, המנועים, הקישורים הצולבים וכו 'כך שיתאימו לתוצאות הרצויות להם, בין אם זה לחקות תהליך תאי מסוים או להנדס חומר בעל פונקציונליות או תכונות מכניות ספציפיות. מגבלות על טווח הריכוזים של אקטין וטובולין נקבעות בגבול התחתון על ידי הריכוז הקריטי הדרוש לפולימריזציה של אקטין (~0.2 μM)57,58,59 וטובולין (~3 - 4 μM)60, ובגבול העליון על ידי המעבר ליישור נמטי של חוטי אקטין (~90 μM)61,62 או מיקרוטובולים (~35 μM)63 . מונומרים של אקטין ודימרים של טובולין צריכים להיות מפולמרים לחוטים יחד, ולא מעורבבים יחד לאחר פילמור, כדי להבטיח שהם יוצרים רשתות מחלחלות הומוגניות התומכות זו בזו באופן סינרגטי. הדינמיקה החדשנית שמציגים החומרים המרוכבים מסתמכת על אינטראקציה זו. בעוד שבדרך כלל חשוב לבצע את כל השלבים כמתואר בפרוטוקול כדי לשחזר בהצלחה את התוצאות המוצגות, שלבים מסוימים מדויקים יותר, בעוד שלאחרים יש מקום לשנות ולהתאים כדי להתאים לצרכים ספציפיים ולמשאבים זמינים.
לדוגמה, שלב חשוב אחד בהבטחת תוצאות הניתנות לשחזור הוא הכנה ואחסון נכונים של הריאגנטים בהתאם להנחיות המופיעות בטבלת החומרים. חלבונים ציטוסקטליים (אקטין, טובולין, מיוזין, קינזין) הם לביליים ויש לצטט אותם, להקפיא הבזק עם חנקן נוזלי, ולאחסן אותם בטמפרטורה של -80 מעלות צלזיוס באליקוטים חד-פעמיים. לאחר הסרת מ -80 מעלות צלזיוס, aliquots צריך להישמר על קרח. חלבונים ציטוסקטליים אינם שומרים באופן אמין על תפקודם לאחר מחזורי הפשרה נוספים בהקפאה.
מיקרוטובולים רגישים יותר לדה-פולימריזציה ולדנטורציה מאשר אקטין. לאחר הסרת מ -80 מעלות צלזיוס, טובולין צריך להישמר על קרח לפני פילמור, ולהשתמש בתוך 12 שעות. לאחר פולימריזציה, מיקרוטובולים צריכים להישמר בטמפרטורת החדר. כמו כן, חיוני לייצב מיקרוטובולים באמצעות טקסול כדי למנוע דה-פולימריזציה. ייצוב פאלוידין של חוטי אקטין חשוב גם הוא כדי לדכא את הליכון האקטין הצורכת ATP המתחרה בפעילות מיוזין וקינסין.
Ultracentrifugation של מנועי מיוזין הוא צעד קריטי נוסף, כפי שהוא מסיר ראשים מתים מיוזין לא פעיל. אי הסרת המונומרים הלא פעילים מבחינה אנזימטית גורמת להצלבה פסיבית של רשת האקטין ולאובדן פעילות. כדי להאריך את פעילות ה-ATPase של מנועים, ניתן לשלב מערכת התחדשות ATP כגון קריאטין פוספט וקריאטין פוספוקינאז64 .
לבסוף, שמירה על פעילות מרוכבת דורשת עיכוב ספיחה של חוטים ומנועים לקירות תא הדגימה, אשר ניתן להשיג על ידי פסיבציה של כיסויי המיקרוסקופ והשקופיות. חלבונים מוטוריים מועדים במיוחד לספיחה, וכתוצאה מכך הקומפוזיט נמשך אל פני השטח של תא הדגימה, נע מחוץ לשדה הראייה, קורס לדו-ממד, ואינו עובר עוד פעילות. סילוניזציה של הכיסויים והמגלשות היא דרך יעילה להעביר את המשטחים ולמנוע ספיחה (ראו שלב 1). שיטת פסיבציה חלופית המשמשת ביעילות בניסויי שלד במבחנה היא ציפוי פני השטח בדו-שכבת שומנים, בדומה לקרום התא18. שיטה זו היא יתרון אם רוצים לקשור חלבונים לפני השטח או להציג אינטראקציות ספציפיות אחרות בין חלבון למשטח, מכיוון שניתן לתפקד את הדו-שכבתי. עבור ניסויי פינצטה אופטיים, פסיבציה של המיקרוספרות היא גם קריטית, וניתן להשיג אותה על ידי ציפוי מיקרוספרות קרבוקסיליות עם BSA או PEG באמצעות כימיה של קרבודימיד קרוסלינקר48.
