
Summary
Abstract
Introduction
Protocol
Representative Results
Discussion
Acknowledgements
Materials
References
Biology
Восстановление и характеристика актин-микротрубочных композитов с перестраиваемой динамикой и механикой с приводом от двигателя
ERRATUM NOTICE
Important: There has been an erratum issued for this article. Read more …В данной работе представлены протоколы для проектирования и характеристики перестраиваемых трехмерных композитных сетей совместно запутанных актиновых нитей и микротрубочек. Композиты подвергаются активной реструктуризации и баллистическому движению, приводимому в действие двигателями миозина II и кинезина, и настраиваются относительными концентрациями актина, микротрубочек, моторных белков и пассивных сшивающих факторов.
Композитный цитоскелет, включающий взаимодействующие сети полугибких актиновых нитей и жестких микротрубочек, реструктурируется и генерирует силы, используя моторные белки, такие как миозин II и кинезин, для управления ключевыми процессами, такими как миграция, цитокинез, адгезия и механозондирование. В то время как взаимодействия актин-микротрубочки являются ключом к универсальности и адаптивности цитоскелета, понимание их взаимодействия с активностью миозина и кинезина все еще зарождается. В этой работе описывается, как спроектировать перестраиваемые трехмерные композитные сети из совместно запутанных актиновых нитей и микротрубочек, которые подвергаются активной реструктуризации и баллистическому движению, приводимые в действие двигателями миозина II и кинезина, и настраиваются относительными концентрациями актина, микротрубочек, моторных белков и пассивных сшивающих устройств. Также подробно описаны протоколы флуоресцентной маркировки микротрубочек и актиновых нитей для наиболее эффективной визуализации композитной реструктуризации и движения с использованием многоспектральной конфокальной визуализации. Наконец, представлены результаты методов анализа данных, которые могут быть использованы для количественной характеристики неравновесной структуры, динамики и механики. Воссоздание и исследование этой перестраиваемой биомиметической платформы дает ценную информацию о том, как связанная двигательная активность, композитная механика и динамика нитей могут привести к множеству клеточных процессов от митоза до поляризации и механо-ощущения.
Цитоскелет представляет собой динамическую композитную сеть взаимодействующих биополимеров, которая обеспечивает структурную и механическую поддержку клеток. Связанные молекулярные двигатели и связывающие белки реструктурируют и адаптируют цитоскелет, чтобы позволить клеткам расти, изменять форму, жесткость, двигаться и даже самовосстанавливаться, обеспечивая множество клеточных процессов, начиная от миграции и деления до механозондирования 1,2. Помимо своего значения в клеточной биофизике, цитоскелет также является квинтэссенцией активного вещества с потенциальными применениями материалов, начиная от заживления ран и доставки лекарств до фильтрации и мягкой робототехники 1,3,4,5,6,7,8,9.
Двумя ключевыми характеристиками, которые наделяют цитоскелет уникальным структурным и механическим разнообразием и многофункциональностью, являются: 1) его композитная природа, включающая множественные взаимодействующие белковые нити, такие как полугибкие актиновые нити и жесткие микротрубочки, а также связанные с ними связывающие и сшивающие белки 3,5,10; и 2) его способность непрерывно реструктурироваться, перемещаться, грубеть и выполнять работу с помощью энергоемких двигателей, таких как миозины и кинезины, толкая и вытягивая нитевидные белки 1,7,11,12,13. Хотя эта элегантная сложность позволяет цитоскелету опосредуть такие разнообразные процессы, как подвижность клеток, цитокинез и заживление ран 3,6,7,11, она препятствует способности исследователей воспроизводить сигнатурные характеристики цитоскелета in vivo в восстановленных системах in vitro.
Текущие усилия по восстановлению границ сосредоточены на композитах запутанных и сшитых актиновых нитей и микротрубочек 3,10,14,15,16,17, силообразующих актимозиновых сетях 2,8,18,19,20,21 и активных нематицах, управляемых кинезин-микротрубочками. взаимодействия 22,23,24,25,26. Было показано, что стационарные актин-микротрубочки демонстрируют эмерджентные механические свойства 15,16,27, такие как повышенная подвижность нити накаливания и повышенная жесткость по сравнению с однокомпонентными системами 27. Исследования систем актионатомизина in vitro сообщили о широком диапазоне структурных и динамических свойств, которые зависят от концентраций актина, миозина и сшивающихвеществ 28,29,30,31. Например, при достаточном сшивании сети актомиозина подвергаются крупномасштабному сокращению иогрублению 2,28,30,32,33,34,35,36, тогда как без сшивок сети демонстрируют быстрый, дестабилизирующий поток и разрыв 19,29 . Сообщалось, что восстановленные активные нематики на основе микротрубочек, которые используют кластеры кинезиновых двигателей для сшивания и вытягивания пучков микротрубочек, демонстрируют длительные турбулентные потоки, расширение, изгиб, разрыв и исцеление 12,22,23,24,25,37,38,39,40,41, 42,43,44,45,46,47.
Совсем недавно было показано, что композиты актин-микротрубочек, приводимые в действие мини-нитями миозина II, приводят к более упорядоченному сокращению и целостности сети по сравнению с неупорядоченным потоком и разрывом сети, которые сети актомиозина без сшивок демонстрируют 17,26,48. Кроме того, сочетание прочности композита и генерации силы оптимизируется, когда актин и микротрубочки присутствуют в сопоставимых концентрациях. Ключевые возникающие особенности в этой области пространства рецептур включают повышенную механическую прочность26, скоординированное движение актина и микротрубочек26, устойчивое устойчивое сжатие и мезомасштабную реструктуризацию17.
Здесь описаны протоколы проектирования и настройки совместно запутанных и сшитых композитов микротрубочек и актиновых нитей, которые выталкиваются из равновесия мини-нитями миозина II и кинезиновыми кластерами, действующими на актиновые нити и микротрубочки соответственно (рисунок 1). Динамика, структура и механика этого класса композитов могут быть настроены относительными концентрациями нитей, двигателей и сшивателей, чтобы продемонстрировать богатое фазовое пространство адвективного и турбулентного потока, изотропного сжатия, ускорения, замедления, размешивания, жесткости, релаксации и разрыва. Основное внимание в данной работе уделяется подготовке и настройке этого класса активных цитоскелетных композитов. Однако, чтобы помочь исследователям в бенчмаркинге и характеристике описанных активных композитов, эффективные методы визуализации с использованием мультиспектральной конфокальной микроскопии также детализированы. Наконец, представлены результаты ключевых методов вычислительного анализа, которые могут быть использованы для измерения динамики, структуры и механики композитов. Исследователям рекомендуется принять эти методы, которые включают дифференциальную динамическую микроскопию (DDM), автокорреляцию пространственных изображений (SIA) и велоциметрию изображения частиц (PIV), поскольку они были оптимизированы для характеристики сложной динамики и структурного разнообразия композитов 17,26,49.
Шаги, описанные ниже, сосредоточены на подготовке композитов и их визуализации с помощью конфокальной микроскопии. Протоколы, описывающие анализ данных после сбора и измерения оптического пинцета, можно найти в предыдущих работах 17,26,48,50 и предоставить по запросу. Все материалы перечислены в Таблице материалов.
1. Подготовьте силанизированные крышки и слайды микроскопа для предотвращения адсорбции белков на поверхности камеры
ПРИМЕЧАНИЕ: Это 2-дневный процесс. Силанизированные слайды могут быть приготовлены за 1 месяц до использования.
- Поместите крышки No 1 (24 мм х 24 мм) и слайды микроскопа (1 дюйм х 3 дюйма) в специальную стойку, которая поместится в плазменный очиститель. Поместите стойку в плазменный очиститель и работайте в течение 20 минут.
- Переложите крышки и слайды на новую стойку, предназначенную только для использования с силаном, и поместите стойку в стеклянную емкость для очистки стекол, как описано ниже.
- Погружайте крышки и слайды в 100% ацетон в течение 1 ч. Погружайте крышки и слайды в 100% этанол в течение 10 минут.
- Погружайте крышки и горки в деионизированную воду (DI) на 5 минут. Повторите шаги очистки еще два раза.
- Погружайте крышки и слайды в свежеприготовленные 0,1 М КОН в течение 15 мин. Погружайте крышки и слайды в свежий DI на 5 минут. Повторите этот шаг еще два раза.
- Сухие на воздухе крышки и горки в течение 10 мин. Обработайте очищенные крышки и слайды силаном для получения гидрофобных поверхностей, как описано ниже.
ПРИМЕЧАНИЕ: Выполните следующие действия в вытяжке.- Погружайте высушенные крышки и слайды в 2% силан (растворенный в толуоле) в течение 5 мин. Используйте воронку, чтобы налить силан обратно в назначенную бутылку для повторного использования до пяти раз.
- Погружайте крышки и слайды в 100% этанол на 5 мин. Замените этанол свежим этанолом. Погружайте крышки и слайды на 5 минут.
- Погружайте крышки и слайды в свежий DI на 5 минут. Повторите этап промывки этанола и DI еще два раза, используя свежий этанол и DI каждый раз. Сухие на воздухе крышки и горки в течение 10 мин.
2. Получение активного актин-микротрубочного композита, приводимого в действие мини-нитями миозина
- Удалите неактивный миозин с помощью связывания актиновой нити и выполните вытягивание с помощью ультрацентрифугирования, как описано ниже.
- Полимеризуйте актин в нити. Используя прецизионную микропипетку и стерильные наконечники пипетки, соедините в микроцентрифужной трубке: 1,87 мкл DI, 1,3 мкл 10x G-буфера, 1,3 мкл 10x F-буфера, 1,63 мкл 4 M KCl, 4,53 мкл актина (47,6 мкМ) и 1,08 мкл 100 мкМ фаллоидина.
ПРИМЕЧАНИЕ: Для обеспечения достаточной полимеризации концентрация актина и молярное соотношение актин:фаллоидин должны составлять соответственно 18,4 мкМ и 2:1. - Аккуратно пипетируйте раствор вверх и вниз, чтобы перемешать, а затем положите на лед в темноте в течение ≥1 ч. Прохладная ультрацентрифуга до 4 °C. Удалить аликвоту миозина с -80 °C и положить на лед.
ПРИМЕЧАНИЕ: Завершите этап 2.2 на этом этапе, пока актин полимеризуется. - После ≥1 ч полимеризации актина добавляют к полимеризованному актину 1,3 мкл 10 мМ АТФ и 2 мкл миозина 19 мкМ.
ПРИМЕЧАНИЕ: Молярное соотношение актин:миозин должно быть >5 для обеспечения достаточного удаления неактивных миозиновых двигателей (т.е. мертвых головок). - Аккуратно пипетируйте раствор вверх и вниз, чтобы перемешать. Перенос в ультрацентрифужную трубку.
Центрифуга при 4 °C и 121 968 х г в течение 30 мин.
- Полимеризуйте актин в нити. Используя прецизионную микропипетку и стерильные наконечники пипетки, соедините в микроцентрифужной трубке: 1,87 мкл DI, 1,3 мкл 10x G-буфера, 1,3 мкл 10x F-буфера, 1,63 мкл 4 M KCl, 4,53 мкл актина (47,6 мкМ) и 1,08 мкл 100 мкМ фаллоидина.
- Подготовьте совместно запутанную композитную сеть актиновых нитей и микротрубочек, как описано ниже.
ПРИМЕЧАНИЕ: Начните за 30 минут до отката миозина (шаг 2.1.4).- Установите тепловой блок на 37 °C. Используйте прецизионные наконечники микропипетки и стерильные пипетки, чтобы добавить в микроцентрифужную трубку следующее: 13,9 мкл PEM, 3 мкл 1% Tween20, 1,55 мкл 47,6 мкМ актина, 0,36 мкл 34,8 мкМ R-актина, 0,3 мкл 250 мМ АТФ, 0,87 мкл 100 мкМ фаллоидина, 1,91 мкл 5-488-тубулина, 0,3 мкл 100 мМ ГТФ, и 0,75 мкл 200 мкМ таксола, до общего объема 23 мкл.
