
Summary
Abstract
Introduction
Protocol
Representative Results
Discussion
Acknowledgements
Materials
References
Biology
Rekonstituering og karakterisering av aktin-mikrotubulikompositter med justerbar motordrevet dynamikk og mekanikk
ERRATUM NOTICE
Important: There has been an erratum issued for this article. Read more …Dette papiret presenterer protokoller for prosjektering og karakterisering av justerbare tredimensjonale sammensatte nettverk av sammenfiltrede aktinfilamenter og mikrotubuli. Kompositter gjennomgår aktiv restrukturering og ballistisk bevegelse, drevet av myosin II og kinesinmotorer, og er innstilt av de relative konsentrasjonene av aktin, mikrotubuli, motorproteiner og passive tverrbindinger.
Det sammensatte cytoskelettet, som består av interagerende nettverk av semifleksible aktinfilamenter og stive mikrotubuli, omstrukturerer og genererer krefter ved hjelp av motorproteiner som myosin II og kinesin for å drive viktige prosesser som migrasjon, cytokinese, adhesjon og mekanosensing. Mens aktin-mikrotubuli-interaksjoner er nøkkelen til cytoskelettets allsidighet og tilpasningsevne, er en forståelse av samspillet med myosin og kinesinaktivitet fortsatt begynnende. Dette arbeidet beskriver hvordan man konstruerer justerbare tredimensjonale sammensatte nettverk av sammenfiltrede aktinfilamenter og mikrotubuli som gjennomgår aktiv restrukturering og ballistisk bevegelse, drevet av myosin II- og kinesinmotorer, og er innstilt av de relative konsentrasjonene av aktin, mikrotubuli, motorproteiner og passive tverrbindinger. Protokoller for fluorescensmerking av mikrotubuli og aktinfilamenter for mest effektivt å visualisere kompositt restrukturering og bevegelse ved hjelp av multispektral konfokal avbildning er også detaljert. Til slutt presenteres resultatene av dataanalysemetoder som kan brukes til å kvantitativt karakterisere ikke-likevektsstruktur, dynamikk og mekanikk. Å gjenskape og undersøke denne justerbare biomimetiske plattformen gir verdifull innsikt i hvordan koblet motoraktivitet, komposittmekanikk og filamentdynamikk kan føre til utallige cellulære prosesser fra mitose til polarisering til mekano-sensasjon.
Cytoskelettet er et dynamisk sammensatt nettverk av samvirkende biopolymerer som gir strukturell og mekanisk støtte til celler. Assosierte molekylære motorer og bindende proteiner omstrukturerer og tilpasser cytoskelettet slik at cellene kan vokse, endre form, stivne, bevege seg og til og med helbrede seg selv, noe som muliggjør utallige cellulære prosesser som spenner fra migrasjon og deling til mekanosens 1,2. Utover sin betydning i cellulær biofysikk, er cytoskelettet også et typisk eksempel på aktiv materie med potensielle materialapplikasjoner som spenner fra sårheling og medikamentlevering til filtrering og myk robotikk 1,3,4,5,6,7,8,9.
De to nøkkelegenskapene som gir cytoskelettet sitt unike strukturelle og mekaniske mangfold og multifunksjonalitet er: 1) dets sammensatte natur, bestående av flere samvirkende proteinfilamenter, slik som semifleksible aktinfilamenter og stive mikrotubuli, samt deres tilknyttede bindings- og tverrbindingsproteiner 3,5,10; og 2) dens evne til kontinuerlig å omstrukturere, bevege seg, grov og utføre arbeid via energikrevende motorer, som myosiner og kinesiner, skyve og trekke på filamentøse proteiner 1,7,11,12,13. Selv om denne elegante kompleksiteten gjør det mulig for cytoskjelettet å formidle så forskjellige prosesser som cellemotilitet, cytokinese og sårheling 3,6,7,11, hemmer det forskernes evne til å reprodusere signaturen in vivo egenskaper av cytoskjelettet i rekonstituerte in vitro-systemer.
Nåværende grenserekonstitueringsarbeid fokuserer på kompositter av sammenfiltrede og tverrbundne aktinfilamenter og mikrotubuli 3,10,14,15,16,17, kraftgenererende aktomyosinnettverk 2,8,18,19,20,21, og aktiv nematikk drevet av kinesin-mikrotubuli interaksjoner 22,23,24,25,26. Steady-state aktin-mikrotubulikompositter har vist seg å vise fremvoksende mekaniske egenskaper15,16,27, for eksempel forbedret filamentmobilitet og økt stivhet sammenlignet med enkeltkomponentsystemer 27. Studier på in vitro aktomyosinsystemer har rapportert et bredt spekter av strukturelle og dynamiske egenskaper som avhenger av konsentrasjonene av aktin, myosin og tverrbindinger 28,29,30,31. For eksempel, med tilstrekkelig tverrbinding, gjennomgår aktomyosinnettverk storskala sammentrekning og grovhet 2,28,30,32,33,34,35,36, mens uten tverrbindinger viser nettverk rask, destabiliserende strømning og brudd 19,29 . Rekonstituert mikrotubulibasert aktiv nematikk som bruker klynger av kinesinmotorer til å krysslenke og trekke på mikrotubulibunter, har blitt rapportert å vise langvarige turbulente strømmer, forlengelse, knekking, oppsprekking og helbredelse 12,22,23,24,25,37,38,39,40,41, 42,43,44,45,46,47.
Mer nylig har aktin-mikrotubulikompositter drevet av myosin II minifilamenter vist seg å føre til mer bestilt sammentrekning og nettverksintegritet sammenlignet med den uordnede strømmen og nettverksbruddet som aktomyosinnettverk uten tverrbindinger viser 17,26,48. Videre optimaliseres kombinasjonen av komposittrobusthet og kraftgenerering når aktin og mikrotubuli er tilstede ved sammenlignbare konsentrasjoner. Viktige fremvoksende funksjoner i denne regionen av formuleringsrommet inkluderer forbedret mekanisk styrke 26, koordinert bevegelse av aktin og mikrotubuli26, jevn vedvarende sammentrekning og mesoskala restrukturering17.
Her beskrives protokoller for å konstruere og justere sammenfiltrede og tverrbundne kompositter av mikrotubuli og aktinfilamenter som skyves ut av likevekt av myosin II minifilamenter og kinesinklynger som virker på henholdsvis aktinfilamenter og mikrotubuli (figur 1). Dynamikken, strukturen og mekanikken til denne klassen av kompositter kan justeres av de relative konsentrasjonene av filamenter, motorer og tverrbindinger for å vise et rikt faserom med advektiv og turbulent strømning, isotropisk sammentrekning, akselerasjon, retardasjon, de-blanding, stivning, avslapning og brudd. Fokuset i dette arbeidet er å forberede og justere denne klassen av aktive cytoskeletale kompositter. Men for å hjelpe forskere med benchmarking og karakterisering av de beskrevne aktive komposittene, er effektive bildebehandlingsmetoder ved bruk av multispektral konfokalmikroskopi også detaljert. Til slutt presenteres resultater av viktige beregningsanalysemetoder som kan brukes til å måle dynamikken, strukturen og mekanikken til komposittene. Forskere oppfordres til å ta i bruk disse metodene - som inkluderer differensiell dynamisk mikroskopi (DDM), romlig bildeautokorrelasjon (SIA) og partikkelbildevelocimetry (PIV) - da de er optimalisert for å karakterisere den komplekse dynamikken og strukturelle mangfoldet av komposittene 17,26,49.
Trinnene beskrevet nedenfor fokuserer på å forberede komposittene og avbilde dem ved hjelp av konfokalmikroskopi. Protokoller som beskriver dataanalyse etter innsamling og optiske pinsettmålinger finnes i tidligere arbeider 17,26,48,50, og leveres på forespørsel. Alle materialer er oppført i tabellen over materialer som følger med.
1. Forbered silaniserte deksler og mikroskopglass for å forhindre adsorpsjon av proteiner til kammeroverflater
MERK: Dette er en 2-dagers prosess. Silaniserte lysbilder kan tilberedes opptil 1 måned før bruk.
- Plasser nr. 1 deksler (24 mm x 24 mm) og mikroskopglass (1 i x 3 tommer) i et angitt stativ som passer i plasmarenseren. Plasser rack i plasmarenser og kjør i 20 minutter.
- Overfør deksler og lysbilder til et nytt stativ som kun er beregnet for bruk med silan, og plasser stativet i glassbeholderen for å rengjøre brillene som beskrevet nedenfor.
- Senk omslag og lysbilder i 100% aceton i 1 time. Senk omslag og lysbilder i 100% etanol i 10 minutter.
- Senk deksler og sklier i avionisert vann (DI) i 5 minutter. Gjenta rengjøringstrinnene to ganger til.
- Senk coverslips og sklier i nyopparbeidede 0,1 M KOH i 15 min. Senk coverslips og lysbilder i fersk DI i 5 min. Gjenta dette trinnet to ganger til.
- Lufttørre deksler og lysbilder i 10 min. Behandle rensede deksler og skred med silan for å produsere hydrofobe overflater som beskrevet nedenfor.
MERK: Fullfør følgende trinn i en avtrekksvifte.- Senk tørkede deksler og lysbilder i 2% silan (oppløst i toluen) i 5 minutter. Bruk en trakt til å helle silan tilbake i den angitte flasken for å gjenbruke opptil fem ganger.
- Senk deksler og lysbilder i 100% etanol i 5 minutter. Erstatt etanol med fersk etanol. Senk omslagsslipp og lysbilder i 5 min.
- Senk coverslips og lysbilder i fersk DI i 5 min. Gjenta etanol- og DI-vasketrinnet to ganger til med fersk etanol og DI hver gang. Lufttørre deksler og lysbilder i 10 min.
2. Fremstilling av aktiv aktin-mikrotubulikompositt drevet av myosin minifilamenter
- Fjern inaktiv myosin via aktinfilamentbinding og utfør nedtrekk via ultrasentrifugering som beskrevet nedenfor.
- Polymeriser aktin i filamenter. Bruk en presisjonsmikropipette og sterile pipetspisser, kombiner i et mikrosentrifugerør: 1,87 μL DI, 1,3 μL 10x G-buffer, 1,3 μL 10x F-buffer, 1,63 μL av 4 M KCl, 4,53 μL aktin (47,6 μM) og 1,08 μL av 100 μM phalloidin.
MERK: For å sikre tilstrekkelig polymerisasjon, bør aktinkonsentrasjonen og aktin: phalloidinmolforholdet være henholdsvis 18,4 μM og 2: 1. - Pipet oppløsningen forsiktig opp og ned for å blande og sett deretter på is i mørket i ≥1 time. Avkjøl ultracentrifuge til 4 ° C. Fjern myosin aliquot fra -80 °C og legg på is.
MERK: Fullfør trinn 2.2 på dette tidspunktet mens aktin polymeriserer. - Etter ≥1 time aktinpolymerisasjon, tilsett 1,3 μL 10 mM ATP og 2 μL 19 μM myosin til det polymeriserte aktinet.
MERK: Forholdet mellom aktin: myosin skal være >5 for å sikre tilstrekkelig fjerning av inaktive myosinmotorer (dvs. døde hoder). - Pipet oppløsningen forsiktig opp og ned for å blande. Overfør til en ultracentrifuge grade tube.
Sentrifuge ved 4 °C og 121 968 x g i 30 minutter.
- Polymeriser aktin i filamenter. Bruk en presisjonsmikropipette og sterile pipetspisser, kombiner i et mikrosentrifugerør: 1,87 μL DI, 1,3 μL 10x G-buffer, 1,3 μL 10x F-buffer, 1,63 μL av 4 M KCl, 4,53 μL aktin (47,6 μM) og 1,08 μL av 100 μM phalloidin.
- Forbered sammenfiltret sammensatt nettverk av aktinfilamenter og mikrotubuli som beskrevet nedenfor.
MERK: Begynn 30 min før myosin spin-down (trinn 2.1.4).- Sett en varmeblokk til 37 °C. Bruk en presisjonsmikropipette og sterile pipetspisser for å legge til følgende i et mikrosentrifugerør: 13,9 μL PEM, 3 μL av 1% Tween20, 1,55 μL av 47,6 μM aktin, 0,36 μL av 34,8 μM R-aktin, 0,3 μL av 250 mM ATP, 0,87 μL av 100 μM phalloidin, 1,91 μL av 5-488-tubulin, 0,3 μL av 100 mM GTP, og 0,75 μL av 200 μM Taxol, til et totalt volum på 23 μL.
