
Summary
Abstract
Introduction
Protocol
Representative Results
Discussion
Acknowledgements
Materials
References
Biology
Aktin-Mikrotübül Kompozitlerinin Ayarlanabilir Motor Tahrikli Dinamiği ve Mekaniği ile Yeniden Yapılandırılması ve Karakterize Edilmesi
ERRATUM NOTICE
Important: There has been an erratum issued for this article. Read more …Bu yazıda, birlikte dolaşık aktin filamentleri ve mikrotübüllerinin ayarlanabilir üç boyutlu kompozit ağlarının mühendisliği ve karakterizasyonu için protokoller sunulmaktadır. Kompozitler, miyozin II ve kinesin motorları tarafından tahrik edilen aktif yeniden yapılanma ve balistik harekete uğrar ve aktin, mikrotübüller, motor proteinler ve pasif çapraz bağlayıcıların göreceli konsantrasyonları tarafından ayarlanır.
Yarı esnek aktin filamentlerinin ve sert mikrotübüllerin etkileşimli ağlarından oluşan kompozit sitoiskelet, göç, sitokinezi, yapışma ve mekanosensing gibi kilit süreçleri yürütmek için miyozin II ve kinesin gibi motor proteinleri kullanarak kuvvetleri yeniden yapılandırır ve üretir. Aktin-mikrotübül etkileşimleri, sitoiskeletin çok yönlülüğü ve uyarlanabilirliğinin anahtarı olsa da, miyozin ve kinesin aktivitesi ile etkileşimlerinin anlaşılması hala yenidir. Bu çalışma, miyozin II ve kinesin motorları tarafından tahrik edilen ve aktin, mikrotübüller, motor proteinleri ve pasif çapraz bağlayıcıların göreceli konsantrasyonları tarafından ayarlanan, aktif yeniden yapılanma ve balistik harekete geçirilen ko-dolaşık aktin filamentlerinin ve mikrotübüllerin ayarlanabilir üç boyutlu kompozit ağlarının nasıl tasarlanacağını açıklamaktadır. Çok spektral konfokal görüntüleme kullanarak kompozit yeniden yapılanmayı ve hareketi en etkili şekilde görselleştirmek için mikrotübüllerin ve aktin filamentlerinin floresan etiketlemesi için protokoller de detaylandırılmıştır. Son olarak, denge dışı yapıyı, dinamiği ve mekaniği nicel olarak karakterize etmek için kullanılabilecek veri analizi yöntemlerinin sonuçları sunulmaktadır. Bu ayarlanabilir biyomimetik platformun yeniden oluşturulması ve araştırılması, bağlı motor aktivitenin, kompozit mekaniğin ve filament dinamiklerinin mitozdan polarizasyona ve mekano-duyuma kadar sayısız hücresel sürece nasıl yol açabileceğine dair değerli bilgiler sağlar.
Hücre iskeleti, hücrelere yapısal ve mekanik destek sağlayan etkileşimli biyopolimerlerin dinamik bir kompozit ağıdır. İlişkili moleküler motorlar ve bağlayıcı proteinler, hücrelerin büyümesine, şeklini değiştirmesine, sertleşmesine, hareket etmesine ve hatta kendi kendini iyileştirmesine izin vermek için sitoiskeleti yeniden yapılandırır ve uyarlar, göç ve bölünmeden mekanosensing'e kadar sayısız hücresel süreci mümkün kılar 1,2. Hücresel biyofizikteki öneminin ötesinde, sitoiskelet aynı zamanda yara iyileşmesi ve ilaç dağıtımından filtrasyona ve yumuşak robotik 1,3,4,5,6,7,8,9'a kadar değişen potansiyel malzeme uygulamaları ile aktif maddenin mükemmel bir örneğidir.
Hücre iskeletine benzersiz yapısal ve mekanik çeşitliliği ve çok işlevliliği kazandıran iki temel özellik şunlardır: 1) yarı esnek aktin filamentleri ve sert mikrotübüller gibi çoklu etkileşimli protein filamentlerinin yanı sıra bunlarla ilişkili bağlanma ve çapraz bağlanma proteinlerini içeren kompozit doğası 3,5,10; ve 2) filamentli proteinleriiterek ve çekerek, miyosinler ve kinezinler gibi enerji tüketen motorlar aracılığıyla sürekli olarak yeniden yapılandırma, hareket etme, kabalaştırma ve iş yapma yeteneği 1,7,11,12,13. Bu zarif karmaşıklık, hücre iskeletinin hücre hareketliliği, sitokinezi ve yara iyileşmesi 3,6,7,11 gibi çeşitli süreçlere aracılık etmesini sağlarken, araştırmacıların in vitro sistemlerde hücre iskeletinin imzasını in vivo özelliklerini yeniden üretme yeteneğini engellemektedir.
Mevcut sınır sulandırma çabaları, dolaşık ve çapraz bağlı aktin filamentlerinin ve mikrotübüllerinin 3,10,14,15,16,17, kuvvet üreten aktomiyozin ağlarının 2,8,18,19,20,21 kompozitlerine ve kinesin-mikrotübül tarafından tahrik edilen aktif nematiklere odaklanmaktadır. etkileşimler 22,23,24,25,26. Kararlı hal aktin-mikrotübül kompozitlerinin, tek bileşenli sistemlere kıyasla gelişmiş filament hareketliliği ve artan sertlik gibi15,16,27 gibi acil mekanik özellikler gösterdiği gösterilmiştir 27. İn vitro aktomiyozin sistemleri üzerine yapılan çalışmalar, aktin, miyozin ve çapraz bağlayıcıların konsantrasyonlarına bağlı olarak çok çeşitli yapısal ve dinamik özellikler bildirmiştir 28,29,30,31. Örneğin, yeterli çapraz bağlama ile, aktomiyozin ağları büyük ölçekli büzülmeye ve kabalaşmaya maruz kalır 2,28,30,32,33,34,35,36, çapraz bağlayıcılar olmadan, ağlar hızlı, istikrarsızlaştırıcı akış ve kopma gösterir 19,29 . Mikrotübül demetlerini çapraz bağlamak ve çekmek için kinesin motor kümelerini kullanan yeniden yapılandırılmış mikrotübül bazlı aktif nematiklerin, uzun süreli türbülanslı akışlar, uzatma, burkulma, kırılma ve iyileşme sergilediği bildirilmiştir 12,22,23,24,25,37,38,39,40,41, 42,43,44,45,46,47.
Daha yakın zamanlarda, miyozin II mini-filamentleri tarafından tahrik edilen aktin-mikrotübül kompozitlerinin, çapraz bağlayıcıları olmayan aktomiyozin ağlarının 17,26,48 sergilediği düzensiz akış ve ağ kopmasına kıyasla daha düzenli büzülme ve ağ bütünlüğüne yol açtığı gösterilmiştir. Ayrıca, kompozit sağlamlık ve kuvvet oluşturma kombinasyonu, aktin ve mikrotübüller karşılaştırılabilir konsantrasyonlarda mevcut olduğunda optimize edilir. Formülasyon uzayının bu bölgesinde ortaya çıkan temel özellikler arasında gelişmiş mekanik mukavemet 26, aktin ve mikrotübüllerin koordineli hareketi26, sürekli sürekli büzülme ve mezoölçekli yeniden yapılanma17 bulunmaktadır.
Burada, sırasıyla aktin filamentleri ve mikrotübülleri üzerinde etkili olan miyozin II mini filamentleri ve kinesin kümeleri tarafından dengenin dışına itilen mikrotübüllerin ve aktin filamentlerinin birlikte dolaşık ve çapraz bağlı kompozitlerini tasarlamak ve ayarlamak için protokoller tanımlanmıştır (Şekil 1). Bu kompozit sınıfının dinamikleri, yapısı ve mekaniği, filamentlerin, motorların ve çapraz bağlayıcıların nispi konsantrasyonları tarafından, advektif ve türbülanslı akış, izotropik büzülme, hızlanma, yavaşlama, de-karıştırma, sertleşme, gevşeme ve kopma gibi zengin bir faz alanı sergilemek üzere ayarlanabilir. Bu çalışmanın odak noktası, bu aktif sitoiskelet kompozitleri sınıfının hazırlanması ve ayarlanmasıdır. Bununla birlikte, araştırmacıların tarif edilen aktif kompozitleri kıyaslamalarına ve karakterize etmelerine yardımcı olmak için, multi-spektral konfokal mikroskopi kullanan etkili görüntüleme yöntemleri de detaylandırılmıştır. Son olarak, kompozitlerin dinamiklerini, yapısını ve mekaniğini ölçmek için kullanılabilecek temel hesaplamalı analiz yöntemlerinin sonuçları sunulmaktadır. Araştırmacılar, diferansiyel dinamik mikroskopi (DDM), uzamsal görüntü otokorelasyonu (SIA) ve parçacık görüntü velosimetrisini (PIV) içeren bu yöntemleri benimsemeye teşvik edilmektedir, çünkü kompozitlerin karmaşık dinamiklerini ve yapısal çeşitliliğini karakterize etmek için optimize edilmiştir 17,26,49.
Aşağıda açıklanan adımlar, kompozitlerin hazırlanmasına ve konfokal mikroskopi kullanılarak görüntülenmesine odaklanmaktadır. Elde etme sonrası veri analizini ve optik cımbız ölçümlerini açıklayan protokoller önceki çalışmalarda bulunabilir 17,26,48,50 ve talep üzerine sağlanabilir. Tüm malzemeler sağlanan Malzeme Tablosunda listelenmiştir.
1. Proteinlerin oda yüzeylerine adsorpsiyonunu önlemek için silanjörlü kapaklar ve mikroskop slaytları hazırlayın
NOT: Bu 2 günlük bir işlemdir. Silanize slaytlar kullanımdan 1 ay öncesine kadar hazırlanabilir.
- 1 numaralı kapak kapaklarını (24 mm x 24 mm) ve mikroskop slaytlarını (1 inç x 3 inç) plazma temizleyiciye sığacak şekilde belirlenmiş bir rafa yerleştirin. Rafı plazma temizleyiciye yerleştirin ve 20 dakika çalıştırın.
- Kapakları ve kızakları yalnızca silan ile kullanılmak üzere tasarlanmış yeni bir rafa aktarın ve aşağıda açıklandığı gibi camları temizlemek için rafı cam kaba yerleştirin.
- Kapakları ve kızakları 1 saat boyunca% 100 asetona batırın. Kapakları ve slaytları 10 dakika boyunca %100 etanol içine daldırın.
- Kapakları ve kaydırakları 5 dakika boyunca deiyonize suya (DI) batırın. Temizleme adımlarını iki kez daha tekrarlayın.
- Kapakları ve slaytları taze hazırlanmış 0,1 M KOH'a 15 dakika bekletin. Kapakları ve slaytları 5 dakika boyunca taze DI'ye batırın. Bu adımı iki kez daha tekrarlayın.
- Hava kuru kapaklar 10 dakika boyunca kayar ve kaydırılır. Aşağıda açıklandığı gibi hidrofobik yüzeyler üretmek için temizlenmiş kapaklara ve kızaklara silan uygulayın.
NOT: Bir duman davlumbazında aşağıdaki adımları tamamlayın.- Kurutulmuş örtüleri ve slaytları 5 dakika boyunca% 2 silan (toluen içinde çözülmüş) içine batırın. Beş defaya kadar yeniden kullanmak üzere belirlenen şişeye silan dökmek için bir huni kullanın.
- Kapakları ve slaytları 5 dakika boyunca %100 etanol içine daldırın. Etanol'ü taze etanol ile değiştirin. Kapakları ve slaytları 5 dakika boyunca daldırın.
- Kapakları ve slaytları 5 dakika boyunca taze DI'ye batırın. Her seferinde taze etanol ve DI kullanarak etanol ve DI yıkama adımını iki kez daha tekrarlayın. Hava kuru kapaklar 10 dakika boyunca kayar ve kaydırılır.
2. Miyozin mini filamentleri tarafından tahrik edilen aktif aktin-mikrotübül kompozitinin hazırlanması
- Aktif olmayan miyozini aktin filament bağlama yoluyla çıkarın ve aşağıda açıklandığı gibi ultrasantrifüjleme yoluyla aşağı çekme işlemi gerçekleştirin.
- Aktin'i filamentlere polimerize edin. Hassas bir mikropipet ve steril pipet uçları kullanarak bir mikrosantrifüj tüpünde birleştirin: 1,87 μL DI, 1,3 μL 10x G-tampon, 1,3 μL 10x F-tampon, 1,63 μL 4 M KCl, 4,53 μL aktin (47,6 μM) ve 1,08 μL 100 μM falloidin.
NOT: Yeterli polimerizasyonu sağlamak için, aktin konsantrasyonu ve aktin:faloidin molar oranı sırasıyla 18.4 μM ve 2:1 olmalıdır. - Karıştırmak için çözeltiyi yavaşça yukarı ve aşağı doğru pipetleyin ve ardından karanlıkta ≥1 saat boyunca buz üzerine koyun. 4 °C'ye kadar serin ultrasantrifüj. Miyozin aliquot'u -80 ° C'den çıkarın ve buz üzerine koyun.
NOT: Aktin polimerize olurken bu noktada adım 2.2'yi tamamlayın. - ≥1 saat aktin polimerizasyonundan sonra, polimerize aktine 1.3 μL 10 mM ATP ve 2 μL 19 μM miyozin ekleyin.
NOT: Aktin:miyozin molar oranı, inaktif miyozin motorlarının (yani ölü kafaların) yeterli şekilde çıkarılmasını sağlamak için >5 olmalıdır. - Karıştırmak için çözeltiyi yavaşça yukarı ve aşağı doğru pipetleyin. Ultra santrifüj sınıfı bir tüpe aktarın.
30 dakika boyunca 4 °C ve 121.968 x g'de santrifüj.
- Aktin'i filamentlere polimerize edin. Hassas bir mikropipet ve steril pipet uçları kullanarak bir mikrosantrifüj tüpünde birleştirin: 1,87 μL DI, 1,3 μL 10x G-tampon, 1,3 μL 10x F-tampon, 1,63 μL 4 M KCl, 4,53 μL aktin (47,6 μM) ve 1,08 μL 100 μM falloidin.
