
Summary
Abstract
Introduction
Protocol
Representative Results
Discussion
Acknowledgements
Materials
References
Immunology and Infection
הערכה סימולטנית של קרבה, מספר חלוקה ופנוטיפ באמצעות ציטומטריית זרימה עבור תאי גזע המטופויטיים ותאי אב
מוצגת כאן טכניקה מבוססת ציטומטריית זרימה המאפשרת למדוד בו זמנית את מספר חלוקות התא, פנוטיפ תאי פני השטח וקרבה תאית. ניתן לבדוק תכונות אלה באופן סטטיסטי באמצעות מסגרת מבוססת תמורות.
טכניקות מעטות יכולות להעריך פנוטיפ וגורל עבור אותו תא בו זמנית. רוב הפרוטוקולים הנוכחיים המשמשים לאפיון פנוטיפ, למרות שהם מסוגלים ליצור מערכי נתונים גדולים, מחייבים את הרס התא המעניין, מה שהופך אותו לבלתי אפשרי להעריך את גורלו התפקודי . לכן קשה לתאר מערכות התמיינות ביולוגיות הטרוגניות כמו המטופויזה. בהתבסס על צבעי מעקב אחר חלוקת תאים, פיתחנו פרוטוקול לקביעת קרבה, מספר חלוקה ומצב התמיינות בו זמנית עבור אבות המטופויטיים בודדים רבים. פרוטוקול זה מאפשר להעריך את פוטנציאל התמיינות ex vivo של אבות המטופויטיים מורינים ובני אדם, מבודדים ממקורות ביולוגיים שונים. יתר על כן, מכיוון שהוא מבוסס על ציטומטריית זרימה ומספר מוגבל של ריאגנטים, הוא יכול לייצר במהירות כמות גדולה של נתונים, ברמת התא הבודד, באופן זול יחסית. אנו מספקים גם את הצינור האנליטי לניתוח תא יחיד, בשילוב עם מסגרת סטטיסטית חזקה. מכיוון שפרוטוקול זה מאפשר קישור של חלוקת תאים והתמיינות ברמת התא הבודד, ניתן להשתמש בו כדי להעריך כמותית מחויבות גורל סימטרית וא-סימטרית, את האיזון בין התחדשות עצמית להתמיינות, ואת מספר החלוקות עבור גורל מחויבות נתון. בסך הכל, פרוטוקול זה יכול לשמש בתכנונים ניסיוניים שמטרתם לפענח את ההבדלים הביולוגיים בין אבות המטופויטיים, מנקודת מבט של תא יחיד.
העשור האחרון עמד בסימן התפשטות עולמית של גישות חד-תאיות לביולוגיה תאית ומולקולרית. בעקבות השלבים של גנומיקה של תא בודד1,2, כיום ניתן לחקור מרכיבים רבים של תא בודד (למשל, DNA, RNA, חלבונים), עם טכניקות חדשניות של תא בודד -omics המתפתחות מדי שנה. טכניקות אלה שפכו אור על שאלות ישנות וחדשות בתחומי האימונולוגיה, נוירוביולוגיה, אונקולוגיה ועוד, הן באמצעות תאי אורגניזם אנושיים והן באמצעות תאי אורגניזם מודל3. על ידי הדגשת ההבדלים בין תאים בודדים, תא בודד -omics הניע את ההגדרה של מודל חדש של hematopoiesis, שבמרכזו ההטרוגניות של תאי גזע ותאי אב hematopoietic (HSPCs) והתרחקות מהמודל הקלאסי של אוכלוסיות הומוגניות בדידות 4,5.
אחד החסרונות המעטים של כל טכניקות -omics הוא הרס התא של עניין, מניעת האפשרות להעריך את הפונקציונליות שלה. לעומת זאת, שיטות חד-תאיות אחרות, כגון בדיקת השתלה של תא בודד וטכנולוגיות למעקב אחר שושלת, מספקות קריאה של הפונקציונליות של תא האב הקדמון על ידי הערכת גורלם של תאים בודדים in vivo 6,7. טכנולוגיות מעקב אחר שושלת כוללות תיוג התא המעניין עם גנטיקה תורשתית7 או תווית פלואורסצנטית8,9, מה שמאפשר לעקוב אחר גורלם של תאים בודדים מרובים בו זמנית. עם זאת, אפיון התאים ההתחלתיים מוגבל בדרך כלל למספר מוגבל של פרמטרים, כגון ביטוי של כמה חלבוני שטח המוערכים על ידי ציטומטריית זרימה10. בנוסף, טכנולוגיות מעקב אחר שושלת חד-תאית דורשות זיהוי מייגע של התווית התאית, בדרך כלל באמצעות ריצוף DNA/RNA או הדמיה. נקודה אחרונה זו מגבילה במיוחד את מספר התנאים שניתן לבחון בניסוי יחיד.
סוג נוסף של שיטות המשמשות לחקר הפונקציונליות של תאים בודדים הן מערכות תרבית תאים ex vivo של HSPCs בודדים. קל לבצע, בדיקות תקן זהב אלה כוללות מיון של תאים בודדים לתוך 96-בארות כלי תרבית התא, ולאחר תרבית, אפיון פנוטיפ צאצאי התא, בדרך כלל על ידי ציטומטריה זרימה או ניתוח מורפולוגי. בדיקות אלה שימשו בעיקר כדי לאפיין את ההתמיינות ארוכת הטווח של HSPCs לתאים בוגרים, בדרך כלל לאחר 2-3 שבועות של תרבית11,12. לחלופין, הם שימשו כדי לנסות לשמר ולהרחיב את Ex vivo HSPCs 13,14,15,16,17,18, עם הבטחה לתועלת רפואית להשתלת תאי גזע אנושיים 19. לבסוף, הם שימשו לחקר המחויבות המוקדמת של HSPCs באמצעות תרבית לטווח קצר20, כאשר המספר הנמוך של תאים שנוצרו בתרבית זו הוא הגורם המגביל העיקרי. חסרון אחד של סוגים שונים אלה של מבחני ex vivo הוא שהם משקפים רק באופן חלקי את מורכבות in vivo; ובכל זאת, הם אחת הדרכים הנדירות לחקור התמיינות HSPC אנושית.