ПРИМЕЧАНИЕ: Перечисленные концентрации актина и тубулина относятся к композиту с актином 2,9 мкМ и тубулином 2,9 мкМ. Общая концентрация белка составляет c = cA + cT = 5,8 мкМ, а молярная фракция актина составляет cA/(cA + cT) = ΦA = 0,5. См. шаг 2.5, чтобы настроить эти значения. - Аккуратно пипетируйте раствор вверх и вниз, чтобы перемешать и поместить на тепловой блок с температурой 37 °C, защищенный от света, в течение 1 ч.
- Установите тепловой блок на 37 °C. Используйте прецизионные наконечники микропипетки и стерильные пипетки, чтобы добавить в микроцентрифужную трубку следующее: 13,9 мкл PEM, 3 мкл 1% Tween20, 1,55 мкл 47,6 мкМ актина, 0,36 мкл 34,8 мкМ R-актина, 0,3 мкл 250 мМ АТФ, 0,87 мкл 100 мкМ фаллоидина, 1,91 мкл 5-488-тубулина, 0,3 мкл 100 мМ ГТФ, и 0,75 мкл 200 мкМ таксола, до общего объема 23 мкл.
- Подготовьте камеры для конфокальных экспериментов по визуализации, как описано ниже.
ПРИМЕЧАНИЕ: Выполните шаги 2.1.4 и 2.2.2 в периоды ожидания.- Поместите два силанизированных слайда бок о бок на конфорку (выключено), положите две полоски термопластичной уплотнительной пленки на слайды на расстоянии ~ 3 мм друг от друга и поместите два силанизированных обтекателя поверх термопластичной уплотнительной пленки, чтобы сформировать камеру для образцов.
- Поверните конфорку на низкой установке до тех пор, пока крышки прочно не свяжутся с слайдами с расплавленной термопластичной уплотнительной пленкой (~1-2 мин). Прижмите с равномерным давлением, чтобы обеспечить склеивание при сохранении расстояния ~100 мкм между двумя поверхностями.
- Снимите камеры и выключите конфорку. Этикетировочные камеры с (+) и (-). Камера (+) будет для активного образца (с миозином), а (-) камера будет контрольной (без миозина). Убедитесь, что каждая камера может вместить ≤10 мкл жидкости.
- Подготовьте образцы к изображению, как описано ниже.
ПРИМЕЧАНИЕ: Важно выполнить этот шаг сразу после завершения шагов 2.1 и 2.2.- Осторожно извлеките образец миозина-актина из ультрацентрифуги (этап 2.1.4) и немедленно пипетируйте верхние 7,5 мкл супернатанта и перенесите в новую микроцентрифужную трубку.
- Извлеките образец актин-микротрубочки из теплового блока и осторожно перемешайте 1,5 мкл 10x D-глюкозы, 1,5 мкл 10x GOC и 1,5 мкл 1 мМ блеббистатина. Разделите раствор на две аликвоты по 13,7 мкл и пометьте как (+) и (-).
- Смешайте 1,28 мкл супернатанта со стадии 2.4.1 до (+) аликвоты. Смешать в 1,28 мкл DI с (-) аликвотой. Медленно перетекайте каждый раствор в соответствующую камеру (этап 2.3) посредством капиллярного действия. Будьте осторожны, чтобы не вводить пузырьки воздуха в канал.
- Запечатайте два открытых конца каждого канала быстросохнущим эпоксидным или ультрафиолетовым клеем. Убедитесь, что клей полностью высох, прежде чем помещать его на микроскоп. Изображение сразу, как описано в шаге 3.
ПРИМЕЧАНИЕ: УФ-клей выгоден, потому что он отверждается почти мгновенно при воздействии ультрафиолета. Однако, поскольку блеббистатин чувствителен к ультрафиолетовому излучению, важно только локально освещать клей (по краям камеры образца) с помощью небольшой УФ-палочки, чтобы избежать деактивации блеббистатина.
- Необязательно: изменяйте концентрации белка для настройки динамики и структуры композитов.
ПРИМЕЧАНИЕ: На следующих этапах предлагается внести изменения в вышеуказанные этапы для изменения концентраций актина, микротрубочек и миозина, если это необходимо.- Выполните описанные выше действия, за исключением следующих изменений на этапах 2.2.1 и 2.4.3.
- Для изменения концентраций актина и микротрубочек, тем самым регулируя c и ΦA, увеличивают или уменьшают объем актина, R-актина и 5-488-тубулина используют на стадии 2.2.1, по желанию26. При изменении концентрации актина регулируйте молярные концентрации R-актина и фаллоидина пропорционально, чтобы поддерживать одинаковые молярные соотношения с актином. Отрегулируйте объем PEM таким образом, чтобы конечный объем смеси оставался 23 мкл. Все остальные объемы и концентрации компонентов остаются прежними.
- Чтобы изменить концентрацию миозина, отрегулируйте объем миозина, добавленного к (+) аликвоте на шаге 2.4.3, по желанию. Отрегулируйте громкость DI, добавленную к (-) аликвоте, соответствующим образом. Отрегулируйте объем PEM на шаге 2.2.1, чтобы учесть увеличение или уменьшение объема миозина (+) и DI (-), гарантируя, что конечный объем каждого образца ((+) и (-)) составляет 14,98 мкл.
3. Визуализация и характеристика активных композитов с помощью конфокальной микроскопии
- Для получения изображения композитов actomyosin-microtubule, подготовленных на этапе 2, используйте лазерный сканирующий конфокальный микроскоп (LSCM) или аналогичный микроскоп с масляным иммерсионным объективом 60x 1,4 NA. Для одновременной визуализации актиновых нитей и микротрубочек в отдельных флуоресцентных каналах используют лазер 561 нм с фильтрами возбуждения/излучения 565/591 нм и лазер 488 нм с фильтрами возбуждения/излучения 488/525 нм.
- Поместите камеру образца на микроскоп таким образом, чтобы контрольный канал располагался непосредственно над объективом. Убедитесь, что между объективом и крышкой есть масляный интерфейс.
- Используйте элементы управления каскадом, чтобы сфокусировать управляющий композит, а затем найдите обе поверхности камеры для отбора проб. Переместите положение z в центр камеры отбора проб. Проверьте наличие прозрачных нитевидных сетей, как показано на рисунке 2.
- По-прежнему визуализируя контрольную камеру, регулируйте интенсивность каждого лазера, чтобы обеспечить одновременную визуализацию актиновых нитей и микротрубочек. Поддерживайте самую низкую возможную интенсивность лазера для предотвращения фотоотбеливания (более распространенного в актиновом канале) и кровотечения (обычно из микротрубочек в актиновый канал).
- Чтобы охарактеризовать неактивный контрольный образец, соберите три временных ряда (видео) из 256 x 256 квадратных пикселей (213 мкм x 213 мкм) изображений со скоростью 2,65 кадра в секунду в общей сложности ≥1000 кадров. Соберите каждый временной ряд в другой области камеры образца, разделенной ≥500 мкм. Обеспечьте минимальное обнаруживаемое движение и отсутствие потока или реструктуризации.
- Отключите лазер 488 нм и используйте элементы управления сценой для перемещения в камеру (+).
- Используя лазер 568 нм, визуализируйте микротрубочки в канале (+) для обеспечения надлежащего формирования сети (фиг.2) и идентифицируйте осевой центр камеры образца (который может отличаться от центрального z-положения управляющей камеры).
- Включите лазер 488 нм и повторите шаг 3.5 выше со следующими изменениями. Собирайте временные ряды в течение 45 минут, останавливая захват, когда образец либо перемещается из поля зрения, либо разрывается, либо фотоблокируется. Записывайте 5-10 временных рядов и отслеживайте время, в которое начинается каждый временной ряд относительно начала первого временного ряда.
- Анализируйте данные с помощью DDM, SIA и PIV, как описано на рисунке 3, рисунке 4, рисунке 5, и ранее 17,48,50,51.
ПРИМЕЧАНИЕ: Лазер 488 нм локально активирует активность АТФазы миозина путем деактивации блеббистатина, поэтому его следует включать только в начале сбора данных таким образом, чтобы t = 0 находилось в начале временного ряда. Эти параметры сбора оптимизированы для анализа дифференциальной динамической микроскопии (DDM), как это было сделано ранее26.
4. Получение активных актин-микротрубочных композитов, приводимых в действие кинезиновыми двигателями
ПРИМЕЧАНИЕ: Следующие этапы создают актин-микротрубочки композитов, которые выводятся из равновесия кинезиновыми двигателями или комбинацией кинезина и миозина50.
- Подготовьте кинезиновые и миозиновые моторы, как описано ниже.
- При включении миозина следуйте шагу 2.1.
- Чтобы сформировать кинезиновые моторные кластеры, которые связывают и оказывают силы между парами микротрубочек, используйте микропипетку и стерильные наконечники пипетки, чтобы добавить следующие в стерильную микроцентрифужную трубку 1,5 мл: 1,16 мкл PEM, 2,74 мкл 8,87 мкМ кинезиновые димеры, 7,29 мкл 83,3 мкМ НейтрАвидин, 0,81 мкл 2 мМ DTT. Аккуратно перемешайте, пипетируя раствор вверх и вниз, и инкубируйте защищенный от света (используйте черную микроцентрифужную трубку или заверните в фольгу) в течение 30 мин при 4 °C.
ПРИМЕЧАНИЕ: Молярное отношение кинезиновых димеров к NA составляет 1:25.
- Следуйте шагу 2.3, чтобы подготовить камеры для отбора проб и сделать три камеры вместо двух. Проводят эту стадию во время инкубации кинезина (стадия 4.1.2) и ультрацентрифугирования миозина (стадия 4.1.1).
- Готовят совместно запутанную композитную сеть актиновых нитей и микротрубочек.
- Установите тепловой блок на 37 °C. Используйте микропипетку и стерильные наконечники пипетки, чтобы добавить следующее в стерильную микроцентрифужную трубку 1,5 мл: 3,21 мкл PEM, 4,5 мкл 1% Tween20, 2,18 мкл 47,6 мкм актина, 3,46 мкл 5-R-тубулина, 4,5 мкл 100 мМ АТФ, 4,5 мкл 10 мМ ГТФ, 1,13 мкл 200 мкМ таксола и 1,57 мкл 20 мкМ 488-фаллоидина. Убедитесь, что общий объем составляет 25 мкл.
- Аккуратно пипетируйте раствор вверх и вниз, чтобы перемешать и поместить на термоблок 37 °C, защищенный от света, в течение 1 ч. Извлеките трубку из теплового блока и используйте микропипетку, чтобы аккуратно смешать в 0,84 мкл 100 мкМ фаллоидина. Инкубировать в течение 5-10 мин при комнатной температуре, защищенной от света.
ПРИМЕЧАНИЕ: Добавление фаллоидина на этом этапе, а не на этапе 4.3.1, улучшает флуоресцентную маркировку актиновых нитей, поскольку 488-фаллоидин не должен конкурировать с немаркированным фаллоидином за сайты связывания актина.
- Подготовьте активные композиты для конфокальной визуализации.
- Добавьте 1,13 мкл 200 мкМ блеббистатина, 1,35 мкл 10x Glu и 1,35 мкл 10x GOC в раствор со стадии 4.3.2 и осторожно перемешайте путем пипетки вверх и вниз. Разделите раствор на три аликвоты по 10 мкл и пометьте как (K), (K+M) и (-).
- Смешайте 2,54 мкл миозина со стадии 2,1,4 до аликвоты (K+M). Смешать в 2,54 мкл PEM с (K) и (-) аликвотами.
- Используйте микропипетку и стерильные наконечники пипетки, чтобы добавить 2,5 мкл кинезиновых кластеров от шага 4.1.2 до (K) и (K + M) аликвот. Пипетку вверх и вниз для смешивания. Смешайте в 2,5 мкл PEM до (-), используя ту же технику.
ПРИМЕЧАНИЕ: Перечисленные концентрации актина и тубулина относятся к композиту с актином 2,32 мкМ и тубулином 3,48 мкМ. Общая концентрация белка составляет c = cA + cT = 5,8 мкМ, а молярная фракция актина составляет cA/(cA + cT) = ΦA = 0,4. Концентрации кинезина и миозина составляют 0,35 мкМ и 0,47 мкМ соответственно. Общие рекомендации по корректировке cA, cT, c и ΦA см. на этапе 2.5. - Используя микропипетку, медленно перетекайте каждый раствор в соответствующий канал подготовленных камер образца (этап 4.2) посредством капиллярного действия. Надавливайте очень медленно и осторожно на пипетку, чтобы не вводить пузырьки воздуха в канал.