- Aşağıda açıklandığı gibi aktin filamentleri ve mikrotübüllerin birlikte dolaşık kompozit ağını hazırlayın.
NOT: Miyozin spin-down'dan 30 dakika önce başlayın (adım 2.1.4).- Bir ısı bloğunu 37 °C'ye ayarlayın. Bir mikrosantrifüj tüpüne aşağıdakileri eklemek için hassas bir mikropipet ve steril pipet uçları kullanın: 13,9 μL PEM, 3 μL% 1 Tween20, 1,55 μL 47,6 μM aktin, 0,36 μL 34,8 μM R-aktin, 0,3 μL 250 mM ATP, 0,87 μL 100 μM falloidin, 1,91 μL 5-488-tübülin, 0,3 μL 100 mM GTP, ve 0.75 μL 200 μM Taksol, toplam 23 μL hacme kadar.
NOT: Listelenen aktin ve tübülin konsantrasyonları, 2.9 μM aktin ve 2.9 μM tübülin içeren bir kompozit içindir. Toplam protein konsantrasyonu c = c A + c T = 5.8 μM ve molar aktin fraksiyonu cA / (c A + cT) = Φ A = 0.5'tir. Bu değerleri ayarlamak için adım 2.5'e bakın. - Karıştırmak ve 1 saat boyunca ışıktan korunan 37 °C'lik bir ısı bloğuna yerleştirmek için çözeltiyi yavaşça yukarı ve aşağı pipetleyin.
- Bir ısı bloğunu 37 °C'ye ayarlayın. Bir mikrosantrifüj tüpüne aşağıdakileri eklemek için hassas bir mikropipet ve steril pipet uçları kullanın: 13,9 μL PEM, 3 μL% 1 Tween20, 1,55 μL 47,6 μM aktin, 0,36 μL 34,8 μM R-aktin, 0,3 μL 250 mM ATP, 0,87 μL 100 μM falloidin, 1,91 μL 5-488-tübülin, 0,3 μL 100 mM GTP, ve 0.75 μL 200 μM Taksol, toplam 23 μL hacme kadar.
- Aşağıda açıklandığı gibi konfokal görüntüleme deneyleri için örnek odalar hazırlayın.
NOT: Bekleme süreleri boyunca 2.1.4 ve 2.2.2 adımlarını tamamlayın.- İki silanize slaytı bir sıcak plaka üzerine yan yana yerleştirin (kapalı), slaytlar boyunca ~ 3 mm aralıklarla iki termoplastik sızdırmazlık filmi şeridi yerleştirin ve bir örnek odası oluşturmak için termoplastik sızdırmazlık filminin üzerine iki silanize kapak kayması yerleştirin.
- Kapaklar erimiş termoplastik sızdırmazlık filmi (~1-2 dakika) ile kızaklara sıkıca bağlanana kadar sıcak plakayı düşük ayarda çevirin. İki yüzey arasında ~100 μm aralığı korurken yapışmayı sağlamak için eşit basınçla bastırın.
- Odaları çıkarın ve sıcak plakayı kapatın. Etiket odaları (+) ve (-) ile. (+) odası aktif numune (miyozlu) için olacak ve (-) odası kontrol (miyozun içermeyen) olacaktır. Her odanın ≤10 μL sıvı barındırabildiğinden emin olun.
- Aşağıda açıklandığı gibi örnekleri görüntüye hazırlayın.
NOT: Adım 2.1 ve 2.2 tamamlandıktan hemen sonra bu adımı tamamlamak önemlidir.- Miyosin-aktin örneğini ultrasantrifüjden dikkatlice çıkarın (adım 2.1.4) ve hemen süpernatantın üst 7.5 μL'sini pipetleyin ve yeni bir mikrosantrifüj tüpüne aktarın.
- Aktin-mikrotübül örneğini ısı bloğundan çıkarın ve 1.5 μL 10x D-Glikoz, 1.5 μL 10x GOC ve 1.5 μL 1 mM blebbistatin ile hafifçe karıştırın. Çözeltiyi iki adet 13,7 μL alikotaya bölün ve (+) ve (-) olarak etiketleyin.
- Süpernatantın 1.28 μL'sini adım 2.4.1'den (+) aliquot'a karıştırın. 1.28 μL DI ile (-) aliquot'a karıştırın. Her bir çözeltiyi kılcal hareket yoluyla karşılık gelen odaya (adım 2.3) yavaşça akıtın. Kanala hava kabarcıkları sokmamaya dikkat edin.
- Her kanalın iki açık ucunu hızlı kuruyan epoksi veya UV yapıştırıcı ile kapatın. Mikroskopa yerleştirmeden önce yapıştırıcının tamamen kuru olduğundan emin olun. 3. adımda açıklandığı gibi hemen görüntüleyin.
NOT: UV yapıştırıcı avantajlıdır çünkü UV ışınlarına maruz kaldığında neredeyse anında kürlenir. Bununla birlikte, blebbistatin UV ışınlarına duyarlı olduğundan, blebbistatinin devre dışı bırakılmasını önlemek için tutkalı (numune odasının kenarlarında) küçük bir UV çubuğu kullanarak sadece yerel olarak aydınlatmak önemlidir.
- İsteğe bağlı: Kompozitlerin dinamiklerini ve yapısını ayarlamak için protein konsantrasyonlarını değiştirin.
NOT: İstenirse aktin, mikrotübüller ve miyozin konsantrasyonlarını değiştirmek için yukarıdaki adımlarda aşağıdaki adımlarda değişiklikler yapılması önerilmektedir.- Adım 2.2.1 ve 2.4.3'teki aşağıdaki değişiklikler dışında yukarıda açıklanan adımları izleyin.
- Aktin ve mikrotübüllerin konsantrasyonlarını değiştirmek, böylece c ve ΦA'yı ayarlamak için, adım 2.2.1'de kullanılan aktin, R-aktin ve 5-488-tübülin hacmini istediğiniz gibi artırın veya azaltın26. Aktin konsantrasyonunu değiştirirken, aktin ile aynı molar oranları korumak için R-aktin ve faloidin molar konsantrasyonlarını orantılı olarak ayarlayın. PEM hacmini, karışımın son hacmi 23 μL kalacak şekilde ayarlayın. Diğer tüm bileşen hacimleri ve konsantrasyonları aynı kalır.
- Miyozin konsantrasyonunu değiştirmek için, adım 2.4.3'te (+) alikot'a eklenen miyozin hacmini istediğiniz gibi ayarlayın. (-) aliquot'a eklenen DI ses seviyesini buna göre ayarlayın. Adım 2.2.1'deki PEM hacmini, miyozin (+) ve DI (-) hacmindeki artışı veya azalmayı hesaba katacak şekilde ayarlayın ve her numunenin ((+) ve (-)) son hacminin 14.98 μL olmasını sağlayın.
3. Konfokal mikroskopi kullanılarak aktif kompozitlerin görüntülenmesi ve karakterizasyonu
- Adım 2'de hazırlanan aktomiyosin-mikrotübül kompozitlerini görüntülemek için, 60x 1.4 NA yağ daldırma hedefine sahip bir lazer taramalı konfokal mikroskop (LSCM) veya benzeri bir mikroskop kullanın. Aktin filamentlerini ve mikrotübülleri ayrı floresan kanallarında aynı anda görselleştirmek için, 565/591 nm uyarma/emisyon filtreli 561 nm lazer ve 488/525 nm uyarma/emisyon filtreli 488 nm lazer kullanın.
- Numune odasını mikroskop üzerine, kontrol kanalı doğrudan hedefin üzerine yerleştirilecek şekilde yerleştirin. Hedef ve kapak kayması arasında bir yağ arayüzü olduğundan emin olun.
- Kontrol kompozitini odaklamak için sahne alanı kontrollerini kullanın, ardından numune odasının her iki yüzeyini de bulun. Z konumunu numune odasının merkezine taşıyın. Şekil 2'de gösterildiği gibi berrak filamentli ağların varlığını kontrol edin.
- Kontrol odasını hala görselleştirerek, aktin filamentlerinin ve mikrotübüllerin eşzamanlı olarak görselleştirilmesine izin vermek için her lazerin yoğunluğunu ayarlayın. Fotobeyazlatmayı (aktin kanalında daha yaygın) ve kanamayı (tipik olarak mikrotübüllerden aktin kanalına) önlemek için mümkün olan en düşük lazer yoğunluğunu koruyun.
- Etkin olmayan kontrol örneğini karakterize etmek için toplam 1000 kare için 2,65 fps'de 256 x 256 piksel kare (213 μm x 213 μm) görüntüden ≥oluşan üç zaman serisi (video) toplayın. Her zaman serisini, numune odasının ≥500 μm ile ayrılmış farklı bir bölgesinde toplayın. Minimum algılanabilir hareket olduğundan ve akış veya yeniden yapılandırma olmadığından emin olun.
- 488 nm lazeri kapatın ve (+) bölmesine gitmek için sahne alanı kontrollerini kullanın.
- 568 nm lazeri kullanarak, uygun ağ oluşumunu sağlamak için (+) kanaldaki mikrotübülleri görselleştirin (Şekil 2) ve numune odasının eksenel merkezini tanımlayın (kontrol odasının merkez z-konumundan farklı olabilir).
- 488 nm lazeri açın ve aşağıdaki değişikliklerle yukarıdaki 3.5. adımı tekrarlayın. 45 dakikaya kadar zaman serileri toplayın, numune görüş alanının dışına çıktığında, yırtılmalarda veya fotoağartıcılarda alımı durdurun. 5-10 zaman serisi kaydedin ve her zaman serisinin ilk zaman serisinin başlangıcına göre başladığı zamanı takip edin.
- DDM, SIA ve PIV kullanarak verileri Şekil 3, Şekil 4, Şekil 5 ve daha önce 17,48,50,51'de açıklandığı gibi analiz edin.
NOT: 488 nm lazer, blebbistatini devre dışı bırakarak miyozin ATPaz aktivitesini lokal olarak aktive eder, bu nedenle yalnızca veri toplamanın başlangıcında, t = 0 zaman serisinin başlangıcında olacak şekilde açılmalıdır. Bu edinme parametreleri, daha önce yapıldığı gibi diferansiyel dinamik mikroskopi (DDM) analizi için optimize edilmiştir26.
4. Kinesin motorları tarafından tahrik edilen aktif aktin-mikrotübül kompozitlerinin hazırlanması
NOT: Aşağıdaki adımlar, kinesin motorları veya kinesin ve miyozin50'nin bir kombinasyonu tarafından dengeden çıkarılan aktin-mikrotübül kompozitleri oluşturur.
- Kinesin ve miyozin motorlarını aşağıda açıklandığı gibi hazırlayın.
- Miyozin içeriyorsa, adım 2.1'i izleyin.
- Mikrotübül çiftleri arasında bağlanan ve kuvvet uygulayan kinesin motor kümeleri oluşturmak için, steril bir 1,5 mL mikrosantrifüj tüpüne aşağıdakileri eklemek için bir mikropipet ve steril pipet uçları kullanın: 1,16 μL PEM, 2,74 μL 8,87 μM kinesin dimer, 7,29 μL 83,3 μM NeutrAvidin, 0,81 μL 2mM DTT. Çözeltiyi yukarı ve aşağı pipetleyerek, hafifçe karıştırın ve 4 °C'de 30 dakika boyunca ışıktan korunan inkübe edin (siyah bir mikrosantrifüj tüpü kullanın veya folyoya sarın).
NOT: Kinesin dimerlerinin NA'ya molar oranı 1:25'tir.
- Örnek odaları hazırlamak ve iki yerine üç oda yapmak için adım 2.3'ü izleyin. Bu adımı kinesin inkübasyonu (adım 4.1.2) ve miyozin ultrasantrifüjleme (adım 4.1.1) sırasında gerçekleştirin.
- Aktin filamentleri ve mikrotübüllerinin birlikte dolaşık kompozit ağını hazırlayın.
- Isı bloğunu 37 °C'ye ayarlayın. Steril 1,5 mL mikrosantrifüj tüpüne aşağıdakileri eklemek için bir mikropipet ve steril pipet uçları kullanın: 3,21 μL PEM, 4,5 μL %1 Tween20, 2,18 μL 47,6 μM aktin, 3,46 μL 5-R-tübülin, 4,5 μL 100 mM ATP, 4,5 μL 10 mM GTP, 1,13 μL 200 μM Taxol ve 1,57 μL 20 μM 488-faloidin. Toplam hacmin 25 μL olduğundan emin olun.
- Karıştırmak ve ışıktan korunan 37 °C ısı bloğuna 1 saat boyunca yerleştirmek için çözeltiyi yavaşça yukarı ve aşağı doğru pipetleyin. Tüpü ısı bloğundan çıkarın ve 0,84 μL 100 μM falloidin ile hafifçe karıştırmak için bir mikropipet kullanın. Oda sıcaklığında 5-10 dakika boyunca inkübe edin, ışıktan koruyun.
NOT: Adım 4.3.1 yerine bu adımda falloidin eklenmesi, aktin filamentlerinin floresan etiketlemesini iyileştirir, çünkü 488-faloidin, aktin bağlanma bölgeleri için etiketlenmemiş faloidin ile rekabet etmek zorunda değildir.
- Konfokal görüntüleme için aktif kompozitler hazırlayın.
- 4.3.2. adımdan itibaren çözeltiye 1,13 μL 200 μM blebbistatin, 1,35 μL 10x Glu ve 1,35 μL 10x GOC'yi ekleyin ve yukarı ve aşağı pipetleyerek yavaşça karıştırın. Çözeltiyi üç adet 10 μL alikota bölün ve (K), (K+M) ve (-) olarak etiketleyin.
- Adım 2.1.4'ten (K + M) alikotuna kadar 2.54 μL miyozin ile karıştırın. 2.54 μL PEM'de (K) ve (-) alikotlara karıştırın.