פיסת מידע אחת חסרה משיטות קיימות של תא בודד (אומיקה של תא בודד, מעקב אחר שושלת ותרבית ex vivo) היא זיהוי מדויק של חלוקות תאים, פרמטר חיוני שיש לקחת בחשבון כאשר חוקרים דינמיקה של HSPC21. דרך פשוטה להעריך את מספר החלוקות באמצעות ציטומטריית זרימה היא שימוש ב"צבעי חלבון" מסיסים, כמו 5-(ו-6)-carboxyfluorescein diacetate succinimidyl ester (CFSE)22. צבעי חלוקה אלה מתפזרים בתוך הציטופלסמה של התאים המוכתמים, מדוללים בחצי ומועברים לשני תאי הבת בכל חלוקת תא, מה שמאפשר למנות עד 10 חלוקות. שילוב של מספר צבעי חלוקה, ניתן לזרוע מספר אבות בודדים באותה באר, שכן כל צבע בודד מאפשר הפרדה בין הצאצאים השונים. זהו העיקרון מאחורי השימוש בצבעי תאים למעקב אחר שיבוט וחלוקה מרובה שהוצג לראשונה עבור לימפוציטים מורינים23,24.
כאן, אנו מציגים את הפיתוח של בדיקת MultiGen לשימוש עם HSPCs מורין ובני אדם. הוא מאפשר בדיקה של תאים בודדים רבים בו זמנית עבור תכונות ההתמיינות, החלוקה והקרבה שלהם ex vivo. בדיקה זו, בעלת תפוקה גבוהה, קלה לביצוע וזולה זו מאפשרת למדוד את הפנוטיפ התאי, את מספר החלוקות שבוצעו, ואת קרבת התאים ויחסי השבט עם התאים האחרים בבאר, והכל בו זמנית. ניתן להשתמש בו כדי להעריך כמותית מחויבות גורל סימטרית וא-סימטרית, את האיזון בין התחדשות עצמית לבין דיפרנציאציה, ואת מספר החלוקות הדרושות לגורל מחויבות נתון. הפרוטוקול דורש ממיין תאים מופעל פלואורסצנטי (FACS) וציטומטר זרימה עם קורא לוחות, בתוספת הציוד הדרוש לביצוע תרבית תאים. בנוסף לפרוטוקול הטכני לביצוע הבדיקה על HSPCs אנושיים, אנו מספקים גם את מסגרת הניתוח המפורטת, כולל הבדיקות הסטטיסטיות הדרושות להערכת תכונות תאיות הקשורות למושג משפחת תאים25. פרוטוקול זה כבר שימש בהצלחה לתיאור תא HSPC מורין26,27.
הפרוטוקול הבא משתמש בתאי CD34+ מועשרים מגנטית כחומר המוצא28. בדרך זו, ניתן להכתים ולבודד ביעילות את HSPCs האנושיים ממקורות דם שונים (למשל, דם טבורי, מח עצם, דם היקפי). חשוב לא להשליך את שבר CD34, מכיוון שהוא ישמש כחלק מהפרוטוקול להגדרת סוגים שונים של בקרות ניסיוניות. ניתן להגדיל או להקטין את כמויות התאים והנפחים שהוזכרו, בהתאם לזרימת העבודה הניסיונית ולצרכים. באופן דומה, ניתן להתאים את הפרוטוקול לחקר סוגים שונים של אבות, פשוט על ידי שינוי הנוגדנים המשמשים למיון התאים ולשלבי ציטומטריית הזרימה.
עבור הפרוטוקול הבא, דם טבורי ללא זיהוי שימש כמקור HSPC ונאסף בהתאם להנחיות שהוגדרו על ידי ביובנק הדם הטבורי של בית החולים סנט לואי (אישור AC-2016-2759) ועם הצהרת הלסינקי.
הערה: לפני שתתחיל, ודא שכל הריאגנטים והציוד הדרושים לפרוטוקול זה זמינים, כמפורט בטבלת החומרים ומוזכר בפרוטוקול. הכינו את הריאגנטים הרלוונטיים טריים ואל תאחסנו אותם, אלא אם צוין זאת במפורש.
1. צביעת צבע התא
הערה: סעיף זה מתאר צביעה עם ארבעה שילובים של צבעי חלוקת תאים CFSE וצבע סגול (CTV). מעבדים את כל הצינורות בו זמנית, גם אם לא מוסיפים תמיסת צבע לתאים. כל השלבים מבוצעים בתנאים סטריליים כדי לאפשר את שלב תרבית התאים הבא. הזמן הנדרש: כ-100 דקות.
- מעבד את מנת הדם הטבורי על פי פרוטוקול מיון מגנטי29. ודא ששני שברים זמינים: שבר CD34 גדול ושבר CD34+ קטן יותר. סובב את שני הצינורות במשך 5 דקות ב 300 x גרם. שאפו את הסופרנאטנט מבלי להפריע לכדור.
- עבור חלק CD34+ , להשעות אותו ב 1 מ"ל של מדיום הנשר של Dulbecco שונה (DMEM) ללא נסיוב בקר עוברי (FBS). לספור את התאים באמצעות hemocytometer; צפיפות התא לא צריכה להיות גבוהה מ 3 x 106 תאים / מ"ל. אם זה המקרה, התאימו את עוצמת הקול בהתאם. עבור שבר CD34, השהה מחדש ב- DMEM עם FBS, וכוונן את עוצמת הקול למקסימום של 6 x 106 תאים/מ"ל.
- Aliquot 250 μL של חלק CD34+ לתוך ארבעה צינורות פוליפרופילן 15 מ"ל. תייג את הצינורות באופן הבא: CD34+/CF (CFSE_only), CD34+/CV (CFSE_high CTV_low), CD34+/VC (CFSE_low CTV_high) ו-CD34+/VI (CTV_high). Aliquot 250 μL של CD34- שבר לתוך עוד ארבעה צינורות פוליפרופילן 15 מ"ל. תייג את הצינורות באופן הבא: CD34-/CF (CFSE_only), CD34-/CV (CFSE_high CTV_low), CD34-/VC (CFSE_low CTV_high) ו-CD34-/VI (CTV_high). ניתן להשליך את התאים הנותרים מהשבר CD34.
- הכן שני פתרונות CFSE, הנקראים CFSE_high ו- CFSE_low. עבור CFSE_high (10 מיקרומטר), ערבב 1.1 מ"ל של DMEM עם FBS עם 2.2 מיקרוליטר של מלאי CFSE (5 מ"מ). עבור CFSE_low (5 מיקרומטר), ערבב 550 μL של DMEM עם FBS ו- 0.55 μL של מלאי CFSE (5 mM).