- Запечатайте два открытых конца каждого канала быстросохнущим эпоксидным или УФ-отверждаемым клеем. Убедитесь, что клей полностью высох, прежде чем помещать его на микроскоп.
ПРИМЕЧАНИЕ: Важно, чтобы этот шаг был выполнен быстро, чтобы свести к минимуму время, в течение которого кинезин действует без контроля. По этой причине рекомендуется эпоксидная смола, которая отверждается за 1 мин (а не за 5 или 10 мин). УФ-отверждаемый клей выгоден в этом отношении, потому что он затвердевает почти мгновенно при воздействии ультрафиолета.
- Изображение подготовлено образцами сразу после шага 3, за исключением следующих важных модификаций. Поскольку кинезин не контролируется световой активацией, он начинает работать сразу после шага 4.4.3, поэтому отметьте это время как t = 0. Чтобы изобразить композит как можно ближе к начальному неактивному состоянию (t = 0), сначала изобразите каналы (K) и (K+M) и запишите время, прошедшее между шагом 4.4.3 и началом сбора данных (шаг 3.8). На практике это время составляет ~5 мин.
5. Включение пассивных сшивателей в активные композиты
ПРИМЕЧАНИЕ: Эти шаги описывают, как использовать биотинилированные субъединицы актина и тубулина и нейтравидин (NA) для пассивного сшивания актина с актином (A-A) или микротрубочек с микротрубочками (M-M) в активных композитах, описанных на этапе 4.
- Готовят сшивающие комплексы А-А или М-М с биотинилированными белками (биотин-актин или биотин-тубулин), NA и биотином в соотношении 2:2:1 биотин-актин/тубулин:биотин:NA. Запустите этот процесс до шага 4 .
- Для сшивки А-А используйте микропипетку и стерильные наконечники пипетки, чтобы добавить 2 мкл биотин-актина 11,6 мкМ, 1,39 мкл 8,33 мкМ NA, 2,27 мкл 1,02 мкМ биотина и 4,34 мкл PEM в микроцентрифужную трубку. Аккуратно перемешайте, пипеткой вверх и вниз.
- Для сшивающих устройств M-M используйте микропипетку и стерильные наконечники пипетки, чтобы добавить 1,86 мкл биотина-тубулина 4,55 мкМ, 1,11 мкл 8,33 мкМ NA, 1,82 мкл биотина 1,02 мкМ и 5,21 мкл PEM в микроцентрифужную трубку. Аккуратно перемешайте, пипеткой вверх и вниз.
- Оберните трубку (трубки) со стадии 5.1.1 и/или 5.1.2 термопластичной уплотнительной пленкой для создания водонепроницаемого уплотнения. Поместите во флотационный плот в ванну с ультразвуковым аппаратом с регулируемой температурой, установленной на 4 °C.
- Ультразвук в течение 90 мин при 4 °C. На практике лучше всего поместить ультразвуковой аппарат в холодное помещение и добавить пакеты со льдом в ванну с ультразвуком для поддержания низкой температуры.
- Для включения сшивающих комплексов в образцы для получения изображений выполните этап 4.3, изменив этап 4.3.1, как описано ниже для сшивки А-А (этап 5.2.1) или сшивки М-М (этап 5.2.2).
- Для сшивки А-А объедините в микроцентрифужной трубке следующее: 1,94 мкл PEM, 4,50 мкл 1% Tween20, 2,18 мкл 47,6 мкм актина, 3,46 мкл 45,5 мкМ 5-R-тубулина, 1,13 мкл сшивающих А-А (стадия 5,1,1), 4,50 мкл 100 мМ АТФ, 4,50 мкл 10 мМ ГТФ, 1,13 мкл 200 мкМ таксола, и 1,57 мкл 20 мкМ 488-фаллоидина. Убедитесь, что общий объем составляет 25 мкл.
- Для сшивания M-M объедините в микроцентрифужной трубке следующее: 1,97 мкл PEM, 4,50 мкл 1% Tween20, 2,18 мкл 47,6 мкМ актина, 3,76 мкл 45,5 мкМ 5-R-тубулина, 1,13 мкл разбавления 1:4 сшивающих M-M (стадия 5,1.2), 4,50 мкл 100 мМ АТФ, 4,50 мкл 10 мМ ГТФ, 1,13 мкл 200 мкМ таксола, и 1,57 мкл 20 мкМ 488-фаллоидина. Убедитесь, что общий объем составляет 25 мкл.
- Следуйте шагам 4.3.2-4.5 с указанием удельных концентраций для молярного соотношения сшитый ликер:актин RA = 0,02 и молярного отношения сшиватель:тубулин RT = 0,005. Эти значения RA и RT приводят к одинаковой длине между сшивающими устройствами вдоль актиновых нитей и микротрубочек (dA
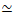 60 нм и dMT 67 нм), рассчитанных с использованием dA = Iмономер/2RA, где Iмономер - длина мономера актина, а dMT = Iкольцо / 26RT, где Iкольцо - длина кольца из 13 тубулинов15, 17.
60 нм и dMT 67 нм), рассчитанных с использованием dA = Iмономер/2RA, где Iмономер - длина мономера актина, а dMT = Iкольцо / 26RT, где Iкольцо - длина кольца из 13 тубулинов15, 17.
Для определения успешного получения активных композитов (Фиг.1), а также для характеристики их динамики и структуры используется лазерный сканирующий флуоресцентный микроскоп с по меньшей мере двумя флуоресцентными каналами для визуализации актиновых нитей и микротрубочек одновременно (Фиг.2 и Фиг.6). Все актиновые нити и микротрубочки в композитах слабо маркируются, а не легируются в индикаторных ярких нитях, как это часто делается в исследованиях in vitro . Этот метод гарантирует, что измеренная динамика и структура являются репрезентативными для самого композита, а не для индикаторов, которые образуются в условиях, отличных от композитов. По этой причине отдельные актиновые нити и микротрубочки обычно не могут быть разрешены, скорее изображения изображают мезомасштабную сетевую структуру (рисунок 2 и рисунок 6).
Этот подход к маркировке был оптимизирован для анализа автокорреляции пространственных изображений (SIA) и дифференциальной динамической микроскопии (DDM), которые исследуют динамику и структуру в обратном пространстве Фурье (рисунок 4, рисунок 5 и рисунок 8)52,53,54,55. Велоциметрия изображений частиц (PIV) также может использоваться для изображения и характеристики динамики и полей потока (рисунок 3 и рисунок 7), но она требует объединения пикселей (более низкое пространственное разрешение) и больших приращений времени задержки (более низкое временное разрешение), чем SIA и DDM, чтобы устранить ошибочные векторы, возникающие из-за шума в плотных изображениях с низким уровнем сигнала. Тем не менее, PIV рекомендуется для качественного изучения полей потока и подтверждения результатов DDM (рисунок 4 и рисунок 8)26,50.
Выборочная характеристика описанных сетей с использованием этих анализов (т.е. DDM, SIA, PIV) предоставляется для того, чтобы помочь исследователям в принятии аналогичных анализов для сравнения и характеристики их образцов. Однако подробные описания этих методов выходят за рамки данной работы. Для подробного описания того, как выполнять DDM на этих и других подобных системах, включая удобный код Python, обратитесь к предыдущим работам 17,26,49,50 и ссылкам там внутри. Для получения подробной информации о том, как выполнять SIA и PIV на системах, описанных здесь, читатель направляется к предыдущим работам17,50.
Необходимо выполнить несколько мер контроля, описанных ниже, чтобы обеспечить функционирование композитов должным образом. Композит без миозина или кинезина должен казаться по существу статическим с минимальными тепловыми колебаниями или дрейфом. Актиновые нити и микротрубочки должны казаться совместно запутанными и однородно распределенными, с минимальным объединением, агрегацией или фазовым разделением актина и микротрубочек по всему полю зрения ~200 мкм х 200 мкм (рисунок 2, крайний слева)17. Следует ожидать аналогичного результата для композитов, которые содержат миозин, но не подвергаются воздействию света 488 нм (для деактивации блеббистатина).
При включении миозина и воздействии света 488 нм композиты подвергаются сокращению, которое в значительной степени изотропно и аналогично для актина и микротрубочек, как видно на микроскопических изображениях, сделанных до и после активности миозина (рисунок 2), а также соответствующих полей потока PIV в течение различного времени во время активности (рисунок 3). Чтобы определить, является ли движение баллистическим, диффузным, субдиффузивным и т.д., характеристическое время декорирования τ(q), определяемое из DDM, оценивается как функция волнового вектора (т.е. обратного пространства). См. как подробно описано ранее 17,26,49. На рисунке 4 также показано, как использовать DDM для характеристики этих композитов. Степенное масштабирование τ(q)~1/vqβ, с β = 1, указывает на баллистическое движение со скоростью v. Для справки, β = 2 представляет диффузионную динамику, а v является коэффициентом диффузии. Все активные композиты демонстрируют баллистическое масштабирование (рисунок 4A) со скоростями, которые настроены концентрациями актина и миозина (рисунок 4B), и которые могут изменяться во времени во время активности, ускоряясь или замедляясь (рисунок 4C, D).
Реструктуризацию и кластеризацию сети, видимую на рисунке 2 и более очевидную для более высоких концентраций актина и миозина, можно охарактеризовать с помощью SIA, как показано на рисунке 5 и описано ранее 17,48,50. Вкратце, длина корреляции ξ, которая является мерой характеристического размера объектов на изображении, может быть определена путем подгонки каждой кривой автокорреляции пространственной интенсивности g(r) к экспоненциальной функции расстояния r между пикселями. Большие пики g(r), которые сохраняются на больших расстояниях, указывают на более крупные структурные особенности (т.е. объединение, кластеризацию отдельных нитей). Как показано на рисунке 5, для более высоких фракций актина и концентраций миозина значительная реструктуризация и агрегация отражаются в увеличении ξ с течением времени.
Вязкоупругие свойства и нелинейный механический отклик активных композитов также могут быть измерены с помощью оптической микрореологии пинцета (OTM). Однако протоколы и репрезентативные результаты для этих экспериментов выходят за рамки этой работы. Заинтересованные читатели ссылаются на предыдущие работы48,56, в которых подробно описано, как выполнять измерения OTM и ожидаемые результаты.
Используя ту же программу экспериментальных и аналитических инструментов, описанную выше, в следующем разделе описывается, как динамика и структура изменяются при включении в композиты кинезиновых двигателей и сшивающих биотин-NA (рисунок 6, рисунок 7 и рисунок 8). На рисунке 6 показаны репрезентативные конфокальные изображения композитов, приводимые либо только кинезином (K), либо кинезином и миозином (K+M), с пассивным сшиванием (XL) актиновых нитей или микротрубочек и без них.
Включение кинезина в композиты первоначально приводит к аналогичной динамике и реструктуризации, что и композиты, управляемые миозином, как показано в верхней строке рисунка 7 (класс 1). Однако динамика обычно переходит к крупномасштабному анизотропному потоку (рисунок 7 средний ряд, класс 2), ускорению и замедлению (рисунок 7 нижний ряд, класс 3). Эти характеристики сочетаются с мезомасштабной кластеризацией и агрегацией через 5-30 мин (рисунок 6 и рисунок 8B). Поля потока, генерируемые PIV, и временные цветовые карты, показанные на рисунке 7 , показывают примеры изотропной реструктуризации (класс 1, верхняя панель), направленного потока (класс 2, средние панели) и двунаправленного ускорения (класс 3, нижние панели).
