
Summary
Abstract
Introduction
Protocol
Representative Results
Discussion
Acknowledgements
Materials
References
Neuroscience
Frazionamento subcellulare per l'isolamento di componenti sinaptici dal cervello murino
Questo protocollo presenta un metodo robusto e dettagliato per ottenere sinaptosomi altamente puri, vescicole sinaptiche e altre frazioni sinaptiche dal cervello del topo. Questo metodo consente la valutazione dei processi sinaptici, compresa l'analisi biochimica della localizzazione e della funzione delle proteine con risoluzione compartimentale.
I terminali sinaptici sono i siti primari della comunicazione neuronale. La disfunzione sinaptica è un segno distintivo di molti disturbi neuropsichiatrici e neurologici. La caratterizzazione dei sottocompartimenti sinaptici mediante isolamento biochimico è, quindi, un potente metodo per chiarire le basi molecolari dei processi sinaptici, sia in salute che in malattia. Questo protocollo descrive l'isolamento dei terminali sinaptici e dei sottocompartimenti sinaptici dal cervello dei topi mediante frazionamento subcellulare. In primo luogo, le strutture terminali sinaptiche sigillate, note come sinaptosomi, vengono isolate in seguito all'omogeneizzazione del tessuto cerebrale. I sinaptosomi sono compartimenti neuronali pre e post-sinaptici con membrane pizzicate e sigillate. Queste strutture mantengono uno stato metabolicamente attivo e sono preziose per lo studio della struttura e della funzione sinaptica. I sinaptosomi vengono quindi sottoposti a lisi ipotonica e ultracentrifugazione per ottenere sottocompartimenti sinaptici arricchiti per vescicole sinaptiche, citosol sinaptico e membrana plasmatica sinaptica. La purezza della frazione è confermata dalla microscopia elettronica e dall'analisi dell'arricchimento biochimico per proteine specifiche dei compartimenti sub-sinaptici. Il metodo presentato è uno strumento semplice e prezioso per studiare le caratteristiche strutturali e funzionali della sinapsi e l'eziologia molecolare di vari disturbi cerebrali.
Le sinapsi sono le unità computazionali di base del cervello attraverso le quali i neuroni comunicano ed esercitano funzioni diverse e squisitamente complesse. Le sinapsi sono, quindi, fondamentali per la salute del cervello1; La disfunzione sinaptica è implicata come fonte o risultato di molti disturbi2. Le sinapsi sono costituite da terminali pre- e post-sinaptici, estensioni di due diversi neuroni che sono strettamente apposti e separati da una fessura sinaptica attraversata da molecole di adesione sinaptica. Le informazioni fluiscono dal compartimento pre- al post-sinaptico sotto forma di messaggeri chimici chiamati neurotrasmettitori1. I processi molecolari coinvolti nella neurotrasmissione sono aree attive di ricerca 3,4,5. Comprendere i processi patogenetici all'interno dei terminali sinaptici e la risposta delle sinapsi alla patologia in altri sotto-compartimenti neuronali sono passi cruciali per affrontare i disturbi del cervello 1,2. Diversi progressi metodologici, applicati prevalentemente a modelli murini, hanno fatto avanzare questa ricerca6. L'isolamento delle frazioni sinaptiche mediante centrifugazione differenziale è uno di questi metodi di cambiamento di paradigma che ha permesso la valutazione dettagliata dei processi sinaptici in salute e malattia.
Il cervello umano adulto è costituito da 80-90 miliardi di neuroni 7,8. Tra le specie murine, il cervello del ratto contiene circa ~ 200 milioni di neuroni, mentre i topi hanno ~ 70 milioni 9,10. Ogni neurone forma migliaia di connessioni sinaptiche specifiche con una rete di neuroni altamente polarizzati mescolati con cellule gliali e vascolarizzazione densa. In un tessuto così complesso ed eterogeneo, un tempo era impensabile isolare e studiare le sinapsi come un sistema indipendente. Nel 1960, Victor Whittaker, Catherine Hebb e altri hanno reso possibile questo isolando terminali sinaptici intatti usando il frazionamento subcellulare11,12,13,14. Nel tentativo di isolare le vescicole sinaptiche (SV), hanno omogeneizzato i cervelli attraverso la forza di taglio liquido nel saccarosio iso-osmotico (0,32 M) seguita da ultracentrifugazione. Hanno ottenuto terminali nervosi o varicosità intatte, chiusi in membrana plasmatica, che hanno chiamato particelle nervose (NEP)11,13. Poiché le caratteristiche strutturali e funzionali della sinapsi sono state preservate in queste strutture, le NEP sono state successivamente definite "sinaptosomi" per congruenza con altri organelli subcellulari13,15. Vale la pena notare che il lavoro di Eduardo de Robertis e colleghi, che hanno coniato il termine "vescicola sinaptica", si è sovrapposto a quello di Whittaker e colleghi e ha contribuito alla convalida dell'isolamento e della caratterizzazione del "sinaptosoma"16,17,18.
I sinaptosomi sono strutture fisiologicamente attive che contengono tutte le proprietà cellulari e molecolari necessarie per la conservazione, il rilascio e la ricaptazione dei neurotrasmettitori13,18. Anche la conservazione delle caratteristiche sinaptiche chiave in vitro e l'assenza di componenti non sinaptici contribuiscono all'utilità di questo metodo di isolamento. I sinaptosomi hanno contribuito immensamente alla comprensione delle proprietà chimiche e fisiologiche della neurotrasmissione e vengono ora utilizzati per studiare i processi molecolari sinaptici e le loro alterazioni nella malattia 19,20,21,22,23. I sinaptosomi sono anche il materiale di partenza iniziale per isolare componenti sinaptici come SV, vescicole rivestite di clatrina (CCV), citosol sinaptico, membrana plasmatica sinaptica, mitocondri sinaptici, molecole di adesione sinaptica e altri componenti di interesse, che possono facilitare la comprensione dei meccanismi molecolari della funzione sinaptica 18,19,20,24,25,26, 27,28. Questi componenti sub-sinaptici possono essere ottenuti mediante lisi osmotica dei sinaptosomi e ultracentrifugazione del gradiente di densità del saccarosio15,29. Sebbene il metodo di frazionamento subcellulare originale del gruppo di ricerca di Whittaker sia noto per essere efficiente nell'isolare sinaptosomi di qualità e SV 13,30, recenti ottimizzazioni migliorano la purezza delle frazioni subcellulari 22,23,31,32. Questo articolo fornisce una versione altamente dettagliata e accessibile di un protocollo classico per il frazionamento subcellulare del tessuto cerebrale murino per isolare sinaptosomi, SV e altri componenti sub-sinaptici.