- הוסף 250 μL של תמיסת CFSE_high לצינורות CF ו- CV, 250 μL של תמיסת CFSE_low לצינורות VC, ו- 250 μL של DMEM עם FBS לצינור VI. כדי להבטיח שילוב יעיל של תרחיף תאים וצבע תא, הטה את הצינור בכמעט 90 מעלות, והפקיד את תמיסות CFSE על דופן הצינור. לאחר מכן, החזק את הצינור במאונך כדי לערבב את שתי התמיסות, ופיפטה שלוש או ארבע פעמים כדי להבטיח ערבוב מהיר של תמיסות CFSE עם התאים המרחפים מחדש. יש לדגור בטמפרטורה של 37°C למשך 8 דקות בדיוק.
- לאחר הדגירה, להוסיף 5 מ"ל של DMEM + 10% FBS. שמור את הצינורות ב 37 ° C במשך 5 דקות.
- סובב את הצינורות במשך 5 דקות ב 300 x גרם. הסר את supernatant באמצעות שאיפה מבלי להפריע את הכדור, ולשטוף את הגלולה עם 5 מ"ל של פוספט חוצץ מלוחים 1x/ethylenediaminetetraacetic חומצה (PBS 1x / EDTA). סובב שוב במשך 5 דקות ב 300 x גרם. השליכו את הסופרנאטנט מבלי להפריע לכדור, והשהו מחדש את גלולת התא ב-250 מיקרוליטר של 1x PBS /EDTA.
- הכינו שני פתרונות CTV, הנקראים CTV_high ו-CTV_low. עבור CTV_high (10 מיקרומטר), ערבבו 1.1 מ"ל של PBS 1x/EDTA ו-2.2 מיקרוליטר של מלאי CTV (5 מילימול). עבור CTV_low (5 מיקרומטר), ערבב 550 μL של PBS 1x/EDTA עם 0.55 μL של מלאי CTV (5 mM).
- הוסף 250 μL של תמיסת CTV_high לצינורות VC ו- VI, 250 μL של תמיסת CTV_low לצינור CV, ו- 250 μL של 1x PBS / EDTA לצינור CF. השתמש באותה טכניקה כמתואר בשלב 1.5. יש לדגור בטמפרטורה של 37°C למשך 8 דקות בדיוק.
- לאחר הדגירה, להוסיף 5 מ"ל של DMEM + 10% FBS. יש לשמור על טמפרטורה של 37°C למשך 5 דקות.
- סובב את הצינורות במשך 5 דקות ב 300 x גרם, להשליך את supernatant מבלי להפריע את הכדור, ולאחר מכן לשטוף את הגלולה עם 5 מ"ל של 1x PBS / EDTA. סובב שוב במשך 5 דקות ב 300 x גרם.
- השליכו את הסופרנאטנט מבלי להפריע לכדור, והשהו מחדש את שברי CD34 ב-1x PBS /EDTA לריכוז סופי של 1.5 x 106 תאים/מ"ל. השהה מחדש את שברי CD34+ ב 40 μL של חיץ צביעה, ולהעביר את התאים לתוך צינורות 1.5 מ"ל.
2. צביעת נוגדנים
הערה: ניתן להתאים את צביעת הנוגדנים בהתאם לצרכי הניסוי. רק שברי CD34+ עוברים צביעת נוגדנים; שברי CD34 משמשים כבקרת צביעה יחידה עבור שילובי הצבעים של חלוקת התא (שברים CV, VC, CF ו- VI). הפאנל הבא מותאם לזיהוי ארבעה סוגים של HSPCs: תאי גזע המטופויטיים (HSC), אבות רב-פוטנטיים (MPPs), אבות רב-פוטנטיים ראשוניים לימפואידים (LMPPs) ותאי אב המטופויטיים (HPC)12. עם זאת, זיהוי של HSCs ו- MPPs מוצג. הזמן הנדרש: 75 דקות.
- הכינו את הצביעה הבודדת לצביעת פני השטח, בעזרת חרוזי פיצוי. ערבבו חרוזים שליליים וחרוזי אימונוגלובולין G (IgG) ביחס של 1:1, לקבלת נפח כולל שווה ערך ל-20 μL x מספר סמני פני השטח (למשל, 120 μL אם לוח הצביעה מכיל שישה נוגדנים).
- שלח 20 μL של חרוזים לתוך צינורות בודדים 1.5 מ"ל עבור כל סמן. הוסף את הנפח המתאים לגורם הדילול עבור כל נוגדן בצינור המתאים (למשל, אם מקדם הדילול הוא 1:20, הוסף 1 μL).
- כדי להכתים את תאי CD34+ , הכינו תערובת אב של נוגדנים12, המבוססת על טבלה 1. מערבבים את הנוגדנים לתוך צינור יחיד של 0.5 מ"ל. הוסף 7 μL מתערובת הנוגדנים הראשית לכל אחד מארבעת תנאי CD34+.
- דגרו על חרוזי הפיצוי ועל דגימות CD34+ בטמפרטורה של 4 מעלות צלזיוס למשך 30 דקות לפחות.
הערה: יש להתאים את זמן הדגירה לפרטים הטכניים של הנוגדנים המשמשים להכתמה. - במהלך הדגירה, הכינו את הצלחת בעלת 96 הקידוחים בעלי תחתית עגולה שתשמש למיון, והוסיפו 100 מיקרוליטר של מדיום תרבית תאים לכל באר באמצעות פיפטה רב ערוצית.
הערה: השאר בארות H8-H12 ריקות. - תווית 5 מ"ל צינורות פוליפרופילן עבור פקדי צביעת פני השטח (5, באמצעות חרוזים), פקדי חלוקת התא (4, באמצעות שברים CD34 ), ואת דגימות CD34+ (4).
- בסוף הדגירה, לשטוף את התאים ואת החרוזים עם 1 מ"ל של חיץ מכתים. העבר את הנפח הכולל לצינורות פוליפרופילן 5 מ"ל. צנטריפוגו את הצינורות במשך 5 דקות ב 300 x גרם, ואז לשאוף את supernatant מבלי להפריע את הכדור.