Скорости актина и микротрубочек в различных временных точках во время активности, определяемые с помощью подходов к кривым τ(q), иллюстрируют ускорение с последующим замедлением (рисунок 8), которое зависит от сшивки. Как также показано на рисунке 8, когда оба моторных белка включены, динамика на самом деле медленнее, чем композиты, содержащие только кинезин, и происходит задержка начала мезомасштабного потока. Миозин также поддерживает более однородное взаимопроникновение сетей актина и микротрубочек на протяжении всей продолжительности деятельности, а также меньшую агрегацию и реструктуризацию. Эти эффекты можно увидеть на изображениях на рисунке 6 и количественно определить по изменяющимся во времени длинам корреляции, вычисляемым с помощью SIA, которые, как правило, меньше в присутствии миозина (рисунок 8B).
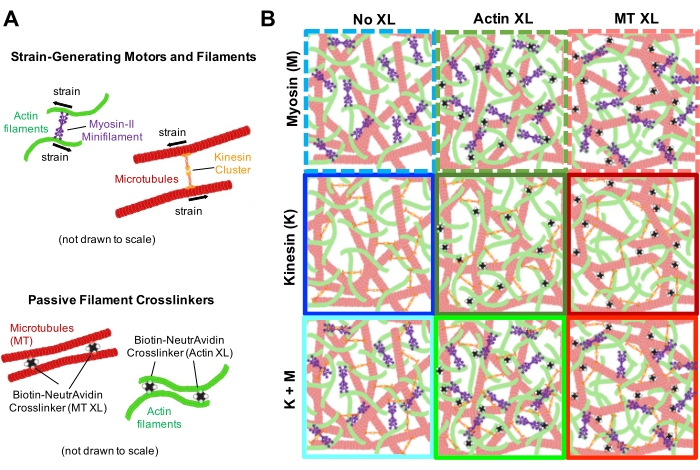
Рисунок 1. Проектирование и характеристика активных актин-микротрубочек с несколькими силообразующими двигателями и пассивными сшивателями. (A) Мономеры актина и димеры тубулина сополимеризуются при молярных концентрациях cAи cT0,73-11,6 мкМ и молярных фракциях актина ΦA= cA/ (cA+ cT) = 0, 0,25, 0,5, 0,75 и 1 с образованием совместно запутанных сетей актиновых нитей (зеленый) и микротрубочек (красный). Пассивное сшивание достигается с использованием NA для связывания биотинилированных актиновых нитей (Actin XL) или микротрубочек (MT XL) при молярных соотношениях сшиватель:белок RA= 0,01-0,08 и RMT= 0,001-0,01 для актина и микротрубочек соответственно. Мини-нити миозина-II (фиолетовые) и кинезиновые кластеры (оранжевый) при концентрациях cM= 0,12 - 0,48 мкМ и cK = 0,2 - 0,7 мкМ, толкают и натягивают нити для вывода композитов из стационарного состояния. (B) Схема пространства формулировки. Мини-нити миозина II (M), кинезиновые кластеры (K) или оба двигателя (K + M) включены в композиты без пассивных сшивателей (No XL), актин-актиновых сшиваний (Actin XL) и сшивок микротрубочек-микротрубочек (MT XL). Все мультфильмы не доведены до масштаба. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
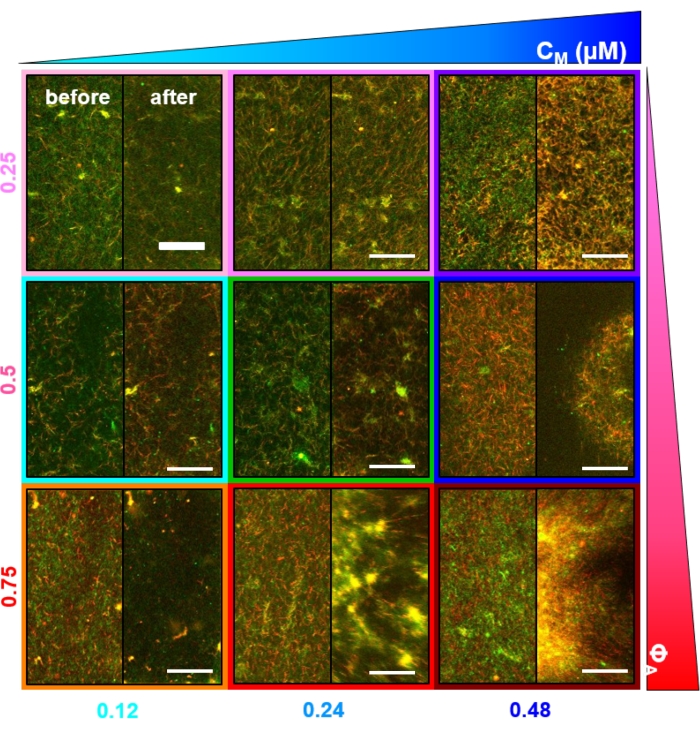
Рисунок 2. Двухцветная конфокальная визуализация миозин-управляемых композитов цитоскелетов с различными концентрациями миозина cM и молярных актиновых фракций ΦA. (A) Двухцветные изображения конфокальной микроскопии размером 256 x 128 квадратных пикселей (212 x 106 мкм2) показывают, как композиты актиновых нитей (зеленый) и микротрубочек (красный) перестраиваются с помощью двигательной активности миозина. Кинезиновые двигатели или пассивные сшиватели отсутствуют. На каждой панели показаны изображения, сделанные в начале (слева, до) и в конце (справа, после) 45-минутной активации миозина (через освещение светом 488 нм для деактивации блеббистатина). Панели упорядочиваются путем увеличения молярной концентрации миозина (cM), идущей слева направо, и увеличения молярной фракции актина (ΦA), идущей сверху вниз. Цвета, обозначающие каждую панель, соответствуют цветовому кодированию, используемому на рисунках 4 и 5. Шкала стержней составляет 50 мкМ. Для наилучшего захвата динамики и структуры для анализа мы используем частоту кадров 1-5 кадров в секунду, ROI со сторонами 50-250 мкм и длительности временных рядов 5-45 мин, в зависимости от скорости сжатия и перестановки. Панели, в которых изображения до и после выглядят одинаково, указывают на минимальную реструктуризацию, как видно на розовых, пурпурных и голубых панелях. Мелкомасштабная кластеризация, о чем свидетельствует повышенная неоднородность и наличие ярких пунктатных особенностей, прослеживается в оранжевых, зеленых и красных панелях. Крупномасштабное сокращение, рассматриваемое как равномерно сжимающаяся сеть, проявляется в синих и фиолетовых панелях. Этот показатель был изменен по сравнению со ссылкой17. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
Рисунок 3. Велоциметрия изображений частиц (PIV) показывает, что активность актомизина вызывает скоординированную сократительную динамику актина и микротрубочек в ко-запутанных композитах. Поля потока PIV для актина (верхний ряд) и микротрубочек (нижний ряд) в композите, управляемом миозином, с (ΦA, cM) = (0,5, 0,24) при увеличении времени в течение 6-минутного временного ряда. Поля потока были сгенерированы с помощью плагина Fiji/ImageJ PIV с временем задержки 20 с и биннингом 2 пикселя x 2 пикселя. Как актин, так и микротрубочки показывают последовательное движение, направленное к центральной области поля зрения на протяжении всей продолжительности фильма. Шкала на всех изображениях составляет 50 мкм. Различные цвета стрелок соответствуют различным скоростям, как указано в цветовой шкале справа от векторных полей. Этот показатель был изменен по сравнению со ссылкой26. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
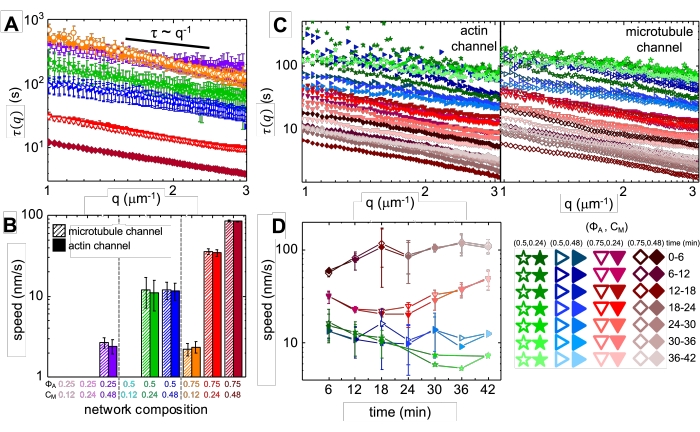
Рисунок 4. Дифференциальная динамическая микроскопия с временным разрешением (DDM) измеряет скорость и тип движения актина и микротрубочек в активных композитах. (A) DDM выполняется на микротрубочках (сверху, открытые символы) и актиновых (внизу, заполненные символы) каналах временных рядов для определения характеристического времени распада τ против волнового числа q как для актина (заполненные символы), так и для микротрубочек (открытые символы), как описано ранее17,26. Все кривые следуют масштабированию τ ~ q-1, указывающему на баллистическое движение, со скоростями v, которые определяются с помощью припадков к τ(q) = (vq)-1. Более высокие скорости соответствуют меньшим значениям τ(q) для любого заданного q. Цвета и фигуры символов соответствуют комбинациям (ΦA, c M), показанным в B. (B) Скорости сжатия v определяются с помощью подгонки к кривым τ(q), показанным в A, которые усредняются по всем временам задержки на протяжении каждого 45-минутного временного ряда. (C) DDM с временным разрешением (trDDM) количественно определяет, как динамика изменяется с течением времени, оценивая τ(q) для актина (заполненные символы, слева) и микротрубочек (открытые символы, справа) для последовательных 6-минутных интервалов (обозначаемых различными оттенками одного и того же цвета) в течение 45-минутного времени активации. trDDM выполняется для каждой (ΦA, cM) комбинации (обозначаемой различными символами и цветами), как описано в легенде в правом нижнем углу. Кривые τ(q), показанные в C, следуют тому же масштабированию и тенденциям, что и в A, но также показывают временную зависимость для определенных (ΦA, cM) композиций, особенно для ΦA= 0,75. (D) Скорости сжатия для актиновых нитей (закрытые символы) и микротрубочек (открытые символы) определяются от прилеганий к соответствующим кривым τ(q). Полосы ошибок на всех графиках представляют стандартную погрешность значений в трех-пяти репликах. Этот показатель был изменен по сравнению со ссылкой17. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
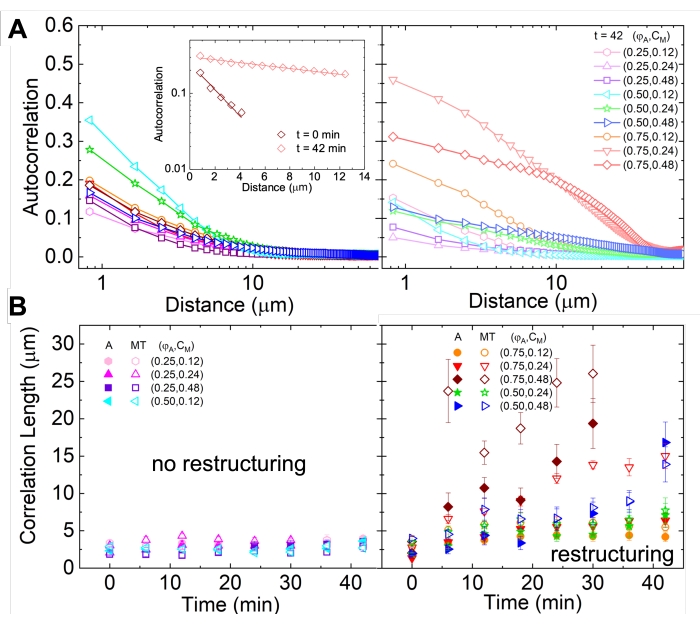
Рисунок 5. Анализ автокорреляции пространственного изображения (SIA) количественно определяет моторную реструктуризацию активных цитоскелетных композитов. (A) Автокорреляция g(r) для микротрубочек в начале (слева, t = 0 мин, темные оттенки) и конце (справа, t = 42 мин, светлые оттенки) эксперимента для (ΦA, cM) составов, перечисленных в легенде. Вставка: пример соответствия данных начальному и конечному 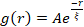 времени для (ΦA, cM) = (0,75, 0,12). (B) Средние длины корреляции ξ для актина (закрытые символы) и микротрубочек (открытые символы) для каждого (ΦA, cM), определяемые с помощью экспоненциальных припадков каждой кривой g(r), как показано во вставке в A. Данные делятся на те, которые демонстрируют минимальную (левую) и существенную (правую) реструктуризацию. Полосы ошибок в A и B представляют стандартную ошибку для трех-пяти реплик. Этот показатель был изменен по сравнению со ссылкой17. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