MERK: Konsentrasjonene av aktin og tubulin som er oppført er for en kompositt med 2,9 μM aktin og 2,9 μM tubulin. Total proteinkonsentrasjon er c = c A + c T = 5,8 μM og molar aktinfraksjon er c A / (cA + c T) = ΦA = 0, 5. Se trinn 2.5 for å justere disse verdiene. - Pip oppløsningen forsiktig opp og ned for å blandes og legg på en 37 °C varmeblokk beskyttet mot lys i 1 time.
- Sett en varmeblokk til 37 °C. Bruk en presisjonsmikropipette og sterile pipetspisser for å legge til følgende i et mikrosentrifugerør: 13,9 μL PEM, 3 μL av 1% Tween20, 1,55 μL av 47,6 μM aktin, 0,36 μL av 34,8 μM R-aktin, 0,3 μL av 250 mM ATP, 0,87 μL av 100 μM phalloidin, 1,91 μL av 5-488-tubulin, 0,3 μL av 100 mM GTP, og 0,75 μL av 200 μM Taxol, til et totalt volum på 23 μL.
- Forbered prøvekamre for konfokale avbildningseksperimenter som beskrevet nedenfor.
MERK: Fullfør trinn 2.1.4 og 2.2.2 i ventetiden.- Plasser to silaniserte lysbilder side om side på en kokeplate (slått av), legg to strimler termoplastisk tetningsfilm over lysbildene ~ 3 mm fra hverandre, og plasser to silaniserte deksler over den termoplastiske tetningsfilmen for å danne et prøvekammer.
- Vri kokeplaten på lav innstilling til dekselene binder seg fast til lysbilder med smeltet termoplastisk tetningsfilm (~ 1-2 min). Trykk ned med jevnt trykk for å sikre binding mens du opprettholder ~ 100 μm avstand mellom de to overflatene.
- Fjern kamrene og slå av kokeplaten. Merk kamre med (+) og (-). (+) kammeret vil være for den aktive prøven (med myosin) og (-) kammeret vil være kontrollen (ingen myosin). Sørg for at hvert kammer har plass til ≤10 μL væske.
- Klargjør prøver til bilde som beskrevet nedenfor.
MERK: Det er viktig å fullføre dette trinnet umiddelbart etter at trinn 2.1 og 2.2 er fullført.- Fjern myosin-aktinprøven forsiktig fra ultracentrifugen (trinn 2.1.4) og pipet umiddelbart opp de øverste 7,5 μL av supernatanten og overfør til et nytt mikrocentrifugerør.
- Fjern aktin-mikrotubuliprøven fra varmeblokken og bland forsiktig i 1,5 μL 10x D-glukose, 1,5 μL 10x GOC og 1,5 μL 1 mM blebbistatin. Del løsningen i to 13,7 μL aliquots og merk som (+) og (-).
- Bland inn 1,28 μL av supernatanten fra trinn 2.4.1 til (+) aliquot. Bland i 1,28 μL DI til (-) aliquot. Flyt hver løsning sakte inn i det tilsvarende kammeret (trinn 2.3) via kapillærvirkning. Vær forsiktig så du ikke introduserer luftbobler i kanalen.
- Forsegl de to åpne endene av hver kanal med hurtigtørkende epoksy- eller UV-lim. Sørg for at limet er helt tørt før du legger på mikroskopet. Bilde umiddelbart som beskrevet i trinn 3.
MERK: UV-lim er fordelaktig fordi det herder nesten umiddelbart ved UV-eksponering. Men fordi blebbistatin er UV-følsomt, er det viktig å bare lokalt belyse limet (ved kantene av prøvekammeret) ved hjelp av en liten UV-stav for å unngå å deaktivere blebbistatinet.
- Valgfritt: Varier proteinkonsentrasjonene for å justere dynamikken og strukturen til komposittene.
MERK: Følgende trinn er foreslåtte endringer i trinnene ovenfor for å variere konsentrasjonen av aktin, mikrotubuli og myosin om ønskelig.- Følg trinnene som er beskrevet ovenfor, bortsett fra følgende endringer i trinn 2.2.1 og 2.4.3.
- For å variere konsentrasjonen av aktin og mikrotubuli, og dermed justere c og ΦA, øke eller redusere volumet av aktin, R-aktin og 5-488-tubulin som brukes i trinn 2.2.1, etter ønske26. Når du varierer aktinkonsentrasjonen, justerer du R-aktin- og phalloidinmolarkonsentrasjonene proporsjonalt for å opprettholde de samme molare forholdene med aktin. Juster volumet av PEM slik at det endelige volumet av blandingen forblir 23 μL. Alle andre komponentvolumer og konsentrasjoner forblir de samme.
- For å variere myosinkonsentrasjonen, juster volumet av myosin tilsatt til (+) aliquot i trinn 2.4.3 etter ønske. Juster DI-volumet som er lagt til (-) aliquot tilsvarende. Juster PEM-volumet i trinn 2.2.1 for å ta hensyn til økningen eller reduksjonen i myosin (+) og DI (-) volum, slik at det endelige volumet av hver prøve ((+) og (-)) er 14,98 μL.
3. Avbildning og karakterisering av aktive kompositter ved hjelp av konfokalmikroskopi
- For å avbilde aktomyosin-mikrotubulikompositter fremstilt i trinn 2, bruk et laserskanning konfokalt mikroskop (LSCM) eller lignende mikroskop, med et 60x 1,4 NA oljenedsenkningsmål. For samtidig å visualisere aktinfilamenter og mikrotubuli i separate fluorescenskanaler, bruk en 561 nm laser med 565/591 nm eksitasjons- / utslippsfiltre og en 488 nm laser med 488/525 nm eksitasjons- / utslippsfiltre.
- Plasser prøvekammeret på mikroskopet slik at kontrollkanalen er plassert direkte over målet. Forsikre deg om at det er et oljegrensesnitt mellom målet og dekselet.
- Bruk scenekontrollene til å sette kontrollkompositten i fokus, og finn deretter begge overflatene på prøvekammeret. Flytt z-posisjonen til midten av prøvekammeret. Kontroller om det finnes klare filamentøse nettverk som vist i figur 2.
- Fremdeles visualisere kontrollkammeret, justere intensiteten av hver laser for å tillate samtidig visualisering av aktinfilamenter og mikrotubuli. Oppretthold lavest mulig laserintensitet for å forhindre fotobleking (mer utbredt i aktinkanalen) og blø gjennom (vanligvis fra mikrotubuli inn i aktinkanalen).
- For å karakterisere den inaktive kontrollprøven, samle tre tidsserier (videoer) på 256 x 256 kvadratpiksler (213 μm x 213 μm) bilder ved 2,65 fps for totalt ≥ 1000 bilder. Samle hver tidsserie i et annet område av prøvekammeret atskilt med ≥500 μm. Sørg for at det er minimal detekterbar bevegelse og ingen flyt eller omstrukturering.
- Lukk av 488 nm laseren og bruk scenekontrollene til å flytte til (+) kammeret.
- Ved hjelp av 568 nm laseren visualiserer du mikrotubuli i (+)-kanalen for å sikre riktig nettverksdannelse (figur 2) og identifiserer prøvekammerets aksiale senter (som kan være forskjellig fra kontrollkammerets midtre z-posisjon).
- Slå på 488 nm laser og gjenta trinn 3.5 ovenfor med følgende modifikasjoner. Samle tidsserier i opptil 45 minutter, og stopp oppkjøpet når prøven enten beveger seg ut av synsfeltet, brudd eller fotobleacher. Ta opp 5-10 tidsserier og hold oversikt over tidspunktet hver tidsserie begynner i forhold til starten av den første tidsserien.
- Analyser data ved hjelp av DDM, SIA og PIV som beskrevet i figur 3, figur 4, figur 5 og tidligere 17,48,50,51.
MERK: 488 nm laseren aktiverer lokalt myosin ATPase-aktivitet ved å deaktivere blebbistatin, så den bør bare slås på ved starten av datainnsamlingen slik at t = 0 er i starten av tidsserien. Disse oppkjøpsparametrene er optimalisert for differensiell dynamisk mikroskopi (DDM) analyse som gjort tidligere26.
4. Fremstilling av aktive aktin-mikrotubulikompositter drevet av kinesinmotorer
MERK: Følgende trinn skaper aktin-mikrotubulikompositter som drives ut av likevekt av kinesinmotorer eller en kombinasjon av kinesin og myosin50.
- Forbered kinesin- og myosinmotorer som beskrevet nedenfor.
- Hvis du inkorporerer myosin, følg trinn 2.1.
- For å danne kinesinmotorklynger som binder og utøver krefter mellom mikrotubulipar, bruk en mikropipette og sterile pipetspisser for å legge til følgende til et sterilt 1,5 ml mikrosentrifugerør: 1,16 μL PEM, 2,74 μL 8,87 μM kinesindimerer, 7,29 μL 83,3 μM NeutrAvidin, 0,81 μL 2mM DTT . Bland forsiktig ved å pipettere oppløsningen opp og ned og inkuber beskyttet mot lys (bruk et svart mikrosentrifugerør eller pakk inn folie) i 30 minutter ved 4 °C.
MERK: Det molare forholdet mellom kinesindimerer og NA er 1:25.
- Følg trinn 2.3 for å forberede prøvekamre og lage tre kamre i stedet for to. Utfør dette trinnet under kinesininkubasjon (trinn 4.1.2) og myosin ultracentrifugation (trinn 4.1.1).
- Forbered co-entangled kompositt nettverk av aktinfilamenter og mikrotubuli.
- Sett varmeblokken til 37 °C. Bruk en mikropipette og sterile pipetspisser for å legge til følgende til et sterilt 1,5 ml mikrosentrifugerør: 3,21 μL PEM, 4,5 μL av 1% Tween20, 2,18 μL av 47,6 μM aktin, 3,46 μL av 5-R-tubulin, 4,5 μL av 100 mM ATP, 4,5 μL av 10 mM GTP, 1,13 μL av 200 μM Taxol og 1,57 μL av 20 μM 488-phalloidin. Sørg for at totalvolumet er 25 μL.
- Pip oppløsningen forsiktig opp og ned for å blandes, og legg på 37 °C varmeblokken beskyttet mot lys i 1 time. Fjern røret fra varmeblokken og bruk en mikropipette til å blande forsiktig i 0,84 μL 100 μM phalloidin. Inkuber i 5-10 minutter ved romtemperatur, beskyttet mot lys.
MERK: Tilsetning av phalloidin på dette trinnet, i stedet for i trinn 4.3.1, forbedrer fluorescensmerkingen av aktinfilamenter, da 488-phalloidin ikke trenger å konkurrere med umerket phalloidin for aktinbindingssteder.
- Forbered aktive kompositter for konfokal avbildning.
- Tilsett 1,13 μL 200 μM blebbistatin, 1,35 μL 10x Glu og 1,35 μL 10x GOC til løsningen fra trinn 4.3.2 og bland forsiktig ved pipettering opp og ned. Del løsningen i tre 10 μL aliquots og merk som (K), (K + M) og (-).
- Bland inn 2,54 μL myosin fra trinn 2.1.4 til (K+M) aliquot. Bland i 2,54 μL PEM til (K) og (-) aliquoter.
- Bruk en mikropipette og sterile pipetspisser for å tilsette 2,5 μL kinesinklynger fra trinn 4.1.2 til (K) og (K+M) aliquoter. Pipet opp og ned for å blande. Bland i 2,5 μL PEM til (-) med samme teknikk.
MERK: Konsentrasjonene av aktin og tubulin som er oppført er for en kompositt med 2,32 μM aktin og 3,48 μM tubulin. Total proteinkonsentrasjon er c = c A + c T = 5,8 μM og molar aktinfraksjon er c A / (c A + cT) = Φ A = 0, 4. Kinesin- og myosinkonsentrasjonene er henholdsvis 0,35 μM og 0,47 μM. Se trinn 2.5 for generelle retningslinjer for justering av c A, cT, c og ΦA. - Ved hjelp av en mikropipette strømmer du hver oppløsning sakte inn i den tilsvarende kanalen til de forberedte prøvekamrene (trinn 4.2) via kapillærvirkning. Skyv pipet veldig sakte og forsiktig ned for ikke å føre luftbobler inn i kanalen.
- Forsegl de to åpne endene av hver kanal med hurtigtørkende epoksy eller UV-herdbart lim. Sørg for at limet er helt tørt før du legger på mikroskopet.