Tutti gli esperimenti con i topi sono stati approvati dall'Institutional Animal Care and Use Committee (IACUC) dell'Università di Yale (protocollo 2021-11117) ed eseguiti in una struttura accreditata dall'Associazione per la valutazione e l'accreditamento di Laboratory Animal Care International (AAALAC). La cura e la stabulazione degli animali erano conformi alla Guida per la cura e l'uso degli animali da laboratorio33 e sono state fornite dallo Yale Animal Resource Center (YARC). Gli animali sono stati mantenuti in un ciclo luce/buio di 12 ore con accesso ad libitum al cibo e all'acqua. Sono necessari da cinque a otto topi o da due a quattro ratti per genotipo o condizione per il seguente protocollo. Sono necessari meno ratti a causa dei loro maggiori volumi cerebrali. Allo stesso modo, l'età degli animali da esperimento può influenzare la resa frazionaria; Altri topi possono essere necessari per età inferiori a 2 mesi. In caso contrario, le procedure descritte si applicano sia alle specie murine che agli animali adulti sani di qualsiasi età. I dati rappresentativi presentati in questo studio hanno utilizzato topi wild-type (C57BL / 6J) (età = 2 mesi; quattro maschi e quattro femmine per replica) ottenuti da una fonte commerciale (vedi Tabella dei materiali).
1. Preparazione sperimentale
NOTA: Questo protocollo richiede ~ 11 h per un singolo ricercatore da completare. Si consiglia vivamente di completare la configurazione da banco (Figura 1), la preparazione del buffer (Tabella 1), il preraffreddamento di centrifughe e rotori a 4 °C e la raccolta e l'etichettatura dei materiali e delle attrezzature necessari (vedere Tabella dei materiali) il giorno prima dell'esecuzione del protocollo, ove applicabile.
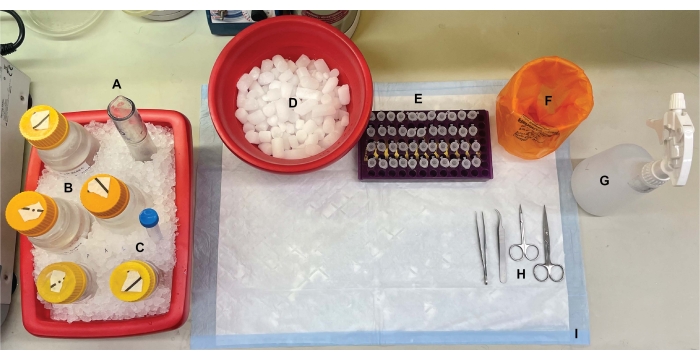
Figura 1: Configurazione da banco. Prima delle dissezioni cerebrali, (A) gli omogeneizzatori di vetro Dounce e (B) tutti i tamponi sono stati raffreddati su ghiaccio. (C) Le soluzioni madre di inibitori della proteasi sono state scongelate sul ghiaccio. Sono stati ottenuti un secondo contenitore di ghiaccio umido per provette da centrifuga, un Dewar di azoto liquido (non mostrato) e (D) un contenitore di ghiaccio secco per la conservazione a breve termine dei campioni congelati in azoto liquido. (E) Le provette per microcentrifuga sono state pre-marcate per tutti i campioni, poiché durante questa procedura sono state raccolte quattro aliquote di ciascun campione di frazione subcellulare per genotipo o condizione (suggerimento che consente di risparmiare tempo: etichettare accuratamente tutte le provette il giorno prima dell'esecuzione dell'esperimento). (F) Un contenitore per rifiuti a rischio biologico appropriato, (G) etanolo al 70%, (H) strumenti chirurgici e (I) un tampone assorbente. Le provette da centrifuga e gli articoli monouso necessari sono stati accantonati per un accesso efficiente durante l'implementazione del protocollo (non mostrato). Fare clic qui per visualizzare una versione ingrandita di questa figura.
- Preparare il piano di lavoro per l'intervento chirurgico e raccogliere le forbici e le pinze necessarie per l'escissione cerebrale (vedi Tabella dei materiali). Pre-etichettare provette da microcentrifuga da 1,5 ml per biopsie a coda di topo e quattro provette per frazione raccolta, come descritto nella Figura 2.
- Procurarsi due contenitori di ghiaccio umido, un contenitore di ghiaccio secco e un matraccio Dewar di azoto liquido da banco.
- Scongelare le soluzioni madre di fenilmetilsulfonilfluoruro (PMSF), pepstatina A, aprotinina e leupeptin su ghiaccio (vedere Tabella dei materiali). Preparare i buffer necessari (tabella 1).
NOTA: Le soluzioni di saccarosio possono essere preparate in anticipo e conservate a 4 °C. Tuttavia, gli inibitori della proteasi (scongelamenti e compresse) devono essere aggiunti freschi a tutti i tamponi all'inizio dell'esperimento a causa dell'instabilità di questi reagenti in soluzioni acquose. Inoltre, tutti i tamponi devono essere preparati con bicchieri senza detersivo e acqua senza detergente per consentire la raccolta di sinaptosomi intatti. - Raffreddare tutti i tamponi e gli omogeneizzatori Dounce in vetro (vedi Tabella dei materiali) sul ghiaccio. Impostare le centrifughe a 4 °C e raffreddare i rotori a 4 °C.
- Aggiungere 14 mL di tampone A (Tabella 1) a un omogeneizzatore Dounce su ghiaccio.
Tabella 1: Composizione dei tamponi di frazionamento subcellulare. Clicca qui per scaricare questa tabella.
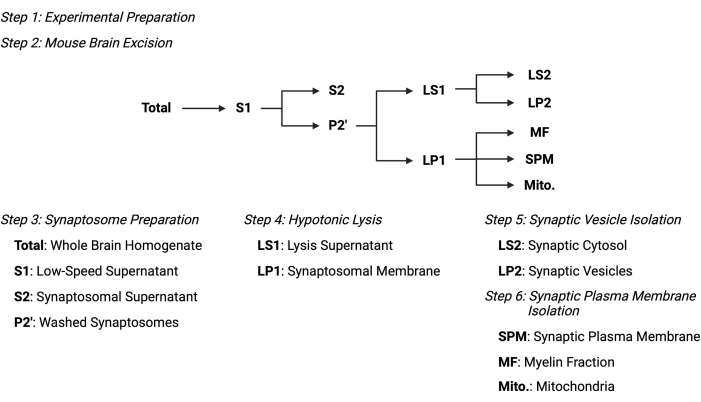
Figura 2: Panoramica del protocollo di frazionamento subcellulare. Schema riassuntivo delle fasi di frazionamento subcellulare e dei campioni raccolti. Fare clic qui per visualizzare una versione ingrandita di questa figura.