- Adım 4.1.2'den (K) ve (K+M) alikotlarına kadar 2,5 μL kinesin kümeleri eklemek için bir mikropipet ve steril pipet uçları kullanın. Pipet yukarı ve aşağı karıştırmak için. Aynı tekniği kullanarak 2,5 μL PEM'de (-) karıştırın.
NOT: Listelenen aktin ve tübülin konsantrasyonları, 2.32 μM aktin ve 3.48 μM tübülin içeren bir kompozit içindir. Toplam protein konsantrasyonu c = c A + c T = 5.8 μM ve molar aktin fraksiyonu cA / (c A + cT) = Φ A = 0.4'tür. Kinesin ve miyozin konsantrasyonları sırasıyla 0.35 μM ve 0.47 μM'dir. c A, cT, c ve ΦA'yı ayarlamaya yönelik genel yönergeler için adım 2.5'e bakın. - Bir mikropipet kullanarak, her bir çözeltiyi kılcal etki yoluyla hazırlanan numune odalarının ilgili kanalına (adım 4.2) yavaşça akıtın. Kanala hava kabarcıkları sokmamak için pipet üzerinde çok yavaş ve nazikçe aşağı doğru itin.
- Her kanalın iki açık ucunu hızlı kuruyan epoksi veya UV kürlenebilir yapıştırıcı ile kapatın. Mikroskopa yerleştirmeden önce yapıştırıcının tamamen kuru olduğundan emin olun.
NOT: Kinezinin izlenmeden hareket ettiği süreyi en aza indirmek için bu adımın hızlı bir şekilde yapılması önemlidir. Bu nedenle 1 dakikada (5 veya 10 dakika yerine) kürlenen epoksi önerilir. UV kürlenebilir yapıştırıcı bu konuda avantajlıdır, çünkü UV ışınlarına maruz kaldığında neredeyse anında kürlenir.
- Aşağıdaki önemli değişiklikler dışında, 3. adımı hemen takip eden görüntü örnekleri hazırlanmıştır. Kinesin ışık aktivasyonu ile kontrol edilmediğinden, adım 4.4.3'ten hemen sonra çalışmaya başlar, bu nedenle bu süreyi t = 0 olarak işaretleyin. Kompoziti ilk etkin olmayan duruma (t = 0) mümkün olduğunca yakın görüntülemek için, önce (K) ve (K+M) kanallarını görüntüleyin ve adım 4.4.3 ile veri toplamanın başlangıcı (adım 3.8) arasında geçen süreyi not edin. Uygulamada, geçen bu süre ~ 5 dakikadır.
5. Pasif çapraz bağlayıcıların aktif kompozitlere dahil edilmesi
NOT: Bu adımlarda, adım 4'te açıklanan aktif kompozitlerde biyotinile aktin ve tübülin alt birimlerinin ve NeutrAvidin'in (NA) aktinin aktine (A-A) veya mikrotübüllerin mikrotübüllere (M-M) pasif olarak çapraz bağlanması için nasıl kullanılacağı açıklanmaktadır.
- A-A veya M-M çapraz bağlayıcı komplekslerini biyotinile proteinler (biyotin-aktin veya biyotin-tübülin), NA ve biyotin ile 2: 2: 1 biyotin-aktin / tübülin: biyotin: NA oranında hazırlayın. Adım 4'ten önce bu işlemi başlatın.
- A-A çapraz bağlayıcılar için, bir mikrosantrifüj tüpüne 2 μL 11,6 μM biyotin-aktin, 1,39 μL 8,33 μM NA, 2,27 μL 1,02 μM biyotin ve 4,34 μL PEM eklemek için bir mikropipet ve steril pipet uçları kullanın. Yukarı ve aşağı pipetle çekerek hafifçe karıştırın.
- M-M çapraz bağlayıcılar için, bir mikrosantrifüj tüpüne 1,86 μL 4,55 μM biyotin-tübülin, 1,11 μL 8,33 μM NA, 1,82 μL 1,02 μM biyotin ve 5,21 μL PEM eklemek için bir mikropipet ve steril pipet uçları kullanın. Yukarı ve aşağı pipetle çekerek hafifçe karıştırın.
- Su geçirmez bir sızdırmazlık oluşturmak için boruları adım 5.1.1 ve/veya 5.1.2'den termoplastik sızdırmazlık filmine sarın. 4 ° C'ye ayarlanmış sıcaklık kontrollü bir sonikatör banyosunda yüzdürme salına yerleştirin.
- 90 ° C'de 4 dakika boyunca sonikat. Uygulamada, sonikatöre soğuk bir odaya koymak ve düşük sıcaklığı korumak için sonikasyon banyosuna buz paketleri eklemek en iyisidir.
- Çapraz bağlayıcı komplekslerini görüntüleme için örneklere dahil etmek üzere adım 4.3'ü izleyin ve aşağıda açıklandığı gibi adım 4.3.1'i A-A çapraz bağlama (adım 5.2.1) veya M-M çapraz bağlama (adım 5.2.2) için değiştirin.
- A-A çapraz bağlama için, aşağıdakileri bir mikrosantrifüj tüpünde birleştirin: 1.94 μL PEM, 4.50 μL% 1 Tween20, 2.18 μL 47.6 μM aktin, 3.46 μL 45.5 μM 5-R-tübülin, 1.13 μL A-A çapraz bağlayıcılar (adım 5.1.1), 4.50 μL 100 mM ATP, 4.50 μL 10 mM GTP, 1.13 μL 200 μM Taksol, ve 1.57 μL 20 μM 488-falloidin. Toplam hacmin 25 μL olduğundan emin olun.
- M-M çapraz bağlama için, aşağıdakileri bir mikrosantrifüj tüpünde birleştirin: 1.97 μL PEM, 4.50 μL% 1 Tween20, 2.18 μL 47.6 μM aktin, 3.76 μL 45.5 μM 5-R-tübülin, M-M çapraz bağlayıcıların 1.13 μL seyreltilmesi (adım 5.1.2), 4.50 μL 100 mM ATP, 4.50 μL 10 mM GTP, 1.13 μL 200 μM Taksi, ve 1.57 μL 20 μM 488-falloidin. Toplam hacmin 25 μL olduğundan emin olun.
- Bir çapraz bağlayıcı için spesifik konsantrasyonlarla 4.3.2-4.5 adımlarını izleyin: RA = 0.02'nin aktin molar oranı ve çapraz bağlayıcı: RT = 0.005'in tübülin molar oranı. Bu R A veR T değerleri, aktin filamentleri ve mikrotübüller (d A 60 nm ve d MT 67 nm) boyunca çapraz bağlayıcılar arasında benzer uzunluklarla sonuçlanır, dA
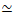 = I monomer / 2RA kullanılarak tahmin edilir, burada I monomer bir aktinmonomerinin uzunluğudur ve dMT = Iring / 26R T, buradahalka, 13 tübülinlik bir halkanın uzunluğudur 15, 17.
= I monomer / 2RA kullanılarak tahmin edilir, burada I monomer bir aktinmonomerinin uzunluğudur ve dMT = Iring / 26R T, buradahalka, 13 tübülinlik bir halkanın uzunluğudur 15, 17.
Aktif kompozitlerin başarılı bir şekilde hazırlanmasını belirlemek (Şekil 1) ve dinamiklerini ve yapılarını karakterize etmek için, aktin filamentlerini ve mikrotübüllerini aynı anda görselleştirmek için en az iki floresan kanallı bir lazer taramalı floresan mikroskobu kullanılır (Şekil 2 ve Şekil 6). Kompozitlerdeki tüm aktin filamentleri ve mikrotübülleri, in vitro çalışmalarda sıklıkla yapıldığı gibi, izleyici parlak filamentlerde doping yapmak yerine, seyrek olarak etiketlenir. Bu yöntem, ölçülen dinamiklerin ve yapının, kompozitlerden farklı koşullar altında oluşan izleyicilerden ziyade kompozitin kendisini temsil etmesini sağlar. Bu nedenle, bireysel aktin filamentleri ve mikrotübülleri tipik olarak çözülemez, bunun yerine görüntüler mezoölçekli ağ yapısını tasvir eder (Şekil 2 ve Şekil 6).
Bu etiketleme yaklaşımı, karşılıklı Fourier uzayındaki dinamikleri ve yapıyı inceleyen uzamsal görüntü otokorelasyonu (SIA) ve diferansiyel dinamik mikroskopi (DDM) analizleri için optimize edilmiştir (Şekil 4, Şekil 5 ve Şekil 8)52,53,54,55. Parçacık görüntü velosimetrisi (PIV), dinamikleri ve akış alanlarını (Şekil 3 ve Şekil 7) tasvir etmek ve karakterize etmek için de kullanılabilir, ancak yoğun, düşük sinyalli görüntülerdeki gürültüden kaynaklanan hatalı vektörleri ortadan kaldırmak için SIA ve DDM'den daha büyük gecikme süresi artışları (daha düşük zamansal çözünürlük) ve piksel bağlama (daha düşük uzamsal çözünürlük) gerektirir. Bununla birlikte, akış alanlarının kalitatif incelemesi ve DDM sonuçlarının doğrulanması için PIV önerilmektedir (Şekil 4 ve Şekil 8)26,50.
Bu analizleri (yani, DDM, SIA, PIV) kullanarak tanımlanan ağların örneklem karakterizasyonu, araştırmacıların örneklerini karşılaştırmak ve karakterize etmek için benzer analizleri benimsemelerine yardımcı olmak için sağlanmıştır. Ancak bu tekniklerin detaylı açıklamaları bu çalışmanın kapsamı dışındadır. Kullanıcı dostu Python kodu da dahil olmak üzere bu ve diğer benzer sistemlerde DDM'nin nasıl gerçekleştirileceğine dair ayrıntılı açıklamalar için, önceki çalışmalara 17,26,49,50 ve içindeki referanslara bakın. Burada açıklanan sistemlerde SIA ve PIV'in nasıl gerçekleştirileceğine ilişkin ayrıntılar için, okuyucu öncekiçalışmalara yönlendirilir 17,50.
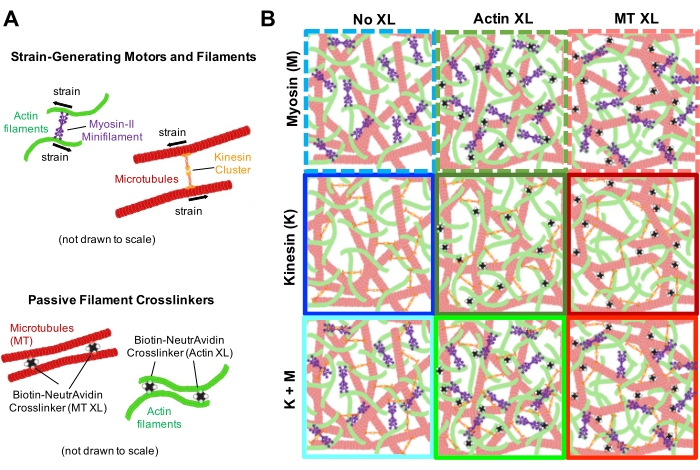
Kompozitlerin beklendiği gibi çalıştığından emin olmak için aşağıda açıklanan çeşitli kontroller yapılmalıdır. Miyozin veya kinesin içermeyen bir kompozit, minimum termal dalgalanmalar veya sürüklenme ile esasen statik görünmelidir. Aktin filamentleri ve mikrotübülleri, ~200 μm x 200 μm'lik bir görüş alanı boyunca aktin ve mikrotübüllerin minimum demetlenmesi, toplanması veya faz ayrımı ile birlikte dolaşmış ve homojen olarak dağılmış görünmelidir (Şekil 2, en solda)17. Miyozin içeren ancak 488 nm ışığa maruz kalmayan kompozitler için de benzer bir sonuç beklenmelidir (blebbistatini devre dışı bırakmak için).
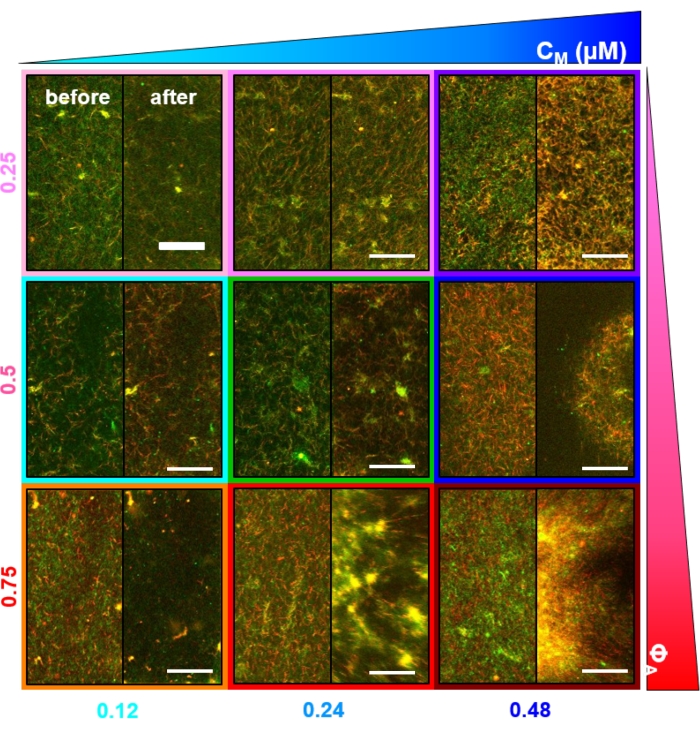
Miyozin dahil edilmesi ve 488 nm ışığa maruz kalınması üzerine, kompozitler, miyozin aktivitesinden önce ve sonra alınan mikroskop görüntülerinde görüldüğü gibi (Şekil 2) ve ayrıca aktivite sırasında değişen zamanlar için karşılık gelen PIV akış alanlarında (Şekil 3) görüldüğü gibi, aktin ve mikrotübüller için büyük ölçüde izotropik ve benzer bir kasılmaya uğrar. Hareketin balistik, difüzyon, subdifüzyon vb. olup olmadığını belirlemek için, DDM'den belirlenen karakteristik korelasyon süresi τ (q), dalga vektörünün (yani karşılıklı uzayın) bir fonksiyonu olarak değerlendirilir. Daha önce ayrıntılı olarak açıklandığı gibi bakınız 17,26,49. Şekil 4 ayrıca bu kompozitleri karakterize etmek için DDM'nin nasıl kullanılacağını da göstermektedir. Güç yasası ölçeklemesi τ(q)~1/vq β, β = 1 ile, v hızıyla balistik hareketi gösterir. Referans olarak, β = 2, v difüzyon katsayısı olan difüzyon dinamiklerini temsil eder. Tüm aktif kompozitler, aktin ve miyozin konsantrasyonları (Şekil 4B) tarafından ayarlanan hızlara sahip balistik ölçekleme (Şekil 4A) sergiler ve aktivite sırasında hızlanarak veya yavaşlayarak zaman içinde değişebilir (Şekil 4C, D).