MERK: Det er viktig at dette trinnet gjøres raskt for å minimere tiden kinesinet virker uten å bli overvåket. Av denne grunn anbefales epoksy som herder på 1 minutt (i stedet for 5 eller 10 minutter). UV-herdbart lim er fordelaktig i denne forbindelse fordi det herder nesten umiddelbart ved UV-eksponering.
- Bilde forberedte prøver umiddelbart, etter trinn 3, bortsett fra følgende viktige modifikasjoner. Fordi kinesin ikke styres av lysaktivering, begynner det å fungere umiddelbart etter trinn 4.4.3, så merk denne gangen som t = 0. Hvis du vil avbilde sammensetningen så nær den opprinnelige inaktive tilstanden (t = 0) som mulig, kan du først se for deg kanalene (K) og (K+M) og notere tiden som har gått mellom trinn 4.4.3 og begynnelsen av datainnsamlingen (trinn 3.8). I praksis er denne forløpte tiden ~ 5 min.
5. Innlemme passive tverrbindinger i aktive kompositter
MERK: Disse trinnene beskriver hvordan biotinylerte aktin- og tubulinunderenheter og NeutrAvidin (NA) passivt kryssbinder aktin til aktin (AA) eller mikrotubuli til mikrotubuli (M-M) i de aktive komposittene beskrevet i trinn 4.
- Forbered A-A eller M-M tverrbindingskomplekser med biotinylerte proteiner (biotin-aktin eller biotin-tubulin), NA og biotin i forholdet 2: 2: 1 biotin-aktin / tubulin: biotin: NA. Start denne prosessen før trinn 4 .
- For A-A tverrbindinger, bruk en mikropipette og sterile pipetspisser for å legge til 2 μL 11,6 μM biotin-aktin, 1,39 μL av 8,33 μM NA, 2,27 μL av 1,02 μM biotin og 4,34 μL PEM til et mikrosentrifugerør. Bland forsiktig ved pipettering opp og ned.
- For M-M tverrbindinger, bruk en mikropipette og sterile pipetspisser for å legge til 1,86 μL av 4,55 μM biotin-tubulin, 1,11 μL av 8,33 μM NA, 1,82 μL av 1,02 μM biotin og 5,21 μL PEM til et mikrosentrifugerør. Bland forsiktig ved pipettering opp og ned.
- Pakk røret/rørene fra trinn 5.1.1 og/eller 5.1.2 i termoplastisk tetningsfilm for å skape en vanntett forsegling. Plasser i en flotasjonsflåte i et temperaturkontrollert sonikerbad satt til 4 ° C.
- Sonikere i 90 min ved 4 ° C. I praksis er det best å sette sonikatoren i et kaldt rom og legge ispakker til sonikeringsbadet for å opprettholde lav temperatur.
- Hvis du vil innlemme krysskoblingskomplekser i eksempler for bildebehandling, følger du trinn 4.3 og endrer trinn 4.3.1 som beskrevet nedenfor for A-A-kryssbinding (trinn 5.2.1) eller M-M-kryssbinding (trinn 5.2.2).
- For A-A tverrbinding, kombiner følgende i et mikrosentrifugerør: 1,94 μL PEM, 4,50 μL av 1% Tween20, 2,18 μL av 47,6 μM aktin, 3,46 μL av 45,5 μM 5-R-tubulin, 1,13 μL av A-A tverrbindinger (trinn 5,1,1), 4,50 μL av 100 mM ATP, 4,50 μL av 10 mM GTP, 1,13 μL av 200 μM Taxol, og 1,57 μL av 20 μM 488-phalloidin. Sørg for at totalvolumet er 25 μL.
- For M-M tverrbinding, kombiner følgende i et mikrosentrifugerør: 1,97 μL PEM, 4,50 μL av 1% Tween20, 2,18 μL av 47,6 μM aktin, 3,76 μL av 45,5 μM 5-R-tubulin, 1,13 μL av 1:4 fortynning av M-M tverrbindinger (trinn 5,1,2), 4,50 μL av 100 mM ATP, 4,50 μL av 10 mM GTP, 1,13 μL av 200 μM Taxol, og 1,57 μL av 20 μM 488-phalloidin. Sørg for at totalvolumet er 25 μL.
- Følg trinn 4.3.2-4.5 med de spesifikke konsentrasjonene for et tverrbinding: aktinmolforhold på RA = 0,02 og tverrbinding: tubulinmolarforhold på RT = 0,005. Disse R A- og R T-verdiene resulterer i lignende lengder mellom tverrbindinger langs aktinfilamenter og mikrotubuli (d A 60 nm og d MT 67 nm), estimert ved bruk av d A = I monomer / 2R A
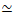 , hvorI-monomer er lengden på en aktinmonomer, og dMT = I ring / 26R T, hvor jegringer er lengden på enring på 13 tubuliner15, 17.
, hvorI-monomer er lengden på en aktinmonomer, og dMT = I ring / 26R T, hvor jegringer er lengden på enring på 13 tubuliner15, 17.
For å bestemme vellykket fremstilling av aktive kompositter (figur 1), og for å karakterisere deres dynamikk og struktur, brukes et laserskanningsfluorescensmikroskop med minst to fluorescenskanaler for å visualisere aktinfilamentene og mikrotubuli samtidig (figur 2 og figur 6). Alle aktinfilamenter og mikrotubuli i komposittene er sparsomt merket, i stedet for doping i sporstofflyse filamenter, slik det ofte gjøres i in vitro-studier. Denne metoden sikrer at den målte dynamikken og strukturen er representativ for selve kompositten i stedet for sporstoffene som dannes under andre forhold enn komposittene. Av denne grunn kan individuelle aktinfilamenter og mikrotubuli vanligvis ikke løses, snarere viser bilder mesoskala nettverksstruktur (figur 2 og figur 6).
Denne merkingsmetoden ble optimalisert for romlig bildeautokorrelasjon (SIA) og differensialdynamisk mikroskopi (DDM) analyser som undersøker dynamikken og strukturen i gjensidig Fourier-rom (figur 4, figur 5 og figur 8) 52,53,54,55. Partikkelbilde-velocimetry (PIV) kan også brukes til å skildre og karakterisere dynamikk og strømningsfelt (figur 3 og figur 7), men det krever pikselbinning (lavere romlig oppløsning) og større forsinkelsestidsintervaller (lavere tidsmessig oppløsning) enn SIA og DDM for å eliminere feilaktige vektorer som oppstår fra støy i de tette lavsignalbildene. Likevel anbefales PIV for kvalitativ undersøkelse av strømningsfelt og bekreftelse av DDM-resultater (figur 4 og figur 8)26,50.
Prøvekarakterisering av de beskrevne nettverkene ved hjelp av disse analysene (dvs. DDM, SIA, PIV) er gitt for å hjelpe forskere med å vedta lignende analyser for å benchmark og karakterisere sine prøver. Detaljerte beskrivelser av disse teknikkene ligger imidlertid utenfor rammen av dette arbeidet. For detaljerte beskrivelser av hvordan du utfører DDM på disse og andre lignende systemer, inkludert brukervennlig Python-kode, se tidligere verk 17,26,49,50 og referansene der innenfor. For detaljer om hvordan du utfører SIA og PIV på systemene beskrevet her, blir leseren henvist til tidligere arbeider17,50.
Flere kontroller, beskrevet nedenfor, bør gjøres for å sikre at komposittene fungerer som forventet. En kompositt uten myosin eller kinesin skal virke i hovedsak statisk med minimale termiske svingninger eller drift. Aktinfilamenter og mikrotubuli skal virke sammenfiltret og homogent fordelt, med minimal bunting, aggregering eller faseseparasjon av aktin og mikrotubuli i et synsfelt på ~200 μm x 200 μm (figur 2, helt til venstre)17. Man bør forvente et lignende resultat for kompositter som inneholder myosin, men som ikke er utsatt for 488 nm lys (for å deaktivere blebbistatin).
Ved inkorporering av myosin og eksponering for 488 nm lys gjennomgår komposittene sammentrekninger som i stor grad er isotrope og like for aktin og mikrotubuli, som vist på mikroskopbilder tatt før og etter myosinaktivitet (figur 2), samt tilsvarende PIV-strømningsfelt for varierende tider under aktivitet (figur 3). For å avgjøre om bevegelsen er ballistisk, diffusiv, subdiffusiv, etc., evalueres den karakteristiske dekorrelasjonstiden τ (q) bestemt fra DDM som en funksjon av bølgevektor (dvs. gjensidig rom). Se som beskrevet i detalj tidligere 17,26,49. Figur 4 viser også hvordan man bruker DDM til å karakterisere disse komposittene. Power-law skalering τ (q) ~ 1 / vq β, med β = 1, indikerer ballistisk bevegelse med hastighet v. Til referanse representerer β = 2 diffusiv dynamikk med v som diffusjonskoeffisienten. Alle aktive kompositter viser ballistisk skalering (figur 4A) med hastigheter som er innstilt av konsentrasjonene av aktin og myosin (figur 4B), og som kan variere i tid under aktivitet, enten akselerere eller bremse (figur 4C,D).
Nettverksrestrukturering og klynger, synlig i figur 2 og tydeligere for høyere aktin- og myosinkonsentrasjoner, kan karakteriseres ved hjelp av SIA, som vist i figur 5, og beskrevet tidligere17,48,50. Kort fortalt kan en korrelasjonslengde ξ, som er et mål på den karakteristiske størrelsen på funksjoner i et bilde, bestemmes ved å tilpasse hver romlige intensitetsautokorrelasjonskurve g (r) til en eksponentiell funksjon av avstand r mellom piksler. Større g (r) topper som vedvarer i lengre avstander indikerer større strukturelle egenskaper (dvs. bunting, klynger av de enkelte filamenter). Som vist i figur 5, for høyere aktinfraksjoner og myosinkonsentrasjoner, gjenspeiles signifikant restrukturering og aggregering i økningen i ξ over tid.
De viskoelastiske egenskapene og den ikke-lineære mekaniske responsen til de aktive komposittene kan også måles ved hjelp av optisk pinsettmikrorheologi (OTM). Protokoller og representative resultater for disse eksperimentene er imidlertid utenfor omfanget av dette arbeidet. Interesserte lesere henvises til tidligere arbeider48,56 som grundig beskriver hvordan man utfører OTM-målinger og forventede resultater.
Ved å bruke det samme programmet med eksperimentelle og analyseverktøy som er beskrevet ovenfor, beskriver følgende avsnitt hvordan dynamikken og strukturen endres når kinesinmotorer og biotin-NA-tverrbindinger er innlemmet i komposittene (figur 6, figur 7 og figur 8). Figur 6 viser representative konfokale bilder av kompositter drevet av enten kinesin-only (K) eller kinesin og myosin (K+M), med og uten passiv tverrbinding (XL) av aktinfilamenter eller mikrotubuli.
Inkorporering av kinesin i kompositter resulterer i utgangspunktet i lignende dynamikk og restrukturering som myosindrevne kompositter som vist i den øverste raden i figur 7 (klasse 1). Dynamikken går imidlertid vanligvis over til storskala anisotropisk strømning (figur 7 midtre rad, klasse 2), akselerasjon og retardasjon (figur 7 nederste rad, klasse 3). Disse egenskapene kombineres med mesoskala clustering og aggregering etter 5-30 min (figur 6 og figur 8B). PIV-genererte strømningsfelt og tidsmessige fargekart vist i figur 7 viser eksempler på isotropisk restrukturering (klasse 1, topppanel), rettet flyt (klasse 2, midtpaneler) og toveis akselerasjon (klasse 3, bunnpaneler).
Hastigheter av aktin og mikrotubuli ved varierende tidspunkter under aktivitet, bestemt via passer til τ (q) kurver, illustrerer akselerasjon etterfulgt av retardasjon (figur 8), som avhenger av tverrbinding. Som også vist i figur 8, når begge motoriske proteiner er innlemmet, er dynamikken faktisk langsommere enn kinesin-bare kompositter, og det er forsinket utbrudd av mesoskala strømning. Myosin støtter også mer homogen interpenetrasjon av aktin- og mikrotubulinettverk gjennom hele aktiviteten, samt mindre aggregering og restrukturering. Disse effektene kan ses på bildene i figur 6 og kvantifiseres av de tidsvarierende korrelasjonslengdene beregnet via SIA, som vanligvis er mindre i nærvær av myosin (figur 8B).