2. Escissione cerebrale del topo
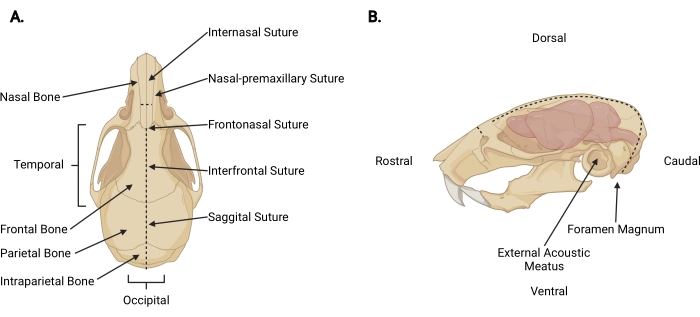
Figura 3: Anatomia craniofacciale . (A) Vista dorsale di un cranio di topo con le relative strutture craniche indicate. (B) Vista laterale sinistra del cranio e del cervello di un topo con le relative strutture craniche e direzioni anatomiche indicate. Le linee tratteggiate rappresentano le posizioni in cui devono essere eseguite le incisioni. Fare clic qui per visualizzare una versione ingrandita di questa figura.
- Anestetizzare profondamente ogni topo con isofluorano al 100% in una camera di anestesia situata in una cappa aspirante o in un armadio di biosicurezza utilizzando un metodo a goccia aperta34. Sacrificare ogni topo con la lussazione della colonna cervicale seguita rapidamente dalla decapitazione. Alternare genotipi o gruppi sperimentali per ogni sacrificio e dissezione12. Ottenere biopsie della coda dopo l'eutanasia asportando 2 mm della punta distale della coda con forbici sottili. Conservare il tessuto per la genotipizzazione.
- Spruzzare la testa decapitata con etanolo al 70% per evitare che i capelli aderiscano al tessuto e agli strumenti chirurgici durante la dissezione.
- Inserire delle forbici sottili sotto la pelle all'altezza dell'incisione di decapitazione a una profondità pericranica e fare un'incisione mediosagittale fino alla sutura interna (Figura 3A) per ritrarre il cuoio capelluto dal cranio.
- Lavorando dall'area occipitale verso ogni aspetto temporale, tagliare la fascia e il muscolo per esporre la superficie esterna del cranio oltre ogni meato acustico esterno (Figura 3B).
- Fissare il cuoio capelluto e l'aspetto rostrale del cranio con la mano non dominante. Con l'altro, inserire forbici sottili di 2 mm nel lato caudale del forame magno, dove il midollo spinale è visibile in uscita. Fare un'incisione della linea mediana fino a quando le forbici raggiungono la superficie interna dell'osso intraparietale (Figura 3; linee tratteggiate).
NOTA: Durante l'incisione iniziale, le forbici devono essere parallele al midollo spinale con pressione applicata verso la superficie interna del cranio per evitare danni al tronco cerebrale e al cervelletto. - Cambia l'angolo delle forbici in modo che le lame corrano parallele alla superficie dorsale del cranio. Continuare a far avanzare rostralmente l'incisione mediosagittale attraverso le ossa parietali e frontali, usando le suture sagittali e interfrontali come guida. Utilizzare una pressione costante verso l'alto per evitare danni alla corteccia. Terminare l'incisione appena oltre la sutura interna (Figura 3A).
- Fare una piccola incisione perpendicolare (~3 mm) all'osso nasale, rostrale alla sutura interna, posizionando le forbici perpendicolari al cranio con ogni lama posizionata su una sutura nasale-premascellare e facendone un taglio uniforme (Figura 3; linee tratteggiate).
NOTA: Questo passaggio aumenterà la facilità di retrazione del cranio e sarà fondamentale per la raccolta del bulbo olfattivo se quest'area è di interesse. - Mentre si fissa l'aspetto rostrale, utilizzare un lato di un paio di pinze strutturate per sollevare delicatamente il cranio dal cervello, quindi lateralmente e ventralmente. Ripeti lungo la linea mediana secondo necessità, poi per l'altro emisfero fino a quando l'intera superficie del cervello è esposta.
- Usando una pinza curva o una spatola fine, sollevare delicatamente il lato rostrale del cervello. Tagliare i nervi ottici e cranici per completare l'escissione dal cranio.
- Per ogni condizione, raccogliere da cinque a otto cervelli di topo insieme nell'omogeneizzatore Dounce in vetro refrigerato contenente 14 ml di tampone A (Tabella 1).
3. Preparazione del sinaptosoma
Nota : gli schemi di questa procedura sono illustrati nella Figura 4.
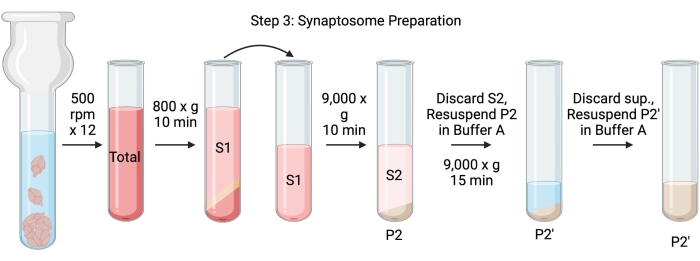
Figura 4: Preparazione del sinaptosoma. Schema della fase 3, la generazione dei sinaptosomi (P2'). Fare clic qui per visualizzare una versione ingrandita di questa figura.
- Omogeneizzare il cervello usando un omogeneizzatore Dounce in vetro in 12 passaggi up-down a 500 rpm (totale). Fare una breve pausa ad ogni discesa per garantire un'omogeneizzazione completa del tessuto. Omogeneizzare preferenzialmente in un bagno di ghiaccio per evitare il riscaldamento e la denaturazione delle proteine. Prelevare 5 μL di aliquote per la determinazione della concentrazione proteica mediante il saggio dell'acido bicinconinico (BCA, vedere Tabella dei materiali). Prendi 100 μL di aliquote di lisato del cervello intero per western blot (WB). Per questo e tutti i campioni successivi (Figura 2), prendere due aliquote per BCA e due aliquote per WB. Congelare tutte le aliquote raccolte in azoto liquido e conservarle a -80 °C.
- Centrifugare l'omogenato cerebrale totale in una provetta centrifuga a fondo rotondo ad alta velocità (14 ml) (vedi tabella dei materiali) a 800 x g per 10 minuti a 4 °C per ottenere il surnatante (S1). Trasferire S1 in una nuova provetta da centrifuga, lasciando dietro di sé il pellet (P1), che contiene cellule e nuclei intatti. Evitare di pipettare il pellet soffice, bianco, sciolto e superficiale. Prendi 2 x 5 μL di S1 per BCA e 2 x 100 μL di S1 per WB.
- Spingere S1 a 9.000 x g per 15 minuti a 4 °C per ottenere il surnatante sinaptosomiale (S2) e il pellet di sinaptosoma grezzo (P2). Prendi 2 x 10 μL di S2 per BCA e 2 x 500 μL di S2 per WB. Scartare il surnatante dopo aver ottenuto le aliquote e procedere alla fase successiva con il pellet.