Şekil 2'de görülebilen ve daha yüksek aktin ve miyozin konsantrasyonları için daha belirgin olan ağ yeniden yapılanması ve kümelenmesi, Şekil 5'te gösterildiği ve daha önce 17,48,50'de açıklandığı gibi SIA kullanılarak karakterize edilebilir. Kısaca, bir görüntüdeki özelliklerin karakteristik boyutunun bir ölçüsü olan ξ korelasyon uzunluğu, her bir uzamsal yoğunluk otokorelasyon eğrisi g(r)'nin pikseller arasındaki r uzaklığının üstel bir fonksiyonuna sığdırılmasıyla belirlenebilir. Daha uzun mesafeler boyunca devam eden daha büyük g(r) pikleri, daha büyük yapısal özellikleri gösterir (yani, bireysel filamentlerin paketlenmesi, kümelenmesi). Şekil 5'te gösterildiği gibi, daha yüksek aktin fraksiyonları ve miyozin konsantrasyonları için, önemli yeniden yapılanma ve agregasyon, zaman içinde ξ'deki artışa yansır.
Aktif kompozitlerin viskoelastik özellikleri ve doğrusal olmayan mekanik tepkisi, optik cımbız mikroreolojisi (OTM) kullanılarak da ölçülebilir. Bununla birlikte, bu deneyler için protokoller ve temsili sonuçlar bu çalışmanın kapsamı dışındadır. İlgilenen okuyucular, OTM ölçümlerinin nasıl yapılacağını ve beklenen sonuçları ayrıntılı olarak açıklayan önceki48,56 numaralı çalışmalara atıfta bulunulmaktadır.
Yukarıda açıklanan aynı deney ve analiz araçları programını kullanarak, aşağıdaki bölüm, kinesin motorları ve biotin-NA çapraz bağlayıcıları kompozitlere dahil edildiğinde dinamiklerin ve yapının nasıl değiştiğini açıklamaktadır (Şekil 6, Şekil 7 ve Şekil 8). Şekil 6 , aktin filamentlerinin veya mikrotübüllerinin pasif çapraz bağlanması (XL) ile ve olmadan, sadece kinesin (K) veya kinesin ve miyozin (K + M) tarafından tahrik edilen kompozitlerin temsili konfokal görüntülerini göstermektedir.
Kinezinin kompozitlere dahil edilmesi, başlangıçta Şekil 7'nin (Sınıf 1) en üst satırında görüldüğü gibi miyozin tahrikli kompozitlerle benzer dinamikler ve yeniden yapılanma ile sonuçlanır. Bununla birlikte, dinamikler tipik olarak büyük ölçekli anizotropik akışa (Şekil 7 orta sıra, Sınıf 2), hızlanmaya ve yavaşlamaya (Şekil 7 alt sıra, Sınıf 3) geçiş yapar. Bu özellikler 5-30 dakika sonra mezoölçek kümeleme ve toplama ile birleşir (Şekil 6 ve Şekil 8B). PIV tarafından oluşturulan akış alanları ve Şekil 7'de gösterilen zamansal renk haritaları, izotropik yeniden yapılanma (Sınıf 1, üst panel), yönlendirilmiş akış (Sınıf 2, orta paneller) ve çift yönlü ivme (Sınıf 3, alt paneller) örneklerini göstermektedir.
Aktivite sırasında değişen zaman noktalarında aktin ve mikrotübüllerin hızları, τ (q) eğrilerine uyumla belirlenir, hızlanmayı ve ardından çapraz bağlamaya bağlı olan yavaşlamayı gösterir (Şekil 8). Şekil 8'de de gösterildiği gibi, her iki motor protein de dahil edildiğinde, dinamikler aslında sadece kinesin kompozitlerinden daha yavaştır ve mezoölçekli akışın gecikmeli başlangıcı vardır. Myosin ayrıca aktivite süresince aktin ve mikrotübül ağlarının daha homojen bir şekilde iç içe geçmesini, ayrıca daha az toplama ve yeniden yapılandırmayı destekler. Bu etkiler Şekil 6'daki görüntülerde görülebilir ve miyozin varlığında genellikle daha küçük olan SIA aracılığıyla hesaplanan zamanla değişen korelasyon uzunlukları ile ölçülür (Şekil 8B).
Şekil 1. Çoklu kuvvet üreten motorlar ve pasif çapraz bağlayıcılar ile aktif aktin-mikrotübül kompozitlerinin tasarımı ve karakterizasyonu. (A) Aktin monomerleri ve tübülin dimerleri, aktin filamentlerinin (yeşil) ve mikrotübüllerin (kırmızı) birlikte dolaşık ağlarını oluşturmak için 0.73-11.6 μM'lik c A ve c Tmolar konsantrasyonlarında ve aktin Φ A = c A/ (cA + cT) = 0, 0.25, 0.5, 0.75 ve 1'in molar fraksiyonlarında kopolimerize edilir. Pasif çapraz bağlama, biyotinile aktin filamentlerini (Aktin XL) veya mikrotübülleri (MT XL) çapraz bağlayıcıda bağlamak için NA kullanılarak elde edilir: aktin ve mikrotübüller için sırasıyla R A = 0.01-0.08 ve RMT= 0.001-0.01 protein molar oranları. Miyosin-II mini filamentler (mor) ve kinesin kümeleri (turuncu), c M = 0.12 - 0.48 μM ve cK = 0.2 - 0.7 μM konsantrasyonlarında, kompozitleri kararlı durumdan çıkarmak için filamentleri itin ve çekin. (B) Formülasyon uzayının şeması. Myosin II mini filamentleri (M), kinesin kümeleri (K) veya her iki motor (K + M), pasif çapraz bağlayıcılar (XL yok), aktin-aktin çapraz bağlantıları (Actin XL) ve mikrotübül-mikrotübül çapraz bağlantıları (MT XL) içermeyen kompozitlere dahil edilmiştir. Tüm karikatürler ölçeğe göre çizilmez. Bu şeklin daha büyük bir versiyonunu görüntülemek için lütfen buraya tıklayın.
Şekil 2. Değişen miyozin konsantrasyonları c M ve molar aktin fraksiyonları Φ A olan miyozin güdümlü sitoiskelet kompozitlerinin iki renkli konfokal görüntülemesi. (A) 256 x 128 kare piksel (212 x 106 μm2) iki renkli konfokal mikroskopi görüntüleri, aktin filamentlerinin (yeşil) ve mikrotübüllerin (kırmızı) kompozitlerinin miyozin motor aktivitesi yoluyla nasıl yeniden düzenlendiğini göstermektedir. Kinesin motorları veya pasif çapraz bağlayıcılar mevcut değildir. Her panelde, 45 dakikalık miyozin aktivasyonunun başında (solda, öncesinde) ve sonunda (sağda, sonra) çekilen görüntüler (blebbistatini devre dışı bırakmak için 488 nm ışıkla aydınlatma yoluyla) gösterilir. Paneller, miyozinin molar konsantrasyonunu (cM) artırarak, soldan sağa doğru giderek ve aktinin (ΦA) molar fraksiyonunu artırarak, yukarıdan aşağıya doğru giderek sıralanır. Her paneli özetleyen renkler, Şekil 4 ve Şekil 5'te kullanılan renk kodlamasıyla eşleşir. Ölçek çubukları 50 μM'dir. Analiz için dinamikleri ve yapıyı en iyi şekilde yakalamak için 1-5 fps kare hızlarını, 50-250 μm kenarlı yatırım getirilerini ve büzülme ve yeniden düzenleme hızına bağlı olarak 5-45 dakikalık zaman serisi sürelerini kullanıyoruz. Önceki ve sonraki görüntülerin benzer göründüğü paneller, pembe, macenta ve camgöbeği panellerinde görüldüğü gibi minimum yeniden yapılanmayı gösterir. Artan heterojenlik ve parlak noktalama özelliklerinin varlığı ile kanıtlanan küçük ölçekli kümeleme, turuncu, yeşil ve kırmızı panellerde görülebilir. Düzgün bir şekilde küçülen bir ağ olarak görülen büyük ölçekli büzülme, mavi ve mor panellerde belirgindir. Bu şekil referans17'den değiştirilmiştir. Bu şeklin daha büyük bir versiyonunu görüntülemek için lütfen buraya tıklayın.
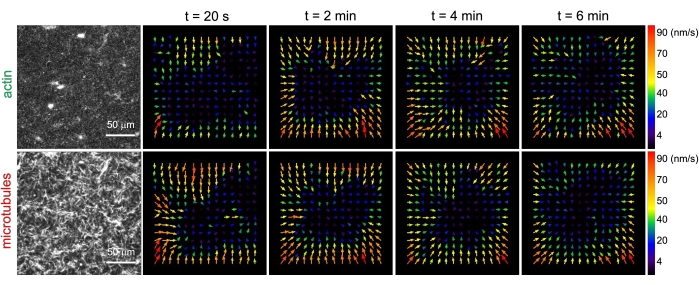
Şekil 3. Partikül görüntü velosimetrisi (PIV), aktomiyozin aktivitesinin, birlikte dolaşık kompozitlerde aktin ve mikrotübüllerin koordineli kontraktil dinamiklerini tetiklediğini göstermektedir. Miyozin tahrikli bir kompozitte aktin (üst sıra) ve mikrotübüller (alt sıra) için PIV akış alanları, 6 dakikalık bir zaman serisi boyunca artan zamanlarda (ΦA, cM) = (0.5, 0.24) ile. Akış alanları, Fiji/ImageJ PIV eklentisi kullanılarak 20 s gecikme süresi ve 2 piksel x 2 piksel gruplama ile oluşturulmuştur. Hem aktin hem de mikrotübüller, film süresi boyunca görüş alanının merkez bölgesine doğru yönlendirilmiş tutarlı bir hareket gösterir. Tüm görüntülerdeki ölçek çubukları 50 μm'dir. Farklı ok renkleri, vektör alanlarının sağındaki renk ölçeğinde belirtilen farklı hızlara karşılık gelir. Bu şekil referans26'dan değiştirilmiştir. Bu şeklin daha büyük bir versiyonunu görüntülemek için lütfen buraya tıklayın.
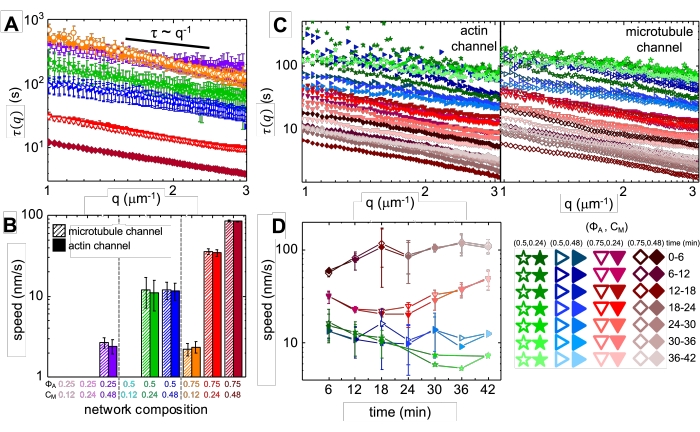
Şekil 4. Zaman çözümlü diferansiyel dinamik mikroskopi (DDM), aktif kompozitlerdeki aktin ve mikrotübüllerin hareket hızını ve türünü ölçer. (A) DDM, daha önce 17,26'da tanımlandığı gibi hem aktin (doldurulmuş semboller) hem de mikrotübüller (açık semboller) için karakteristik bozunma sürelerini τ vs dalga sayısı q'yu belirlemek için zaman serilerinin mikrotübül (üst, açık semboller) ve aktin (alt, dolgulu semboller) kanalları üzerinde gerçekleştirilir. Tüm eğriler, balistik hareketi gösteren τ ~ q-1 ölçeklendirmesini takip eder ve τ(q) = (vq)-1'e sığdırılarak belirlenen v hızları vardır. Daha yüksek hızlar, herhangi bir q için daha küçük τ (q) değerlerine karşılık gelir. Sembol renkleri ve şekilleri, B'de gösterilen (Φ A, cM) kombinasyonlarına karşılık gelir. (B) Büzülme hızları v, A'da gösterilen τ(q) eğrilerine uyarak belirlenir ve bunlar her 45 dakikalık zaman serisi boyunca tüm gecikme sürelerinin ortalaması alınır. (C) Zaman çözümlü DDM (trDDM), 45 dakikalık aktivasyon süresi boyunca ardışık 6 dakikalık aralıklarla (aynı rengin farklı tonlarıyla gösterilir) aktin (doldurulmuş semboller, sol) ve mikrotübüller (açık semboller, sağ) için τ(q) değerlendirerek dinamiklerin zaman içinde nasıl değiştiğini ölçer. trDDM, sağ alttaki göstergede açıklandığı gibi her (ΦA, cM) kombinasyonu (farklı semboller ve renklerle gösterilir) için gerçekleştirilir. C'de gösterilen τ(q) eğrileri, A'dakilerle benzer ölçekleme ve eğilimleri takip eder, ancak aynı zamanda belirli (Φ A, cM) bileşimler için, özellikle de Φ A = 0.75 için zamana bağımlılık gösterir. (D) Aktin filamentleri (kapalı semboller) ve mikrotübüller (açık semboller) için büzülme hızları, uyumlardan karşılık gelen τ (q) eğrilerine göre belirlenir. Tüm grafiklerdeki hata çubukları, üç ila beş çoğaltmadaki değerlerin standart hatasını temsil eder. Bu şekil referans17'den değiştirilmiştir. Bu şeklin daha büyük bir versiyonunu görüntülemek için lütfen buraya tıklayın.