ישנם כמה היבטים של הפרוטוקולים המוצגים שחוקרים עשויים לשקול לשנות בהתאם לצרכים שלהם. ראשית, חוקרים עשויים לבחור להחליף קרוסלינקרים של ביוטין-NA שאינם מקומיים בהצלבות ביולוגיות, כגון אלפא-אקטינין או MAP65 שמצליבים אקטין ומיקרוטובולים,בהתאמה 28,65,66. השימוש בהצלבות שאינן ילידיות במרוכבים המתוארים כאן מונע על ידי יכולת השחזור, היציבות והכוונון המשופרות שלהם בהשוואה להצלבות מקומיות. בגלל קשר הביוטין-NA החזק, ניתן להניח ש-crosslinkers הם קבועים, ולא רוב ה-crosslinkers המקומיים שנקשרים באופן ארעי עם שיעורי תחלופה רחבים. הדינמיקה של crosslinking חולף מסבכת את ניתוח התרומות של crosslinkers ומנועים לדינמיקה. יתר על כן, ניתן להשתמש באופן רב-תכליתי במקשרי ביוטין-NA כדי להצליב גם אקטין וגם מיקרוטובולים, כמו גם להצליב אקטין למיקרוטובולים. בדרך זו, ניתן לבצע השוואה חד משמעית בין מוטיבים של crosslinking, תוך שמירה על כל המשתנים האחרים (למשל, גודל crosslinker, זיקה מחייבת, stoichiometry, וכו ') קבוע. לבסוף, הריאגנטים הדרושים לשילוב מקשרי ביוטין-NA זמינים באופן מסחרי, מאופיינים היטב ונפוצים במעבדות ביופיזיקה רבות. עם זאת, אחת החוזקות המרכזיות של פלטפורמת המבחנה המתוארת כאן היא המודולריות שלה, כך שחוקרים צריכים להיות מסוגלים להחליף בצורה חלקה מקשרי ביוטין-NA במקשרים מקומיים אם יבחרו.
שנית, בפרוטוקול הנוכחי, מונומרים של אקטין ודימרים של טובולין מתפלמרים לחוטים יחד בצינור צנטריפוגה לפני שהם מתווספים לתא הדגימה. הזרמת התמיסה של חלבוני נימה סבוכים לתוך תא הדגימה עלולה לגרום ליישור זרימה, במיוחד של המיקרוטובולים, מה ששובר את האיזוטרופיה וההומוגניות הרצויות של החומרים המרוכבים. ואכן, התקדמות משמעותית בעבודה קודמת על מרוכבים מסוג אקטין-מיקרוטובול במצב יציב הייתה היכולת לבצע פולימריזציה משותפת של אקטין ומיקרוטובולים באתרם (בתא הדגימה) כדי להבטיח היווצרות של רשתות איזוטרופיות של אקטין ומיקרוטובולים15,16,27. עם זאת, הרחבת גישה זו לחומרים מרוכבים פעילים תדרוש הוספת המנועים לדגימה לפני פילמור אקטין וטובולין ודגירה של כל הדגימה יחד בטמפרטורה של 37 מעלות צלזיוס לפני הניסויים. בדיקות של וריאציה זו לפרוטוקול הביאו לפולימריזציה מופחתת של אקטין וללא פעילות מוטורית ניכרת, ככל הנראה בשל פעילות ATPase מתחרה והדגירה הממושכת של 37 מעלות צלזיוס של המנועים. למרבה המזל, לא ניתן להבחין ביישור זרימה של חומרים מרוכבים כאשר עוקבים אחר הפרוטוקולים הנוכחיים, כפי שניתן לראות באיור 2, איור 3 ואיור 6. עם זאת, החוקרים מוזמנים לתכנן פרוטוקולים המאפשרים היווצרות באתרם של חומרים מרוכבים פעילים.
נקודת שיקול נוספת היא ערכת הסימון הפלואורסצנטית, הכוללת תיוג דליל של כל חוטי האקטין והמיקרוטובולים ברשת. גישת תיוג זו עברה אופטימיזציה כדי להמחיש באופן ישיר את מבנה הרשת במקום להסיק מבנה ודינמיקה באמצעות חוטי מעקב או מיקרוספרות. עם זאת, הפשרה היא כי חוטים בודדים אינם מסומנים בבהירות וניתן לפתרון. גישה אחת שחוקרים יכולים לנקוט כדי לפתור גם חוטים בודדים וגם לדמיין את מבנה הרשת היא להשתמש בחוטים שנוצרו מראש המסומנים בפלואורופור אחר, כך שגם הרשת הסובבת וגם חוטים בודדים יכולים להיות מצולמים בו זמנית. עם זאת, כאשר משתמשים ביותר משני פלואורופורים וערוצי עירור/פליטה, לעתים קרובות קשה לבטל את הדימום בין הערוצים, ולכן יש להקפיד על בחירת הפלואורופורים, המסננים ועוצמות הלייזר.
מגבלה קשורה היא חוסר היכולת לדמיין את מנועי המיוזין או הקינסין בחומרים המרוכבים. מונומרים של אקטין עם תווית פלואורסצנטית ודימרים של טובולין המשמשים זמינים באופן מסחרי, בעוד שהדמיה של מיוזין או קינזין בחומרים מרוכבים דורשת תיוג פנימי. החוקרים מוזמנים לעשות את הצעד הבא כדי לתייג מנועים, כפי שנעשה בעבר18,67, כדי להיות מסוגלים לקשר באופן חד משמעי את הפעילות המוטורית ואת הקישור לדינמיקה ולמבנים שהחומרים המרוכבים שלנו מציגים.