Figur 1. Design og karakterisering av aktive aktin-mikrotubulikompositter med flere kraftgenererende motorer og passive tverrbindinger. (A) Aktinmonomerer og tubulindimerer kopolymeriseres ved molare konsentrasjoner c Aog c T på 0,73-11,6 μM og molare fraksjoner av aktin Φ A = c A / (c A + c T) = 0, 0,25, 0,5, 0,75 og 1, for å danne sammenfiltrede nettverk av aktinfilamenter (grønn) og mikrotubuli (rød). Passiv tverrbinding oppnås ved bruk av NA for å koble biotinylerte aktinfilamenter (Actin XL) eller mikrotubuli (MT XL) ved tverrbinding: proteinmolare forhold på R A = 0,01-0,08 og RMT = 0,001-0,01 for henholdsvis aktin og mikrotubuli. Myosin-II minifilamenter (lilla) og kinesinklynger (oransje), i konsentrasjoner av c M = 0,12 - 0,48 μM og cK = 0,2 - 0,7 μM, skyv og trekk i filamentene for å drive komposittene ut av steady-state. (B) Skjematisk formuleringsrom. Myosin II minifilamenter (M), kinesinklynger (K) eller begge motorene (K+M) er inkorporert i kompositter uten passive tverrbindinger (No XL), aktin-aktin tverrbindinger (Actin XL) og mikrotubuli-mikrotubuli tverrbindinger (MT XL). Alle tegneserier er ikke tegnet i skala. Vennligst klikk her for å se en større versjon av denne figuren.
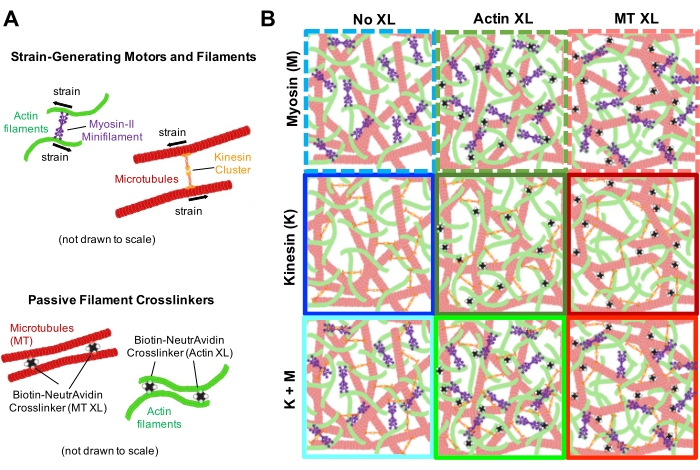
Figur 2. Tofarget konfokal avbildning av myosindrevne cytoskelettkompositter med varierende myosinkonsentrasjoner cM og molare aktinfraksjoner ΦA. (A) 256 x 128 kvadratpiksler (212 x 106 μm2) tofargede konfokale mikroskopibilder viser hvordan kompositter av aktinfilamenter (grønn) og mikrotubuli (rød) omorganiseres via myosinmotorisk aktivitet. Ingen kinesinmotorer eller passive tverrbindinger er til stede. I hvert panel vises bilder tatt i begynnelsen (venstre, før) og slutten (høyre, etter) av 45 min myosinaktivering (via belysning med 488 nm lys for å deaktivere blebbistatin). Paneler bestilles ved å øke molar konsentrasjon av myosin (cM), gå fra venstre til høyre, og øke molar fraksjon av aktin (ΦA), går fra topp til bunn. Fargene som beskriver hvert panel, samsvarer med fargekodingen som brukes i figur 4 og figur 5. Skala barer er 50 μM. For best å fange dynamikk og struktur for analyse, bruker vi bildefrekvenser på 1-5 fps, avkastning med 50-250 μm sider og tidsserievarigheter på 5-45 min, avhengig av hastigheten på sammentrekning og omorganisering. Paneler der før- og etterbildene ser like ut, indikerer minimal restrukturering, som sett i rosa-, magenta- og cyanpanelene. Småskala klynger, dokumentert av økt heterogenitet og tilstedeværelsen av lyse punktlige funksjoner, kan ses i de oransje, grønne og røde panelene. Storskala sammentrekning, sett på som et jevnt krympende nettverk, er tydelig i de blå og lilla panelene. Dette tallet er endret fra referanse17. Vennligst klikk her for å se en større versjon av denne figuren.
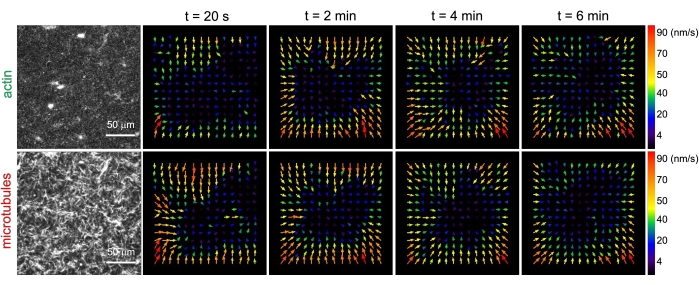
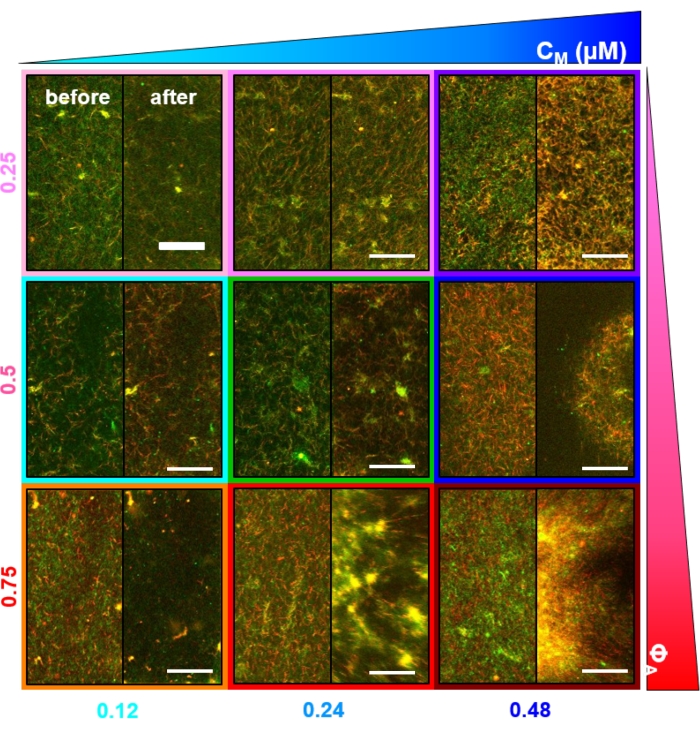
Figur 3. Partikkelbilde velocimetry (PIV) viser at actomyosinaktivitet utløser koordinert kontraktil dynamikk av aktin og mikrotubuli i sammenfiltrede kompositter. PIV strømningsfelt for aktin (øverste rad) og mikrotubuli (nederste rad) i en myosindrevet kompositt med (ΦA, cM) = (0,5, 0,24) ved økende tider i løpet av en 6 min tidsserie. Flytfelt ble generert ved hjelp av Fiji / ImageJ PIV-plugin med en forsinkelsestid på 20 s og 2 piksler x 2 piksel binning. Både aktin og mikrotubuli viser konsekvent bevegelse rettet mot sentrum av synsfeltet gjennom hele filmens varighet. Skalalinjer i alle bilder er 50 μm. Ulike pilfarger tilsvarer forskjellige hastigheter som angitt i fargeskalaen til høyre for vektorfelt. Dette tallet er endret fra referanse26. Vennligst klikk her for å se en større versjon av denne figuren.
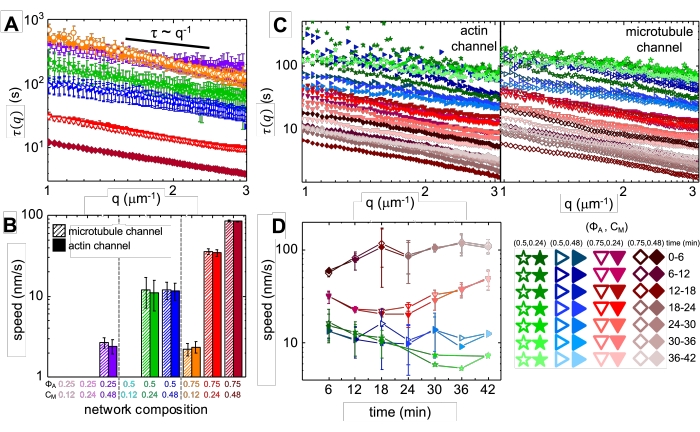
Figur 4. Tidsoppløst differensiell dynamisk mikroskopi (DDM) måler hastigheten og typen bevegelse av aktin og mikrotubuli i aktive kompositter. (A) DDM utføres på mikrotubuli (øverste, åpne symboler) og aktin (nederst, fylte symboler) kanaler av tidsserier for å bestemme karakteristiske forfallstider τ vs bølgetall q for både aktin (fylte symboler) og mikrotubuli (åpne symboler) som beskrevet tidligere17,26. Alle kurver følger τ ~ q-1 skalering, noe som indikerer ballistisk bevegelse, med hastigheter v som bestemmes via passer til τ (q) = (vq) -1. Raskere hastigheter tilsvarer mindre τ(q)-verdier for en gitt q. Symbolfarger og former tilsvarer (Φ A, cM) kombinasjoner vist i B. (B) Kontraksjonshastigheter v bestemmes via passer til τ(q)-kurver vist i A, som er gjennomsnitt over alle forsinkelsestider for varigheten av hver 45 min tidsserie. (C) Time-resolved DDM (trDDM) kvantifiserer hvordan dynamikken varierer over tid ved å evaluere τ(q) for aktin (fylte symboler, venstre) og mikrotubuli (åpne symboler, høyre) i påfølgende 6 min intervaller (betegnet med forskjellige nyanser av samme farge) i løpet av 45 min aktiveringstid. trDDM utføres for hver (ΦA, cM) kombinasjon (betegnet med forskjellige symboler og farger) som beskrevet i forklaringen nederst til høyre. τ(q)-kurver vist i C følger lignende skalering og trender som de i A, men viser også tidsavhengighet for visse (Φ A, cM) komposisjoner, spesielt for ΦA= 0,75. (D) Kontraksjonshastigheter for aktinfilamenter (lukkede symboler) og mikrotubuli (åpne symboler) bestemmes fra tilpasninger til tilsvarende τ(q)-kurver. Feilfelt i alle plott representerer standardfeilen for verdier på tvers av tre til fem replikaer. Dette tallet er endret fra referanse17. Vennligst klikk her for å se en større versjon av denne figuren.
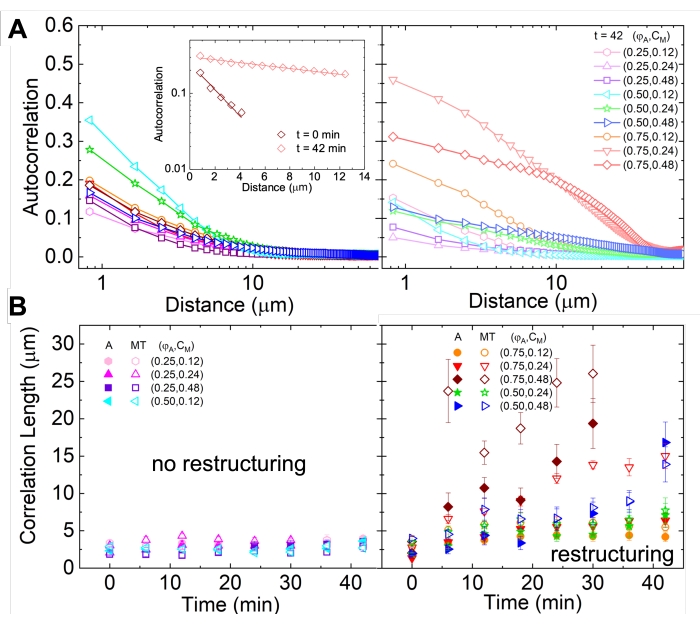
Figur 5. Romlig autokorrelasjonsanalyse (SIA) kvantifiserer den motordrevne restruktureringen av aktive cytoskeletale kompositter. (A) Autokorrelasjon g(r) for mikrotubuli i begynnelsen (venstre, t = 0 min, mørke nyanser) og slutten (høyre, t = 42 min, lyse nyanser) av eksperimentet for (ΦA, cM) formuleringer oppført i forklaringen. Innfelt: eksempel passer på data til 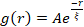 ved innledende og endelige tider for (ΦA, cM) = (0,75, 0,12). (B) Gjennomsnittlige korrelasjonslengder ξ for aktin (lukkede symboler) og mikrotubuli (åpne symboler) for hver (Φ A, cM) bestemt via eksponentielle tilpasninger av hver g (r) kurve, som vist i innfelt i A. Data er delt inn i de som viser minimal (venstre) versus betydelig (høyre) restrukturering. Feilfelt i A og B representerer standardfeilen på tvers av tre til fem replikaer. Dette tallet er endret fra referanse17. Vennligst klikk her for å se en større versjon av denne figuren.
ved innledende og endelige tider for (ΦA, cM) = (0,75, 0,12). (B) Gjennomsnittlige korrelasjonslengder ξ for aktin (lukkede symboler) og mikrotubuli (åpne symboler) for hver (Φ A, cM) bestemt via eksponentielle tilpasninger av hver g (r) kurve, som vist i innfelt i A. Data er delt inn i de som viser minimal (venstre) versus betydelig (høyre) restrukturering. Feilfelt i A og B representerer standardfeilen på tvers av tre til fem replikaer. Dette tallet er endret fra referanse17. Vennligst klikk her for å se en større versjon av denne figuren.