времени для (ΦA, cM) = (0,75, 0,12). (B) Средние длины корреляции ξ для актина (закрытые символы) и микротрубочек (открытые символы) для каждого (ΦA, cM), определяемые с помощью экспоненциальных припадков каждой кривой g(r), как показано во вставке в A. Данные делятся на те, которые демонстрируют минимальную (левую) и существенную (правую) реструктуризацию. Полосы ошибок в A и B представляют стандартную ошибку для трех-пяти реплик. Этот показатель был изменен по сравнению со ссылкой17. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
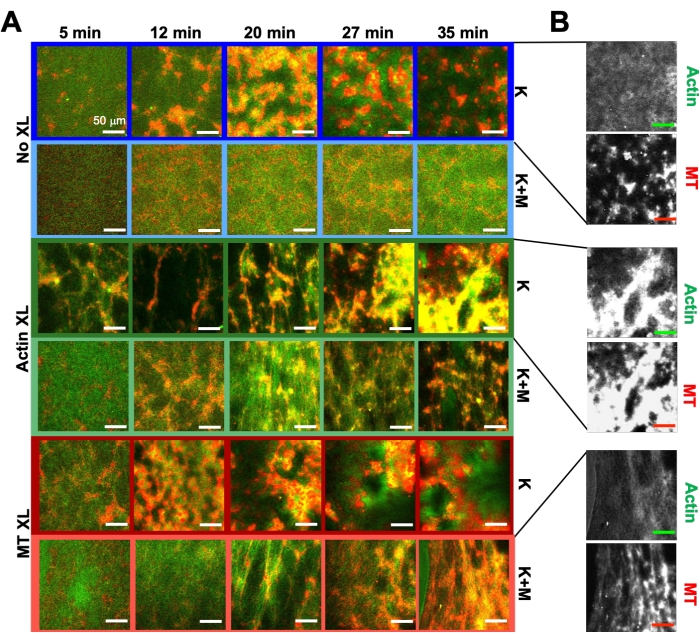
Рисунок 6. Включение кинезиновых двигателей и пассивных сшивателей в активные композиты для повышения программируемости и расширения фазового пространства динамики и структуры. (A) Двухцветные конфокальные изображения актина (зеленый) и микротрубочек (красный) в активных композитах показывают сложную рецептур-зависимую реструктуризацию с течением времени (перечислены в мин). Пять изображений в каждом ряду соответствуют пяти кадрам временного ряда 2000 кадров, полученных для композита, приводимого кинезином (K, строки 1, 3, 5) или кинезином и миозином (K+M, строки 2, 4, 6), и включающими либо отсутствие пассивных сшивателей (No XL, строки 1, 2), актин-актиновые сшивки (Actin XL, строки 3, 4) или сшивки микротрубочек-микротрубочек (MT XL, ряды 5, 6). Все шкалы составляют 50 мкм. Цвета контура соответствуют цветовой схеме на рисунке 8. (B) Отдельные каналы флуоресценции актина и микротрубочек для композитов, содержащих только кинезин, имеют различные структуры как с совместной локализацией актин-МТ, так и с микрофазным разделением. Показанные изображения предназначены для композитов с cA= 2,32 мкМ, cT= 3,48 мкМ, cK = 0,35 мкМ, cM= 0,47 мкМ (строки 2, 4, 6), RA = 0,02 (строки 3, 4) и RMT = 0,005 (строки 5, 6). Все композиты начинаются с равномерно распределенных взаимопроникающих сетей актина и микротрубочек (колонка 1). Кинезин-управляемые композиты без сшивающихся (строка 1) образуют слабо связанные аморфные кластеры, богатые MT. Актин сначала совместно локализуется в центрах этих агрегатов, но затем выдавливается из богатых МТ регионов, которые продолжают сокращаться и отключаться друг от друга. Сшивание актин-актин (строка 3) препятствует этому микромасштабному разделению актина и МТ, и вместо этого богатые МТ агрегаты соединяются через длинные нити актина. Сшивание актина также позволяет медленно поглощать актин в богатые MT области, так что композит становится связанной сетью колокализованных кластеров актина и MT. Сшивание микротрубочек (строка 5) приводит к аморфной кластеризации МТ, которые объединяются с течением времени, что приводит к более масштабному фазовому разделению актина и МТ. Добавление миозина (строки 2, 4, 6) уменьшает кинезиновое размешивание и реструктуризацию. Без сшивок (строка 2) композиты показывают небольшую перестановку в течение нескольких часов. Сшивание увеличивает реструктуризацию и колокализацию актина и микротрубочек (строки 4, 6). В частности, когда микротрубочки сшиты (строка 6), происходит значительное взаимопроникновение и реорганизация в паутиноподобные сети волокон. Этот показатель был изменен по сравнению со ссылкой50. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
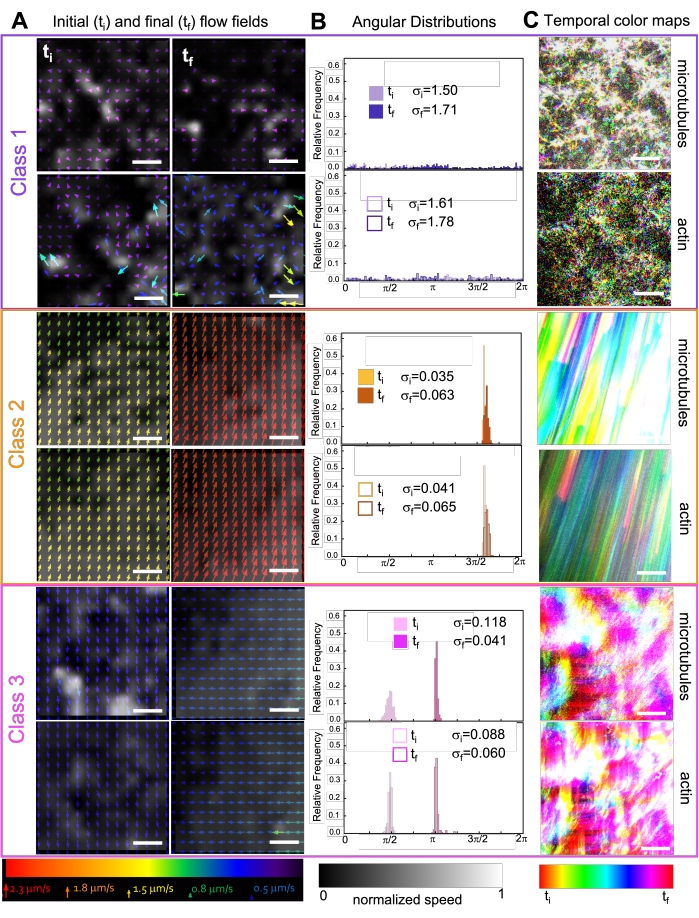
Рисунок 7. PIV показывает, что активные композиты демонстрируют три класса пространственно-временных различных полей потока. (A) Поля потока PIV для первого (ti) и последнего (tf) кадров трех репрезентативных временных рядов, показывающие различные динамические классы, которые составляются на рисунке 6 . Поля потока PIV для микротрубочек (вверху) и актина (внизу) для видео класса 1 (сверху, фиолетовый), класса 2 (средний, оранжевый) и класса 3 (внизу, пурпурный) с цветами стрелок, соответствующими универсальной шкале скорости внизу, и цветовой картой в градациях серого, показывающей пространственное распределение скорости, нормализованное отдельно для каждого поля потока в соответствии с масштабом, показанным внизу. Все шкалы составляют 50 мкМ. (B) Угловые распределения векторов скорости от A (в единицах радианов) с перечисленными начальными и конечными стандартными отклонениями σi и σf. (C) Временные цветовые карты для видео, проанализированных в форматах A и B, показывают покадровое положение каждого пикселя относительно его начальной точки. Карты класса 1 показывают мелкомасштабное случайное движение; карты класса 2 изображают быстрое однонаправленное движение с минимальными пространственными или временными вариациями; карты класса 3 демонстрируют особенности как класса 1, так и класса 2. Этот показатель был изменен по сравнению со ссылкой50. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
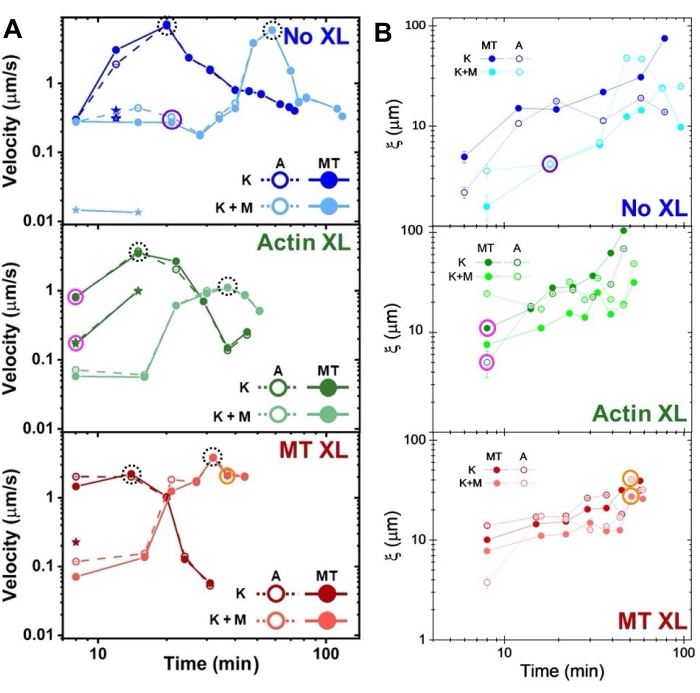
Рисунок 8. DDM и SIA измеряют изменяющуюся во времени динамику и структуру двухмоторных актин-микротрубочных композитов. (A) Скорости для композитов, описанные на рисунке 6 и рисунке 7, измеренные с помощью DDM, показывают ускорение и замедление композитов, запрограммированных сшиванием и активностью миозина. Скорости микротрубочек (МТ, замкнутые круги) и актина (А, открытые круги) строятся как функция времени активности в композитах без сшивки (верхняя, синяя), актиновое сшивание (середина, зеленый), сшивка микротрубочек (нижняя, красная), без миозина (К, более темные оттенки) и с миозином (К+М, более светлые оттенки). Для случаев класса 3, которые имеют две скорости, более медленная скорость обозначается звездой. Точки данных, заключенные пунктирными черными кругами, соответствуют максимальной скорости vmaxдля каждой формулировки. Полосы погрешностей (большинство из которых слишком малы, чтобы их видеть) являются стандартной ошибкой по степенному закону соответствия τ(q). (B) Структурные корреляционные длины ξ, определяемые с помощью SIA, по сравнению со временем активности для того же набора временных рядов, оцененных в A. Каждая точка данных представляет собой среднее значение длин корреляции, определенной для первого и последнего кадра соответствующего временного ряда. В общем, ξ увеличивается во времени как для актина, так и для микротрубочек во всех композитных системах, а композиты, приводимые исключительно кинезином, имеют большую длину корреляции, чем те, в которых миозин также присутствует. Точки данных в форматах A и B, соответствующие трем временным рядам, проанализированным на рисунке 7 , обведены соответствующим классом цвета (1 = фиолетовый, 2 = оранжевый, 3 = пурпурный). Этот показатель был изменен по сравнению со ссылкой50. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
Ключевым достижением восстановленной системы, описанной выше, является ее модульность и настраиваемость, поэтому пользователям рекомендуется изменять концентрации белков, двигателей, сшивателей и т. Д. В соответствии с их желаемыми результатами, будь то эмуляция конкретного клеточного процесса или разработка материала с определенной функциональностью или механическими свойствами. Ограничения диапазона концентраций актина и тубулина устанавливаются на нижнем пределе критической концентрацией, необходимой для полимеризации актина (~0,2 мкМ)57,58,59 и тубулина (~3 - 4 мкМ)60, и на верхнем пределе путем перехода к нематическому выравниванию актиновых нитей (~90 мкМ)61,62 или микротрубочек (~35 мкМ)63 . Мономеры актина и димеры тубулина должны полимеризоваться в нити вместе, а не смешиваться вместе после полимеризации, чтобы гарантировать, что они образуют однородно взаимопроникающие просачивающиеся сети, которые синергетически поддерживают друг друга. Новая динамика, которую демонстрируют композиты, опирается на это взаимодействие. Хотя, как правило, важно следовать всем шагам, изложенным в протоколе, для успешного воспроизведения показанных результатов, некоторые шаги являются более точными, в то время как другие имеют возможность модифицировать и корректировать в соответствии с конкретными потребностями и имеющимися ресурсами.