- השהה מחדש את התאים במאגר הצביעה, באמצעות כ- 500 μL כל אחד עבור החרוזים ותאי CD34+ , ו- 1 מ"ל עבור צינורות CD34.
טבלה 1: תבנית להכנת תערובת האב של הנוגדנים לניסוי מיון תאים. אנא לחץ כאן כדי להוריד טבלה זו.
3. מיון תאים
הערה: מספרי התאים הממוינים עשויים להשתנות בהתאם לכמות התאים הכוללת הזמינה. בפרוטוקול, מסופק מספר תא מינימלי עבור כל פקד. הזמן הדרוש (לצלחת בודדת): 100 דקות.
- פתח את ניסוי התבנית או הגדר ניסוי חדש. צור דגימה אחת וצינורות מרובים, אחד לכל תנאי.
- הגדר את אסטרטגיית ה- gating המפורטת באיור 1, וצור דיאגרמות תרשים של שש נקודות. ראשית, דמיינו את התאים בתרשים נקודות FSC-A/SSC-A, ולחצו פעמיים על הכלי INTERGON GATING כדי לבחור אוכלוסייה עם פיזור צדדי נמוך (איור 1A). בתרשים הנקודות הבא (FSC-A / FSC-H), לחץ לחיצה ימנית על העלילה ובחר את השער "תאים" מהתפריט הנפתח על ידי לחיצה עליו. השתמשו באותו כלי גאטינג כדי לבחור אוכלוסייה הדוקה באלכסון שבין שני הצירים (איור 1B).
- בתרשים הנקודות השלישי (APC לעומת FSH-H), הציגו את האוכלוסייה "תאים בודדים" ושער התאים שלילי לביטוי של שושלת APC (Lin) (איור 1C). בחלקה הרביעית (CFSE לעומת CTV), הציגו את האוכלוסייה "לין" וצרו ארבעה שערים נפרדים, אחד לכל שילוב צבעים (איור 1D).
הערה: שערים אלה חייבים להיות הדוקים, כדי לבחור רק חלק קטן של תאים מוכתמים הומוגניים. - השתמש בתרשים החמישי והשישי (APC-Cy7 לעומת BV650 ו- PE-Cy7 לעומת PE) כדי לזהות את אבות העניין. שער נדיב את אוכלוסיית CD34+CD38 ואת אוכלוסיית CD34+CD38+ בחלקה החמישית (איור 1E). לאחר מכן, בחר את אוכלוסיית CD34+CD38- בחלקה השישית, וצייר שלושה שערים, לפי איור 1F.
- הפעל את צינורות הצביעה הבודדים המכילים את חרוזי הפיצוי, ולחץ על כפתור הרכישה . התאם את מתחי צינור מכפיל האור (PMT) מהתפריט הנפתח פרמטרים , במיוחד עבור צבעי חלוקת התא (בין 104 ל- 105 בקנה מידה דו-מעריכי).
- לטש את מטריצת הפיצוי בהתאם ללוח המשמש למיון, באמצעות הכרטיסייה פיצוי . הקלט לפחות 5,000 אירועים בשער החרוזים, ולחץ על כפתור ההקלטה .
- הפעל את שברי CD34 ובדוק שוב את מטריצת הפיצוי. תעד לפחות 10,000 אירועים בשער החד-תא.
- הפעל את שברי CD34+ , והקלט לפחות 5,000 אירועים בשער התא היחיד. התאימו את השער לכל שילוב צבעים, והגדירו שער הדוק לבחירת אוכלוסייה הומוגנית (איור 1D). באופן דומה, התאם את ה- gating לבחירת HSCs ו- MPPs.
- לאחר השלמת הניתוח וכל הצינורות נרשמים, הכנס את הצלחת במחזיק המתאים, לאחר ביצוע כיול אריה סטנדרטי למיון על לוחות 96 בארות. מומלץ לקרר את הצלחת.
- הכן את תבנית מיון הלוחות בהתאם לסכמה המוצגת בטבלה 2, באמצעות פריסת מיון הניסוי. הבארות הנקראות "CD34-" מכילות 5,000-10,000 תאים, ממוינים בשער CF/CV/VC/VI. הבארות "בתפזורת" מכילות לפחות 500 תאים, ממוינים על השער CD34+CD38-. לבסוף, הבארות החד-תאיות מכילות רק אירוע אחד לכל שילוב צבע חלוקת תאים לכל באר, כך שבסך הכל ארבעה אירועים לכל באר.
הערה: ניתן להתאים את אוכלוסיות ה"תפזורת" לתת-קבוצה ספציפית של אבות; אל תמיין פחות מ- 500 תאים. - עבור המיון, להמשיך לפי הסדר, להשלים כל שילוב צבע חלוקת התא לפני המעבר הבא. לדוגמה, התחל במיון CD34- CF, במצב טוהר תפוקה . לחץ על כפתור הרכישה ולאחר מכן על כפתור המיון.
- בסוף מיון CD34, הכנס את צינור CF CD34+ . לרכוש, ולאחר מכן ללחוץ על כפתור המיון, לוודא שיש לסמן 0/ 16/0 כמו ציון טוהר. לבסוף, מיין את התאים המעניינים, תא אחד לכל באר, בטוהר של תא יחיד , והקפד לסמן את אפשרות מיון האינדקס .
- עבור לשילוב הצבעים הבא של חלוקת התא, וחזור על אותו סדר. כהפניה, טבלה 2 מספקת דוגמה ללוח ממוין.
הערה: פונקציית מיון האינדקס יוצרת קבצים נפרדים עבור כל תנאי ממוין. - בסוף המיון, יצא את הקבצים כקבצי .fcs 3.0. שים את התאים 37 ° C, 5% CO2 אינקובטור. התאים מתורבתים במשך מספר ימים, על פי תכנון הניסוי, במשך 24 שעות26 לפחות.
טבלה 2: תבנית עבור לוח תא הממיין 96 בארות, בהתבסס על הדרישות הספציפיות לניתוח ציטומטריית זרימה עוקבת. אנא לחץ כאן כדי להוריד טבלה זו.
4. ניתוח נתוני מיון תאים
הערה: כדי לאמת את איכות מיון התאים, ניתוח נתוני FACS נחוץ לפני שמתקדמים הלאה. הפלט העיקרי של שלב זה הוא יצירת גיליון אלקטרוני המכיל את עוצמות הסמן של כל תא בודד ממוין.