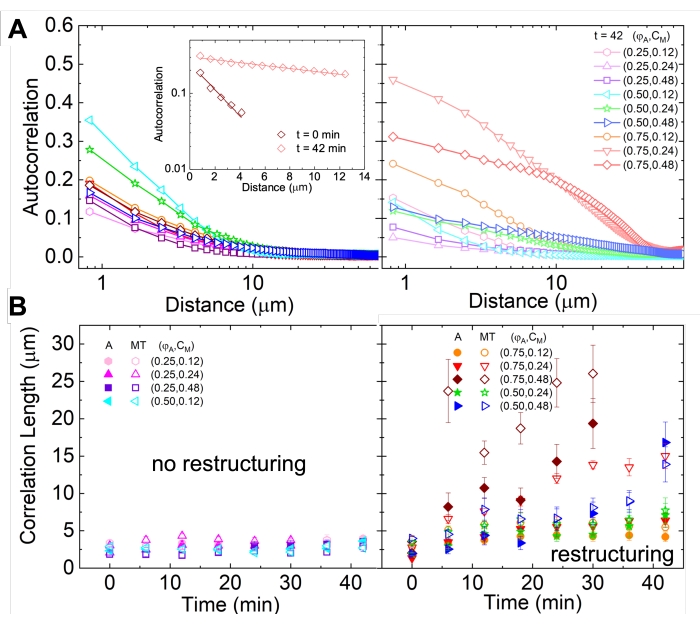
Şekil 5. Uzamsal görüntü otokorelasyonu (SIA) analizi, aktif sitoiskelet kompozitlerinin motor güdümlü yeniden yapılandırılmasını ölçer. (A) Efsanede listelenen (ΦA, cM) formülasyonları için deneyin başlangıcındaki (sol, t = 0 dakika, koyu tonlar) ve sonundaki (sağ, t = 42 dakika, açık tonlar) mikrotübüller için otokorelasyon g(r). Giriş: (ΦA, cM) = (0,75, 0,12) için başlangıç ve son zamanlardaki veri 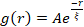 uyumu örneği. (B) A'daki girişte gösterildiği gibi, her bir g(r) eğrisinin üstel uyumları ile belirlenen her (ΦA, cM) için aktin (kapalı semboller) ve mikrotübüller (açık semboller) için ξ ortalama korelasyon uzunlukları. A ve B'deki hata çubukları, üç ila beş çoğaltmadaki standart hatayı temsil eder. Bu şekil referans17'den değiştirilmiştir. Bu şeklin daha büyük bir versiyonunu görüntülemek için lütfen buraya tıklayın.
uyumu örneği. (B) A'daki girişte gösterildiği gibi, her bir g(r) eğrisinin üstel uyumları ile belirlenen her (ΦA, cM) için aktin (kapalı semboller) ve mikrotübüller (açık semboller) için ξ ortalama korelasyon uzunlukları. A ve B'deki hata çubukları, üç ila beş çoğaltmadaki standart hatayı temsil eder. Bu şekil referans17'den değiştirilmiştir. Bu şeklin daha büyük bir versiyonunu görüntülemek için lütfen buraya tıklayın.
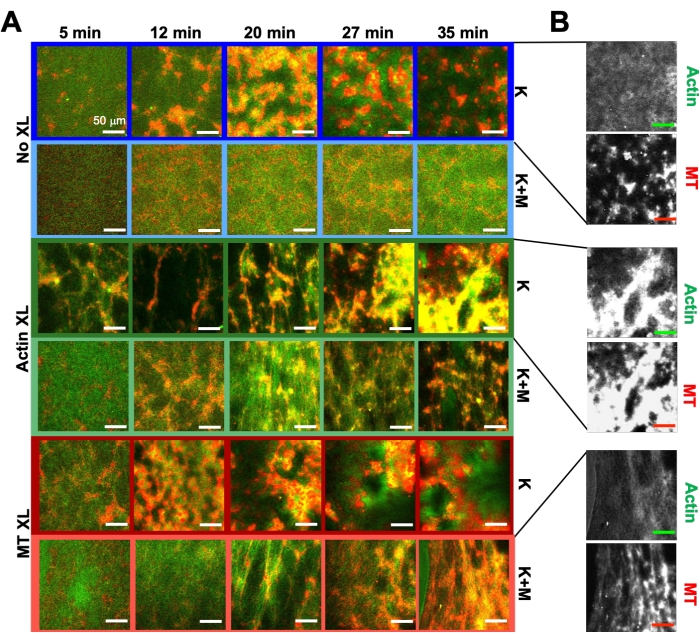
Şekil 6. Kinesin motorlarını ve pasif çapraz bağlayıcıları, programlanabilirliği artırmak ve dinamiklerin ve yapının faz alanını genişletmek için aktif kompozitlere dahil etmek. (A) Aktif kompozitlerdeki aktin (yeşil) ve mikrotübüllerin (kırmızı) iki renkli konfokal görüntüleri, zaman içinde karmaşık formülasyona bağlı yeniden yapılanma gösterir (min'de listelenmiştir). Her satırdaki beş görüntü, kinesin (K, satır 1, 3, 5) veya kinesin ve miyosin (K + M, satır 2, 4, 6) tarafından tahrik edilen bir kompozit için elde edilen 2000 kare zaman serisinin beş karesine karşılık gelir ve pasif çapraz bağlayıcılar (XL yok, satır 1, 2), aktin-aktin çapraz bağlantıları (Actin XL, satır 3, 4) veya mikrotübül-mikrotübül çapraz bağlantıları (MT XL, satır 5, 6). Ölçek çubuklarının tümü 50 μm'dir. Anahat renkleri, Şekil 8'deki renk düzeniyle eşleşir. (B) Sadece kinezin içeren kompozitler için ayrı aktin ve mikrotübül floresan kanalları, hem aktin-MT ko-lokalizasyonu hem de mikro-faz ayrımı ile çeşitli yapılar gösterir. Gösterilen görüntüler c A= 2,32 μM, c T= 3,48 μM, c K = 0,35 μM, cM= 0,47 μM (satır 2, 4, 6), RA= 0,02 (satır 3, 4) ve RMT = 0,005 (satır 5, 6) olan kompozitler içindir. Tüm kompozitler, düzgün dağılmış iç içe geçmiş aktin ve mikrotübül ağları (sütun 1) ile başlar. Çapraz bağlayıcıları olmayan Kinesin tahrikli kompozitler (satır 1), MT bakımından zengin olan gevşek bağlı amorf kümeler oluşturur. Aktin, başlangıçta bu agregaların merkezlerinde birlikte lokalize olur, ancak daha sonra büzülmeye ve birbirlerinden kopmaya devam eden MT bakımından zengin bölgelerden sıkılır. Aktin-aktin çapraz bağlama (satır 3) bu mikro ölçekli aktin-MT ayrımını engeller ve bunun yerine MT bakımından zengin agregalar uzun aktin iplikçikleri aracılığıyla bağlanır. Aktin çapraz bağlama ayrıca aktinin MT açısından zengin bölgelere yavaş alımını sağlar, böylece kompozit birlikte lokalize aktin ve MT kümelerinin bağlı bir ağı haline gelir. Mikrotübül çapraz bağlama (satır 5), zamanla birleşen MT'lerin amorf kümelenmesine yol açaraktin ve MT'lerin daha büyük ölçekli faz ayrımına neden olur. miyozin eklenmesi (satır 2, 4, 6) kinezin güdümlü de-karıştırma ve yeniden yapılandırmayı azaltır. Çapraz bağlayıcılar olmadan (satır 2), kompozitler saatler boyunca çok az yeniden düzenleme gösterir. Çapraz bağlama, aktin ve mikrotübüllerin yeniden yapılandırılmasını ve birlikte lokalizasyonunu arttırır (satır 4, 6). Spesifik olarak, mikrotübüller çapraz bağlandığında (satır 6), web benzeri fiber ağlarına önemli ölçüde iç içe geçme ve yeniden yapılanma vardır. Bu rakam referans50'den değiştirilmiştir. Bu şeklin daha büyük bir versiyonunu görüntülemek için lütfen buraya tıklayın.
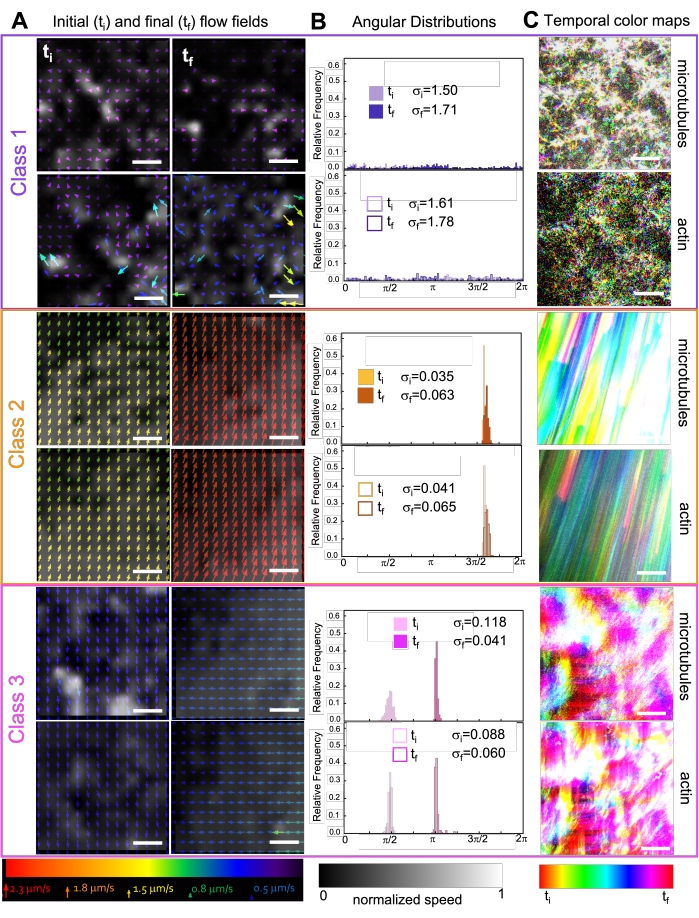
Şekil 7. PIV, aktif kompozitlerin üç sınıf mekansal olarak farklı akış alanları sergilediğini göstermektedir. (A) Üç temsili zaman serisinin ilk (t i) ve son (tf) çerçeveleri için PIV akış alanları, Şekil 6'da gösterilen kompozitlerin sergilediği farklı dinamik sınıfları gösterir. Sınıf 1 (üst, mor), sınıf 2 (orta, turuncu) ve sınıf 3 (alt, macenta) için mikrotübüller (üstte) ve aktin (altta) için PIV akış alanları, alttaki evrensel hız ölçeğine karşılık gelen ok renkleri ve uzamsal hız dağılımını gösteren gri tonlamalı renk haritası, altta gösterilen ölçeğe göre her akış alanı için ayrı ayrı normalleştirilmiştir. Ölçek çubuklarının tümü 50 μM'dir. (B) A'dan (radyan birimleri cinsinden) hız vektörlerinin açısal dağılımları, i ve σ f σ listelenen başlangıç ve son standart sapmalarla. (C) A ve B'de analiz edilen videoların zamansal renk haritaları, her pikselin başlangıç noktasına göre kare-kare konumunu gösterir. Sınıf 1 haritaları küçük ölçekli rastgele hareket gösterir; sınıf 2 haritaları, minimum mekansal veya zamansal varyasyonla hızlı tek yönlü hareketi gösterir; Sınıf 3 haritaları hem sınıf 1 hem de 2'nin özelliklerini sergiler. Bu rakam referans50'den değiştirilmiştir. Bu şeklin daha büyük bir versiyonunu görüntülemek için lütfen buraya tıklayın.
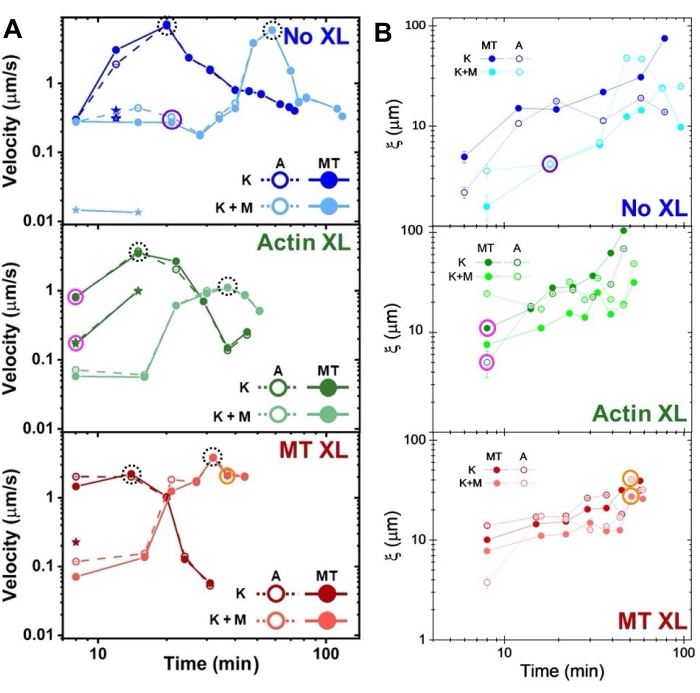
Şekil 8. DDM ve SIA, iki motorlu aktin-mikrotübül kompozitlerinin zamanla değişen dinamiklerini ve yapısını ölçer. (A) DDM ile ölçülen Şekil 6 ve Şekil 7'de tanımlanan kompozitlerin hızları, çapraz bağlama ve miyozin aktivitesi ile programlanan kompozitlerin hızlanmasını ve yavaşlamasını gösterir. Mikrotübüllerin (MT, kapalı daireler) ve aktin (A, açık daireler) hızları, çapraz bağlama (üst, mavi), aktin çapraz bağlama (orta, yeşil), mikrotübül çapraz bağlama (alt, kırmızı), miyozin içermeyen (K, koyu tonlar) ve miyozin (K + M, açık tonlar) ile kompozitlerde aktivite süresinin bir fonksiyonu olarak çizilir. İki hıza sahip sınıf 3 durumlar için, daha yavaş hız bir yıldızla gösterilir. Kesikli siyah dairelerle çevrili veri noktaları, her formülasyon için maksimum hız vmaksimumakarşılık gelir. Hata çubukları (çoğu görmek için çok küçük), karşılık gelen τ (q) 'nun güç yasası uyumları üzerindeki standart hatadır. (B) A'da değerlendirilen aynı zaman serileri kümesi için SIA ile belirlenen yapısal korelasyon uzunlukları ξ ile aktivite süresi. Her veri noktası, karşılık gelen zaman serisinin ilk ve son karesi için belirlenen korelasyon uzunluklarının ortalamasıdır. Genel olarak, ξ, tüm kompozit sistemlerde hem aktin hem de mikrotübüller için zaman içinde artar ve yalnızca kinesin tarafından tahrik edilen kompozitler, miyozinin de mevcut olanlardan daha büyük korelasyon uzunluklarına sahiptir. Şekil 7'de analiz edilen üç zaman serisine karşılık gelen A ve B'deki veri noktaları, karşılık gelen sınıf renginde daire içine alınır (1 = mor, 2 = turuncu, 3 = macenta). Bu rakam referans50'den değiştirilmiştir. Bu şeklin daha büyük bir versiyonunu görüntülemek için lütfen buraya tıklayın.