- Risospendere P2 in 3 mL di Tampone A ghiacciato con inibitori della proteasi e centrifugare a 9.000 x g per 15 minuti a 4 °C per ottenere il surnatante (S2') e i sinaptosomi lavati (P2'). Scartare il surnatante e conservare il pellet.
- Risospendere P2' in 3 ml di tampone A. Evitare di risospendere la porzione rosso scuro sul fondo del pellet, che contiene principalmente mitocondri. Prendi 2 x 20 μL di P2' per BCA e 2 x 100 μL di P2' per WB.
NOTA: Questo può essere ottenuto mescolando delicatamente i bordi e la superficie del pellet per risospendere i sinaptosomi lavati di bianco mentre si dirige la punta della pipetta lontano dal centro rosso del pellet.
4. Lisi ipotonica
Nota : gli schemi di questa procedura sono illustrati nella Figura 5.
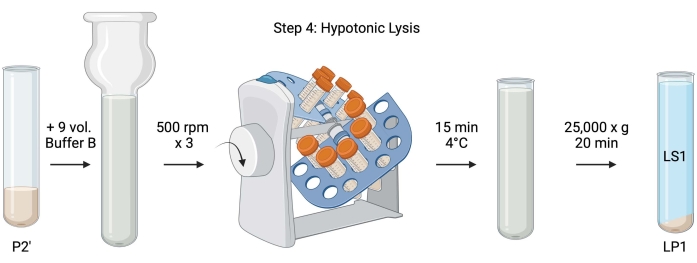
Figura 5: Lisi ipotonica. Schema della fase 4, la lisi ipotonica dei sinaptosomi per generare la lisi supernatante (LS1) e le frazioni di membrana sinaptosomiale (LP1). Fare clic qui per visualizzare una versione ingrandita di questa figura.
- Per la lisi ipotonica dei sinaptosomi lavati, aggiungere 9 volumi di Buffer B raffreddato (Tabella 1) a P2' risospeso (~27 ml). Omogeneizzare i sinaptosomi in un omogeneizzatore Dounce di vetro (tre passaggi su-giù a 500 giri / min).
- Trasferire i campioni in provette coniche da centrifuga con tappo da 50 ml. Ruotarli su un revolver tubolare in una cella frigorifera a 4 °C per 15 minuti.
- Centrifugare lisato P2' a 25.000 x g per 20 minuti a 4 °C per ottenere il surnatante di lisi (LS1) e il pellet di lisi contenente membrane sinaptosomiali (LP1). Prendi 2 x 50 μL di LS1 per BCA e 2 x 400 μL di LS1 per WB. Trasferire LS1 in una provetta da centrifuga con tappo per l'ultracentrifugazione (vedere Tabella dei materiali).
5. Isolamento della vescicola sinaptica
Nota : gli schemi di questa procedura sono illustrati nella Figura 6.
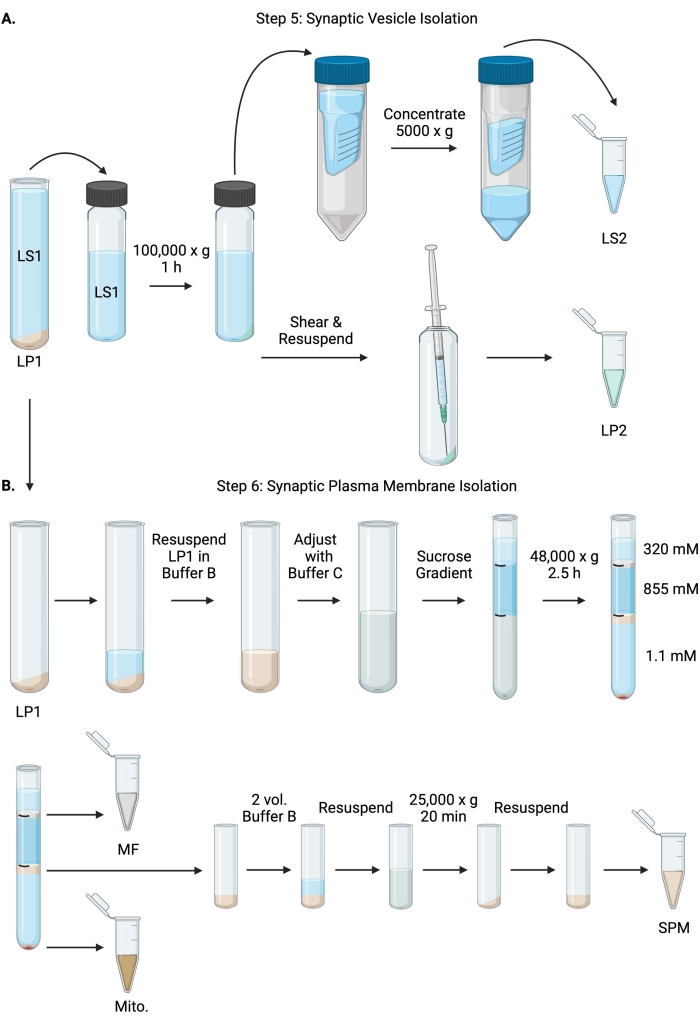
Figura 6: Isolamento della vescicola sinaptica e isolamento della membrana plasmatica sinaptica. (A) Schema della fase 5, l'isolamento delle frazioni di citosol sinaptico (LS2) e vescicola sinaptica (LP2) e (B) fase 6, la generazione di mielina (MF), membrana plasmatica sinaptica (SPM) e frazioni mitocondriali (Mito.) dopo l'ultracentrifugazione dei gradienti di saccarosio. Fare clic qui per visualizzare una versione ingrandita di questa figura.
- Centrifugare LS1 in rotore ultracentrifugo ad angolo fisso (vedi Tabella dei materiali) a 100.000 x g per 60 minuti a 4 °C per ottenere surnatante citosol sinaptico (LS2) e pellet di vescicola sinaptica (LP2). LP2 sarà piccolo, traslucido e fortemente aderente al lato del tubo della centrifuga.
- Risospendere LP2 in 500 μL di tampone A. Utilizzando un ago da 23 G e una siringa da 1 ml, tagliare LP2 con triturazione delicata. Prendi 2 x 10 μL di LP2 per BCA e 2 x 250 μL di LP2 per WB.
- Trasferire LS2 (~30 ml) alle unità filtranti centrifughe con un cutoff di 10 kDa (vedere Tabella dei materiali).
NOTA: Se le proteine inferiori a 10 kDa sono di interesse, sono disponibili unità di filtro centrifughe di taglio di 4 kDa, ma comporteranno tempi di rotazione più lunghi. - Concentrare LS2 a circa 0,5 mL ruotando a 5000 x g per un massimo di 1 ora a 4 °C. Prendere 2 x 10 μL di LS2 concentrato per BCA e 2 x 250 μL di LS2 concentrato per WB. Dopo aver avviato la rotazione, procedere direttamente al passaggio 6.1.