- העלה את קבצי .fcs 3.0 בתוכנת הניתוח.
- אמת את הגדרת הפיצוי המשמשת במהלך מיון תאים, באמצעות קבצי הצביעה הבודדים שהוקלטו לפני המיון בפועל.
- הגדר את אסטרטגיית ה- gating המתוקנת באמצעות הקבצים המתאימים לצובר השונה. העתק והדבק שערים אלה בקבצי מיון האינדקס.
- ודא שהתאים הממוינים באינדקס נפלו בשער שנקבע. אם ישנם כמה תאים ממוינים שגודרו באופן שגוי, ניתן לזהות אותם על-ידי ייצוא קואורדינטות הלוחות שנרשמו במהלך מיון האינדקס, ולהסיר אותם מאוחר יותר בניתוח.
- יצא את האירוע מקבצי מיון האינדקס כפרמטרים מפוזרים. לייצא אותם כמו קבצי .csv, מתקתק את האפשרויות "קנה מידה ערכים" ו "פרמטרים פיצוי". יש לייצא קבצים אלה בתיקייה בשם "קבצים מיוצאים".
- שלב את כל הקבצים בקובץ .csv אחד, באמצעות הסקריפט בקובץ משלים 1. הגדר את הנתיב הנכון עם הפונקציה "setwd". הפלט של סקריפט זה הוא גיליון אלקטרוני המכיל את כל האירועים המגודרים השונים ואת העוצמות היחסיות עבור כל הפרמטרים.
5. לאחר הכתמת נוגדנים בתרבית
הערה: בצע חלק זה של הפרוטוקול בתנאים סטריליים; מספר ריאגנטים משותפים עם השלבים הראשונים, וצריכים להישאר סטריליים. לניתוח ציטומטריית זרימה, השתמש בציטומטר זרימה עם קורא לוחות. זה מאפשר לבצע את הצביעה ישירות בצלחת תרבית הרקמה, הפחתת אובדן התא למינימום על ידי הגבלת כמות הצנרת והספינינג. הכינו את צביעת סמן המשטח בצבע יחיד באמצעות חרוזי הפיצוי, למעט בארות A1-A4, המייצגות את הצביעה היחידה לצבעי CF/CV/VC/VI וכבר קיימות בלוח 96 הקידוחים. אוכלוסיות הצובר הממוינות על פי צבע התא עוזרות לקבוע את אסטרטגיית הגאטינג למספר החטיבות ולגידור הכללי. זמן נדרש: 120 דקות.
- לפני תחילת הפרוטוקול, סמן את הבארות המכילות לפחות תא אחד על ידי בדיקת הצלחת תחת מיקרוסקופ הפוך. שלב זה מאפשר לייעל את כמות הנוגדנים המשמשים להכתמה ומאיץ את ההליך.
- הכינו את תערובת הנוגדנים, לפי טבלה 3. מכיוון שיש כמות משמעותית של צנרת, הטבלה לוקחת בחשבון את התקלה הטכנית עקב הפיפט, כולל תוספת של 5% נפח. הנוגדנים המתוארים בטבלה מאפשרים לאפיין מגוון של HSPCs מדגימות דם טבורי אנושיות12.
- צנטריפוגה את הצלחת במשך 5 דקות ב 300 x גרם. הפוך במהירות את הצלחת מתחת למכסה המנוע ומעל מגבת נייר, כדי להסיר את הסופרנטנט.
- הוסף 8 μL של חיץ צביעה לבארות A1-A4. מוסיפים 8 μL מהתערובת לבארות האחרות.
- ערבבו את החרוזים השליליים ואת חרוזי פיצוי IgG ביחס של 1:1, לקבלת נפח כולל שווה ערך ל-120 μL. יש לשלוח 20 μL בצינור אחד של 1.5 מ"ל לכל סמן. הוסף את נפח הנוגדנים המתאים לגורם הדילול (למשל, אם מקדם הדילול הוא 1:20, הוסף 1 μL).
הערה: התאם את הנפח הכולל למספר הסמנים המשמשים לצביעה (לדוגמה, 100 μL אם לוח הצביעה מכיל חמישה נוגדנים). - יש לדגור על הצלחת ועל בקרת פיצוי הצביעה היחידה בטמפרטורה של +4°C, למשך 30 דקות לפחות.
הערה: יש להתאים את זמן הדגירה לפרטים הטכניים של הנוגדנים המשמשים להכתמה. - לשטוף את החרוזים עם 1 מ"ל של חיץ מכתים. העבר את הנפח הכולל לצינורות פוליפרופילן 5 מ"ל שסומנו בעבר. צנטריפוגה את הצינורות במשך 5 דקות ב 300 x גרם, ולאחר מכן להסיר את supernatant באמצעות שאיפה.
- שטפו את התאים בצלחת על ידי הוספת 100 מיקרוליטר של חיץ צביעה לכל באר באמצעות פיפטה רב ערוצית. צנטריפוגו את הצלחת ב 300 x גרם במשך 5 דקות, ולאחר מכן במהירות להפוך את הצלחת מתחת למכסה המנוע מעל מגבת נייר, כדי להסיר את supernatant.
- להשהות מחדש את התאים ב 85 μL של חיץ צביעה, באמצעות פיפטה רב ערוצית.
- התחל את הניתוח על ציטומטר הזרימה (מצב רכישה), באמצעות התבנית הייעודית ולחץ על מותאם אישית. תבנית מותאמת אישית זו לוקחת בחשבון את התכונות הטכניות של לוח 96 בארות עגול תחתון, במיוחד את המידות של כל באר (קוטר, עומק ועובי). הגשושית חייבת להגיע לתחתית הבאר, לכן יש למקם אותה בדיוק במרכז הבארות A1 ו-H12.
- לאחר בחירת הפלואורופורים המעניינים מהרשימה המוצעת על ידי התוכנה, הגדר את הגדרת הלוח לפי תבנית הלוחות של טבלה 2, מתוקנת למספר הבארות המכילות תא אחד לפחות.
- בחר 100 μL כמגבלת נפח הרכישה. סמן את אפשרות התסיסה . הגדר את קצב הרכישה למקסימום של 1 μL/s, מכיוון שמהירות נמוכה יותר משפרת את הנפח הכולל המנותח לכל באר.