Yukarıda açıklanan yeniden yapılandırılmış sistemin önemli bir ilerlemesi, modülerliği ve ayarlanabilirliğidir, bu nedenle kullanıcılar, belirli bir hücresel süreci taklit etmek veya belirli işlevsellik veya mekanik özelliklere sahip bir malzemeyi tasarlamak olsun, istenen sonuçlara uyacak şekilde proteinlerin, motorların, çapraz bağlayıcıların vb. konsantrasyonlarını değiştirmeye teşvik edilir. Aktin ve tübülin konsantrasyon aralığındaki sınırlamalar, aktin (~0.2 μM)57,58,59 ve tübülini (~3 - 4 μM)60 polimerize etmek için gereken kritik konsantrasyon ile alt sınırda ve aktin filamentlerinin (~90 μM) 61,62 veya mikrotübüllerin (~35 μM)63 nematik hizalamasına geçişle üst sınırda belirlenir. . Aktin monomerleri ve tübülin dimerleri, birbirlerini sinerjik olarak destekleyen homojen olarak birbirine nüfuz eden sızmış ağlar oluşturmalarını sağlamak için polimerizasyondan sonra birlikte karıştırılmak yerine birlikte filamentlere polimerize edilmelidir. Kompozitlerin sergilediği yeni dinamikler bu etkileşime dayanıyor. Gösterilen sonuçları başarılı bir şekilde yeniden oluşturmak için protokolde özetlenen tüm adımları izlemek genellikle önemli olsa da, bazı adımlar daha kesindir, diğerleri ise belirli ihtiyaçlara ve mevcut kaynaklara uyacak şekilde değiştirmek ve ayarlamak için alana sahiptir.
Örneğin, tekrarlanabilir sonuçların sağlanmasında önemli bir adım, Malzeme Tablosunda sağlanan yönergeleri izleyerek reaktiflerin uygun şekilde hazırlanması ve depolanmasıdır. Sitoiskelet proteinleri (aktin, tübülin, miyosin, kinesin) kararsızdır ve aliquoted edilmeli, sıvı azotla dondurulmalı ve tek kullanımlık alikotlarda -80 ° C'de saklanmalıdır. -80 ° C'den çıkarıldıktan sonra, alikotlar buz üzerinde tutulmalıdır. Sitoiskelet proteinleri, ek donma-çözülme döngülerinden sonra işlevi güvenilir bir şekilde korumazlar.
Mikrotübüller depolimerizasyona ve denatüre karşı aktin'den daha hassastır. -80 ° C'den çıkarıldıktan sonra, tübülin polimerizasyondan önce buz üzerinde tutulmalı ve 12 saat içinde kullanılmalıdır. Polimerize edildikten sonra, mikrotübüller oda sıcaklığında tutulmalıdır. Depolimerizasyonu önlemek için mikrotübüllerin taksol ile stabilize edilmesi de kritik öneme sahiptir. Aktin filamentlerinin falloidin stabilizasyonu, miyozin ve kinesin aktivitesi ile rekabet eden ATP tüketen aktin koşu bandını baskılamak için de önemlidir.
Miyozin motorlarının ultrasantrifüjlenmesi, aktif olmayan miyozin ölü kafalarını ortadan kaldırdığı için bir başka kritik adımdır. Enzimatik olarak inaktif monomerlerin çıkarılmaması, aktin ağının pasif çapraz bağlanmasına ve aktivite kaybına neden olur. Motorların ATPaz aktivitesini uzatmak için, kreatin fosfat ve kreatin fosfokinaz64 gibi bir ATP rejenerasyon sistemi dahil edilebilir.
Son olarak, kompozit aktivitenin sürdürülmesi, filamentlerin ve motorların numune odasının duvarlarına adsorpsiyonunun inhibe edilmesini gerektirir; bu, mikroskop kapaklarının ve slaytlarının pasivasyonu ile elde edilebilir. Motor proteinler özellikle adsorpsiyona eğilimlidir, bu da kompozitin numune odasının yüzeyine çekilmesine, görüş alanından çıkmasına, 2D'ye çökmesine ve artık faaliyete geçmemesine neden olur. Kapak kaymalarını ve kızakları silanize etmek, yüzeyleri pasifleştirmenin ve adsorpsiyonu önlemenin etkili bir yoludur (bkz. adım 1). İn vitro sitoiskelet deneylerinde etkili bir şekilde kullanılan alternatif bir pasivasyon yöntemi, yüzeyin hücre zarı18'e benzer bir lipit çift katmanı ile kaplanmasıdır. Bu yöntem, proteinleri yüzeye bağlamak veya diğer spesifik protein-yüzey etkileşimlerini tanıtmak isterse avantajlıdır, çünkü çift katman işlevselleştirilebilir. Optik cımbız deneyleri için, mikrosferlerin pasivasyonu da kritiktir ve karbodiimid çapraz bağlayıcı kimyası48 yoluyla karboksillenmiş mikrosferlerin BSA veya PEG ile kaplanmasıyla elde edilebilir.
Sunulan protokollerin, araştırmacıların ihtiyaçlarına uyacak şekilde değiştirmeyi düşünebilecekleri birkaç yönü vardır. İlk olarak, araştırmacılar doğal olmayan biyotin-NA çapraz bağlayıcılarını, sırasıyla 28,65,66 aktin ve mikrotübülleri çapraz bağlayan alfa-aktinin veya MAP65 gibi biyolojik çapraz bağlayıcılarla değiştirmeyi seçebilirler. Burada açıklanan kompozitlerde yerel olmayan çapraz bağlayıcıların kullanımı, yerel çapraz bağlayıcılara kıyasla gelişmiş tekrarlanabilirlik, kararlılık ve ayarlanabilirlikleri ile motive edilir. Güçlü biyotin-NA bağı nedeniyle, çapraz bağlayıcıların, geniş kapsamlı ciro oranlarıyla geçici olarak bağlanan çoğu yerel çapraz bağlayıcı yerine, kalıcı olduğu varsayılabilir. Geçici çapraz bağlamanın dinamikleri, çapraz bağlayıcılardan ve motorlardan dinamiklere yapılan katkıların ayrıştırılmasını zorlaştırır. Ayrıca, biyotin-NA bağlayıcıları hem aktin hem de mikrotübülleri çapraz bağlamak ve aktini mikrotübüllere çapraz bağlamak için çok yönlü olarak kullanılabilir. Bu şekilde, çapraz bağlama motifleri arasında açık bir karşılaştırma yapılabilir ve diğer tüm değişkenler (örneğin, çapraz bağlayıcı boyutu, bağlama afinitesi, stokiyometri, vb.) sabit tutulur. Son olarak, biyotin-NA bağlayıcılarını dahil etmek için gereken reaktifler yaygın olarak ticari olarak temin edilebilir, iyi karakterize edilmiştir ve birçok biyofizik laboratuvarında yaygın olarak kullanılmaktadır. Bununla birlikte, burada açıklanan in vitro platformun en önemli güçlü yönlerinden biri modülerliğidir, bu nedenle araştırmacılar, seçtikleri takdirde biyotin-NA bağlayıcılarını yerel bağlayıcılarla sorunsuz bir şekilde değiştirebilmelidir.
İkincisi, mevcut protokolde, aktin monomerleri ve tübülin dimerleri, numune odasına eklenmeden önce bir santrifüj tüpünde birlikte filamentlere polimerize edilir. Dolaşık filamentli proteinlerin çözeltisinin numune odasına akıtılması, özellikle kompozitlerin istenen izotropisini ve homojenliğini kıran mikrotübüllerin akış hizalamasına neden olabilir. Gerçekten de, kararlı hal aktin-mikrotübül kompozitleri üzerindeki önceki çalışmalarda önemli bir ilerleme, aktin ve mikrotübüllerin izotropik nüfuz eden ağlarının oluşumunu sağlamak için aktin ve mikrotübülleri in situ (numune odasında) birlikte polimerize etme yeteneğiydi15,16,27. Bununla birlikte, bu yaklaşımın aktif kompozitlere genişletilmesi, aktin ve tübülin polimerizasyonundan önce motorların numuneye eklenmesini ve deneylerden önce tüm numunenin 37 ° C'de birlikte inkübe edilmesini gerektirecektir. Protokoldeki bu varyasyonun testleri, muhtemelen rakip ATPaz aktivitesi ve motorların uzun süreli 37 ° C inkübasyonu nedeniyle azalmış aktin polimerizasyonu ve fark edilebilir motor aktivitesi ile sonuçlanmamıştır. Neyse ki, Şekil 2, Şekil 3 ve Şekil 6'da görülebileceği gibi, mevcut protokolleri takip ederken kompozitlerin fark edilebilir bir akış hizalaması yoktur. Bununla birlikte, araştırmacıların aktif kompozitlerin yerinde oluşumuna izin veren protokoller tasarlamaları teşvik edilmektedir.
Göz önünde bulundurulması gereken bir diğer nokta, ağdaki tüm aktin filamentlerinin ve mikrotübüllerinin seyrek olarak etiketlenmesini gerektiren floresan etiketleme şemasıdır. Bu etiketleme yaklaşımı, izleyici filamentleri veya mikrosferleri aracılığıyla yapı ve dinamikleri çıkarmak yerine ağın yapısını doğrudan görselleştirmek için optimize edilmiştir. Bununla birlikte, ödünleşim, bireysel filamentlerin parlak bir şekilde etiketlenmemiş ve çözülebilir olmamasıdır. Araştırmacıların hem tek filamentleri çözmek hem de ağ yapısını görselleştirmek için alabilecekleri bir yaklaşım, başka bir florofor ile etiketlenmiş önceden oluşturulmuş filamentlere doping yapmaktır, böylece hem çevreleyen ağ hem de bireysel filamentler aynı anda görüntülenebilir. Bununla birlikte, ikiden fazla florofor ve uyarma / emisyon kanalı kullanıldığında, kanallar arasındaki kanamanın ortadan kaldırılması genellikle zordur, bu nedenle floroforların, filtrelerin ve lazer yoğunluklarının seçiminde dikkatli olunmalıdır.
İlgili bir sınırlama, kompozitlerdeki miyozin veya kinesin motorlarının görselleştirilememesidir. Kullanılan floresan etiketli aktin monomerleri ve tübülin dimerleri ticari olarak temin edilebilirken, kompozitlerde miyozin veya kinezinin görselleştirilmesi kurum içi etiketleme gerektirir. Araştırmacılar, daha önce 18,67'de yapıldığı gibi, motor aktivitesini ve kompozitlerimizin sergilediği dinamiklere ve yapılara bağlanmayı kesin olarak bağlayabilmek için motorları etiketlemek için bir sonraki adımı atmaya teşvik edilmektedir.
Son olarak, mevcut protokolde, kinesin aktivitesinin başlangıcının ve süresinin kontrol edilmediğine dikkat etmek önemlidir. Miyozin aktivitesi, yukarıda tarif edildiği gibi, kinezinin benzer ışık aktivasyonunu oluşturmak için blebbistatinin foto-deaktivasyonu kullanılarak kontrol edildiğinden, ışıkla aktive edilen ATP dahil edilebilir.
Burada açıklanan tasarımların karmaşıklığını artırmak, hücresel koşulları daha iyi taklit etmek ve dinamik-yapı-fonksiyon parametre alanını genişletmek için, gelecekteki çalışmalar vimentin68,69 gibi ara filamentlerin yanı sıra dynein 13,70 gibi diğer motorları da dahil etmeye odaklanacaktır. Gelsolin ayrıca aktin uzunluğu14'ü kontrol etmek için farklı konsantrasyonlarda ve ayrıca mikrotübül sertliğini kontrol etmek için tau proteininde dahil edilecektir.
Özetle, sunulan protokoller, tek bir sistemde farklı substratlar üzerinde hareket eden iki ayrı aktif kuvvet üreten bileşen içeren sitoiskeletten ilham alan aktif madde sistemlerinin dinamiklerinin, yapısının ve mekaniğinin nasıl tasarlanacağını, oluşturulacağını ve karakterize edileceğini açıklamaktadır. Bu ayarlanabilir ve modüler platform, yeniden yapılanma çabalarını hücresel sitoiskeleti taklit etmeye önemli bir adım daha yaklaştırır ve farklı bileşenleri bağımsız olarak dahil ederek, çıkararak ve ayarlayarak özelliklerini geniş bir faz alanı boyunca programlamak için benzersiz bir yetenek sunar. Ayrıca, bu çok yönlü sistemin tüm bileşenleri, daha önce50'de açıklandığı gibi Ross Laboratuvarında saflaştırılan ve talep üzerine temin edilebilen kinesin dimerleri hariç, ticari olarak temin edilebilir (Malzeme Tablosuna bakınız). Son olarak, tüm analiz kodları GitHub49 aracılığıyla ücretsiz olarak kullanılabilir ve ücretsiz programlama dillerine ve yazılımlarına (Python ve Fiji) dayanmaktadır. Bu sistemleri tasarlamak için protokollerin şeffaf bir şekilde yayılması, bu platformu farklı uzmanlıklara, geçmişlere, kurumsal bağlantılara ve araştırma hedeflerine sahip çeşitli bir kullanıcı grubu için daha erişilebilir hale getirecektir.