לבסוף, חשוב לציין כי, בפרוטוקול הנוכחי, את ההתחלה ואת משך הפעילות kinesin אינו נשלט. מאחר שפעילות המיוזין נשלטת באמצעות השבתה פוטו-נטרוטית של בלביסטטין, כפי שתואר לעיל, כדי לבנות הפעלת אור דומה של קינזין, ניתן לשלב ATP המופעל על-ידי אור.
כדי לבנות את המורכבות של העיצובים המתוארים כאן, כדי לחקות טוב יותר את התנאים התאיים ולהרחיב את מרחב הפרמטרים של מבנה-פונקציה דינמית, העבודה העתידית תתמקד בשילוב חוטי ביניים, כגון vimentin68,69, כמו גם מנועים אחרים כגון dynein13,70. גלסולין ישולב גם בריכוזים שונים כדי לשלוט באורך אקטין14, כמו גם חלבון טאו כדי לשלוט בנוקשות המיקרוטובול.
לסיכום, הפרוטוקולים המוצגים מתארים כיצד לתכנן, ליצור ולאפיין את הדינמיקה, המבנה והמכניקה של מערכות חומר פעיל בהשראת ציטוסקלטון, המכילות שני רכיבים פעילים נפרדים המייצרים כוח הפועל על מצעים שונים במערכת אחת. פלטפורמה מתכווננת ומודולרית זו מקרבת את מאמצי השיקום צעד חשוב אחד קרוב יותר לחיקוי השלד התאי ומציעה את היכולת הייחודית לתכנת את תכונותיו על פני מרחב פאזה רחב על ידי שילוב עצמאי, הסרה וכוונון של הרכיבים השונים. יתר על כן, כל הרכיבים של מערכת רב-תכליתית זו זמינים באופן מסחרי (ראו טבלת חומרים), למעט דימרים קינזין אשר מטוהרים במעבדת רוס, כפי שתואר קודםלכן 50, וזמינים על פי בקשה. לבסוף, כל קוד הניתוח זמין באופן חופשי דרך GitHub49 ומבוסס על שפות תכנות ותוכנות חופשיות (Python ו- Fiji). ההפצה השקופה של פרוטוקולים לתכנון מערכות אלה תהפוך את הפלטפורמה הזו לנגישה יותר לקבוצה מגוונת של משתמשים בעלי מומחיות, רקע, שיוך מוסדי ומטרות מחקר שונות.
למחברים אין מה לחשוף
אנו מודים למאיה הנדייה ולד"ר ג'ונתן מישל על סיוע בניתוח נתונים, ולד"ר ג'נט שונג, ד"ר מומיטה דאס וד"ר מייקל רוסט על דיונים והדרכה מועילים. מחקר זה נתמך על ידי מענק מחקר של קרן ויליאם מ. קק ופרס NSF DMREF (DMR 2119663) המוענקים למענקי RMRA ו-JLR והמכונים הלאומיים לבריאות R15 (R15GM123420, 2R15GM123420-02) שהוענקו ל-RMR-A ול-RJM.
| Name | Company | Catalog Number | Comments |
| (-)-Blebbistatin Abbreviation used in paper: blebbistatin | Sigma Aldrich | B0560 | Stock Concentration: 200 μM in DMSO Storage: dessicated, in DMSO, -20ºC Stock and Experiment Recipes: dissolve 1 mg of powder to 200 μM in DMSO Storage, Handling, Troubleshooting Notes: limited shelf-life, typically stops functioning reliably after 3-4 months. purchase and prepare new solution every 3 months. |
| 1:20 488-tubulin:tubulin mixture Abbreviation used in paper: 5-488-tubulin | NA | NA | Stock Concentration: 5 mg/ml in PEM Storage: single use aliquots, -80ºC, avoid freeze-thaw cycles Stock and Experiment Recipes: mix tubulin and 488-tubulin at a 20:1 ratio, flash freeze with LN2 Storage, Handling, Troubleshooting Notes: each aliquot can be used for up to 12 hrs stored on ice at 4ºC, protect from light |
| 1:20 R-tubulin:tubulin mixture Abbreviation used in paper: 5-R-tubulin | NA | NA | Stock Concentration: 5 mg/ml in PEM Storage: single use aliquots, -80ºC, avoid freeze-thaw cycles Stock and Experiment Recipes: mix tubulin and rhodamine tubulin at a 20:1 ratio, flash freeze with LN2 Storage, Handling, Troubleshooting Notes: each aliquot can be used for up to 12 hrs stored on ice at 4ºC, protect from light |
| actin (biotin): skeletal muscle Abbreviation used in paper: biotin-actin | Cytoskeleton | AB07 | Stock Concentration: 1 mg/ml in G-buffer Storage: single use aliquots, -80ºC, avoid freeze-thaw cycles Stock and Experiment Recipes: reconstitute to 1 mg/ml in G-buffer, flash freeze with LN2 Storage, Handling, Troubleshooting Notes: (1) immediately prior to use dilute to 0.5 mg/ml in PEM, (2) once removed from -80ºC, store aliquot on ice at 4ºC for up to 1 week |
| actin (rhodamine): rabbit skeletal muscle Abbreviation used in paper: R-actin | Cytoskeleton | AR05 | Stock Concentration: 1.5 mg/ml in G-buffer Storage: single use aliquots, -80ºC, avoid freeze-thaw cycles Stock and Experiment Recipes: reconstitute to 1.5 mg/ml in G-buffer, flash freeze with LN2 Storage, Handling, Troubleshooting Notes: once removed from -80ºC, store aliquot on ice at 4ºC, can be used for up to 1 week |
| adenosine triphosphate Abbreviation used in paper: ATP | Thermo Fisher Scientific | A1048 | Stock Concentration: 100 mM Storage: in solution (pH 7), -20ºC Stock and Experiment Recipes: reconsitute in DI H20, bring pH to 7 with NaOH Storage, Handling, Troubleshooting Notes: routinely check pH and adjust as needed, hydrolyzes over time, replace every ~6-12 months |