- הוסף את פתרונות הניקוי והשטיפה המתאימים לבארות H8-H12. התבנית בטבלה 2 משאירה באופן ספציפי בארות H8-H12 ריקות, מכיוון שציטומטר הזרימה צריך להפעיל מגוון תנאי שטיפה בסוף הניתוח.
הערה: שלב זה מותאם לפרטים של ציטומטר הזרימה שבו נעשה שימוש. - במקטע חלקות ושערים, הגדר תחילה את השער החד-תאי, באמצעות FSC-A/SSC-A scatterplot ולאחר מכן את FSC-H/FSC-A scatterplot. צור היסטוגרמה לכל סמן עניין.
- לאחר אישור ההגדרות, המשך לסעיף ניתוח . נתחו תחילה את הצביעה הבודדת, ותיעדו לא פחות מ-5,000 אירועים (טווח אופטימלי: 5,000-15,000 אירועים), הן עבור חרוזי הפיצוי והן עבור השברים המוכתמים ב-CD34. כוונן את המתחים במידת הצורך.
- לאחר שכל הכתמים הבודדים נרשמים, ניתן להתחיל את הרכישה בפועל, תוך לחיצה על פונקציית הרכישה .
טבלה 3: תערובת מופת של נוגדנים לניסוי ציטומטריית זרימה, במיוחד לזיהוי HSPCs מדם טבורי אנושי. אנא לחץ כאן כדי להוריד טבלה זו.
6. ניתוח נתוני ציטומטריה של זרימה לאחר תרבית
הערה: ניתוח הנתונים המתואר הוא ספציפי עבור התוכנה המוזכרת בטבלת החומרים. הפלט העיקרי הוא יצירת גיליון אלקטרוני המכיל מידע על עוצמת סמן פני השטח, מספר החלוקות וקרבה לכל תא מנותח. חלק זה של הפרוטוקול כלול סקריפט שנכתב ב-R, הדרוש לזרימת עבודה זו כדי ליצור את גיליון הניתוח הסופי.
- יצא קבצים מציטומטר הזרימה כקובצי .fcs. העלה אותם לתוכנת הניתוח, וקבץ אותם יחד כ"צביעה יחידה", "בתפזורת" ו"תא בודד".
- הכינו מטריצת פיצוי באמצעות קובצי הצביעה הבודדים, והחילו אותה על שתי הקבוצות האחרות בגרירה ושחרור.
הערה: אם נעשה שימוש בכלי פיצוי אוטומטי, בדוק את האיכות ביד לפני שתמשיך הלאה. - כדי שיהיה נציג של גטינג, שרשרו את הבארות השונות בתפזורת בקובץ אחד. שלב זה מדגיש במהירות אם שני צבעים חופפים (בדרך כלל CV ו- VC) או חריגות אחרות, ולכן יש להחריג אותם. לאחר לחיצה על אפשרות אוכלוסיות השרשור , בחר את כל הפרמטרים ללא פיצוי מתפריט "פרמטרים" ולאחר מכן לחץ על שרשור.
- העלה את הקובץ המשורשר לסביבת העבודה, ולאחר מכן החל את מטריצת הפיצוי באמצעות גרירה ושחרור.
- הכינו את אסטרטגיית ה-gating שהוגדרה באיור 2 באמצעות הקובץ המשורשר. בשער התא היחיד, הצג את האירועים בתרשים פיזור עם CFSE ו- CTV. צרו שער ראשון שנקרא 'מסומן', כולל כל ארבעת הצבעים ולא כולל פלואורסצנטיות אוטומטית אפשרית (איור 2C). לאחר מכן, שער כל צבע בנפרד.
- תאים המסומנים עם CV ו- VC זקוקים לערך מותמר, בהתחשב בכך שהצבע הוא תוצאה של אותות CFSE ו- CTV. שני האותות המתואמים מסובבים אפוא בסקאלה לוגריתמית ב-45°, כדי לאפשר לדילול החלוקה להתקדם במקביל לציר ה-x. ערך זה שעבר המרה נגזר באופן ידני, לחיצה על כלים ולאחר מכן על פרמטר נגזרת. הדבק את הנוסחה הבאה בתיבת הנוסחאות :

הערה: המשוואה26 מניחה כי CFSE ו- CTV הם פרמטרים 03 ו- 17. - כדי להמחיש נכונה את הפרמטר החדש הזה בשם פרמטר נגזר, הגדר ציר ליניארי בטווח ~3-7, לחץ על אפשרות פרמטר ציר ובחר התאמה אישית של ציר.
- החילו את הגאטינג על כל צבע בנפרד כתרשים היסטוגרמה: עבור CF ו- VI, קבעו את CFSE-A ו- CTV-A על ציר x, בהתאמה. עבור CV ו- VC, הגדר את הפרמטר הנגזר החדש על ציר x. הגדר שערים המתאימים לכל פסגה, כפי שמוצג באיור 3.
- החל את gating על כל תא בודד בודד היטב. הקפד להוסיף את הפרמטר הנגזר לכל באר מנותחת. אמת ידנית כל שער צבע עבור כל באר, כדי לזהות אירועים שהוקצו באופן שגוי לשיא נתון. דוגמאות ל-gating מוצגות באיור 4.
- לאחר השלמת הניתוח, וכל הבארות אומתו, בחר את כל שערי CF/CV/VC/VI המכילים תא אחד לפחות. לייצא אותם כמו קבצים .csv, מתקתק את האפשרויות "קנה מידה ערכים" ו "פרמטרים פיצוי". קבצים אלה מיוצאים לתיקייה בשם "קבצים מיוצאים".
- שלב את כל הקבצים .csv אחת, באמצעות סקריפט R בקובץ משלים 1. זכור להגדיר את הנתיב הנכון עם הפונקציה "setwd". הפלט של סקריפט זה הוא גיליון אלקטרוני המכיל את כל האירועים המגודרים השונים ואת העוצמות היחסיות עבור כל הפרמטרים.
- פתח את הגיליון האלקטרוני ושנה את שמות העמודות עבור כל פרמטר, לדוגמה, באמצעות השמות הבאים: CFSE, CTV, CD90, CD123, CD45RA, CD34, CD38. שמות אלה ישמשו לזיהוי סף ה- gating כדי להקצות נכונה לכל תא את זהותו.