Yazarların açıklayacak hiçbir şeyi yoktur
Veri analizi konusunda yardımcı oldukları için Maya Hendija ve Dr. Jonathan Michel'e ve yararlı tartışmalar ve rehberlik için Dr. Janet Sheung, Dr. Moumita Das ve Dr. Michael Rust'a teşekkür ederiz. Bu araştırma, RMRA ve JLR'ye verilen William M. Keck Vakfı Araştırma Bursu ve NSF DMREF Ödülü (DMR 2119663) ve RMR-A ve RJM'ye verilen Ulusal Sağlık Enstitüleri R15 Hibeleri (R15GM123420, 2R15GM123420-02) ile desteklenmiştir.
| Name | Company | Catalog Number | Comments |
| (-)-Blebbistatin Abbreviation used in paper: blebbistatin | Sigma Aldrich | B0560 | Stock Concentration: 200 μM in DMSO Storage: dessicated, in DMSO, -20ºC Stock and Experiment Recipes: dissolve 1 mg of powder to 200 μM in DMSO Storage, Handling, Troubleshooting Notes: limited shelf-life, typically stops functioning reliably after 3-4 months. purchase and prepare new solution every 3 months. |
| 1:20 488-tubulin:tubulin mixture Abbreviation used in paper: 5-488-tubulin | NA | NA | Stock Concentration: 5 mg/ml in PEM Storage: single use aliquots, -80ºC, avoid freeze-thaw cycles Stock and Experiment Recipes: mix tubulin and 488-tubulin at a 20:1 ratio, flash freeze with LN2 Storage, Handling, Troubleshooting Notes: each aliquot can be used for up to 12 hrs stored on ice at 4ºC, protect from light |
| 1:20 R-tubulin:tubulin mixture Abbreviation used in paper: 5-R-tubulin | NA | NA | Stock Concentration: 5 mg/ml in PEM Storage: single use aliquots, -80ºC, avoid freeze-thaw cycles Stock and Experiment Recipes: mix tubulin and rhodamine tubulin at a 20:1 ratio, flash freeze with LN2 Storage, Handling, Troubleshooting Notes: each aliquot can be used for up to 12 hrs stored on ice at 4ºC, protect from light |
| actin (biotin): skeletal muscle Abbreviation used in paper: biotin-actin | Cytoskeleton | AB07 | Stock Concentration: 1 mg/ml in G-buffer Storage: single use aliquots, -80ºC, avoid freeze-thaw cycles Stock and Experiment Recipes: reconstitute to 1 mg/ml in G-buffer, flash freeze with LN2 Storage, Handling, Troubleshooting Notes: (1) immediately prior to use dilute to 0.5 mg/ml in PEM, (2) once removed from -80ºC, store aliquot on ice at 4ºC for up to 1 week |
| actin (rhodamine): rabbit skeletal muscle Abbreviation used in paper: R-actin | Cytoskeleton | AR05 | Stock Concentration: 1.5 mg/ml in G-buffer Storage: single use aliquots, -80ºC, avoid freeze-thaw cycles Stock and Experiment Recipes: reconstitute to 1.5 mg/ml in G-buffer, flash freeze with LN2 Storage, Handling, Troubleshooting Notes: once removed from -80ºC, store aliquot on ice at 4ºC, can be used for up to 1 week |
| adenosine triphosphate Abbreviation used in paper: ATP | Thermo Fisher Scientific | A1048 | Stock Concentration: 100 mM Storage: in solution (pH 7), -20ºC Stock and Experiment Recipes: reconsitute in DI H20, bring pH to 7 with NaOH Storage, Handling, Troubleshooting Notes: routinely check pH and adjust as needed, hydrolyzes over time, replace every ~6-12 months |
| AlexaFluor488 Phalloidin Abbreviation used in paper: 488-phalloidin | Thermo Fisher Scientific | A12379 | Stock Concentration: 100 μM DMSO Storage: protected from light, dessicated, -20ºC Stock and Experiment Recipes: reconstitute to 100 μM with DMSO Storage, Handling, Troubleshooting Notes: immediately prior to use dilute to 20 μM in PEM (1 μL in 4 μL PEM) |
| AlexaFluor488–labeled actin Abbreviation used in paper: 488-actin | Thermo Fisher Scientific | A12373 | Stock Concentration: 1.5 mg/ml in G-buffer Storage: single use aliquots, -80ºC, avoid freeze-thaw cycles Stock and Experiment Recipes: reconstitute to 1.5 mg/ml in G-buffer, flash freeze with LN2 Storage, Handling, Troubleshooting Notes: this item has been discontinued |
| Basic Plasma Cleaner Abbreviation used in paper: plasma cleaner | Harrick Plasma | PDC-32G | |
| Bemis Parafilm M Laboratory Wrapping Film Abbreviation used in paper: transparent film | Thermo Fisher Scientific | 13-374-5 | |
| D-(+)-Glucose Abbreviation used in paper: | Thermo Fisher Scientific | A1682836 | Stock Concentration: 100x Storage: store at stock concentration (100x) or 10x concentration, dessicated, at -20ºC Stock and Experiment Recipes: reconstitute powder to 4.5 mg/ml in DI H20 Storage, Handling, Troubleshooting Notes: final concentration in solution should 45 μg/mL |
| D-Biotin Abbreviation used in paper: biotin | Fisher Scientific | BP232-1 | Stock Concentration: 1.02 mM in PEM Storage: dessicated, 4ºC |
| deionized nanopure water Abbreviation used in paper: DI | |||
| Dimethyldichlorosilane Abbreviation used in paper: silane | Thermo Fisher Scientific | D/3820/PB05 | Stock Concentration: 2% dissolved in Toulene |
| Dithiothreitol Abbreviation used in paper: DTT | Thermo Fisher Scientific | R0861 | Stock Concentration: 1 M in DMSO Storage: dessicated, -20ºC Stock and Experiment Recipes: dilute to 2 mM in PEM immediately before each experiment |
| DMSO Anhydrous Abbreviation used in paper: DMSO | Thermo Fisher Scientific | D12345 | |
| F-Buffer Abbreviation used in paper: F-buffer | NA | NA | Stock Concentration: 10x Storage: dessicated, -20ºC Stock and Experiment Recipes: 10 mM Imidazole (pH 7.0), 50 mM KCl, 1 mM MgCl2, 1 mM EGTA, 0.2 mM ATP |
| G-Buffer Abbreviation used in paper: G-buffer | NA | NA | Stock Concentration: 10x Storage: dessicated, -20ºC Stock and Experiment Recipes: 2.0 mM Tris (pH 8), 0.2 mM ATP, 0.5 mM DTT, 0.1 mM CaCl2. Store at -20°C. |
| glass microscope slide Abbreviation used in paper: slide | Thermo Fisher Scientific | 22-310397 | |
| Glucose oxidase + catalase + β-mercaptoethanol Abbreviation used in paper: GOC | Sigma Aldrich | G2133-250KU, C1345, 63689 | Stock Concentration: 100x Storage: store at stock concentration (100x) or 10x concentration, dessicated, at -20ºC Stock and Experiment Recipes: For 100x: 4.3 mg/ml glucose oxidase, 0.7 mg/ml catalase, 0.5% v/v β-mercaptoethanol in DI H20 Storage, Handling, Troubleshooting Notes: final concentration in solution should be: 0.005% β-mercaptoethanol, 43 μg/mL glucose oxidase, 7 μg/mL catalase |
| glu-GOC oxygen scavenging system Abbreviation used in paper: glu-GOC | NA | NA | Stock Concentration: 100x Storage: prepare fresh each time Stock and Experiment Recipes: mix equal parts Glu and GOC and add at 1/100 final sample volume immediately before imaging Storage, Handling, Troubleshooting Notes: prepare from Glu and GOC immediately before imaging |
| Guanosine triphosphate Abbreviation used in paper: GTP | Thermo Fisher Scientific | R0461 | Stock Concentration: 100 mM Storage: 100 μL aliquots at -20ºC |
| Instant Mix 1-minute epoxy Abbreviation used in paper: epoxy | Loctite | 1366072 | |
| Kinesin-1 401 BIO 6x HIS Abbreviation used in paper: kinesin | Prepared in JL Ross Lab at Syracuse University | NA | Stock Concentration: 8.87 μM in PEM Storage: single use aliquots, -80ºC, avoid freeze-thaw cycles Storage, Handling, Troubleshooting Notes: biotinylated dimers form kinesin clusters, each aliquot can be used for up to 12 hrs stored on ice at 4ºC |
| NeutrAvidin Abbreviation used in paper: NA | Thermo Fisher Scientific | 31000 | Stock Concentration: 5 mg/ml in PEM Storage: dessicated, -20ºC Stock and Experiment Recipes: reconstitute powder to 5 mg/ml in PEM |
| No 1. glass coverslips (24 mm x 24 mm) Abbreviation used in paper: coverslip | Thermo Fisher Scientific | 12-548-CP | |
| Paclitaxel Abbreviation used in paper: Taxol | Thermo Fisher Scientific | P3456 | Stock Concentration: 2 mM in DMSO Storage: protected from light, dessicated, -20ºC Stock and Experiment Recipes: reconstitute to 2 mM with DMSO Storage, Handling, Troubleshooting Notes: immediately prior to use dilute to 200 μM in DMSO (0.4 μL in 3.6 μL DMSO) |
| PEM-100 Abbreviation used in paper: PEM | NA | NA | Stock Concentration: 1x Storage: room temperature (RT) Stock and Experiment Recipes: 100 mM K-PIPES (pH 6.8), 2 mM EGTA, 2 mM MgCl2 Storage, Handling, Troubleshooting Notes: use KOH to adjust pH to 6.8, recheck pH often and adjust accordingly |
| phalloidin Abbreviation used in paper: phalloidin | Thermo Fisher Scientific | P3457 | Stock Concentration: 100 μM in DMSO Storage: protected from light, dessicated, -20ºC, adhere closely to storage/handling conditions Stock and Experiment Recipes: reconstitute to 100 μM with DMSO Storage, Handling, Troubleshooting Notes: susceptible to impurities in its preparation and denaturing, identifiable as large amorphous aggregates of actin in samples |
| porcine brain tubulin Abbreviation used in paper: tubulin | Cytoskeleton | T240 | Stock Concentration: 5 mg/ml in PEM Storage: single use aliquots, -80ºC, avoid freeze-thaw cycles Stock and Experiment Recipes: reconstitute powder to 5 mg/ml in PEM, flash freeze with LN2 Storage, Handling, Troubleshooting Notes: each aliquot can be used for up to 12 hrs stored on ice at 4ºC |
| Potassium Chloride Abbreviation used in paper: KCl | Thermo Fisher Scientific | AM9640G | Stock Concentration: 4 M Storage: RT |
| Rabbit skeletal actin Abbreviation used in paper: actin | Cytoskeleton | AKL99 | Stock Concentration: 2 mg/ml in G-buffer Storage: single use aliquots, -80ºC, avoid freeze-thaw cycles Stock and Experiment Recipes: reconstitute to 2 mg/ml in G-buffer, flash freeze with LN2 Storage, Handling, Troubleshooting Notes: once removed from -80ºC, store aliquot on ice at 4ºC, can be used for up to 1 week |
| Rabbit skeletal myosin II Abbreviation used in paper: myosin | Cytoskeleton | MY02 | Stock Concentration: 10 mg/ml in PEM Storage: single use aliquots, -80ºC, avoid freeze-thaw cycles Stock and Experiment Recipes: reconstitute powder to 10 mg/ml in PEM, flash freeze with LN2 Storage, Handling, Troubleshooting Notes: monomers form minifilaments at low KCl, each aliquot can be used for up to 12 hrs stored on ice at 4ºC |
| Tubulin (biotin): porcine brain Abbreviation used in paper: biotin-tubulin | Cytoskeleton | T333P | Stock Concentration: 5 mg/ml in PEM Storage: single use aliquots, -80ºC, avoid freeze-thaw cycles Stock and Experiment Recipes: reconstitute powder to 5 mg/ml in PEM, flash freeze with LN2 Storage, Handling, Troubleshooting Notes: immediately prior to use dilute to 0.5 mg/ml in PEM |
| Tubulin (fluorescent HiLyte 488): porcine brain Abbreviation used in paper: 488-tubulin | Cytoskeleton | TL488M | Stock Concentration: 5 mg/ml in PEM Storage: single use aliquots, -80ºC, avoid freeze-thaw cycles Stock and Experiment Recipes: reconstitute powder to 5 mg/ml in PEM, flash freeze with LN2 Storage, Handling, Troubleshooting Notes: each aliquot can be used for up to 12 hrs stored on ice at 4ºC, protect from light |
| tubulin (rhodamine): porcine brain Abbreviation used in paper: R-tubulin | Cytoskeleton | TL590M | Stock Concentration: 5 mg/ml in PEM Storage: single use aliquots, -80ºC, avoid freeze-thaw cycles Stock and Experiment Recipes: reconstitute powder to 5 mg/ml in PEM, flash freeze with LN2 Storage, Handling, Troubleshooting Notes: each aliquot can be used for up to 12 hrs stored on ice at 4ºC, protect from light |
| Tween 20 Abbreviation used in paper: Tween20 | Thermo Fisher Scientific | J20605.AP | Stock Concentration: 1% v/v in DI H20 Storage: RT |
| ultracentrifuge grade microtubes Abbreviation used in paper: Beckman-Coulter Optima Max XP | Beckman Coultier | 343776 | Storage, Handling, Troubleshooting Notes: 8x34 mm PC |
| UV light curing glue Abbreviation used in paper: UV glue | Pharda | SKG-2869 |
- Fletcher, D. A., Mullins, R. D. Cell mechanics and the cytoskeleton. Nature. 463 (7280), 485-492 (2010).