| AlexaFluor488 Phalloidin Abbreviation used in paper: 488-phalloidin | Thermo Fisher Scientific | A12379 | Stock Concentration: 100 μM DMSO Storage: protected from light, dessicated, -20ºC Stock and Experiment Recipes: reconstitute to 100 μM with DMSO Storage, Handling, Troubleshooting Notes: immediately prior to use dilute to 20 μM in PEM (1 μL in 4 μL PEM) |
| AlexaFluor488–labeled actin Abbreviation used in paper: 488-actin | Thermo Fisher Scientific | A12373 | Stock Concentration: 1.5 mg/ml in G-buffer Storage: single use aliquots, -80ºC, avoid freeze-thaw cycles Stock and Experiment Recipes: reconstitute to 1.5 mg/ml in G-buffer, flash freeze with LN2 Storage, Handling, Troubleshooting Notes: this item has been discontinued |
| Basic Plasma Cleaner Abbreviation used in paper: plasma cleaner | Harrick Plasma | PDC-32G | |
| Bemis Parafilm M Laboratory Wrapping Film Abbreviation used in paper: transparent film | Thermo Fisher Scientific | 13-374-5 | |
| D-(+)-Glucose Abbreviation used in paper: | Thermo Fisher Scientific | A1682836 | Stock Concentration: 100x Storage: store at stock concentration (100x) or 10x concentration, dessicated, at -20ºC Stock and Experiment Recipes: reconstitute powder to 4.5 mg/ml in DI H20 Storage, Handling, Troubleshooting Notes: final concentration in solution should 45 μg/mL |
| D-Biotin Abbreviation used in paper: biotin | Fisher Scientific | BP232-1 | Stock Concentration: 1.02 mM in PEM Storage: dessicated, 4ºC |
| deionized nanopure water Abbreviation used in paper: DI | |||
| Dimethyldichlorosilane Abbreviation used in paper: silane | Thermo Fisher Scientific | D/3820/PB05 | Stock Concentration: 2% dissolved in Toulene |
| Dithiothreitol Abbreviation used in paper: DTT | Thermo Fisher Scientific | R0861 | Stock Concentration: 1 M in DMSO Storage: dessicated, -20ºC Stock and Experiment Recipes: dilute to 2 mM in PEM immediately before each experiment |
| DMSO Anhydrous Abbreviation used in paper: DMSO | Thermo Fisher Scientific | D12345 | |
| F-Buffer Abbreviation used in paper: F-buffer | NA | NA | Stock Concentration: 10x Storage: dessicated, -20ºC Stock and Experiment Recipes: 10 mM Imidazole (pH 7.0), 50 mM KCl, 1 mM MgCl2, 1 mM EGTA, 0.2 mM ATP |
| G-Buffer Abbreviation used in paper: G-buffer | NA | NA | Stock Concentration: 10x Storage: dessicated, -20ºC Stock and Experiment Recipes: 2.0 mM Tris (pH 8), 0.2 mM ATP, 0.5 mM DTT, 0.1 mM CaCl2. Store at -20°C. |
| glass microscope slide Abbreviation used in paper: slide | Thermo Fisher Scientific | 22-310397 | |
| Glucose oxidase + catalase + β-mercaptoethanol Abbreviation used in paper: GOC | Sigma Aldrich | G2133-250KU, C1345, 63689 | Stock Concentration: 100x Storage: store at stock concentration (100x) or 10x concentration, dessicated, at -20ºC Stock and Experiment Recipes: For 100x: 4.3 mg/ml glucose oxidase, 0.7 mg/ml catalase, 0.5% v/v β-mercaptoethanol in DI H20 Storage, Handling, Troubleshooting Notes: final concentration in solution should be: 0.005% β-mercaptoethanol, 43 μg/mL glucose oxidase, 7 μg/mL catalase |
| glu-GOC oxygen scavenging system Abbreviation used in paper: glu-GOC | NA | NA | Stock Concentration: 100x Storage: prepare fresh each time Stock and Experiment Recipes: mix equal parts Glu and GOC and add at 1/100 final sample volume immediately before imaging Storage, Handling, Troubleshooting Notes: prepare from Glu and GOC immediately before imaging |
| Guanosine triphosphate Abbreviation used in paper: GTP | Thermo Fisher Scientific | R0461 | Stock Concentration: 100 mM Storage: 100 μL aliquots at -20ºC |
| Instant Mix 1-minute epoxy Abbreviation used in paper: epoxy | Loctite | 1366072 | |
| Kinesin-1 401 BIO 6x HIS Abbreviation used in paper: kinesin | Prepared in JL Ross Lab at Syracuse University | NA | Stock Concentration: 8.87 μM in PEM Storage: single use aliquots, -80ºC, avoid freeze-thaw cycles Storage, Handling, Troubleshooting Notes: biotinylated dimers form kinesin clusters, each aliquot can be used for up to 12 hrs stored on ice at 4ºC |
| NeutrAvidin Abbreviation used in paper: NA | Thermo Fisher Scientific | 31000 | Stock Concentration: 5 mg/ml in PEM Storage: dessicated, -20ºC Stock and Experiment Recipes: reconstitute powder to 5 mg/ml in PEM |