- הוסף שש עמודות בשם "ובכן", "מצב", "צבע", "דור", "Original_cell" ו- "Culture_time". משתנים אלה הם אלה שהוגדרו בניסוי ומוסקים מכל שורה:
export_A10 CD34 + PBS_CV_Peak 1.csv.1 = A10 (טוב), CD34+ (Original_cell), PBS (מצב), CV (צבע), Peak_1 (דור). - ייצא את הבארות בתפזורת כדי לזהות את ערכי הסף לגידור: ייצא את אוכלוסיית העניין המפוצה (למשל, CD34+CD38-) כקבצי .csv, תוך סימון האפשרויות "ערכי קנה מידה" ו"פרמטרים מפוצים". יצא קבצים אלה בתיקייה בשם "קבצים מיוצאים".
- כדי למצוא את הסף עבור CD38, זהה את הערך המספרי הגדול ביותר עבור פרמטר זה. לעומת זאת, כדי למצוא את הסף עבור CD34, זהה את הערך המספרי הקטן ביותר עבור פרמטר זה. חזור על תהליך זה עבור כל הפרמטרים המעניינים.
הערה: עבור הניתוח המוצג בפרוטוקול, הסמן CD45RA משמש הן לזיהוי LMPPs בשער CD34+CD38- והן CMP/GMP בשער CD34+CD38+. משמעות הדבר היא שיש לחלץ שני ערכי סף שונים עבור סמן זה. - העתק והדבק את ערכי הסף בקובץ Excel בשם "gating_matrix". קובץ זה מאורגן לפי טבלה 4, ומאפשר ניתוח של ניסויים עצמאיים מרובים. חשוב מאוד לתת שם לכל עמודה בדיוק עם סכימה זו: XXYYMMDD_xxh, כאשר XX מייצג את שני ראשי התיבות של המפעיל, YY את שני המספרים האחרונים עבור השנה, MM עבור החודש, DD עבור היום, ו- xx עבור נקודת הזמן של הניתוח.
טבלה 4: מטריצת גטינג להקצאת גורל התא, לפני הניתוח הסטטיסטי. CD45h מתייחס לעוצמה של CD45RA עבור תת-קבוצה של HPC, בעוד CD45l מתייחס לעוצמה של CD45RA עבור תת-קבוצות CD34+CD38. אנא לחץ כאן כדי להוריד טבלה זו.
7. ניתוח סטטיסטי
הערה: הבדיקה הסטטיסטית של הנתונים המופקים כוללת צינור ניתוח מותאם אישית, המקודד באמצעות שפת התכנות Python (קובץ משלים 2, קובץ משלים 3 וקובץ משלים 4). הסקריפט מאורגן בשלושה בלוקים: הראשון לעיבוד הגיליון האלקטרוני, הבלוק השני ליצירת מפת החום להדמיית נתונים, והבלוק האחרון ליצירת היסטוגרמות מרובות לניתוח ובדיקה של מאפייני בידול וחלוקה.
- החל מהבלוק "0_process_data" (קובץ משלים 2), ודא שנתיבי gating_matrix והגיליון האלקטרוני של הנתונים מוגדרים כראוי בסקריפט.
- הגדר את מילון "cell_cols", כדי להקצות את גורלות התאים הרלוונטיים לכל תא. במקרה הספציפי, הגורלות הם HSCs, MPPs, LMPPs, אבות מיאלואידים נפוצים (CMPs), אבות גרנולו-מונוציטיים (GMPs), אבות מגה-קריוציטים-אריתרואידים (MEPs) ו- CD34-.
- באמצעות ערכי הסף שהוגדרו מבארות הצובר (שלב 6.16), הגדר את הפונקציה "cell_class_exp_time". חיוני להיות עקבי במתן שמות לעמודות, להגדיר נכון ערכי סף אלה, תוך שימוש באותו שם המשמש להגדרת כל עמודה בשלב 6.12.
- הפנוטיפים של התא מוגדרים בסקריפט באמצעות סדרה של משפטי "אם-אחר", בהתבסס על ערכי הסף שזוהו במהלך ניתוח ציטומטריית הזרימה.
הערה: ניתן להציג פנוטיפים שונים על-ידי שינוי הצהרות אלה כדי להתאים לשילובי סמנים אחרים. - ציין את התנאים הספציפיים לניסוי, באמצעות הפונקציה "cond_rule" (למשל, טיפולים ניסיוניים שונים). עבור ערכת הנתונים שסופקה, התנאים נקראים "GT" ו- "Diff". תאר את שתי תרביות התאים השונות המשמשות לתרבית התאים. מידע זה ישמש את הבלוק "1_dot_plot" (קובץ משלים 3) לשרטוט מפת החום.
- בבלוק "2_bar_plot" (קובץ משלים 4), הגדר את המילון "class_dct", כולל גורלות התאים הנפרדים המעניינים. עבור מערך הנתונים שסופק, גורלות התאים המעניינים זהים למילון "cell_cols".
- הגדר "conds" (תנאים), "or_cells" (התא המקורי), "sym_labs" (תוויות סימטריה) ו- "times" (נקודת הזמן של הניסוי). אלה מסננים חוזרים ונשנים הנחוצים לתכנון. "conds" לוקחים שוב את התנאים המוגדרים ב-"cond_rule", "or_cells" הם HSCs ו-MPPs, ו-"sym_labs" מתארים את סוג החטיבות.
- בבלוק "2_bar_plot", ניתן להתוות תאים שהתקדמו עד חלוקה 6.
הערה: ערכת הנתונים שסופקה כוללת רק תאים עד חטיבה 4, כך שהודעת שגיאה מוקפצת, אך פעולה זו אינה מונעת מקובץ ה- Script לפעול. - ניתן לאחזר את האיורים שנוצרו על ידי הסקריפט בתיקייה בשם "איורים" כקובצי PDF. הקבצים בשם "Test" מייצגים את הבדיקות הסטטיסטיות השונות שבוצעו עבור ההיסטוגרמה המתאימה.