6. Isolamento della membrana plasmatica sinaptica
- Risospendere LP1 (punto 4.3) in 1 mL di buffer B (Tabella 1). Prendi 2 x 10 μL di LP1 per BCA e 2 x 50 μL di LP1 per WB. Regolare l'LP1 rimanente su un volume finale di 7,5 mL e una concentrazione finale di saccarosio di 1,1 M con il tampone B e il tampone C (tabella 1).
- Trasferire 7,5 mL di LP1 risospeso in un tubo da ultracentrifuga da 14 mL (vedere Tabella dei materiali). Sovrapporre attentamente LP1 con 3,75 mL di tampone D (Tabella 1), quindi sovrapporre con 1,25 mL di tampone A (o un volume maggiore da riempire appena sotto la parte superiore del tubo della centrifuga). Evitare il pipettaggio lungo il lato del tubo, che interromperà le interfacce del gradiente di saccarosio. Dopo aver sovrapposto ogni frazione di saccarosio, contrassegnare la parte superiore della soluzione con una penna. Bilanciare i tubi per l'ultracentrifugazione in base al peso, non al volume, con l'aggiunta a goccia del tampone A entro 10 mg. Centrifugare a 48.000 x g per 2,5 ore a 4 °C in un rotore ultracentrifugo a benna oscillante (vedi Tabella dei materiali).
- Acquisire immagini dei gradienti intatti dopo l'ultracentrifugazione per documentare la distinzione di ciascuna interfaccia di saccarosio e il successo del frazionamento.
- Rimuovere con cautela lo strato superficiale di 320 mM di saccarosio (tampone A). Recuperare la frazione mielinica (MF) all'interfaccia saccarosio 320 mM/855 mM in un volume di 800 μL. Recuperare la frazione della membrana plasmatica sinaptica (SPM) all'interfaccia di saccarosio 855 mM/1,1 M in un volume di 1.000 μL. Pipettare ogni frazione dalla parete del tubo in modo circolare per garantire che la frazione completa venga raccolta. Aspirare con cautela il saccarosio rimanente e recuperare il pellet mitocondriale (Mito.) riassumendo in 200 μL di tampone B. Prendi 2 x 100 μL di MF per BCA e 2 x 10 μL di Mito. per BCA; dividere il resto di MF e Mito. campioni a metà per WB.
- Diluire la frazione SPM con 2 volumi di tampone B (~2 ml), quindi centrifugare in un rotore ad angolo fisso in una provetta da centrifuga da 3,5 mL (vedere Tabella dei materiali) a 25.000 x g per 20 minuti a 4 °C. Scartare il surnatante e risospendere il pellet SPM nel tampone A per un volume finale di 250 μL. Prendi 2 x 5 μL di SPM per BCA e dividi l'SPM rimanente a metà per WB.
- Eseguire un BCA per determinare la concentrazione proteica di ciascun campione, tenendo conto del volume aliquota variabile.
NOTA: Per l'analisi WB, la concentrazione proteica di lavoro suggerita per tutte le frazioni subcellulari è di 2 μg/μL (o massima quanto ottenibile per LS1 e MF).
Il metodo presentato si traduce in 11 frazioni subcellulari cerebrali che possono essere sottoposte a ulteriore purificazione e varie forme di analisi a valle35,36. Il metodo gold standard per valutare la qualità dei sinaptosomi, SVs23 e altri componenti è la microscopia elettronica (EM) (Figura 7). L'immunoblotting quantitativo per proteine presenti in specifiche frazioni subcellulari può anche essere eseguito per valutare i marcatori di purezza della frazione (Figura 8). Ad esempio, l'analisi immunoblot delle frazioni rivela l'arricchimento di N-caderina (CDH2, nome UniProt) nella frazione di membrana plasmatica sinaptica (SPM), α-sinucleina (SYUA) nel citosol sinaptico (LS2), sinaptofisina (SYPH) nella frazione vescicola sinaptica (LP2) e proteina basica della mielina (MBP) nella frazione mielinica (MF) rispetto ai livelli proteici nell'omogenato iniziale dell'intero cervello (totale) (Figura 8 ). Una volta stabilita la purezza della frazione (ad esempio, si noti l'assenza di CDH2 nella frazione LS2 o l'aumento di molte volte di SYPH nella frazione LP2), l'immunoblotting quantitativo può essere utilizzato per determinare la localizzazione delle proteine di interesse o interrogare le differenze nella distribuzione delle proteine tra genotipi o trattamenti. Comprendere la localizzazione subcellulare delle proteine sinaptiche può consentire la dissezione di funzioni proteiche precedentemente non descritte. Inoltre, questo metodo può chiarire i difetti del traffico o la disfunzione sinaptica negli stati patologici, specialmente se abbinato a saggi funzionali. Ad esempio, il nostro team ha utilizzato questo metodo per identificare un pool di tioesterasi 1 della proteina palmitoil enzimaticamente attiva che è arricchita nel citosol sinaptico19.
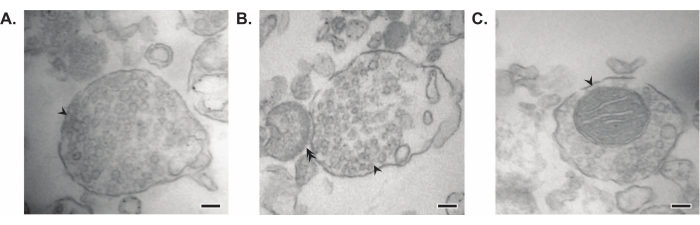
Figura 7: Microscopia elettronica (EM) dei sinaptosomi. (A) Immagine EM rappresentativa di un sinaptosoma contenente vescicole sinaptiche (freccia). (B) Immagine EM rappresentativa di un sinaptosoma con componenti pre- (freccia) e post-sinaptica (doppia freccia). (C) Immagine EM rappresentativa di un sinaptosoma contenente vescicole sinaptiche e un mitocondrio (freccia) (barre della scala = 100 nm). Fare clic qui per visualizzare una versione ingrandita di questa figura.