| No 1. glass coverslips (24 mm x 24 mm) Abbreviation used in paper: coverslip | Thermo Fisher Scientific | 12-548-CP | |
| Paclitaxel Abbreviation used in paper: Taxol | Thermo Fisher Scientific | P3456 | Stock Concentration: 2 mM in DMSO Storage: protected from light, dessicated, -20ºC Stock and Experiment Recipes: reconstitute to 2 mM with DMSO Storage, Handling, Troubleshooting Notes: immediately prior to use dilute to 200 μM in DMSO (0.4 μL in 3.6 μL DMSO) |
| PEM-100 Abbreviation used in paper: PEM | NA | NA | Stock Concentration: 1x Storage: room temperature (RT) Stock and Experiment Recipes: 100 mM K-PIPES (pH 6.8), 2 mM EGTA, 2 mM MgCl2 Storage, Handling, Troubleshooting Notes: use KOH to adjust pH to 6.8, recheck pH often and adjust accordingly |
| phalloidin Abbreviation used in paper: phalloidin | Thermo Fisher Scientific | P3457 | Stock Concentration: 100 μM in DMSO Storage: protected from light, dessicated, -20ºC, adhere closely to storage/handling conditions Stock and Experiment Recipes: reconstitute to 100 μM with DMSO Storage, Handling, Troubleshooting Notes: susceptible to impurities in its preparation and denaturing, identifiable as large amorphous aggregates of actin in samples |
| porcine brain tubulin Abbreviation used in paper: tubulin | Cytoskeleton | T240 | Stock Concentration: 5 mg/ml in PEM Storage: single use aliquots, -80ºC, avoid freeze-thaw cycles Stock and Experiment Recipes: reconstitute powder to 5 mg/ml in PEM, flash freeze with LN2 Storage, Handling, Troubleshooting Notes: each aliquot can be used for up to 12 hrs stored on ice at 4ºC |
| Potassium Chloride Abbreviation used in paper: KCl | Thermo Fisher Scientific | AM9640G | Stock Concentration: 4 M Storage: RT |
| Rabbit skeletal actin Abbreviation used in paper: actin | Cytoskeleton | AKL99 | Stock Concentration: 2 mg/ml in G-buffer Storage: single use aliquots, -80ºC, avoid freeze-thaw cycles Stock and Experiment Recipes: reconstitute to 2 mg/ml in G-buffer, flash freeze with LN2 Storage, Handling, Troubleshooting Notes: once removed from -80ºC, store aliquot on ice at 4ºC, can be used for up to 1 week |
| Rabbit skeletal myosin II Abbreviation used in paper: myosin | Cytoskeleton | MY02 | Stock Concentration: 10 mg/ml in PEM Storage: single use aliquots, -80ºC, avoid freeze-thaw cycles Stock and Experiment Recipes: reconstitute powder to 10 mg/ml in PEM, flash freeze with LN2 Storage, Handling, Troubleshooting Notes: monomers form minifilaments at low KCl, each aliquot can be used for up to 12 hrs stored on ice at 4ºC |
| Tubulin (biotin): porcine brain Abbreviation used in paper: biotin-tubulin | Cytoskeleton | T333P | Stock Concentration: 5 mg/ml in PEM Storage: single use aliquots, -80ºC, avoid freeze-thaw cycles Stock and Experiment Recipes: reconstitute powder to 5 mg/ml in PEM, flash freeze with LN2 Storage, Handling, Troubleshooting Notes: immediately prior to use dilute to 0.5 mg/ml in PEM |
| Tubulin (fluorescent HiLyte 488): porcine brain Abbreviation used in paper: 488-tubulin | Cytoskeleton | TL488M | Stock Concentration: 5 mg/ml in PEM Storage: single use aliquots, -80ºC, avoid freeze-thaw cycles Stock and Experiment Recipes: reconstitute powder to 5 mg/ml in PEM, flash freeze with LN2 Storage, Handling, Troubleshooting Notes: each aliquot can be used for up to 12 hrs stored on ice at 4ºC, protect from light |
| tubulin (rhodamine): porcine brain Abbreviation used in paper: R-tubulin | Cytoskeleton | TL590M | Stock Concentration: 5 mg/ml in PEM Storage: single use aliquots, -80ºC, avoid freeze-thaw cycles Stock and Experiment Recipes: reconstitute powder to 5 mg/ml in PEM, flash freeze with LN2 Storage, Handling, Troubleshooting Notes: each aliquot can be used for up to 12 hrs stored on ice at 4ºC, protect from light |
| Tween 20 Abbreviation used in paper: Tween20 | Thermo Fisher Scientific | J20605.AP | Stock Concentration: 1% v/v in DI H20 Storage: RT |
| ultracentrifuge grade microtubes Abbreviation used in paper: Beckman-Coulter Optima Max XP | Beckman Coultier | 343776 | Storage, Handling, Troubleshooting Notes: 8x34 mm PC |
| UV light curing glue Abbreviation used in paper: UV glue | Pharda | SKG-2869 |
- Fletcher, D. A., Mullins, R. D. Cell mechanics and the cytoskeleton. Nature. 463 (7280), 485-492 (2010).