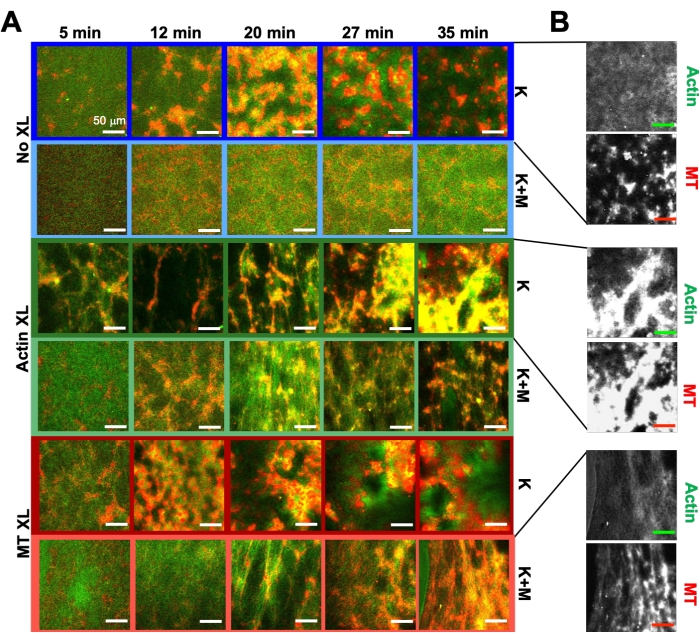
Figur 6. Inkorporering av kinesinmotorer og passive tverrbindinger i aktive kompositter for å øke programmerbarheten og utvide faserommet for dynamikk og struktur. (A) Tofargede konfokale bilder av aktin (grønn) og mikrotubuli (rød) i aktive kompositter viser kompleks formuleringsavhengig restrukturering over tid (oppført i min). De fem bildene i hver rad tilsvarer fem rammer av en tidsserie på 2000 rammer anskaffet for en kompositt drevet av kinesin (K, rad 1, 3, 5) eller kinesin og myosin (K + M, rad 2, 4, 6), og inkluderer enten ingen passive tverrbindinger (Ingen XL, rad 1, 2), aktin-aktin tverrbindinger (Actin XL, rad 3, 4) eller mikrotubuli-mikrotubuli tverrbindinger (MT XL, rad 5, 6). Skala barer er alle 50 μm. Omrissfarger samsvarer med fargevalget i figur 8. (B) Separate aktin- og mikrotubulifluorescenskanaler for kinesinkomposittene viser varierte strukturer med både aktin-MT-kolokalisering og mikrofaseseparasjon. Bildene som vises er for kompositter med c A= 2,32 μM, c T= 3,48 μM, c K = 0,35 μM, cM= 0,47 μM (rad 2, 4, 6), R A= 0,02 (rad 3, 4) og RMT = 0,005 (rad 5, 6). Alle kompositter begynner med jevnt fordelte interpenetrerende nettverk av aktin og mikrotubuli (kolonne 1). Kinesindrevne kompositter uten tverrbindinger (rad 1) danner løst tilkoblede amorfe klynger som er MT-rike. Actin samlokaliserer i sentrene til disse aggregatene i utgangspunktet, men blir deretter presset ut av de MT-rike regionene som fortsetter å trekke seg sammen og koble fra hverandre. Kryssbinding av aktin-aktin (rad 3) hindrer denne mikroskala actin-MT-separasjonen, og i stedet er MT-rike aggregater forbundet via lange tråder av aktin. Aktin kryssbinding muliggjør også langsom opptak av aktin i de MT-rike regionene, slik at kompositten blir et tilkoblet nettverk av samlokaliserte aktin- og MT-klynger. Mikrotubuli kryssbinding (rad 5) fører til amorf klynge av MT som samler seg over tid, noe som resulterer i større skala fase separasjon av aktin og MTs. Tilsetning av myosin (rad 2, 4, 6) reduserer kinesindrevet de-blanding og restrukturering. Uten tverrbindinger (rad 2) viser kompositter lite omorganisering i løpet av timer. Tverrbinding øker restrukturering og samlokalisering av aktin og mikrotubuli (rad 4, 6). Spesielt når mikrotubuli er tverrbundet (rad 6), er det betydelig interpenetrasjon og omorganisering i nettlignende nettverk av fibre. Dette tallet er endret fra referanse50. Vennligst klikk her for å se en større versjon av denne figuren.
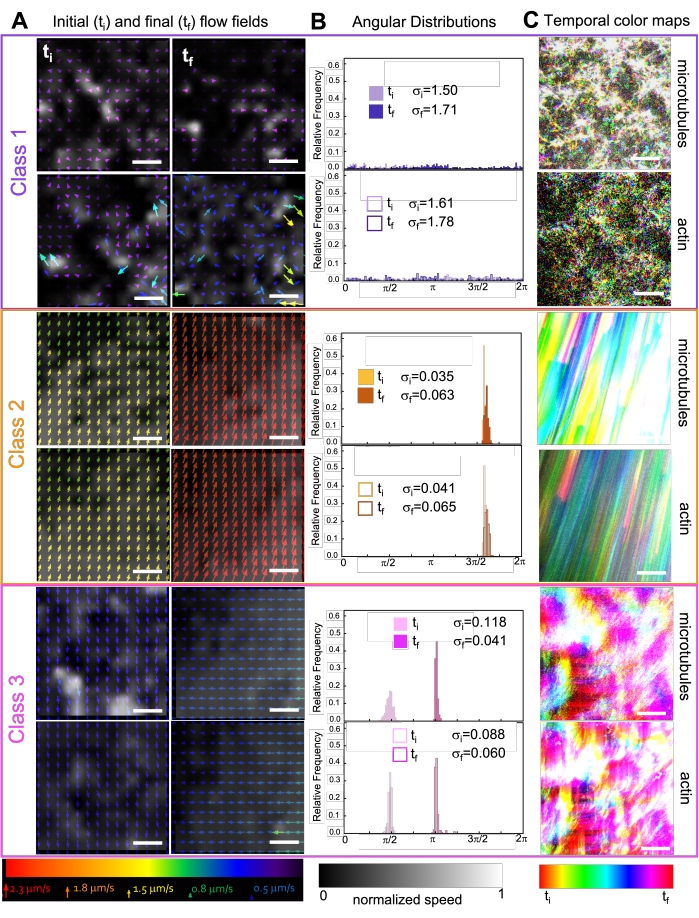
Figur 7. PIV viser at aktive kompositter viser tre klasser av spatiotemporalt distinkte strømningsfelt. (A) PIV-strømningsfelt for den første (t i) og siste (tf) rammen av tre representative tidsserier, som viser de forskjellige dynamiske klassene som sammenstiller vist i figur 6 viser. PIV-strømningsfelt for mikrotubuli (øverst) og aktin (nederst) for eksempelvideoer i klasse 1 (øverst, lilla), klasse 2 (midtre, oransje) og klasse 3 (nederst, magenta), med pilfarger som tilsvarer den universelle hastighetsskalaen nederst, og gråtonefargekartet som viser den romlige hastighetsfordelingen, normalisert separat for hvert flytfelt i henhold til skalaen vist nederst. Skala barer er alle 50 μM. (B) Vinkelfordelinger av hastighetsvektorer fra A (i enheter av radianer) med oppførte innledende og endelige standardavvik σ i og σf. (C) Tidsmessige fargekart for videoene analysert i A og B viser bilde-til-ramme-posisjonen til hver piksel i forhold til utgangspunktet. Klasse 1-kart viser småskala tilfeldig bevegelse; klasse 2-kart viser rask ensrettet bevegelse med minimal romlig eller tidsmessig variasjon; Klasse 3-kart viser trekk fra både klasse 1 og 2. Dette tallet er endret fra referanse50. Vennligst klikk her for å se en større versjon av denne figuren.
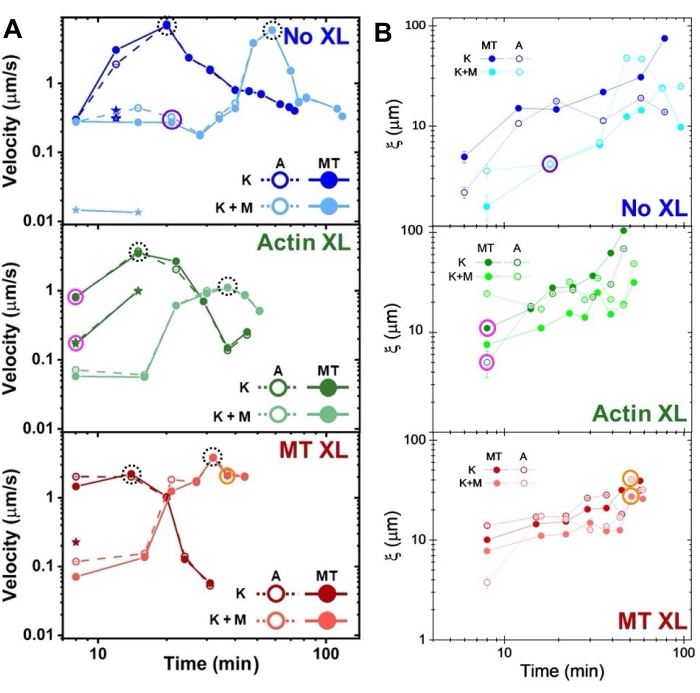
Figur 8. DDM og SIA måler den tidsvarierende dynamikken og strukturen til tomotoriske aktin-mikrotubulikompositter. (A) Hastigheter for kompositter beskrevet i figur 6 og figur 7, målt via DDM, viser akselerasjon og retardasjon av kompositter, programmert ved tverrbinding og myosinaktivitet. Hastigheter av mikrotubuli (MT, lukkede sirkler) og aktin (A, åpne sirkler) er plottet som en funksjon av aktivitetstid i kompositter uten tverrbinding (topp, blå), aktin tverrbinding (midt, grønn), mikrotubuli tverrbinding (bunn, rød), uten myosin (K, mørkere nyanser), og med myosin (K + M, lysere nyanser). For klasse 3-tilfeller, som har to hastigheter, indikeres den langsommere hastigheten av en stjerne. Datapunkter omsluttet av stiplede svarte sirkler tilsvarer maksimal hastighet vmaksfor hver formulering. Feilfelt (de fleste for små til å se) er standardfeilen over strømlovstilpasningene til den tilsvarende τ(q). (B) Strukturelle korrelasjonslengder ξ, bestemt via SIA, versus aktivitetstid, for samme sett med tidsserier evaluert i A. Hvert datapunkt er et gjennomsnitt av korrelasjonslengdene som er bestemt for den første og siste rammen i den tilsvarende tidsserien. Generelt øker ξ i tid for både aktin og mikrotubuli i alle komposittsystemer, og kompositter drevet utelukkende av kinesin har større korrelasjonslengder enn de der myosin også er tilstede. Datapunkter i A og B som tilsvarer de tre tidsseriene som er analysert i figur 7 , er sirklet i tilsvarende klassefarge (1 = lilla, 2 = oransje, 3 = magenta). Dette tallet er endret fra referanse50. Vennligst klikk her for å se en større versjon av denne figuren.