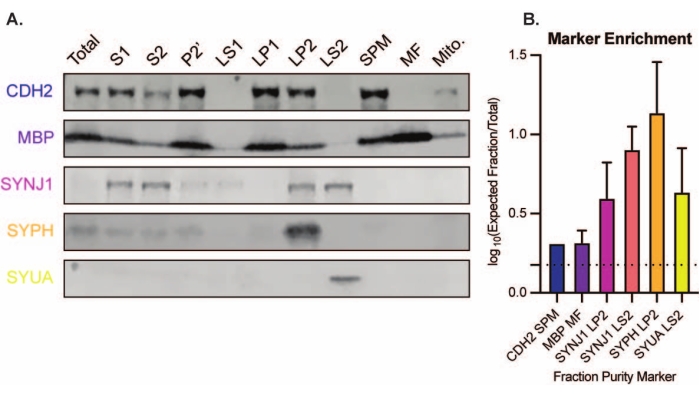
Figura 8: Analisi immunoblot delle frazioni subcellulari. (A) I marcatori di purezza della frazione subcellulare (indicati con la nomenclatura UniProt) sono opportunamente localizzati rispetto all'omogenato dell'intero cervello (totale): N-caderina (CDH2) nella frazione sinaptica della membrana plasmatica (SPM), sinaptofisina 1 (SYPH) e sinaptojanina 1 (SYNJ1) nella frazione arricchita di vescicole sinaptiche (LP2), α-sinucleina (SYUA) nel citosol sinaptico (LS2), e la proteina basica della mielina (MBP) nella frazione mielinica (MF). (B) L'analisi di quantificazione immunoblot rivela l'arricchimento (variazione di piega rispetto al totale) dei marcatori di purezza frazionaria. I dati sono rappresentati come media ± deviazione standard su una scala log10. La linea tratteggiata indica un cambiamento di 1,5 volte (y = 0,176) (n = 3 esperimenti replicati con 8 topi wild-type; età = 2 mesi; n = 4-5 macchie per SYPH, SYUA, MBP, con n = 3 valori tracciati precedentemente pubblicati da Gorenberg et al.19; n = 5 per SYNJ1; n = 1 per CDH2). Fare clic qui per visualizzare una versione ingrandita di questa figura.
Nei loro studi seminali, Whittaker e colleghi hanno utilizzato quattro criteri morfologici per identificare i sinaptosomi: (1) le strutture hanno una membrana plasmatica sigillata; (2) le strutture contengono SV simili a quelle delle terminazioni nervose e varicosità in situ per dimensioni e numero; (3) le strutture possiedono uno o più piccoli mitocondri; e (4) la membrana presinaptica è frequentemente aderente a una componente post-sinaptica11,12,13. Sebbene i primi due criteri si applichino generalmente a ogni metodo di isolamento, nei protocolli più recenti descritti in questo articolo, non tutti i sinaptosomi risultanti avranno mitocondri e terminali post-sinaptici collegati. Circa il 60% dei sinaptosomi avrà mitocondri e si stima che solo fino al 15% abbia attaccato terminali post-sinaptici37. Se le componenti post-sinaptiche sono di particolare interesse, l'uso di un tampone di omogeneizzazione isotonico simile a Krebs e la filtrazione a pressione per l'arricchimento sono noti per produrre alte concentrazioni di sinaptosomi con terminali post-sinaptici (chiamati anche sinaptoneurosomi)22,38.
Il metodo di sacrificare l'animale può influire sulla qualità dei sinaptosomi e delle sottofrazioni sinaptiche. Gli animali adulti sacrificati con un metodo di eutanasia che non richiede l'anestesia si tradurranno nella migliore qualità della frazione. Inoltre, i cervelli dovrebbero essere appena sezionati, non congelati e omogeneizzati usando un rapporto 1:10 di tampone di omogeneizzazione (peso / volume) per le frazioni sinaptiche più vitali22. Il cervello ha una popolazione eterogenea di sinapsi che possono essere differenziate dal tipo di neurotrasmettitori che trasportano. La formazione del sinaptosoma non è generalmente influenzata dal tipo di sinapsi o dal contenuto di neurotrasmettitori13. Un'eccezione sono le fibre muschiose nel cervelletto, che sono note per essere interrotte in condizioni ottimali per ottenere sinaptosomi dal resto del cervello39,40. Pertanto, la rimozione del cervelletto prima dell'omogeneizzazione del cervello è raccomandata se l'esclusione di questa regione non influisce sull'obiettivo sperimentale. Se interessati a isolare i sinaptosomi di un particolare carattere di neurotrasmettitore, le aree del cervello che sono arricchite per i neuroni contenenti il neurotrasmettitore di interesse possono prima essere isolate. Tuttavia, questo approccio imporrà limitazioni alla resa della frazione finale, a seconda delle dimensioni della regione di interesse (anche l'età degli animali è quindi una considerazione). Esistono metodi immunochimici per l'isolamento dei sinaptosomi specifici dei neurotrasmettitori, ma la vitalità e la resa saranno significativamente compromesse22. Se la valutazione della vitalità metabolica del sinaptosoma è importante, può essere impiegata la misurazione del rilascio di neurotrasmettitori 41,42 o alcuni saggi enzimatici43.
I contaminanti comuni nei preparati di sinaptosomi includono microsomi, mitocondri liberi, SV e membrane neuronali e gliali. La contaminazione può essere ridotta aumentando il numero di lavaggi alle frazioni P1 e P222 ed evitando la risospensione del pellet mitocondriale rosso nelle fasi successive. Negli esperimenti in cui la vitalità metabolica e il tempo sono cruciali, ridurre il numero di lavaggi e utilizzare gradienti di Ficoll o Percoll su gradienti di saccarosio sarà utile44,45,46. Questi metodi riducono anche significativamente la contaminazione. Il protocollo originale di Whittaker produceva SV di alta qualità. Un'ulteriore ottimizzazione di Nagy et al.23, inclusa in questo metodo, produce SV con notevole omogeneità e purezza senza compromettere significativamente la resa36. Se specifici sottotipi di SV sono di interesse, come le SV glutammatergiche (contenenti VGLUT-1) o GABAergiche (contenenti VGAT-1), l'immunoisolamento con anticorpi specifici può essere eseguito47,48. Sono disponibili anche metodi alternativi per isolare i CCV dai sinaptosomi, che, a causa della densità differenziale, potrebbero non essere presenti nella stessa interfaccia delle SV ottenute con questo metodo20,49,50.
Nel complesso, l'attuale protocollo per isolare i componenti sinaptici può essere ulteriormente ottimizzato per ottenere frazioni con maggiore omogeneità e vitalità in base alla qualità e alla quantità del tessuto cerebrale sorgente e agli obiettivi sperimentali. Per ulteriori dettagli sulla risoluzione dei problemi, si dovrebbe fare riferimento ai capitoli dei libri di Dunkley e Robinson22 e Ganzella et al.36.
Gli autori non hanno nulla da rivelare.