- Koenderink, G. H., Paluch, E. K. Architecture shapes contractility in actomyosin networks. Current Opinion in Cell Biology. 50, 79-85 (2018).
- Dogterom, M., Koenderink, G. H. Actin-microtubule crosstalk in cell biology. Nature Reviews Molecular Cell Biology. 20 (1), 38-54 (2019).
- Burla, F., Mulla, Y., Vos, B. E., Aufderhorst-Roberts, A., Koenderink, G. H. From mechanical resilience to active material properties in biopolymer networks. Nature Reviews Physics. 1 (4), 249-263 (2019).
- Wen, Q., Janmey, P. A. Polymer physics of the cytoskeleton. Current Opinion in Solid State and Materials Science. 15 (5), 177-182 (2011).
- Xiao, Q., Hu, X., Wei, Z., Tam, K. Y. Cytoskeleton molecular motors: structures and their functions in neuron. International Journal of Biological Sciences. 12 (9), 1083-1092 (2016).
- Ajeti, V. et al. Wound healing coordinates actin architectures to regulate mechanical work. Nature Physics. 15 (7), 696-705 (2019).
- Jung, W. et al. Dynamic motions of molecular motors in the actin cytoskeleton. Cytoskeleton. 76 (11-12), 517-531 (2019).
- Pollard, T. D., O'Shaughnessy, B. Molecular mechanism of cytokinesis. Annual Review of Biochemistry. 88 (1), 661-689 (2019).
- Huber, F., Boire, A., López, M. P., Koenderink, G. H. Cytoskeletal crosstalk: when three different personalities team up. Current Opinion in Cell Biology. 32, 39-47 (2015).
- Rivero, F. et al. The role of the cortical cytoskeleton: F-actin crosslinking proteins protect against osmotic stress, ensure cell size, cell shape and motility, and contribute to phagocytosis and development. Journal of Cell Science. 109 (11), 2679-2691 (1996).
- Duclos, G. et al. Topological structure and dynamics of three-dimensional active nematics. Science. 367 (6482), 1120-1124 (2020).
- Baclayon, M. et al. Optical tweezers-based measurements of forces and dynamics at microtubule ends. Optical Tweezers. 1486, 411-435 (2017).
- Gurmessa, B., Fitzpatrick, R., Falzone, T. T., Robertson-Anderson, R. M. Entanglement density tunes microscale nonlinear response of entangled actin. Macromolecules. 49 (10), 3948-3955 (2016).
- Francis, M. L. et al. Non-monotonic dependence of stiffness on actin crosslinking in cytoskeleton composites. Soft Matter. 15 (44), 9056-9065 (2019).
- Ricketts, S. N. et al. Varying crosslinking motifs drive the mesoscale mechanics of actin-microtubule composites. Scientific Reports. 9 (1), 12831 (2019).
- Lee, G. et al. Active cytoskeletal composites display emergent tunable contractility and restructuring. Soft Matter. 17 (47), 10765-10776 (2021).
- Murrell, M. P., Gardel, M. L. F-actin buckling coordinates contractility and severing in a biomimetic actomyosin cortex. Proceedings of the National Academy of Sciences. 109 (51), 20820-20825 (2012).
- Soares e Silva, M. et al. Active multistage coarsening of actin networks driven by myosin motors. Proceedings of the National Academy of Sciences. 108 (23), 9408-9413 (2011).
- Sonn-Segev, A., Bernheim-Groswasser, A., Roichman, Y. Dynamics in steady state in vitro acto-myosin networks. Journal of Physics: Condensed Matter. 29 (16), 163002 (2017).
- Ideses, Y., Sonn-Segev, A., Roichman, Y., Bernheim-Groswasser, A. Myosin II does it all: assembly, remodeling, and disassembly of actin networks are governed by myosin II activity. Soft Matter. 9 (29), 7127 (2013).
- Fürthauer, S. et al. Self-straining of actively crosslinked microtubule networks. Nature Physics. 15 (12), 1295-1300 (2019).
- Lemma, L. M. et al. Multiscale microtubule dynamics in active nematics. Physical Review Letters. 127 (14), 148001 (2021).
- Fan, Y., Wu, K.-T., Aghvami, S. A., Fraden, S., Breuer, K. S. Effects of confinement on the dynamics and correlation scales in kinesin-microtubule active fluids. Physical Review E. 104 (3), 034601 (2021).
- Triclin, S. et al. Self-repair protects microtubules from destruction by molecular motors. Nature Materials. 20 (6), 883-891 (2021).
- Lee, G. et al. Myosin-driven actin-microtubule networks exhibit self-organized contractile dynamics. Science Advances. 7 (6), eabe4334 (2021).
- Ricketts, S. N., Ross, J. L., Robertson-Anderson, R. M. Co-entangled actin-microtubule composites exhibit tunable stiffness and power-law stress relaxation. Biophysical Journal. 115 (6), 1055-1067 (2018).
- Bendix, P. M. et al. A quantitative analysis of contractility in active cytoskeletal protein networks. Biophysical Journal. 94 (8), 3126-3136 (2008).
- Linsmeier, I. et al. Disordered actomyosin networks are sufficient to produce cooperative and telescopic contractility. Nature Communications. 7 (1), 12615 (2016).
- Stam, S. et al. Filament rigidity and connectivity tune the deformation modes of active biopolymer networks. Proceedings of the National Academy of Sciences. 114 (47), E10037-E10045 (2017).
- Yadav, V. et al. Filament nucleation tunes mechanical memory in active polymer networks. Advanced Functional Materials. 29 (49), 1905243 (2019).
- Ennomani, H. et al. Architecture and connectivity govern actin network contractility. Current Biology. 26 (5), 616-626 (2016).
- Alvarado, J., Sheinman, M., Sharma, A., MacKintosh, F. C., Koenderink, G. H. Molecular motors robustly drive active gels to a critically connected state. Nature Physics. 9 (9), 591-597 (2013).
- Alvarado, J., Cipelletti, L., Koenderink, G. H. Uncovering the dynamic precursors to motor-driven contraction of active gels. Soft Matter. 15 (42), 8552-8565 (2019).
- Jung, W., Murrell, M. P., Kim, T. F-actin cross-linking enhances the stability of force generation in disordered actomyosin networks. Computational Particle Mechanics. 2 (4), 317-327 (2015).
- Lenz, M., Thoresen, T., Gardel, M. L., Dinner, A. R. Contractile units in disordered actomyosin bundles arise from f-actin buckling. Physical Review Letters. 108 (23), 238107 (2012).
- Memarian, F.L. et al. Active nematic order and dynamic lane formation of microtubules driven by membrane-bound diffusing motors. Proceedings of the National Academy of Sciences. 118 (52), e2117107118 (2021).
- Needleman, D., Dogic, Z. Active matter at the interface between materials science and cell biology. Nature Reviews Materials. 2 (9), 17048 (2017).
- Foster, P. J., Fürthauer, S., Shelley, M. J., Needleman, D. J. Active contraction of microtubule networks. eLife. 4, e10837 (2015).
- Thijssen, K. et al. Submersed micropatterned structures control active nematic flow, topology, and concentration. Proceedings of the National Academy of Sciences. 118 (38), e2106038118 (2021).
- Sanchez, T., Chen, D. T. N., DeCamp, S. J., Heymann, M., Dogic, Z. Spontaneous motion in hierarchically assembled active matter. Nature. 491 (7424), 431-434 (2012).
- Colen, J. et al. Machine learning active-nematic hydrodynamics. Proceedings of the National Academy of Sciences. 118 (10), e2016708118 (2021).
- Mitchell, K. A., Tan, A. J., Arteaga, J., Hirst, L. S. Fractal generation in a two-dimensional active-nematic fluid. Chaos: An Interdisciplinary Journal of Nonlinear Science. 31 (7), 073125 (2021).
- Pandolfi, R. J., Edwards, L., Johnston, D., Becich, P., Hirst, L. S. Designing highly tunable semiflexible filament networks. Physical Review E. 89 (6), 062602 (2014).
- Tan, A. J. et al. Topological chaos in active nematics. Nature Physics. 15 (10), 1033-1039 (2019).
- Roostalu, J., Rickman, J., Thomas, C., Nédélec, F., Surrey, T. Determinants of polar versus nematic organization in networks of dynamic microtubules and mitotic motors. Cell. 175 (3), 796-808.e14 (2018).
- Ndlec, F. J., Surrey, T., Maggs, A. C., Leibler, S. Self-organization of microtubules and motors. Nature. 389 (6648), 305-308 (1997).
- Sheung, J. Y. et al. Motor-driven restructuring of cytoskeleton composites leads to tunable time-varying elasticity. ACS Macro Letters. 10 (9), 1151-1158 (2021).
- McGorty, R. PyDDM v0.2.0. Zenodo. (2022).
- Achiriloaie, D. H. et al. Kinesin and myosin motors compete to drive rich multi-phase dynamics in programmable cytoskeletal composites. arXiv. (2021).
- Wulstein, D. M., Regan, K. E., Garamella, J., McGorty, R. J., Robertson-Anderson, R. M. Topology-dependent anomalous dynamics of ring and linear DNA are sensitive to cytoskeleton crosslinking. Science Advances. 5 (12), eaay5912 (2019).
- McGorty, R. Image-Correlation. at <https://github.com/rmcgorty/Image-Correlation>. (2020).
- Robertson, C. Theory and practical recommendations for autocorrelation-based image correlation spectroscopy. Journal of Biomedical Optics. 17 (8), 080801 (2012).
- McGorty, R. Differential Dynamic Microscopy - Python. at <https://github.com/rmcgorty/Differential-Dynamic-Microscopy---Python>. (2021).
- Cerbino, R., Trappe, V. Differential dynamic microscopy: probing wave vector dependent dynamics with a microscope. Physical Review Letters. 100 (18), 188102 (2008).
- Robertson-Anderson, R. M. Optical tweezers microrheology: from the basics to advanced techniques and applications. ACS Macro Letters. 7 (8), 968-975 (2018).
- Pollard, T. D. Polymerization of ADP-actin. Journal of Cell Biology. 99 (3), 769-777 (1984).
- Coué, M., Brenner, S. L., Spector, I., Korn, E. D. Inhibition of actin polymerization by latrunculin A. FEBS Letters. 213 (2), 316-318 (1987).
- Pollard, T. D. Actin and actin-binding proteins. Cold Spring Harbor Perspectives in Biology. 8 (8), a018226 (2016).
- Kumar, N. Taxol-induced polymerization of purified tubulin. Mechanism of action. Journal of Biological Chemistry. 256 (20), 10435-10441 (1981).
- Käs, J. et al. F-actin, a model polymer for semiflexible chains in dilute, semidilute, and liquid crystalline solutions. Biophysical Journal. 70 (2), 609-625 (1996).
- Viamontes, J., Narayanan, S., Sandy, A. R., Tang, J. X. Orientational order parameter of the nematic liquid crystalline phase of F -actin. Physical Review E. 73 (6), 061901 (2006).
- Hitt, A. L., Cross, A. R., Williams, R. C. Microtubule solutions display nematic liquid crystalline structure. Journal of Biological Chemistry. 265 (3), 1639-1647 (1990).
- Andexer, J. N., Richter, M. Emerging enzymes for ATP regeneration in biocatalytic processes. ChemBioChem. 16 (3), 380-386 (2015).
- Farhadi, L. et al. Actin and microtubule crosslinkers tune mobility and control co-localization in a composite cytoskeletal network. Soft Matter. 16 (31), 7191-7201 (2020).
- Falzone, T. T., Lenz, M., Kovar, D. R., Gardel, M. L. Assembly kinetics determine the architecture of α-actinin crosslinked F-actin networks. Nature Communications. 3 (1), 861 (2012).
- Thoresen, T., Lenz, M., Gardel, M. L. Reconstitution of contractile actomyosin bundles. Biophysical Journal. 100 (11), 2698-2705 (2011).
- Sanghvi-Shah, R., Weber, G. F. Intermediate filaments at the junction of mechanotransduction, migration, and development. Frontiers in Cell and Developmental Biology. 5, 81 (2017).
- Shen, Y. et al. Effects of vimentin intermediate filaments on the structure and dynamics of in vitro multicomponent interpenetrating cytoskeletal networks. Physical Review Letters. 127 (10), 108101 (2021).
- Laan, L., Roth, S., Dogterom, M. End-on microtubule-dynein interactions and pulling-based positioning of microtubule organizing centers. Cell Cycle. 11 (20), 3750-3757 (2012).
Erratum
Erratum: Reconstituting and Characterizing Actin-Microtubule Composites with Tunable Motor-Driven Dynamics and MechanicsAn erratum was issued for: Reconstituting and Characterizing Actin-Microtubule Composites with Tunable Motor-Driven Dynamics and Mechanics. The Authors section was updated.
Mehrzad Sasanpour1
Daisy H. Achiriloaie1,2
Gloria Lee1
Gregor Leech1
Christopher Currie1
K. Alice Lindsay3
Jennifer L. Ross3
Ryan J. McGorty1
Rae M. Robertson-Anderson1
1Department of Physics and Biophysics, University of San Diego
2W. M. Keck Science Department, Scripps College, Pitzer College, and Claremont McKenna College
3Department of Physics, Syracuse University
to:
Mehrzad Sasanpour1
Daisy H. Achiriloaie1,2
Gloria Lee1
Gregor Leech1
Maya Hendija1
K. Alice Lindsay3
Jennifer L. Ross3
Ryan J. McGorty1
Rae M. Robertson-Anderson1
1Department of Physics and Biophysics, University of San Diego
2W. M. Keck Science Department, Scripps College, Pitzer College, and Claremont McKenna College
3Department of Physics, Syracuse University
ABOUT JoVE
Copyright © 2024 MyJoVE Corporation. All rights reserved