Et viktig fremskritt for det rekonstituerte systemet beskrevet ovenfor er dets modularitet og tunabilitet, slik at brukerne oppfordres til å endre konsentrasjonene av proteiner, motorer, tverrbindinger, etc. for å passe til de ønskede resultatene, enten det er å etterligne en bestemt cellulær prosess eller konstruere et materiale med spesifikk funksjonalitet eller mekaniske egenskaper. Begrensninger på konsentrasjonsområdet for aktin og tubulin er satt til nedre grense av den kritiske konsentrasjonen som trengs for å polymerisere aktin (~ 0,2 μM) 57,58,59 og tubulin (~ 3 - 4 μM) 60, og ved øvre grense ved overgang til nematisk justering av aktinfilamenter (~ 90 μM) 61,62 eller mikrotubuli (~ 35 μM) 63 . Aktinmonomerer og tubulindimerer bør polymeriseres til filamenter sammen, i stedet for å blandes sammen etter polymerisering, for å sikre at de danner homogent interpenetrerende perkolerte nettverk som synergistisk støtter hverandre. Den nye dynamikken som komposittene viser, er avhengig av dette samspillet. Selv om det generelt er viktig å følge alle trinnene som beskrevet i protokollen for å kunne reprodusere resultatene som vises, er noen trinn mer krevende, mens andre har plass til å endre og justere for å passe til spesifikke behov og tilgjengelige ressurser.
For eksempel er et viktig skritt for å sikre reproduserbare resultater riktig forberedelse og lagring av reagensene i henhold til retningslinjene gitt i materialtabellen. Cytoskeletale proteiner (aktin, tubulin, myosin, kinesin) er labile og skal aliquoteres, flashfryses med flytende nitrogen og lagres ved -80 °C i engangsaliquoter. Når aliquotene er fjernet fra -80 °C, skal de oppbevares på is. Cytoskeletale proteiner beholder ikke pålitelig funksjon etter ytterligere fryse-tine sykluser.
Mikrotubuli er mer følsomme for depolymerisering og denaturering enn aktin. Når tubulin er fjernet fra -80 °C, bør den oppbevares på is før polymerisasjon og brukes innen 12 timer. Når polymerisert, bør mikrotubuli holdes ved romtemperatur. Det er også viktig å stabilisere mikrotubuli med taxol for å forhindre depolymerisering. Phalloidin-stabilisering av aktinfilamenter er også viktig for å undertrykke ATP-konsumerende aktin-tredemølle som konkurrerer med myosin- og kinesinaktivitet.
Ultracentrifugation av myosin motorer er et annet kritisk skritt, da det fjerner inaktive myosin døde hoder. Fjerning av enzymatisk inaktive monomerer resulterer i passiv kryssbinding av aktinnettverket og tap av aktivitet. For å forlenge ATPase-aktiviteten til motorer, kan et ATP-regenereringssystem som kreatinfosfat og kreatinfosfokinase64 inkorporeres.
Til slutt krever opprettholdelse av komposittaktivitet inhibering av adsorpsjon av filamenter og motorer til prøvekammerets vegger, noe som kan oppnås ved passivering av mikroskopdekslene og lysbildene. Motorproteiner er spesielt utsatt for adsorpsjon, noe som resulterer i at kompositten blir trukket til overflaten av prøvekammeret, beveger seg ut av synsfeltet, kollapser til 2D og ikke lenger gjennomgår aktivitet. Silanisering av deksler og lysbilder er en effektiv måte å passivere overflatene og forhindre adsorpsjon (se trinn 1). En alternativ passiveringsmetode som brukes effektivt i in vitro cytoskeletteksperimenter, er å belegge overflaten med et lipid-dobbeltlag, lik cellemembranen18. Denne metoden er fordelaktig hvis man ønsker å binde proteiner til overflaten eller introdusere andre spesifikke protein-overflate-interaksjoner, fordi dobbeltlaget kan funksjonaliseres. For optiske pinsetteksperimenter er passivering av mikrosfærene også kritisk, og kan oppnås ved å belegge karboksylerte mikrosfærer med BSA eller PEG via karbodimidtverrbindingskjemi48.
Det er noen aspekter av de presenterte protokollene som forskere kan vurdere å endre for å passe deres behov. For det første kan forskere velge å erstatte ikke-innfødte biotin-NA-tverrbindinger med biologiske tverrbindinger, for eksempel alfa-aktinin eller MAP65 som tverrbinder aktin og mikrotubuli, henholdsvis 28,65,66. Bruken av ikke-innfødte tverrbindinger i komposittene som er beskrevet her, er motivert av deres forbedrede reproduserbarhet, stabilitet og tunbarhet sammenlignet med innfødte tverrbindinger. På grunn av den sterke biotin-NA-bindingen kan tverrbindinger antas å være permanente, i stedet for de fleste innfødte tverrbindinger som forbigående binder seg til omfattende omsetningshastigheter. Dynamikken i forbigående tverrbinding kompliserer analysen av bidragene fra tverrbindinger og motorer til dynamikken. Videre kan biotin-NA-linkere allsidig brukes til å kryssbinde både aktin og mikrotubuli, samt tverrbindingsaktin til mikrotubuli. På denne måten kan en entydig sammenligning mellom tverrbindingsmotiver gjøres, slik at alle andre variabler (f.eks. Kryssbindingsstørrelse, bindingsaffinitet, støkiometri, etc.) er faste. Endelig er reagensene som trengs for å innlemme biotin-NA-linkere, allment tilgjengelige, godt karakteriserte og ofte brukt i mange biofysikklaboratorier. En av de viktigste styrkene til in vitro-plattformen som er beskrevet her, er imidlertid dens modularitet, så forskere bør kunne sømløst erstatte biotin-NA-linkere med innfødte linkere hvis de velger.
For det andre, i den nåværende protokollen, polymeriseres aktinmonomerer og tubulindimerer til filamenter sammen i et sentrifugerrør før de tilsettes prøvekammeret. Strømning av løsningen av sammenfiltrede filamentøse proteiner inn i prøvekammeret kan forårsake strømningsjustering, spesielt av mikrotubuli, som bryter ønsket isotropi og homogenitet av komposittene. Faktisk var et stort fremskritt i tidligere arbeid med steady-state actin-mikrotubulikompositter evnen til å kopolymerisere aktin og mikrotubuli in situ (i prøvekammeret) for å sikre dannelse av isotrope interpenetrerende nettverk av aktin og mikrotubuli15,16,27. Å utvide denne tilnærmingen til aktive kompositter vil imidlertid kreve å legge motorene til prøven før aktin- og tubulinpolymerisering og få hele prøven til å inkubere sammen ved 37 ° C før eksperimenter. Tester av denne variasjonen til protokollen har resultert i redusert aktinpolymerisasjon og ingen merkbar motoraktivitet, sannsynligvis på grunn av konkurrerende ATPase-aktivitet og den langvarige 37 °C-inkubasjonen av motorene. Heldigvis er det ingen merkbar flytjustering av kompositter når man følger gjeldende protokoller, som det fremgår av figur 2, figur 3 og figur 6. Likevel oppfordres forskere til å designe protokoller som tillater in situ dannelse av aktive kompositter.
Et annet poeng å vurdere er fluorescensmerkingsordningen, som innebærer sparsomt merking av alle aktinfilamenter og mikrotubuli i nettverket. Denne merkingsmetoden ble optimalisert for å direkte visualisere strukturen i nettverket i stedet for å utlede struktur og dynamikk via sporfilamenter eller mikrosfærer. Avveiningen er imidlertid at individuelle filamenter ikke er sterkt merket og løsbare. En tilnærming som forskere kan ta for å både løse enkeltfilamenter og visualisere nettverksstruktur, er å dope i forhåndsformede filamenter merket med en annen fluorofor, slik at både det omkringliggende nettverket og individuelle filamenter kan avbildes samtidig. Men når du bruker mer enn to fluoroforer og eksitasjons- / utslippskanaler, er blødning mellom kanaler ofte vanskelig å eliminere, så det må utvises forsiktighet ved valg av fluoroforer, filtre og laserintensiteter.
En relatert begrensning er manglende evne til å visualisere myosin- eller kinesinmotorene i komposittene. De fluorescerende merkede aktinmonomerer og tubulindimerer som brukes er kommersielt tilgjengelige, mens visualisering av myosin eller kinesin i kompositter krever intern merking. Forskere oppfordres til å ta neste skritt for å merke motorer, som gjort tidligere18,67, for å kunne utvetydig knytte motoraktivitet og binding til dynamikken og strukturene som våre kompositter viser.
Til slutt er det viktig å merke seg at i gjeldende protokoll er utbruddet og varigheten av kinesinaktiviteten ikke kontrollert. Fordi myosinaktiviteten styres ved hjelp av fotodeaktivering av blebbistatin, som beskrevet ovenfor, for å bygge inn lignende lysaktivering av kinesin, kan man innlemme lysaktivert ATP.
For å bygge opp kompleksiteten i designene som er beskrevet her, for bedre å etterligne cellulære forhold og utvide parameterrommet for dynamisk struktur-funksjon, vil fremtidig arbeid fokusere på å inkorporere mellomliggende filamenter, for eksempel vimentin68,69, samt andre motorer som dynein13,70. Gelsolin vil også bli innlemmet i forskjellige konsentrasjoner for å kontrollere aktinlengde14, samt tau-protein for å kontrollere mikrotubulistivhet.
Oppsummert beskriver de presenterte protokollene hvordan man designer, skaper og karakteriserer dynamikken, strukturen og mekanikken til cytoskelettinspirerte aktive materiesystemer, som inneholder to separate aktive kraftgenererende komponenter som virker på forskjellige substrater i et enkelt system. Denne justerbare og modulære plattformen bringer rekonstitueringsarbeidet et viktig skritt nærmere å etterligne det cellulære cytoskelettet og tilbyr den unike muligheten til å programmere egenskapene over et bredt faserom ved uavhengig å innlemme, fjerne og justere de forskjellige komponentene. Videre er alle komponenter i dette allsidige systemet kommersielt tilgjengelige (se materialtabell), bortsett fra kinesindimerene som er renset i Ross Lab, som beskrevet tidligere50, og tilgjengelig på forespørsel. Endelig er all analysekode fritt tilgjengelig gjennom GitHub49 og er basert på gratis programmeringsspråk og programvare (Python og Fiji). Den gjennomsiktige formidlingen av protokoller for å designe disse systemene vil forhåpentligvis gjøre denne plattformen mer tilgjengelig for en mangfoldig gruppe brukere med ulik kompetanse, bakgrunn, institusjonell tilknytning og forskningsmål.
Forfatterne har ingenting å avsløre
Vi anerkjenner Maya Hendija og Dr. Jonathan Michel for hjelp med dataanalyse, og Dr. Janet Sheung, Dr. Moumita Das og Dr. Michael Rust for nyttige diskusjoner og veiledning. Denne forskningen ble støttet av et William M. Keck Foundation Research Grant og NSF DMREF Award (DMR 2119663) tildelt RMRA og JLR og National Institutes of Health R15 Grants (R15GM123420, 2R15GM123420-02) tildelt RMR-A og RJM.