Ringraziamo P. Colosi per la preparazione delle immagini EM. Questo lavoro è stato sostenuto dal National Institutes of Health (R01 NS064963, SSC; R01 NS110354, SSC; R01 NS083846, SSC; R21 NS094971, SSC; T32 NS007224, SMT; T32 NS041228, SMT), Dipartimento della Difesa degli Stati Uniti (W81XWH-17-1-0564, SSC; W81XWH-19-1-0264, VDJ), Aligning Science Across Parkinson's (ASAP) Collaborative Research Network (SSC) e Michael J. Fox Foundation Target Advancement Program (MJFF-020160, SSC & VDJ). Abbiamo creato illustrazioni grafiche utilizzando BioRender.com.
| Name | Company | Catalog Number | Comments |
| 1 mL TB Syringe | BD | 309649 | |
| 1.5 mL Eppendorf Tubes | USA Scientific | 1415-2500 | |
| 14 mL, Open-Top Thinwall Ultra-Clear Tube | Beckman Coulter | 344060 | Compatible with SW 40 Ti |
| 23 Gauge Precision Glide Hypodermic Needle | BD | 305145 | |
| 26.3 mL, Polycarbonate Bottle with Cap Assembly | Beckman Coulter | 355618 | Compatible with Ti70 |
| 3.5 mL, Open-Top Thickwall Polypropylene Tube | Beckman Coulter | 349623 | Compatible with TLA-100.3 |
| 50 mL Falcon Tubes | Fisher Scientific | 14-432-22 | |
| Amicon Ultra-15 Centrifugal Filter Unit | Millipore Sigma | UFC901024 | |
| Aprotinin | Sigma-Aldrich | A6279 | 1 mg/mL in diH2O |
| Avanti J-26 XP Centrifuge | Beckman Coulter | B22984 | <26,000 rpm |
| Benchtop HDPE Dewar Flask | Thermo Scientific | 5028U19 | |
| C57BL/6J Mice | The Jackson Labs | 000664 | |
| Centrifuge 5810R | Eppendorf | EP022628168 | <14,000 rpm |
| complete, Mini, EDTA-free Protease Inhibitor Cocktail Tablets | Roche | 11873580001 | Add 1 tablet per 50 mL of solution |
| Curved Forceps | Fine Science Tools | 11273-20 | |
| Fine Surgical Scissors | Fine Science Tools | 8r | |
| Glas-Col Tissue Homogenizing System | Cole-Parmer | UX-04369-15 | |
| Graefe Forceps | Fine Science Tools | 11650-10 | |
| High-Speed Polycarbonate Round Bottom Centrifuge Tubes | ThermoFisher | 3117-0500 | Compatible with JA20 |
| Isofluorane | Henry Schein Animal Health | NDC 11695-6776-2 | |
| JA-20 Rotor | Beckman Coulter | 334831 | |
| Leupeptin | American Bio | AB01108 | 1 mg/mL in diH2O |
| N-[2-Hydroxyethyl] piperazine-N’-[2-ethanesulfonic acid] (HEPES) | American Bio | AB00892 | |
| Optima L-80 XP Ultracentrifuge | Beckman Coulter | <100,000 rpm | |
| Optima TLX Ultracentrifuge | Beckman Coulter | <120,000 rpm | |
| Pepstatin A | Thermo Scientific | 78436 | 1 mg/mL in DMSO |
| Phenylmethylsulfonyl fluoride (PMSF) | American Bio | AB01620 | |
| Pierce BCA Protein Assay Kit | Thermo Scientific | 23335 | For determination of protein concentration |
| Pipette Tips | |||
| Serological Pipettes | |||
| Sucrose | Sigma-Aldrich | S0389 | |
| Surgical Scissors | Fine Science Tools | 14002-12 | |
| SW 40 Ti Swinging-Bucket Rotor | Beckman Coulter | 331301 | |
| Teflon-Coated Pestle and Mortar Tissue Grinder | Thomas Scientific | 3431D94 | |
| Ti70 Rotor | Beckman Coulter | 337922 | |
| TLA-100.3 Rotor | Beckman Coulter | 349490 | |
| Tube Revolver | Dot Scientific | DTR-02VS |
- Kandel, E. R., Schwartz, J. H., Jessell, T. M., Siegelbaum, S. A., Hudspeth, A. J., Education, A. J. Synaptic Transmission. Principles of Neural Science, Fifth Edition. , (2014).
- Lepeta, K., et al. Synaptopathies: synaptic dysfunction in neurological disorders - A review from students to students. Journal of Neurochemistry. 138 (6), 785-805 (2016).
- Südhof, T. C., Malenka, R. C. Understanding synapses: Past, present, and future. Neuron. 60 (3), 469-476 (2008).
- Südhof, T. C. The molecular machinery of neurotransmitter release (Nobel lecture). Angewandte Chemie International Edition. 53 (47), 12696-12717 (2014).
- Jahn, R., Boyken, J., Pfaff, D. W. Molecular Regulation of Synaptic Release. Neuroscience in the 21st Century: From Basic to Clinical. , 351-401 (2013).
- Xiong, H., Gendelman, H. E. . Current Laboratory Methods in Neuroscience Research. , (2014).
- Azevedo, F. A., et al. Equal numbers of neuronal and nonneuronal cells make the human brain an isometrically scaled-up primate brain. Journal of Comparative Neurology. 513 (5), 532-541 (2009).
- Herculano-Houzel, S. The remarkable, yet not extraordinary, human brain as a scaled-up primate brain and its associated cost. Proceedings of the National Academy of Sciences of the United States of America. 109, 10661-10668 (2012).
- Herculano-Houzel, S., Lent, R. Isotropic fractionator: A simple, rapid method for the quantification of total cell and neuron numbers in the brain. Journal of Neuroscience. 25 (10), 2518-2521 (2005).
- Herculano-Houzel, S., Mota, B., Lent, R. Cellular scaling rules for rodent brains. Proceedings of the National Academy of Sciences of the United States of America. 103 (32), 12138-12143 (2006).
- Gray, E. G., Whittaker, V. P. The isolation of nerve endings from brain: An electron-microscopic study of cell fragments derived by homogenization and centrifugation. Journal of Anatomy. 96, 79-88 (1962).
- Gray, E. G., Whittaker, V. P. The isolation of synaptic vesicles from the central nervous system. Journal of Physiology. 153, 35-37 (1960).
- Whittaker, V. P. Thirty years of synaptosome research. Journal of Neurocytology. 22 (9), 735-742 (1993).
- Jahn, R., Fasshauer, D. Molecular machines governing exocytosis of synaptic vesicles. Nature. 490, 201-207 (2012).
- Whittaker, V. P., Michaelson, I. A., Kirkland, R. J. The separation of synaptic vesicles from nerve-ending particles ('synaptosomes). Biochemical Journal. 90 (2), 293-303 (1964).
- De Robertis, E., Rodriguez De Lores Arnaiz, G., Pellegrino De Iraldi, A. Isolation of synaptic vesicles from nerve endings of the rat brain. Nature. 194, 794-795 (1962).
- De Robertis, E., Pellegrino De Iraldi, A., Rodriguez, G., Gomez, C. J. On the isolation of nerve endings and synaptic vesicles. The Journal of Biophysical and Biochemical Cytology. 9 (1), 229-235 (1961).
- Zimmermann, H., Whittaker, V. P., Murphy, K. M. The Discovery of the Synaptosome and Its Implications. Synaptosomes. , 9-26 (2018).
- Gorenberg, E. L., et al. Identification of substrates of palmitoyl protein thioesterase 1 highlights roles of depalmitoylation in disulfide bond formation and synaptic function. PLoS Biology. 20 (3), 3001590 (2022).