- Koenderink, G. H., Paluch, E. K. Architecture shapes contractility in actomyosin networks. Current Opinion in Cell Biology. 50, 79-85 (2018).
- Dogterom, M., Koenderink, G. H. Actin-microtubule crosstalk in cell biology. Nature Reviews Molecular Cell Biology. 20 (1), 38-54 (2019).
- Burla, F., Mulla, Y., Vos, B. E., Aufderhorst-Roberts, A., Koenderink, G. H. From mechanical resilience to active material properties in biopolymer networks. Nature Reviews Physics. 1 (4), 249-263 (2019).
- Wen, Q., Janmey, P. A. Polymer physics of the cytoskeleton. Current Opinion in Solid State and Materials Science. 15 (5), 177-182 (2011).
- Xiao, Q., Hu, X., Wei, Z., Tam, K. Y. Cytoskeleton molecular motors: structures and their functions in neuron. International Journal of Biological Sciences. 12 (9), 1083-1092 (2016).
- Ajeti, V. et al. Wound healing coordinates actin architectures to regulate mechanical work. Nature Physics. 15 (7), 696-705 (2019).
- Jung, W. et al. Dynamic motions of molecular motors in the actin cytoskeleton. Cytoskeleton. 76 (11-12), 517-531 (2019).
- Pollard, T. D., O'Shaughnessy, B. Molecular mechanism of cytokinesis. Annual Review of Biochemistry. 88 (1), 661-689 (2019).
- Huber, F., Boire, A., López, M. P., Koenderink, G. H. Cytoskeletal crosstalk: when three different personalities team up. Current Opinion in Cell Biology. 32, 39-47 (2015).
- Rivero, F. et al. The role of the cortical cytoskeleton: F-actin crosslinking proteins protect against osmotic stress, ensure cell size, cell shape and motility, and contribute to phagocytosis and development. Journal of Cell Science. 109 (11), 2679-2691 (1996).
- Duclos, G. et al. Topological structure and dynamics of three-dimensional active nematics. Science. 367 (6482), 1120-1124 (2020).
- Baclayon, M. et al. Optical tweezers-based measurements of forces and dynamics at microtubule ends. Optical Tweezers. 1486, 411-435 (2017).
- Gurmessa, B., Fitzpatrick, R., Falzone, T. T., Robertson-Anderson, R. M. Entanglement density tunes microscale nonlinear response of entangled actin. Macromolecules. 49 (10), 3948-3955 (2016).
- Francis, M. L. et al. Non-monotonic dependence of stiffness on actin crosslinking in cytoskeleton composites. Soft Matter. 15 (44), 9056-9065 (2019).
- Ricketts, S. N. et al. Varying crosslinking motifs drive the mesoscale mechanics of actin-microtubule composites. Scientific Reports. 9 (1), 12831 (2019).
- Lee, G. et al. Active cytoskeletal composites display emergent tunable contractility and restructuring. Soft Matter. 17 (47), 10765-10776 (2021).
- Murrell, M. P., Gardel, M. L. F-actin buckling coordinates contractility and severing in a biomimetic actomyosin cortex. Proceedings of the National Academy of Sciences. 109 (51), 20820-20825 (2012).
- Soares e Silva, M. et al. Active multistage coarsening of actin networks driven by myosin motors. Proceedings of the National Academy of Sciences. 108 (23), 9408-9413 (2011).
- Sonn-Segev, A., Bernheim-Groswasser, A., Roichman, Y. Dynamics in steady state in vitro acto-myosin networks. Journal of Physics: Condensed Matter. 29 (16), 163002 (2017).
- Ideses, Y., Sonn-Segev, A., Roichman, Y., Bernheim-Groswasser, A. Myosin II does it all: assembly, remodeling, and disassembly of actin networks are governed by myosin II activity. Soft Matter. 9 (29), 7127 (2013).