| Name | Company | Catalog Number | Comments |
| (-)-Blebbistatin Abbreviation used in paper: blebbistatin | Sigma Aldrich | B0560 | Stock Concentration: 200 μM in DMSO Storage: dessicated, in DMSO, -20ºC Stock and Experiment Recipes: dissolve 1 mg of powder to 200 μM in DMSO Storage, Handling, Troubleshooting Notes: limited shelf-life, typically stops functioning reliably after 3-4 months. purchase and prepare new solution every 3 months. |
| 1:20 488-tubulin:tubulin mixture Abbreviation used in paper: 5-488-tubulin | NA | NA | Stock Concentration: 5 mg/ml in PEM Storage: single use aliquots, -80ºC, avoid freeze-thaw cycles Stock and Experiment Recipes: mix tubulin and 488-tubulin at a 20:1 ratio, flash freeze with LN2 Storage, Handling, Troubleshooting Notes: each aliquot can be used for up to 12 hrs stored on ice at 4ºC, protect from light |
| 1:20 R-tubulin:tubulin mixture Abbreviation used in paper: 5-R-tubulin | NA | NA | Stock Concentration: 5 mg/ml in PEM Storage: single use aliquots, -80ºC, avoid freeze-thaw cycles Stock and Experiment Recipes: mix tubulin and rhodamine tubulin at a 20:1 ratio, flash freeze with LN2 Storage, Handling, Troubleshooting Notes: each aliquot can be used for up to 12 hrs stored on ice at 4ºC, protect from light |
| actin (biotin): skeletal muscle Abbreviation used in paper: biotin-actin | Cytoskeleton | AB07 | Stock Concentration: 1 mg/ml in G-buffer Storage: single use aliquots, -80ºC, avoid freeze-thaw cycles Stock and Experiment Recipes: reconstitute to 1 mg/ml in G-buffer, flash freeze with LN2 Storage, Handling, Troubleshooting Notes: (1) immediately prior to use dilute to 0.5 mg/ml in PEM, (2) once removed from -80ºC, store aliquot on ice at 4ºC for up to 1 week |
| actin (rhodamine): rabbit skeletal muscle Abbreviation used in paper: R-actin | Cytoskeleton | AR05 | Stock Concentration: 1.5 mg/ml in G-buffer Storage: single use aliquots, -80ºC, avoid freeze-thaw cycles Stock and Experiment Recipes: reconstitute to 1.5 mg/ml in G-buffer, flash freeze with LN2 Storage, Handling, Troubleshooting Notes: once removed from -80ºC, store aliquot on ice at 4ºC, can be used for up to 1 week |
| adenosine triphosphate Abbreviation used in paper: ATP | Thermo Fisher Scientific | A1048 | Stock Concentration: 100 mM Storage: in solution (pH 7), -20ºC Stock and Experiment Recipes: reconsitute in DI H20, bring pH to 7 with NaOH Storage, Handling, Troubleshooting Notes: routinely check pH and adjust as needed, hydrolyzes over time, replace every ~6-12 months |
| AlexaFluor488 Phalloidin Abbreviation used in paper: 488-phalloidin | Thermo Fisher Scientific | A12379 | Stock Concentration: 100 μM DMSO Storage: protected from light, dessicated, -20ºC Stock and Experiment Recipes: reconstitute to 100 μM with DMSO Storage, Handling, Troubleshooting Notes: immediately prior to use dilute to 20 μM in PEM (1 μL in 4 μL PEM) |
| AlexaFluor488–labeled actin Abbreviation used in paper: 488-actin | Thermo Fisher Scientific | A12373 | Stock Concentration: 1.5 mg/ml in G-buffer Storage: single use aliquots, -80ºC, avoid freeze-thaw cycles Stock and Experiment Recipes: reconstitute to 1.5 mg/ml in G-buffer, flash freeze with LN2 Storage, Handling, Troubleshooting Notes: this item has been discontinued |
| Basic Plasma Cleaner Abbreviation used in paper: plasma cleaner | Harrick Plasma | PDC-32G | |
| Bemis Parafilm M Laboratory Wrapping Film Abbreviation used in paper: transparent film | Thermo Fisher Scientific | 13-374-5 | |
| D-(+)-Glucose Abbreviation used in paper: | Thermo Fisher Scientific | A1682836 | Stock Concentration: 100x Storage: store at stock concentration (100x) or 10x concentration, dessicated, at -20ºC Stock and Experiment Recipes: reconstitute powder to 4.5 mg/ml in DI H20 Storage, Handling, Troubleshooting Notes: final concentration in solution should 45 μg/mL |
| D-Biotin Abbreviation used in paper: biotin | Fisher Scientific | BP232-1 | Stock Concentration: 1.02 mM in PEM Storage: dessicated, 4ºC |
| deionized nanopure water Abbreviation used in paper: DI | |||
| Dimethyldichlorosilane Abbreviation used in paper: silane | Thermo Fisher Scientific | D/3820/PB05 | Stock Concentration: 2% dissolved in Toulene |
| Dithiothreitol Abbreviation used in paper: DTT | Thermo Fisher Scientific | R0861 | Stock Concentration: 1 M in DMSO Storage: dessicated, -20ºC Stock and Experiment Recipes: dilute to 2 mM in PEM immediately before each experiment |
| DMSO Anhydrous Abbreviation used in paper: DMSO | Thermo Fisher Scientific | D12345 | |
| F-Buffer Abbreviation used in paper: F-buffer | NA | NA | Stock Concentration: 10x Storage: dessicated, -20ºC Stock and Experiment Recipes: 10 mM Imidazole (pH 7.0), 50 mM KCl, 1 mM MgCl2, 1 mM EGTA, 0.2 mM ATP |
| G-Buffer Abbreviation used in paper: G-buffer | NA | NA | Stock Concentration: 10x Storage: dessicated, -20ºC Stock and Experiment Recipes: 2.0 mM Tris (pH 8), 0.2 mM ATP, 0.5 mM DTT, 0.1 mM CaCl2. Store at -20°C. |
| glass microscope slide Abbreviation used in paper: slide | Thermo Fisher Scientific | 22-310397 | |
| Glucose oxidase + catalase + β-mercaptoethanol Abbreviation used in paper: GOC | Sigma Aldrich | G2133-250KU, C1345, 63689 | Stock Concentration: 100x Storage: store at stock concentration (100x) or 10x concentration, dessicated, at -20ºC Stock and Experiment Recipes: For 100x: 4.3 mg/ml glucose oxidase, 0.7 mg/ml catalase, 0.5% v/v β-mercaptoethanol in DI H20 Storage, Handling, Troubleshooting Notes: final concentration in solution should be: 0.005% β-mercaptoethanol, 43 μg/mL glucose oxidase, 7 μg/mL catalase |
| glu-GOC oxygen scavenging system Abbreviation used in paper: glu-GOC | NA | NA | Stock Concentration: 100x Storage: prepare fresh each time Stock and Experiment Recipes: mix equal parts Glu and GOC and add at 1/100 final sample volume immediately before imaging Storage, Handling, Troubleshooting Notes: prepare from Glu and GOC immediately before imaging |
| Guanosine triphosphate Abbreviation used in paper: GTP | Thermo Fisher Scientific | R0461 | Stock Concentration: 100 mM Storage: 100 μL aliquots at -20ºC |
| Instant Mix 1-minute epoxy Abbreviation used in paper: epoxy | Loctite | 1366072 | |
| Kinesin-1 401 BIO 6x HIS Abbreviation used in paper: kinesin | Prepared in JL Ross Lab at Syracuse University | NA | Stock Concentration: 8.87 μM in PEM Storage: single use aliquots, -80ºC, avoid freeze-thaw cycles Storage, Handling, Troubleshooting Notes: biotinylated dimers form kinesin clusters, each aliquot can be used for up to 12 hrs stored on ice at 4ºC |
| NeutrAvidin Abbreviation used in paper: NA | Thermo Fisher Scientific | 31000 | Stock Concentration: 5 mg/ml in PEM Storage: dessicated, -20ºC Stock and Experiment Recipes: reconstitute powder to 5 mg/ml in PEM |
| No 1. glass coverslips (24 mm x 24 mm) Abbreviation used in paper: coverslip | Thermo Fisher Scientific | 12-548-CP | |
| Paclitaxel Abbreviation used in paper: Taxol | Thermo Fisher Scientific | P3456 | Stock Concentration: 2 mM in DMSO Storage: protected from light, dessicated, -20ºC Stock and Experiment Recipes: reconstitute to 2 mM with DMSO Storage, Handling, Troubleshooting Notes: immediately prior to use dilute to 200 μM in DMSO (0.4 μL in 3.6 μL DMSO) |
| PEM-100 Abbreviation used in paper: PEM | NA | NA | Stock Concentration: 1x Storage: room temperature (RT) Stock and Experiment Recipes: 100 mM K-PIPES (pH 6.8), 2 mM EGTA, 2 mM MgCl2 Storage, Handling, Troubleshooting Notes: use KOH to adjust pH to 6.8, recheck pH often and adjust accordingly |
| phalloidin Abbreviation used in paper: phalloidin | Thermo Fisher Scientific | P3457 | Stock Concentration: 100 μM in DMSO Storage: protected from light, dessicated, -20ºC, adhere closely to storage/handling conditions Stock and Experiment Recipes: reconstitute to 100 μM with DMSO Storage, Handling, Troubleshooting Notes: susceptible to impurities in its preparation and denaturing, identifiable as large amorphous aggregates of actin in samples |
| porcine brain tubulin Abbreviation used in paper: tubulin | Cytoskeleton | T240 | Stock Concentration: 5 mg/ml in PEM Storage: single use aliquots, -80ºC, avoid freeze-thaw cycles Stock and Experiment Recipes: reconstitute powder to 5 mg/ml in PEM, flash freeze with LN2 Storage, Handling, Troubleshooting Notes: each aliquot can be used for up to 12 hrs stored on ice at 4ºC |
| Potassium Chloride Abbreviation used in paper: KCl | Thermo Fisher Scientific | AM9640G | Stock Concentration: 4 M Storage: RT |
| Rabbit skeletal actin Abbreviation used in paper: actin | Cytoskeleton | AKL99 | Stock Concentration: 2 mg/ml in G-buffer Storage: single use aliquots, -80ºC, avoid freeze-thaw cycles Stock and Experiment Recipes: reconstitute to 2 mg/ml in G-buffer, flash freeze with LN2 Storage, Handling, Troubleshooting Notes: once removed from -80ºC, store aliquot on ice at 4ºC, can be used for up to 1 week |
| Rabbit skeletal myosin II Abbreviation used in paper: myosin | Cytoskeleton | MY02 | Stock Concentration: 10 mg/ml in PEM Storage: single use aliquots, -80ºC, avoid freeze-thaw cycles Stock and Experiment Recipes: reconstitute powder to 10 mg/ml in PEM, flash freeze with LN2 Storage, Handling, Troubleshooting Notes: monomers form minifilaments at low KCl, each aliquot can be used for up to 12 hrs stored on ice at 4ºC |
| Tubulin (biotin): porcine brain Abbreviation used in paper: biotin-tubulin | Cytoskeleton | T333P | Stock Concentration: 5 mg/ml in PEM Storage: single use aliquots, -80ºC, avoid freeze-thaw cycles Stock and Experiment Recipes: reconstitute powder to 5 mg/ml in PEM, flash freeze with LN2 Storage, Handling, Troubleshooting Notes: immediately prior to use dilute to 0.5 mg/ml in PEM |
| Tubulin (fluorescent HiLyte 488): porcine brain Abbreviation used in paper: 488-tubulin | Cytoskeleton | TL488M | Stock Concentration: 5 mg/ml in PEM Storage: single use aliquots, -80ºC, avoid freeze-thaw cycles Stock and Experiment Recipes: reconstitute powder to 5 mg/ml in PEM, flash freeze with LN2 Storage, Handling, Troubleshooting Notes: each aliquot can be used for up to 12 hrs stored on ice at 4ºC, protect from light |
| tubulin (rhodamine): porcine brain Abbreviation used in paper: R-tubulin | Cytoskeleton | TL590M | Stock Concentration: 5 mg/ml in PEM Storage: single use aliquots, -80ºC, avoid freeze-thaw cycles Stock and Experiment Recipes: reconstitute powder to 5 mg/ml in PEM, flash freeze with LN2 Storage, Handling, Troubleshooting Notes: each aliquot can be used for up to 12 hrs stored on ice at 4ºC, protect from light |
| Tween 20 Abbreviation used in paper: Tween20 | Thermo Fisher Scientific | J20605.AP | Stock Concentration: 1% v/v in DI H20 Storage: RT |
| ultracentrifuge grade microtubes Abbreviation used in paper: Beckman-Coulter Optima Max XP | Beckman Coultier | 343776 | Storage, Handling, Troubleshooting Notes: 8x34 mm PC |
| UV light curing glue Abbreviation used in paper: UV glue | Pharda | SKG-2869 |
- Fletcher, D. A., Mullins, R. D. Cell mechanics and the cytoskeleton. Nature. 463 (7280), 485-492 (2010).
- Koenderink, G. H., Paluch, E. K. Architecture shapes contractility in actomyosin networks. Current Opinion in Cell Biology. 50, 79-85 (2018).
- Dogterom, M., Koenderink, G. H. Actin-microtubule crosstalk in cell biology. Nature Reviews Molecular Cell Biology. 20 (1), 38-54 (2019).
- Burla, F., Mulla, Y., Vos, B. E., Aufderhorst-Roberts, A., Koenderink, G. H. From mechanical resilience to active material properties in biopolymer networks. Nature Reviews Physics. 1 (4), 249-263 (2019).
- Wen, Q., Janmey, P. A. Polymer physics of the cytoskeleton. Current Opinion in Solid State and Materials Science. 15 (5), 177-182 (2011).
- Xiao, Q., Hu, X., Wei, Z., Tam, K. Y. Cytoskeleton molecular motors: structures and their functions in neuron. International Journal of Biological Sciences. 12 (9), 1083-1092 (2016).
- Ajeti, V. et al. Wound healing coordinates actin architectures to regulate mechanical work. Nature Physics. 15 (7), 696-705 (2019).
- Jung, W. et al. Dynamic motions of molecular motors in the actin cytoskeleton. Cytoskeleton. 76 (11-12), 517-531 (2019).