Например, одним из важных шагов в обеспечении воспроизводимых результатов является правильная подготовка и хранение реагентов в соответствии с руководящими принципами, приведенными в Таблице материалов. Цитоскелетные белки (актин, тубулин, миозин, кинезин) являются лабильными и должны быть аликвотированы, заморожены жидким азотом и храниться при -80 °C в одноразовых аликвотах. После удаления при -80 °C аликвоты следует держать на льду. Цитоскелетные белки не сохраняют надежную функцию после дополнительных циклов замораживания-оттаивания.
Микротрубочки более чувствительны к деполимеризации и денатурации, чем актин. После удаления при -80 °C тубулин следует держать на льду перед полимеризацией и использовать в течение 12 ч. После полимеризации микротрубочки следует хранить при комнатной температуре. Также крайне важно стабилизировать микротрубочки с помощью таксола для предотвращения деполимеризации. Фаллоидин-стабилизация актиновых нитей также важна для подавления АТФ-потребляющей беговой дорожки актина, которая конкурирует с активностью миозина и кинезина.
Ультрацентрифугирование миозиновых двигателей является еще одним важным шагом, поскольку оно удаляет неактивные мертвые головки миозина. Неудаление ферментативно неактивных мономеров приводит к пассивному сшиванию актиновой сети и потере активности. Для продления активности АТФазы двигателей может быть включена система регенерации АТФ, такая как креатинфосфат и креатинфосфокиназа64 .
Наконец, поддержание композиционной активности требует ингибирования адсорбции нитей и двигателей к стенкам камеры для отбора проб, что может быть достигнуто путем пассивации крышек и слайдов микроскопа. Моторные белки особенно склонны к адсорбции, в результате чего композит вытягивается на поверхность камеры образца, перемещается из поля зрения, коллапсирует до 2D и больше не подвергается активности. Силанизация обшивки и слайдов является эффективным способом пассивации поверхностей и предотвращения адсорбции (см. шаг 1). Альтернативным методом пассивации, эффективно используемым в экспериментах с цитоскелетом in vitro , является покрытие поверхности липидным бислоем, аналогичным клеточной мембране18. Этот метод является выгодным, если кто-то хочет привязать белки к поверхности или ввести другие специфические белково-поверхностные взаимодействия, потому что бислой может быть функционализирован. Для экспериментов с оптическим пинцетом пассивация микросфер также имеет решающее значение и может быть достигнута путем покрытия карбоксилированных микросфер BSA или PEG с помощью карбодиимидной сшивающей химии48.
Есть несколько аспектов представленных протоколов, которые исследователи могут рассмотреть возможность изменения в соответствии со своими потребностями. Во-первых, исследователи могут выбрать замену ненативных сшивающих биотин-NA биологическими сшивателями, такими как альфа-актинин или MAP65, которые сшивают актин и микротрубочки соответственно 28,65,66. Использование ненативных сшивающих устройств в композитах, описанных здесь, мотивировано их улучшенной воспроизводимостью, стабильностью и настраиваемостью по сравнению с нативными сшивателями. Из-за сильной связи биотин-NA можно считать, что сшивки являются постоянными, а не большинством нативных сшивающихся, которые временно связываются с широкими скоростями оборота. Динамика переходных сшиваний усложняет анализ вклада сшивающих устройств и двигателей в динамику. Кроме того, линкеры биотин-NA могут быть универсально использованы для сшивания как актина, так и микротрубочек, а также сшивания актина с микротрубочками. Таким образом, можно провести однозначное сравнение между мотивами сшивания, сохраняя все другие переменные (например, размер сшивания, сродство связывания, стехиометрию и т. Д.). Наконец, реагенты, необходимые для включения биотин-NA линкеров, широко коммерчески доступны, хорошо характеризуются и обычно используются во многих биофизических лабораториях. Тем не менее, одной из ключевых сильных сторон платформы in vitro, описанной здесь, является ее модульность, поэтому исследователи должны иметь возможность легко заменить линкеры biotin-NA на нативные линкеры, если они захотят.
Во-вторых, в настоящем протоколе мономеры актина и димеры тубулина полимеризуются в нити вместе в центрифужной трубке перед добавлением в камеру для отбора проб. Поступление раствора запутанных нитевидных белков в камеру образца может привести к выравниванию потока, особенно микротрубочек, что нарушает желаемую изотропию и однородность композитов. Действительно, крупным достижением в предыдущих работах по стационарным актин-микротрубочевым композитам была возможность совместной полимеризации актина и микротрубочек in situ (в камере образца) для обеспечения формирования изотропных взаимопроникающих сетей актина и микротрубочек 15,16,27. Однако распространение этого подхода на активные композиты потребует добавления двигателей к образцу до полимеризации актина и тубулина и инкубации всего образца вместе при 37 °C до начала экспериментов. Испытания этого изменения протокола привели к снижению полимеризации актина и отсутствию заметной двигательной активности, вероятно, из-за конкурирующей активности АТФазы и длительной инкубации двигателей при 37 °C. К счастью, при следовании текущим протоколам не наблюдается заметного выравнивания потока композитов, как показано на рисунке 2, рисунке 3 и рисунке 6. Тем не менее, исследователям рекомендуется разрабатывать протоколы, которые позволяют формировать in situ активные композиты.
Другим моментом рассмотрения является схема флуоресцентной маркировки, которая влечет за собой разреженную маркировку всех актиновых нитей и микротрубочек в сети. Этот подход к маркировке был оптимизирован для непосредственной визуализации структуры сети, а не для вывода структуры и динамики с помощью трассирующих нитей или микросфер. Однако компромисс заключается в том, что отдельные нити не имеют яркой маркировки и разрешимости. Один из подходов, который исследователи могут использовать как для разрешения отдельных нитей, так и для визуализации структуры сети, заключается в легировании в предварительно сформированных нитях, помеченных другим флуорофором, поэтому как окружающая сеть, так и отдельные нити могут быть изображены одновременно. Однако при использовании более двух флуорофоров и каналов возбуждения / излучения кровотечение между каналами часто трудно устранить, поэтому необходимо соблюдать осторожность при выборе флуорофоров, фильтров и интенсивности лазера.
Связанным с этим ограничением является неспособность визуализировать миозиновые или кинезиновые двигатели в композитах. Используемые флуоресцентно меченые мономеры актина и димеры тубулина коммерчески доступны, тогда как визуализация миозина или кинезина в композитах требует собственной маркировки. Исследователям предлагается сделать следующий шаг к маркировке двигателей, как это было сделано ранее18,67, чтобы иметь возможность однозначно связать двигательную активность и связывание с динамикой и структурами, которые демонстрируют наши композиты.
Наконец, важно отметить, что в текущем протоколе начало и продолжительность активности кинезина не контролируются. Поскольку активность миозина контролируется с помощью фотодеактивации блеббистатина, как описано выше, для создания аналогичной световой активации кинезина, можно включить светоактивированную АТФ.
Чтобы создать сложность конструкций, описанных здесь, чтобы лучше имитировать клеточные условия и расширить пространство параметров динамической структуры-функции, будущая работа будет сосредоточена на включении промежуточных нитей, таких как виментин 68,69, а также других двигателей, таких как динеин 13,70. Гельсолин также будет включен в различных концентрациях для контроля длины актина14, а также тау-белок для контроля жесткости микротрубочек.
Таким образом, представленные протоколы описывают, как проектировать, создавать и характеризовать динамику, структуру и механику систем активной материи, вдохновленных цитоскелетами, которые содержат два отдельных активных силообразующих компонента, которые действуют на разные субстраты в одной системе. Эта перестраиваемая и модульная платформа приближает усилия по восстановлению на один важный шаг ближе к имитации клеточного цитоскелета и предлагает уникальную возможность программировать его свойства в широком фазовом пространстве путем независимого включения, удаления и настройки различных компонентов. Кроме того, все компоненты этой универсальной системы коммерчески доступны (см. Таблицу материалов), за исключением кинезиновых димеров, которые очищаются в лаборатории Росса, как описано ранее50, и доступны по запросу. Наконец, весь код анализа находится в свободном доступе через GitHub49 и основан на свободных языках программирования и программном обеспечении (Python и Fiji). Мы надеемся, что прозрачное распространение протоколов для разработки этих систем сделает эту платформу более доступной для разнообразной группы пользователей с различным опытом, опытом, институциональной принадлежностью и исследовательскими целями.
Авторам нечего раскрывать
Мы благодарим Майю Хендию и д-ра Джонатана Мишеля за помощь в анализе данных, а также д-ра Джанет Шеунг, д-ра Мумиту Даса и д-ра Майкла Раста за полезные обсуждения и рекомендации. Это исследование было поддержано исследовательским грантом Фонда Уильяма М. Кека и премией NSF DMREF (DMR 2119663), присуждаемой RMRA и JLR и грантами R15 Национальных институтов здравоохранения (R15GM123420, 2R15GM123420-02), присуждаемыми RMR-A и RJM.