- Vidyadhara, D. J., et al. Dopamine transporter and synaptic vesicle sorting defects initiate auxilin-linked Parkinson's disease. bioRxiv. , (2022).
- Schrimpf, S. P., et al. Proteomic analysis of synaptosomes using isotope-coded affinity tags and mass spectrometry. Proteomics. 5 (10), 2531-2541 (2005).
- Dunkley, P. R., Robinson, P. J., Murphy, K. M. Synaptosome Preparations: Which Procedure Should I Use. Synaptosomes. , 27-53 (2018).
- Nagy, A., Baker, R. R., Morris, S. J., Whittaker, V. P. The preparation and characterization of synaptic vesicles of high purity. Brain Research. 109 (2), 285-309 (1976).
- Takamori, S., et al. Molecular anatomy of a trafficking organelle. Cell. 127 (4), 831-846 (2006).
- Wagner, J. A., Kelly, R. B. Topological organization of proteins in an intracellular secretory organelle: the synaptic vesicle. Proceedings of the National Academy of Sciences of the United States of America. 76 (8), 4126-4130 (1979).
- Jahn, R., Schiebler, W., Ouimet, C., Greengard, P. A 38,000-dalton membrane protein (p38) present in synaptic vesicles. Proceedings of the National Academy of Sciences of the United States of America. 82 (12), 4137-4141 (1985).
- Binotti, B., Jahn, R., Pérez-Lara, &. #. 1. 9. 3. ;. An overview of the synaptic vesicle lipid composition. Archives of Biochemistry and Biophysics. 709, 108966 (2021).
- Siegel, D. P., Ware, B. R. Electrokinetic properties of synaptic vesicles and synaptosomal membranes. Biophysical Journal. 30 (1), 159-172 (1980).
- Whittaker, V. P., Michaelson, I. A., Kirkland, R. J. The separation of synaptic vesicles from disrupted nervending particles. Biochemical Pharmacology. 12 (3), 300-302 (1963).
- Clementi, F., Whittaker, V. P., Sheridan, M. N. The yield of synaptosomes from the cerebral cortex of guinea pigs estimated by a polystyrene bead "tagging" procedure. Zeitschrift für Zellforschung und Mikroskopische Anatomie. 72, 126-138 (1966).
- Carlson, S. S., Wagner, J. A., Kelly, R. B. Purification of synaptic vesicles from elasmobranch electric organ and the use of biophysical criteria to demonstrate purity. Biochemistry. 17 (7), 1188-1199 (1978).
- Huttner, W. B., Schiebler, W., Greengard, P., De Camilli, P. Synapsin I (protein I), a nerve terminal-specific phosphoprotein. III. Its association with synaptic vesicles studied in a highly purified synaptic vesicle preparation. Journal of Cell Biology. 96 (5), 1374-1388 (1983).
- Hawkins, P., et al. A guide to defining and implementing protocols for the welfare assessment of laboratory animals: eleventh report of the BVAAWF/FRAME/RSPCA/UFAW Joint Working Group on Refinement. Laboratory Animals. 45 (1), 1-13 (2011).
- Risling, T. E., Caulkett, N. A., Florence, D. Open-drop anesthesia for small laboratory animals. Canadian Veterinary Journal. 53 (3), 299-302 (2012).
- Deutsch, C., Drown, C., Rafalowska, U., Silver, I. A. Synaptosomes from rat brain: Morphology, compartmentation, and transmembrane pH and electrical gradients. Journal of Neurochemistry. 36 (6), 2063-2072 (1981).
- Ganzella, M., Ninov, M., Riedel, D., Jahn, R. Isolation of synaptic vesicles from mammalian brain. Methods in Molecular Biology. 2417, 131-145 (2022).
- Dunkley, P. R., et al. A rapid Percoll gradient procedure for isolation of synaptosomes directly from an S1 fraction: homogeneity and morphology of subcellular fractions. Brain Research. 441 (1-2), 59-71 (1988).
- Schwartz, R. D., Skolnick, P., Hollingsworth, E. B., Paul, S. M. Barbiturate and picrotoxin-sensitive chloride efflux in rat cerebral cortical synaptoneurosomes. FEBS Letters. 175 (1), 193-196 (1984).
- Pittaluga, A., Thellung, S., Maura, G., Raiteri, M. Characterization of two central AMPA-preferring receptors having distinct location, function and pharmacology. Naunyn-Schmiedeberg's Archives of Pharmacology. 349 (6), 555-558 (1994).
- Israël, M., Whittaker, V. P. The isolation of mossy fibre endings from the granular layer of the cerebellar cortex. Experientia. 21 (6), 325-326 (1965).
- Khvotchev, M., Lonart, G., Südhof, T. C. Role of calcium in neurotransmitter release evoked by alpha-latrotoxin or hypertonic sucrose. Neuroscience. 101 (3), 793-802 (2000).
- Lonart, G., Janz, R., Johnson, K. M., Südhof, T. C. Mechanism of action of rab3A in mossy fiber LTP. Neuron. 21 (5), 1141-1150 (1998).
- Nicholls, D. G., Sihra, T. S. Synaptosomes possess an exocytotic pool of glutamate. Nature. 321 (6072), 772-773 (1986).
- Dunkley, P. R., Jarvie, P. E., Robinson, P. J. A rapid Percoll gradient procedure for preparation of synaptosomes. Nature Protocols. 3 (11), 1718-1728 (2008).
- Cotman, C. W., Matthews, D. A. Synaptic plasma membranes from rat brain synaptosomes: Isolation and partial characterization. Biochimica et Biophysica Acta. 249 (2), 380-394 (1971).
- Booth, R. F., Clark, J. B. A rapid method for the preparation of relatively pure metabolically competent synaptosomes from rat brain. Biochemical Journal. 176 (2), 365-370 (1978).
- Takamori, S., Riedel, D., Jahn, R. Immunoisolation of GABA-specific synaptic vesicles defines a functionally distinct subset of synaptic vesicles. Journal of Neuroscience. 20 (3), 4904-4911 (2000).
- Burger, P. M., et al. Synaptic vesicles immunoisolated from rat cerebral cortex contain high levels of glutamate. Neuron. 3 (6), 715-720 (1989).
- Blondeau, F., et al. Tandem MS analysis of brain clathrin-coated vesicles reveals their critical involvement in synaptic vesicle recycling. Proceedings of the National Academy of Sciences of the United States of America. 101 (11), 3833-3838 (2004).
- Maycox, P. R., Link, E., Reetz, A., Morris, S. A., Jahn, R. Clathrin-coated vesicles in nervous tissue are involved primarily in synaptic vesicle recycling. Journal of Cell Biology. 118 (6), 1379-1388 (1992).
ABOUT JoVE
Copyright © 2024 MyJoVE Corporation. All rights reserved