- Pollard, T. D., O'Shaughnessy, B. Molecular mechanism of cytokinesis. Annual Review of Biochemistry. 88 (1), 661-689 (2019).
- Huber, F., Boire, A., López, M. P., Koenderink, G. H. Cytoskeletal crosstalk: when three different personalities team up. Current Opinion in Cell Biology. 32, 39-47 (2015).
- Rivero, F. et al. The role of the cortical cytoskeleton: F-actin crosslinking proteins protect against osmotic stress, ensure cell size, cell shape and motility, and contribute to phagocytosis and development. Journal of Cell Science. 109 (11), 2679-2691 (1996).
- Duclos, G. et al. Topological structure and dynamics of three-dimensional active nematics. Science. 367 (6482), 1120-1124 (2020).
- Baclayon, M. et al. Optical tweezers-based measurements of forces and dynamics at microtubule ends. Optical Tweezers. 1486, 411-435 (2017).
- Gurmessa, B., Fitzpatrick, R., Falzone, T. T., Robertson-Anderson, R. M. Entanglement density tunes microscale nonlinear response of entangled actin. Macromolecules. 49 (10), 3948-3955 (2016).
- Francis, M. L. et al. Non-monotonic dependence of stiffness on actin crosslinking in cytoskeleton composites. Soft Matter. 15 (44), 9056-9065 (2019).
- Ricketts, S. N. et al. Varying crosslinking motifs drive the mesoscale mechanics of actin-microtubule composites. Scientific Reports. 9 (1), 12831 (2019).
- Lee, G. et al. Active cytoskeletal composites display emergent tunable contractility and restructuring. Soft Matter. 17 (47), 10765-10776 (2021).
- Murrell, M. P., Gardel, M. L. F-actin buckling coordinates contractility and severing in a biomimetic actomyosin cortex. Proceedings of the National Academy of Sciences. 109 (51), 20820-20825 (2012).
- Soares e Silva, M. et al. Active multistage coarsening of actin networks driven by myosin motors. Proceedings of the National Academy of Sciences. 108 (23), 9408-9413 (2011).
- Sonn-Segev, A., Bernheim-Groswasser, A., Roichman, Y. Dynamics in steady state in vitro acto-myosin networks. Journal of Physics: Condensed Matter. 29 (16), 163002 (2017).
- Ideses, Y., Sonn-Segev, A., Roichman, Y., Bernheim-Groswasser, A. Myosin II does it all: assembly, remodeling, and disassembly of actin networks are governed by myosin II activity. Soft Matter. 9 (29), 7127 (2013).
- Fürthauer, S. et al. Self-straining of actively crosslinked microtubule networks. Nature Physics. 15 (12), 1295-1300 (2019).
- Lemma, L. M. et al. Multiscale microtubule dynamics in active nematics. Physical Review Letters. 127 (14), 148001 (2021).
- Fan, Y., Wu, K.-T., Aghvami, S. A., Fraden, S., Breuer, K. S. Effects of confinement on the dynamics and correlation scales in kinesin-microtubule active fluids. Physical Review E. 104 (3), 034601 (2021).
- Triclin, S. et al. Self-repair protects microtubules from destruction by molecular motors. Nature Materials. 20 (6), 883-891 (2021).
- Lee, G. et al. Myosin-driven actin-microtubule networks exhibit self-organized contractile dynamics. Science Advances. 7 (6), eabe4334 (2021).
- Ricketts, S. N., Ross, J. L., Robertson-Anderson, R. M. Co-entangled actin-microtubule composites exhibit tunable stiffness and power-law stress relaxation. Biophysical Journal. 115 (6), 1055-1067 (2018).
- Bendix, P. M. et al. A quantitative analysis of contractility in active cytoskeletal protein networks. Biophysical Journal. 94 (8), 3126-3136 (2008).
- Linsmeier, I. et al. Disordered actomyosin networks are sufficient to produce cooperative and telescopic contractility. Nature Communications. 7 (1), 12615 (2016).
- Stam, S. et al. Filament rigidity and connectivity tune the deformation modes of active biopolymer networks. Proceedings of the National Academy of Sciences. 114 (47), E10037-E10045 (2017).
- Yadav, V. et al. Filament nucleation tunes mechanical memory in active polymer networks. Advanced Functional Materials. 29 (49), 1905243 (2019).
- Ennomani, H. et al. Architecture and connectivity govern actin network contractility. Current Biology. 26 (5), 616-626 (2016).
- Alvarado, J., Sheinman, M., Sharma, A., MacKintosh, F. C., Koenderink, G. H. Molecular motors robustly drive active gels to a critically connected state. Nature Physics. 9 (9), 591-597 (2013).
- Alvarado, J., Cipelletti, L., Koenderink, G. H. Uncovering the dynamic precursors to motor-driven contraction of active gels. Soft Matter. 15 (42), 8552-8565 (2019).
- Jung, W., Murrell, M. P., Kim, T. F-actin cross-linking enhances the stability of force generation in disordered actomyosin networks. Computational Particle Mechanics. 2 (4), 317-327 (2015).
- Lenz, M., Thoresen, T., Gardel, M. L., Dinner, A. R. Contractile units in disordered actomyosin bundles arise from f-actin buckling. Physical Review Letters. 108 (23), 238107 (2012).
- Memarian, F.L. et al. Active nematic order and dynamic lane formation of microtubules driven by membrane-bound diffusing motors. Proceedings of the National Academy of Sciences. 118 (52), e2117107118 (2021).
- Needleman, D., Dogic, Z. Active matter at the interface between materials science and cell biology. Nature Reviews Materials. 2 (9), 17048 (2017).
- Foster, P. J., Fürthauer, S., Shelley, M. J., Needleman, D. J. Active contraction of microtubule networks. eLife. 4, e10837 (2015).
- Thijssen, K. et al. Submersed micropatterned structures control active nematic flow, topology, and concentration. Proceedings of the National Academy of Sciences. 118 (38), e2106038118 (2021).
- Sanchez, T., Chen, D. T. N., DeCamp, S. J., Heymann, M., Dogic, Z. Spontaneous motion in hierarchically assembled active matter. Nature. 491 (7424), 431-434 (2012).
- Colen, J. et al. Machine learning active-nematic hydrodynamics. Proceedings of the National Academy of Sciences. 118 (10), e2016708118 (2021).
- Mitchell, K. A., Tan, A. J., Arteaga, J., Hirst, L. S. Fractal generation in a two-dimensional active-nematic fluid. Chaos: An Interdisciplinary Journal of Nonlinear Science. 31 (7), 073125 (2021).
- Pandolfi, R. J., Edwards, L., Johnston, D., Becich, P., Hirst, L. S. Designing highly tunable semiflexible filament networks. Physical Review E. 89 (6), 062602 (2014).
- Tan, A. J. et al. Topological chaos in active nematics. Nature Physics. 15 (10), 1033-1039 (2019).
- Roostalu, J., Rickman, J., Thomas, C., Nédélec, F., Surrey, T. Determinants of polar versus nematic organization in networks of dynamic microtubules and mitotic motors. Cell. 175 (3), 796-808.e14 (2018).
- Ndlec, F. J., Surrey, T., Maggs, A. C., Leibler, S. Self-organization of microtubules and motors. Nature. 389 (6648), 305-308 (1997).
- Sheung, J. Y. et al. Motor-driven restructuring of cytoskeleton composites leads to tunable time-varying elasticity. ACS Macro Letters. 10 (9), 1151-1158 (2021).
- McGorty, R. PyDDM v0.2.0. Zenodo. (2022).
- Achiriloaie, D. H. et al. Kinesin and myosin motors compete to drive rich multi-phase dynamics in programmable cytoskeletal composites. arXiv. (2021).
- Wulstein, D. M., Regan, K. E., Garamella, J., McGorty, R. J., Robertson-Anderson, R. M. Topology-dependent anomalous dynamics of ring and linear DNA are sensitive to cytoskeleton crosslinking. Science Advances. 5 (12), eaay5912 (2019).
- McGorty, R. Image-Correlation. at <https://github.com/rmcgorty/Image-Correlation>. (2020).
- Robertson, C. Theory and practical recommendations for autocorrelation-based image correlation spectroscopy. Journal of Biomedical Optics. 17 (8), 080801 (2012).
- McGorty, R. Differential Dynamic Microscopy - Python. at <https://github.com/rmcgorty/Differential-Dynamic-Microscopy---Python>. (2021).
- Cerbino, R., Trappe, V. Differential dynamic microscopy: probing wave vector dependent dynamics with a microscope. Physical Review Letters. 100 (18), 188102 (2008).
- Robertson-Anderson, R. M. Optical tweezers microrheology: from the basics to advanced techniques and applications. ACS Macro Letters. 7 (8), 968-975 (2018).
- Pollard, T. D. Polymerization of ADP-actin. Journal of Cell Biology. 99 (3), 769-777 (1984).
- Coué, M., Brenner, S. L., Spector, I., Korn, E. D. Inhibition of actin polymerization by latrunculin A. FEBS Letters. 213 (2), 316-318 (1987).
- Pollard, T. D. Actin and actin-binding proteins. Cold Spring Harbor Perspectives in Biology. 8 (8), a018226 (2016).
- Kumar, N. Taxol-induced polymerization of purified tubulin. Mechanism of action. Journal of Biological Chemistry. 256 (20), 10435-10441 (1981).
- Käs, J. et al. F-actin, a model polymer for semiflexible chains in dilute, semidilute, and liquid crystalline solutions. Biophysical Journal. 70 (2), 609-625 (1996).
- Viamontes, J., Narayanan, S., Sandy, A. R., Tang, J. X. Orientational order parameter of the nematic liquid crystalline phase of F -actin. Physical Review E. 73 (6), 061901 (2006).
- Hitt, A. L., Cross, A. R., Williams, R. C. Microtubule solutions display nematic liquid crystalline structure. Journal of Biological Chemistry. 265 (3), 1639-1647 (1990).
- Andexer, J. N., Richter, M. Emerging enzymes for ATP regeneration in biocatalytic processes. ChemBioChem. 16 (3), 380-386 (2015).
- Farhadi, L. et al. Actin and microtubule crosslinkers tune mobility and control co-localization in a composite cytoskeletal network. Soft Matter. 16 (31), 7191-7201 (2020).
- Falzone, T. T., Lenz, M., Kovar, D. R., Gardel, M. L. Assembly kinetics determine the architecture of α-actinin crosslinked F-actin networks. Nature Communications. 3 (1), 861 (2012).
- Thoresen, T., Lenz, M., Gardel, M. L. Reconstitution of contractile actomyosin bundles. Biophysical Journal. 100 (11), 2698-2705 (2011).
- Sanghvi-Shah, R., Weber, G. F. Intermediate filaments at the junction of mechanotransduction, migration, and development. Frontiers in Cell and Developmental Biology. 5, 81 (2017).
- Shen, Y. et al. Effects of vimentin intermediate filaments on the structure and dynamics of in vitro multicomponent interpenetrating cytoskeletal networks. Physical Review Letters. 127 (10), 108101 (2021).
- Laan, L., Roth, S., Dogterom, M. End-on microtubule-dynein interactions and pulling-based positioning of microtubule organizing centers. Cell Cycle. 11 (20), 3750-3757 (2012).
Erratum
Erratum: Reconstituting and Characterizing Actin-Microtubule Composites with Tunable Motor-Driven Dynamics and MechanicsAn erratum was issued for: Reconstituting and Characterizing Actin-Microtubule Composites with Tunable Motor-Driven Dynamics and Mechanics. The Authors section was updated.
Mehrzad Sasanpour1
Daisy H. Achiriloaie1,2
Gloria Lee1
Gregor Leech1
Christopher Currie1
K. Alice Lindsay3
Jennifer L. Ross3
Ryan J. McGorty1
Rae M. Robertson-Anderson1
1Department of Physics and Biophysics, University of San Diego
2W. M. Keck Science Department, Scripps College, Pitzer College, and Claremont McKenna College
3Department of Physics, Syracuse University
to:
Mehrzad Sasanpour1
Daisy H. Achiriloaie1,2
Gloria Lee1
Gregor Leech1
Maya Hendija1
K. Alice Lindsay3
Jennifer L. Ross3
Ryan J. McGorty1
Rae M. Robertson-Anderson1
1Department of Physics and Biophysics, University of San Diego
2W. M. Keck Science Department, Scripps College, Pitzer College, and Claremont McKenna College
3Department of Physics, Syracuse University
ABOUT JoVE
Copyright © 2024 MyJoVE Corporation. All rights reserved