| Name | Company | Catalog Number | Comments |
| (-)-Blebbistatin Abbreviation used in paper: blebbistatin | Sigma Aldrich | B0560 | Stock Concentration: 200 μM in DMSO Storage: dessicated, in DMSO, -20ºC Stock and Experiment Recipes: dissolve 1 mg of powder to 200 μM in DMSO Storage, Handling, Troubleshooting Notes: limited shelf-life, typically stops functioning reliably after 3-4 months. purchase and prepare new solution every 3 months. |
| 1:20 488-tubulin:tubulin mixture Abbreviation used in paper: 5-488-tubulin | NA | NA | Stock Concentration: 5 mg/ml in PEM Storage: single use aliquots, -80ºC, avoid freeze-thaw cycles Stock and Experiment Recipes: mix tubulin and 488-tubulin at a 20:1 ratio, flash freeze with LN2 Storage, Handling, Troubleshooting Notes: each aliquot can be used for up to 12 hrs stored on ice at 4ºC, protect from light |
| 1:20 R-tubulin:tubulin mixture Abbreviation used in paper: 5-R-tubulin | NA | NA | Stock Concentration: 5 mg/ml in PEM Storage: single use aliquots, -80ºC, avoid freeze-thaw cycles Stock and Experiment Recipes: mix tubulin and rhodamine tubulin at a 20:1 ratio, flash freeze with LN2 Storage, Handling, Troubleshooting Notes: each aliquot can be used for up to 12 hrs stored on ice at 4ºC, protect from light |
| actin (biotin): skeletal muscle Abbreviation used in paper: biotin-actin | Cytoskeleton | AB07 | Stock Concentration: 1 mg/ml in G-buffer Storage: single use aliquots, -80ºC, avoid freeze-thaw cycles Stock and Experiment Recipes: reconstitute to 1 mg/ml in G-buffer, flash freeze with LN2 Storage, Handling, Troubleshooting Notes: (1) immediately prior to use dilute to 0.5 mg/ml in PEM, (2) once removed from -80ºC, store aliquot on ice at 4ºC for up to 1 week |
| actin (rhodamine): rabbit skeletal muscle Abbreviation used in paper: R-actin | Cytoskeleton | AR05 | Stock Concentration: 1.5 mg/ml in G-buffer Storage: single use aliquots, -80ºC, avoid freeze-thaw cycles Stock and Experiment Recipes: reconstitute to 1.5 mg/ml in G-buffer, flash freeze with LN2 Storage, Handling, Troubleshooting Notes: once removed from -80ºC, store aliquot on ice at 4ºC, can be used for up to 1 week |
| adenosine triphosphate Abbreviation used in paper: ATP | Thermo Fisher Scientific | A1048 | Stock Concentration: 100 mM Storage: in solution (pH 7), -20ºC Stock and Experiment Recipes: reconsitute in DI H20, bring pH to 7 with NaOH Storage, Handling, Troubleshooting Notes: routinely check pH and adjust as needed, hydrolyzes over time, replace every ~6-12 months |
| AlexaFluor488 Phalloidin Abbreviation used in paper: 488-phalloidin | Thermo Fisher Scientific | A12379 | Stock Concentration: 100 μM DMSO Storage: protected from light, dessicated, -20ºC Stock and Experiment Recipes: reconstitute to 100 μM with DMSO Storage, Handling, Troubleshooting Notes: immediately prior to use dilute to 20 μM in PEM (1 μL in 4 μL PEM) |
| AlexaFluor488–labeled actin Abbreviation used in paper: 488-actin | Thermo Fisher Scientific | A12373 | Stock Concentration: 1.5 mg/ml in G-buffer Storage: single use aliquots, -80ºC, avoid freeze-thaw cycles Stock and Experiment Recipes: reconstitute to 1.5 mg/ml in G-buffer, flash freeze with LN2 Storage, Handling, Troubleshooting Notes: this item has been discontinued |
| Basic Plasma Cleaner Abbreviation used in paper: plasma cleaner | Harrick Plasma | PDC-32G | |
| Bemis Parafilm M Laboratory Wrapping Film Abbreviation used in paper: transparent film | Thermo Fisher Scientific | 13-374-5 | |
| D-(+)-Glucose Abbreviation used in paper: | Thermo Fisher Scientific | A1682836 | Stock Concentration: 100x Storage: store at stock concentration (100x) or 10x concentration, dessicated, at -20ºC Stock and Experiment Recipes: reconstitute powder to 4.5 mg/ml in DI H20 Storage, Handling, Troubleshooting Notes: final concentration in solution should 45 μg/mL |
| D-Biotin Abbreviation used in paper: biotin | Fisher Scientific | BP232-1 | Stock Concentration: 1.02 mM in PEM Storage: dessicated, 4ºC |
| deionized nanopure water Abbreviation used in paper: DI | |||
| Dimethyldichlorosilane Abbreviation used in paper: silane | Thermo Fisher Scientific | D/3820/PB05 | Stock Concentration: 2% dissolved in Toulene |
| Dithiothreitol Abbreviation used in paper: DTT | Thermo Fisher Scientific | R0861 | Stock Concentration: 1 M in DMSO Storage: dessicated, -20ºC Stock and Experiment Recipes: dilute to 2 mM in PEM immediately before each experiment |
| DMSO Anhydrous Abbreviation used in paper: DMSO | Thermo Fisher Scientific | D12345 | |
| F-Buffer Abbreviation used in paper: F-buffer | NA | NA | Stock Concentration: 10x Storage: dessicated, -20ºC Stock and Experiment Recipes: 10 mM Imidazole (pH 7.0), 50 mM KCl, 1 mM MgCl2, 1 mM EGTA, 0.2 mM ATP |
| G-Buffer Abbreviation used in paper: G-buffer | NA | NA | Stock Concentration: 10x Storage: dessicated, -20ºC Stock and Experiment Recipes: 2.0 mM Tris (pH 8), 0.2 mM ATP, 0.5 mM DTT, 0.1 mM CaCl2. Store at -20°C. |
| glass microscope slide Abbreviation used in paper: slide | Thermo Fisher Scientific | 22-310397 | |
| Glucose oxidase + catalase + β-mercaptoethanol Abbreviation used in paper: GOC | Sigma Aldrich | G2133-250KU, C1345, 63689 | Stock Concentration: 100x Storage: store at stock concentration (100x) or 10x concentration, dessicated, at -20ºC Stock and Experiment Recipes: For 100x: 4.3 mg/ml glucose oxidase, 0.7 mg/ml catalase, 0.5% v/v β-mercaptoethanol in DI H20 Storage, Handling, Troubleshooting Notes: final concentration in solution should be: 0.005% β-mercaptoethanol, 43 μg/mL glucose oxidase, 7 μg/mL catalase |
| glu-GOC oxygen scavenging system Abbreviation used in paper: glu-GOC | NA | NA | Stock Concentration: 100x Storage: prepare fresh each time Stock and Experiment Recipes: mix equal parts Glu and GOC and add at 1/100 final sample volume immediately before imaging Storage, Handling, Troubleshooting Notes: prepare from Glu and GOC immediately before imaging |
| Guanosine triphosphate Abbreviation used in paper: GTP | Thermo Fisher Scientific | R0461 | Stock Concentration: 100 mM Storage: 100 μL aliquots at -20ºC |
| Instant Mix 1-minute epoxy Abbreviation used in paper: epoxy | Loctite | 1366072 | |
| Kinesin-1 401 BIO 6x HIS Abbreviation used in paper: kinesin | Prepared in JL Ross Lab at Syracuse University | NA | Stock Concentration: 8.87 μM in PEM Storage: single use aliquots, -80ºC, avoid freeze-thaw cycles Storage, Handling, Troubleshooting Notes: biotinylated dimers form kinesin clusters, each aliquot can be used for up to 12 hrs stored on ice at 4ºC |
| NeutrAvidin Abbreviation used in paper: NA | Thermo Fisher Scientific | 31000 | Stock Concentration: 5 mg/ml in PEM Storage: dessicated, -20ºC Stock and Experiment Recipes: reconstitute powder to 5 mg/ml in PEM |
| No 1. glass coverslips (24 mm x 24 mm) Abbreviation used in paper: coverslip | Thermo Fisher Scientific | 12-548-CP | |
| Paclitaxel Abbreviation used in paper: Taxol | Thermo Fisher Scientific | P3456 | Stock Concentration: 2 mM in DMSO Storage: protected from light, dessicated, -20ºC Stock and Experiment Recipes: reconstitute to 2 mM with DMSO Storage, Handling, Troubleshooting Notes: immediately prior to use dilute to 200 μM in DMSO (0.4 μL in 3.6 μL DMSO) |
| PEM-100 Abbreviation used in paper: PEM | NA | NA | Stock Concentration: 1x Storage: room temperature (RT) Stock and Experiment Recipes: 100 mM K-PIPES (pH 6.8), 2 mM EGTA, 2 mM MgCl2 Storage, Handling, Troubleshooting Notes: use KOH to adjust pH to 6.8, recheck pH often and adjust accordingly |
| phalloidin Abbreviation used in paper: phalloidin | Thermo Fisher Scientific | P3457 | Stock Concentration: 100 μM in DMSO Storage: protected from light, dessicated, -20ºC, adhere closely to storage/handling conditions Stock and Experiment Recipes: reconstitute to 100 μM with DMSO Storage, Handling, Troubleshooting Notes: susceptible to impurities in its preparation and denaturing, identifiable as large amorphous aggregates of actin in samples |
| porcine brain tubulin Abbreviation used in paper: tubulin | Cytoskeleton | T240 | Stock Concentration: 5 mg/ml in PEM Storage: single use aliquots, -80ºC, avoid freeze-thaw cycles Stock and Experiment Recipes: reconstitute powder to 5 mg/ml in PEM, flash freeze with LN2 Storage, Handling, Troubleshooting Notes: each aliquot can be used for up to 12 hrs stored on ice at 4ºC |
| Potassium Chloride Abbreviation used in paper: KCl | Thermo Fisher Scientific | AM9640G | Stock Concentration: 4 M Storage: RT |
| Rabbit skeletal actin Abbreviation used in paper: actin | Cytoskeleton | AKL99 | Stock Concentration: 2 mg/ml in G-buffer Storage: single use aliquots, -80ºC, avoid freeze-thaw cycles Stock and Experiment Recipes: reconstitute to 2 mg/ml in G-buffer, flash freeze with LN2 Storage, Handling, Troubleshooting Notes: once removed from -80ºC, store aliquot on ice at 4ºC, can be used for up to 1 week |
| Rabbit skeletal myosin II Abbreviation used in paper: myosin | Cytoskeleton | MY02 | Stock Concentration: 10 mg/ml in PEM Storage: single use aliquots, -80ºC, avoid freeze-thaw cycles Stock and Experiment Recipes: reconstitute powder to 10 mg/ml in PEM, flash freeze with LN2 Storage, Handling, Troubleshooting Notes: monomers form minifilaments at low KCl, each aliquot can be used for up to 12 hrs stored on ice at 4ºC |
| Tubulin (biotin): porcine brain Abbreviation used in paper: biotin-tubulin | Cytoskeleton | T333P | Stock Concentration: 5 mg/ml in PEM Storage: single use aliquots, -80ºC, avoid freeze-thaw cycles Stock and Experiment Recipes: reconstitute powder to 5 mg/ml in PEM, flash freeze with LN2 Storage, Handling, Troubleshooting Notes: immediately prior to use dilute to 0.5 mg/ml in PEM |
| Tubulin (fluorescent HiLyte 488): porcine brain Abbreviation used in paper: 488-tubulin | Cytoskeleton | TL488M | Stock Concentration: 5 mg/ml in PEM Storage: single use aliquots, -80ºC, avoid freeze-thaw cycles Stock and Experiment Recipes: reconstitute powder to 5 mg/ml in PEM, flash freeze with LN2 Storage, Handling, Troubleshooting Notes: each aliquot can be used for up to 12 hrs stored on ice at 4ºC, protect from light |
| tubulin (rhodamine): porcine brain Abbreviation used in paper: R-tubulin | Cytoskeleton | TL590M | Stock Concentration: 5 mg/ml in PEM Storage: single use aliquots, -80ºC, avoid freeze-thaw cycles Stock and Experiment Recipes: reconstitute powder to 5 mg/ml in PEM, flash freeze with LN2 Storage, Handling, Troubleshooting Notes: each aliquot can be used for up to 12 hrs stored on ice at 4ºC, protect from light |
| Tween 20 Abbreviation used in paper: Tween20 | Thermo Fisher Scientific | J20605.AP | Stock Concentration: 1% v/v in DI H20 Storage: RT |
| ultracentrifuge grade microtubes Abbreviation used in paper: Beckman-Coulter Optima Max XP | Beckman Coultier | 343776 | Storage, Handling, Troubleshooting Notes: 8x34 mm PC |
| UV light curing glue Abbreviation used in paper: UV glue | Pharda | SKG-2869 |
- Fletcher, D. A., Mullins, R. D. Cell mechanics and the cytoskeleton. Nature. 463 (7280), 485-492 (2010).
- Koenderink, G. H., Paluch, E. K. Architecture shapes contractility in actomyosin networks. Current Opinion in Cell Biology. 50, 79-85 (2018).
- Dogterom, M., Koenderink, G. H. Actin-microtubule crosstalk in cell biology. Nature Reviews Molecular Cell Biology. 20 (1), 38-54 (2019).
- Burla, F., Mulla, Y., Vos, B. E., Aufderhorst-Roberts, A., Koenderink, G. H. From mechanical resilience to active material properties in biopolymer networks. Nature Reviews Physics. 1 (4), 249-263 (2019).
- Wen, Q., Janmey, P. A. Polymer physics of the cytoskeleton. Current Opinion in Solid State and Materials Science. 15 (5), 177-182 (2011).
- Xiao, Q., Hu, X., Wei, Z., Tam, K. Y. Cytoskeleton molecular motors: structures and their functions in neuron. International Journal of Biological Sciences. 12 (9), 1083-1092 (2016).
- Ajeti, V. et al. Wound healing coordinates actin architectures to regulate mechanical work. Nature Physics. 15 (7), 696-705 (2019).
- Jung, W. et al. Dynamic motions of molecular motors in the actin cytoskeleton. Cytoskeleton. 76 (11-12), 517-531 (2019).
- Pollard, T. D., O'Shaughnessy, B. Molecular mechanism of cytokinesis. Annual Review of Biochemistry. 88 (1), 661-689 (2019).
- Huber, F., Boire, A., López, M. P., Koenderink, G. H. Cytoskeletal crosstalk: when three different personalities team up. Current Opinion in Cell Biology. 32, 39-47 (2015).