- Fürthauer, S. et al. Self-straining of actively crosslinked microtubule networks. Nature Physics. 15 (12), 1295-1300 (2019).
- Lemma, L. M. et al. Multiscale microtubule dynamics in active nematics. Physical Review Letters. 127 (14), 148001 (2021).
- Fan, Y., Wu, K.-T., Aghvami, S. A., Fraden, S., Breuer, K. S. Effects of confinement on the dynamics and correlation scales in kinesin-microtubule active fluids. Physical Review E. 104 (3), 034601 (2021).
- Triclin, S. et al. Self-repair protects microtubules from destruction by molecular motors. Nature Materials. 20 (6), 883-891 (2021).
- Lee, G. et al. Myosin-driven actin-microtubule networks exhibit self-organized contractile dynamics. Science Advances. 7 (6), eabe4334 (2021).
- Ricketts, S. N., Ross, J. L., Robertson-Anderson, R. M. Co-entangled actin-microtubule composites exhibit tunable stiffness and power-law stress relaxation. Biophysical Journal. 115 (6), 1055-1067 (2018).
- Bendix, P. M. et al. A quantitative analysis of contractility in active cytoskeletal protein networks. Biophysical Journal. 94 (8), 3126-3136 (2008).
- Linsmeier, I. et al. Disordered actomyosin networks are sufficient to produce cooperative and telescopic contractility. Nature Communications. 7 (1), 12615 (2016).
- Stam, S. et al. Filament rigidity and connectivity tune the deformation modes of active biopolymer networks. Proceedings of the National Academy of Sciences. 114 (47), E10037-E10045 (2017).
- Yadav, V. et al. Filament nucleation tunes mechanical memory in active polymer networks. Advanced Functional Materials. 29 (49), 1905243 (2019).
- Ennomani, H. et al. Architecture and connectivity govern actin network contractility. Current Biology. 26 (5), 616-626 (2016).
- Alvarado, J., Sheinman, M., Sharma, A., MacKintosh, F. C., Koenderink, G. H. Molecular motors robustly drive active gels to a critically connected state. Nature Physics. 9 (9), 591-597 (2013).
- Alvarado, J., Cipelletti, L., Koenderink, G. H. Uncovering the dynamic precursors to motor-driven contraction of active gels. Soft Matter. 15 (42), 8552-8565 (2019).
- Jung, W., Murrell, M. P., Kim, T. F-actin cross-linking enhances the stability of force generation in disordered actomyosin networks. Computational Particle Mechanics. 2 (4), 317-327 (2015).
- Lenz, M., Thoresen, T., Gardel, M. L., Dinner, A. R. Contractile units in disordered actomyosin bundles arise from f-actin buckling. Physical Review Letters. 108 (23), 238107 (2012).
- Memarian, F.L. et al. Active nematic order and dynamic lane formation of microtubules driven by membrane-bound diffusing motors. Proceedings of the National Academy of Sciences. 118 (52), e2117107118 (2021).
- Needleman, D., Dogic, Z. Active matter at the interface between materials science and cell biology. Nature Reviews Materials. 2 (9), 17048 (2017).
- Foster, P. J., Fürthauer, S., Shelley, M. J., Needleman, D. J. Active contraction of microtubule networks. eLife. 4, e10837 (2015).
- Thijssen, K. et al. Submersed micropatterned structures control active nematic flow, topology, and concentration. Proceedings of the National Academy of Sciences. 118 (38), e2106038118 (2021).
- Sanchez, T., Chen, D. T. N., DeCamp, S. J., Heymann, M., Dogic, Z. Spontaneous motion in hierarchically assembled active matter. Nature. 491 (7424), 431-434 (2012).
- Colen, J. et al. Machine learning active-nematic hydrodynamics. Proceedings of the National Academy of Sciences. 118 (10), e2016708118 (2021).
- Mitchell, K. A., Tan, A. J., Arteaga, J., Hirst, L. S. Fractal generation in a two-dimensional active-nematic fluid. Chaos: An Interdisciplinary Journal of Nonlinear Science. 31 (7), 073125 (2021).
- Pandolfi, R. J., Edwards, L., Johnston, D., Becich, P., Hirst, L. S. Designing highly tunable semiflexible filament networks. Physical Review E. 89 (6), 062602 (2014).
- Tan, A. J. et al. Topological chaos in active nematics. Nature Physics. 15 (10), 1033-1039 (2019).
- Roostalu, J., Rickman, J., Thomas, C., Nédélec, F., Surrey, T. Determinants of polar versus nematic organization in networks of dynamic microtubules and mitotic motors. Cell. 175 (3), 796-808.e14 (2018).
- Ndlec, F. J., Surrey, T., Maggs, A. C., Leibler, S. Self-organization of microtubules and motors. Nature. 389 (6648), 305-308 (1997).
- Sheung, J. Y. et al. Motor-driven restructuring of cytoskeleton composites leads to tunable time-varying elasticity. ACS Macro Letters. 10 (9), 1151-1158 (2021).
- McGorty, R. PyDDM v0.2.0. Zenodo. (2022).