- Rivero, F. et al. The role of the cortical cytoskeleton: F-actin crosslinking proteins protect against osmotic stress, ensure cell size, cell shape and motility, and contribute to phagocytosis and development. Journal of Cell Science. 109 (11), 2679-2691 (1996).
- Duclos, G. et al. Topological structure and dynamics of three-dimensional active nematics. Science. 367 (6482), 1120-1124 (2020).
- Baclayon, M. et al. Optical tweezers-based measurements of forces and dynamics at microtubule ends. Optical Tweezers. 1486, 411-435 (2017).
- Gurmessa, B., Fitzpatrick, R., Falzone, T. T., Robertson-Anderson, R. M. Entanglement density tunes microscale nonlinear response of entangled actin. Macromolecules. 49 (10), 3948-3955 (2016).
- Francis, M. L. et al. Non-monotonic dependence of stiffness on actin crosslinking in cytoskeleton composites. Soft Matter. 15 (44), 9056-9065 (2019).
- Ricketts, S. N. et al. Varying crosslinking motifs drive the mesoscale mechanics of actin-microtubule composites. Scientific Reports. 9 (1), 12831 (2019).
- Lee, G. et al. Active cytoskeletal composites display emergent tunable contractility and restructuring. Soft Matter. 17 (47), 10765-10776 (2021).
- Murrell, M. P., Gardel, M. L. F-actin buckling coordinates contractility and severing in a biomimetic actomyosin cortex. Proceedings of the National Academy of Sciences. 109 (51), 20820-20825 (2012).
- Soares e Silva, M. et al. Active multistage coarsening of actin networks driven by myosin motors. Proceedings of the National Academy of Sciences. 108 (23), 9408-9413 (2011).
- Sonn-Segev, A., Bernheim-Groswasser, A., Roichman, Y. Dynamics in steady state in vitro acto-myosin networks. Journal of Physics: Condensed Matter. 29 (16), 163002 (2017).
- Ideses, Y., Sonn-Segev, A., Roichman, Y., Bernheim-Groswasser, A. Myosin II does it all: assembly, remodeling, and disassembly of actin networks are governed by myosin II activity. Soft Matter. 9 (29), 7127 (2013).
- Fürthauer, S. et al. Self-straining of actively crosslinked microtubule networks. Nature Physics. 15 (12), 1295-1300 (2019).
- Lemma, L. M. et al. Multiscale microtubule dynamics in active nematics. Physical Review Letters. 127 (14), 148001 (2021).
- Fan, Y., Wu, K.-T., Aghvami, S. A., Fraden, S., Breuer, K. S. Effects of confinement on the dynamics and correlation scales in kinesin-microtubule active fluids. Physical Review E. 104 (3), 034601 (2021).
- Triclin, S. et al. Self-repair protects microtubules from destruction by molecular motors. Nature Materials. 20 (6), 883-891 (2021).
- Lee, G. et al. Myosin-driven actin-microtubule networks exhibit self-organized contractile dynamics. Science Advances. 7 (6), eabe4334 (2021).
- Ricketts, S. N., Ross, J. L., Robertson-Anderson, R. M. Co-entangled actin-microtubule composites exhibit tunable stiffness and power-law stress relaxation. Biophysical Journal. 115 (6), 1055-1067 (2018).
- Bendix, P. M. et al. A quantitative analysis of contractility in active cytoskeletal protein networks. Biophysical Journal. 94 (8), 3126-3136 (2008).
- Linsmeier, I. et al. Disordered actomyosin networks are sufficient to produce cooperative and telescopic contractility. Nature Communications. 7 (1), 12615 (2016).
- Stam, S. et al. Filament rigidity and connectivity tune the deformation modes of active biopolymer networks. Proceedings of the National Academy of Sciences. 114 (47), E10037-E10045 (2017).
- Yadav, V. et al. Filament nucleation tunes mechanical memory in active polymer networks. Advanced Functional Materials. 29 (49), 1905243 (2019).
- Ennomani, H. et al. Architecture and connectivity govern actin network contractility. Current Biology. 26 (5), 616-626 (2016).
- Alvarado, J., Sheinman, M., Sharma, A., MacKintosh, F. C., Koenderink, G. H. Molecular motors robustly drive active gels to a critically connected state. Nature Physics. 9 (9), 591-597 (2013).
- Alvarado, J., Cipelletti, L., Koenderink, G. H. Uncovering the dynamic precursors to motor-driven contraction of active gels. Soft Matter. 15 (42), 8552-8565 (2019).
- Jung, W., Murrell, M. P., Kim, T. F-actin cross-linking enhances the stability of force generation in disordered actomyosin networks. Computational Particle Mechanics. 2 (4), 317-327 (2015).
- Lenz, M., Thoresen, T., Gardel, M. L., Dinner, A. R. Contractile units in disordered actomyosin bundles arise from f-actin buckling. Physical Review Letters. 108 (23), 238107 (2012).
- Memarian, F.L. et al. Active nematic order and dynamic lane formation of microtubules driven by membrane-bound diffusing motors. Proceedings of the National Academy of Sciences. 118 (52), e2117107118 (2021).
- Needleman, D., Dogic, Z. Active matter at the interface between materials science and cell biology. Nature Reviews Materials. 2 (9), 17048 (2017).
- Foster, P. J., Fürthauer, S., Shelley, M. J., Needleman, D. J. Active contraction of microtubule networks. eLife. 4, e10837 (2015).
- Thijssen, K. et al. Submersed micropatterned structures control active nematic flow, topology, and concentration. Proceedings of the National Academy of Sciences. 118 (38), e2106038118 (2021).
- Sanchez, T., Chen, D. T. N., DeCamp, S. J., Heymann, M., Dogic, Z. Spontaneous motion in hierarchically assembled active matter. Nature. 491 (7424), 431-434 (2012).
- Colen, J. et al. Machine learning active-nematic hydrodynamics. Proceedings of the National Academy of Sciences. 118 (10), e2016708118 (2021).
- Mitchell, K. A., Tan, A. J., Arteaga, J., Hirst, L. S. Fractal generation in a two-dimensional active-nematic fluid. Chaos: An Interdisciplinary Journal of Nonlinear Science. 31 (7), 073125 (2021).
- Pandolfi, R. J., Edwards, L., Johnston, D., Becich, P., Hirst, L. S. Designing highly tunable semiflexible filament networks. Physical Review E. 89 (6), 062602 (2014).
- Tan, A. J. et al. Topological chaos in active nematics. Nature Physics. 15 (10), 1033-1039 (2019).
- Roostalu, J., Rickman, J., Thomas, C., Nédélec, F., Surrey, T. Determinants of polar versus nematic organization in networks of dynamic microtubules and mitotic motors. Cell. 175 (3), 796-808.e14 (2018).
- Ndlec, F. J., Surrey, T., Maggs, A. C., Leibler, S. Self-organization of microtubules and motors. Nature. 389 (6648), 305-308 (1997).
- Sheung, J. Y. et al. Motor-driven restructuring of cytoskeleton composites leads to tunable time-varying elasticity. ACS Macro Letters. 10 (9), 1151-1158 (2021).
- McGorty, R. PyDDM v0.2.0. Zenodo. (2022).
- Achiriloaie, D. H. et al. Kinesin and myosin motors compete to drive rich multi-phase dynamics in programmable cytoskeletal composites. arXiv. (2021).
- Wulstein, D. M., Regan, K. E., Garamella, J., McGorty, R. J., Robertson-Anderson, R. M. Topology-dependent anomalous dynamics of ring and linear DNA are sensitive to cytoskeleton crosslinking. Science Advances. 5 (12), eaay5912 (2019).
- McGorty, R. Image-Correlation. at <https://github.com/rmcgorty/Image-Correlation>. (2020).
- Robertson, C. Theory and practical recommendations for autocorrelation-based image correlation spectroscopy. Journal of Biomedical Optics. 17 (8), 080801 (2012).
- McGorty, R. Differential Dynamic Microscopy - Python. at <https://github.com/rmcgorty/Differential-Dynamic-Microscopy---Python>. (2021).
- Cerbino, R., Trappe, V. Differential dynamic microscopy: probing wave vector dependent dynamics with a microscope. Physical Review Letters. 100 (18), 188102 (2008).
- Robertson-Anderson, R. M. Optical tweezers microrheology: from the basics to advanced techniques and applications. ACS Macro Letters. 7 (8), 968-975 (2018).
- Pollard, T. D. Polymerization of ADP-actin. Journal of Cell Biology. 99 (3), 769-777 (1984).
- Coué, M., Brenner, S. L., Spector, I., Korn, E. D. Inhibition of actin polymerization by latrunculin A. FEBS Letters. 213 (2), 316-318 (1987).
- Pollard, T. D. Actin and actin-binding proteins. Cold Spring Harbor Perspectives in Biology. 8 (8), a018226 (2016).
- Kumar, N. Taxol-induced polymerization of purified tubulin. Mechanism of action. Journal of Biological Chemistry. 256 (20), 10435-10441 (1981).
- Käs, J. et al. F-actin, a model polymer for semiflexible chains in dilute, semidilute, and liquid crystalline solutions. Biophysical Journal. 70 (2), 609-625 (1996).
- Viamontes, J., Narayanan, S., Sandy, A. R., Tang, J. X. Orientational order parameter of the nematic liquid crystalline phase of F -actin. Physical Review E. 73 (6), 061901 (2006).
- Hitt, A. L., Cross, A. R., Williams, R. C. Microtubule solutions display nematic liquid crystalline structure. Journal of Biological Chemistry. 265 (3), 1639-1647 (1990).
- Andexer, J. N., Richter, M. Emerging enzymes for ATP regeneration in biocatalytic processes. ChemBioChem. 16 (3), 380-386 (2015).
- Farhadi, L. et al. Actin and microtubule crosslinkers tune mobility and control co-localization in a composite cytoskeletal network. Soft Matter. 16 (31), 7191-7201 (2020).
- Falzone, T. T., Lenz, M., Kovar, D. R., Gardel, M. L. Assembly kinetics determine the architecture of α-actinin crosslinked F-actin networks. Nature Communications. 3 (1), 861 (2012).
- Thoresen, T., Lenz, M., Gardel, M. L. Reconstitution of contractile actomyosin bundles. Biophysical Journal. 100 (11), 2698-2705 (2011).
- Sanghvi-Shah, R., Weber, G. F. Intermediate filaments at the junction of mechanotransduction, migration, and development. Frontiers in Cell and Developmental Biology. 5, 81 (2017).
- Shen, Y. et al. Effects of vimentin intermediate filaments on the structure and dynamics of in vitro multicomponent interpenetrating cytoskeletal networks. Physical Review Letters. 127 (10), 108101 (2021).
- Laan, L., Roth, S., Dogterom, M. End-on microtubule-dynein interactions and pulling-based positioning of microtubule organizing centers. Cell Cycle. 11 (20), 3750-3757 (2012).
Erratum
Erratum: Reconstituting and Characterizing Actin-Microtubule Composites with Tunable Motor-Driven Dynamics and MechanicsAn erratum was issued for: Reconstituting and Characterizing Actin-Microtubule Composites with Tunable Motor-Driven Dynamics and Mechanics. The Authors section was updated.
Mehrzad Sasanpour1
Daisy H. Achiriloaie1,2
Gloria Lee1
Gregor Leech1
Christopher Currie1
K. Alice Lindsay3
Jennifer L. Ross3
Ryan J. McGorty1
Rae M. Robertson-Anderson1
1Department of Physics and Biophysics, University of San Diego
2W. M. Keck Science Department, Scripps College, Pitzer College, and Claremont McKenna College
3Department of Physics, Syracuse University
to:
Mehrzad Sasanpour1
Daisy H. Achiriloaie1,2
Gloria Lee1
Gregor Leech1
Maya Hendija1
K. Alice Lindsay3
Jennifer L. Ross3
Ryan J. McGorty1
Rae M. Robertson-Anderson1
1Department of Physics and Biophysics, University of San Diego
2W. M. Keck Science Department, Scripps College, Pitzer College, and Claremont McKenna College
3Department of Physics, Syracuse University
ABOUT JoVE
Copyright © 2024 MyJoVE Corporation. All rights reserved