- Achiriloaie, D. H. et al. Kinesin and myosin motors compete to drive rich multi-phase dynamics in programmable cytoskeletal composites. arXiv. (2021).
- Wulstein, D. M., Regan, K. E., Garamella, J., McGorty, R. J., Robertson-Anderson, R. M. Topology-dependent anomalous dynamics of ring and linear DNA are sensitive to cytoskeleton crosslinking. Science Advances. 5 (12), eaay5912 (2019).
- McGorty, R. Image-Correlation. at <https://github.com/rmcgorty/Image-Correlation>. (2020).
- Robertson, C. Theory and practical recommendations for autocorrelation-based image correlation spectroscopy. Journal of Biomedical Optics. 17 (8), 080801 (2012).
- McGorty, R. Differential Dynamic Microscopy - Python. at <https://github.com/rmcgorty/Differential-Dynamic-Microscopy---Python>. (2021).
- Cerbino, R., Trappe, V. Differential dynamic microscopy: probing wave vector dependent dynamics with a microscope. Physical Review Letters. 100 (18), 188102 (2008).
- Robertson-Anderson, R. M. Optical tweezers microrheology: from the basics to advanced techniques and applications. ACS Macro Letters. 7 (8), 968-975 (2018).
- Pollard, T. D. Polymerization of ADP-actin. Journal of Cell Biology. 99 (3), 769-777 (1984).
- Coué, M., Brenner, S. L., Spector, I., Korn, E. D. Inhibition of actin polymerization by latrunculin A. FEBS Letters. 213 (2), 316-318 (1987).
- Pollard, T. D. Actin and actin-binding proteins. Cold Spring Harbor Perspectives in Biology. 8 (8), a018226 (2016).
- Kumar, N. Taxol-induced polymerization of purified tubulin. Mechanism of action. Journal of Biological Chemistry. 256 (20), 10435-10441 (1981).
- Käs, J. et al. F-actin, a model polymer for semiflexible chains in dilute, semidilute, and liquid crystalline solutions. Biophysical Journal. 70 (2), 609-625 (1996).
- Viamontes, J., Narayanan, S., Sandy, A. R., Tang, J. X. Orientational order parameter of the nematic liquid crystalline phase of F -actin. Physical Review E. 73 (6), 061901 (2006).
- Hitt, A. L., Cross, A. R., Williams, R. C. Microtubule solutions display nematic liquid crystalline structure. Journal of Biological Chemistry. 265 (3), 1639-1647 (1990).
- Andexer, J. N., Richter, M. Emerging enzymes for ATP regeneration in biocatalytic processes. ChemBioChem. 16 (3), 380-386 (2015).
- Farhadi, L. et al. Actin and microtubule crosslinkers tune mobility and control co-localization in a composite cytoskeletal network. Soft Matter. 16 (31), 7191-7201 (2020).
- Falzone, T. T., Lenz, M., Kovar, D. R., Gardel, M. L. Assembly kinetics determine the architecture of α-actinin crosslinked F-actin networks. Nature Communications. 3 (1), 861 (2012).
- Thoresen, T., Lenz, M., Gardel, M. L. Reconstitution of contractile actomyosin bundles. Biophysical Journal. 100 (11), 2698-2705 (2011).
- Sanghvi-Shah, R., Weber, G. F. Intermediate filaments at the junction of mechanotransduction, migration, and development. Frontiers in Cell and Developmental Biology. 5, 81 (2017).
- Shen, Y. et al. Effects of vimentin intermediate filaments on the structure and dynamics of in vitro multicomponent interpenetrating cytoskeletal networks. Physical Review Letters. 127 (10), 108101 (2021).
- Laan, L., Roth, S., Dogterom, M. End-on microtubule-dynein interactions and pulling-based positioning of microtubule organizing centers. Cell Cycle. 11 (20), 3750-3757 (2012).
Erratum
Erratum: Reconstituting and Characterizing Actin-Microtubule Composites with Tunable Motor-Driven Dynamics and MechanicsAn erratum was issued for: Reconstituting and Characterizing Actin-Microtubule Composites with Tunable Motor-Driven Dynamics and Mechanics. The Authors section was updated.
Mehrzad Sasanpour1
Daisy H. Achiriloaie1,2
Gloria Lee1
Gregor Leech1
Christopher Currie1
K. Alice Lindsay3
Jennifer L. Ross3
Ryan J. McGorty1
Rae M. Robertson-Anderson1
1Department of Physics and Biophysics, University of San Diego
2W. M. Keck Science Department, Scripps College, Pitzer College, and Claremont McKenna College
3Department of Physics, Syracuse University
to:
Mehrzad Sasanpour1
Daisy H. Achiriloaie1,2
Gloria Lee1
Gregor Leech1
Maya Hendija1
K. Alice Lindsay3
Jennifer L. Ross3
Ryan J. McGorty1
Rae M. Robertson-Anderson1
1Department of Physics and Biophysics, University of San Diego
2W. M. Keck Science Department, Scripps College, Pitzer College, and Claremont McKenna College
3Department of Physics, Syracuse University
Tags
ABOUT JoVE
Copyright © 2024 MyJoVE Corporation. All rights reserved