מיון FACS
אסטרטגיות המיון המוצגות בפרוטוקול זה מבוססות על אסטרטגיות מקובלות 12,30,31. עבור אסטרטגיית הגאטינג המוצגת באיור 1, חומר המוצא הוא אבות דם טבורי שטוהרו בעבר באמצעות העשרה מגנטית CD34+, מה שמסביר את האחוז הזניח של תאים חיוביים לשושלת. חיוני להשתמש בשערים הדוקים עבור ארבעת שילובי הצבעים התוך-תאיים (למשל, ה-CTV באיור), כדי לשפר את רזולוציית הפסגות במהלך הניתוח הבא ולשער את אוכלוסיית התאים הנכונה (איור 1D). במקרה המוצג באיור, השערים בוחרים עבור האוכלוסייה הגדולה והמוגדרת יותר. הימצאותן של אוכלוסיות מרובות וקרובות לכל שילוב צבעים של חלוקת תאים אינה מייצגת, מניסיוננו, הבדלים ביולוגיים. במקום זאת, זה יכול להצביע על א) הליך צביעה לא אופטימלי, או ב) הטרוגניות גדולה (במיוחד בגודל) במאגר ההתחלתי של תאים. זה לא בלתי צפוי כאשר מתחילים מדם טבורי או ממקורות ביולוגיים מורכבים אחרים (למשל, מח עצם, דם היקפי). אם השער אינו מוגדר היטב, הדילול ההדרגתי של שילובי הצבעים השונים יכול להוביל למיזוג של הפסגות המאוחרות יותר, במיוחד עבור התנאים CV ו-VC (איור 2D). תוצאה שלילית נוספת של גטינג תת-אופטימלי היא חוסר היכולת להבחין ביעילות בין פסגות שונות לאחר תרבית תאים, שכן אוכלוסייה התחלתית הטרוגנית יכולה להוביל לפסגות רדודות.
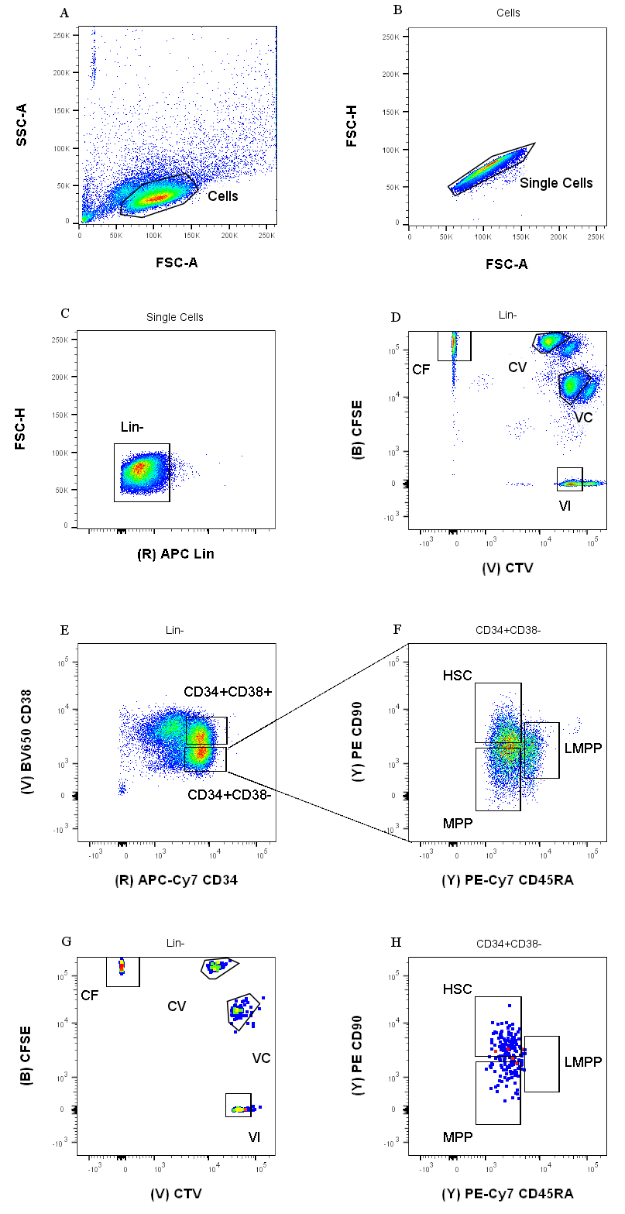
איור 1: אסטרטגיית Gating למיון תאים. (A) FSC-A לעומת SSC-A, כדי למנוע פסולת ותאים מזהמים. (B) FSC-A לעומת FSC-H, כדי לא לכלול כפילויות וגושי תאים. (C) Lin לעומת FSC-H, כדי לא לכלול תאים שהם Lin+. (D) CTV לעומת CFSE, כדי לזהות באופן חד-משמעי את התאים המוכתמים בצירופי הצבעים CF, CV, VC ו-VI. השערים צריכים להיות קפדניים מספיק כדי לכלול אוכלוסייה הומוגנית. (E) CD34 לעומת CD38, כדי להפריד את האבות המוגבלים CD34+CD38+ (הנקראים גם HPC) מהתא הרב-פוטנטי CD34+CD38-. (F) CD45RA לעומת CD90, מאוכלוסיית CD34+CD38, כדי להפריד בין האבות הלא בשלים ביותר המועשרים ב-HSC (CD90+CD45RA-), LMPP (CD90באמצעCD45RA+), וה-MPP המחויב יותר (CD90-CD45RA-). (G) אינדקס ממוין אירועים, המיוצגים כאן עבור צביעת שילוב צבע התא שלהם ו- (H) ביטוי סמני פני השטח CD90 ו- CD45RA. אנא לחץ כאן כדי להציג גרסה גדולה יותר של איור זה.
ניתוח ציטומטריית זרימה לאחר תרבית תאים
הנתונים באיור 2 מייצגים HSCs של דם טבורי אנושי, שנשמרו בתרבית במשך 72 שעות, בנוכחות ציטוקינים מרובים המסוגלים לתמוך במגוון של אבות ומבשרים מיאלואידים. לוחות 2A עד 2D מייצגים את ה-gating הדרוש כדי לבסס את הקרבה של כל אחד מהתאים הבודדים, בעוד שלוחות 2E עד 2G מאפשרים פנוטיפ תאי. הנוכחות המופחתת של חברי פרלמנט באיור היא ככל הנראה תוצאה של תנאי התרבית ששימשו לניסוי המייצג הזה (איור 2F). שימוש בציטוקינים שונים ובתנאי תרבית שונים משנים את האחוז היחסי של כל תת-קבוצה, בדומה לבחירת תאי התחלה שונים לניסוי.
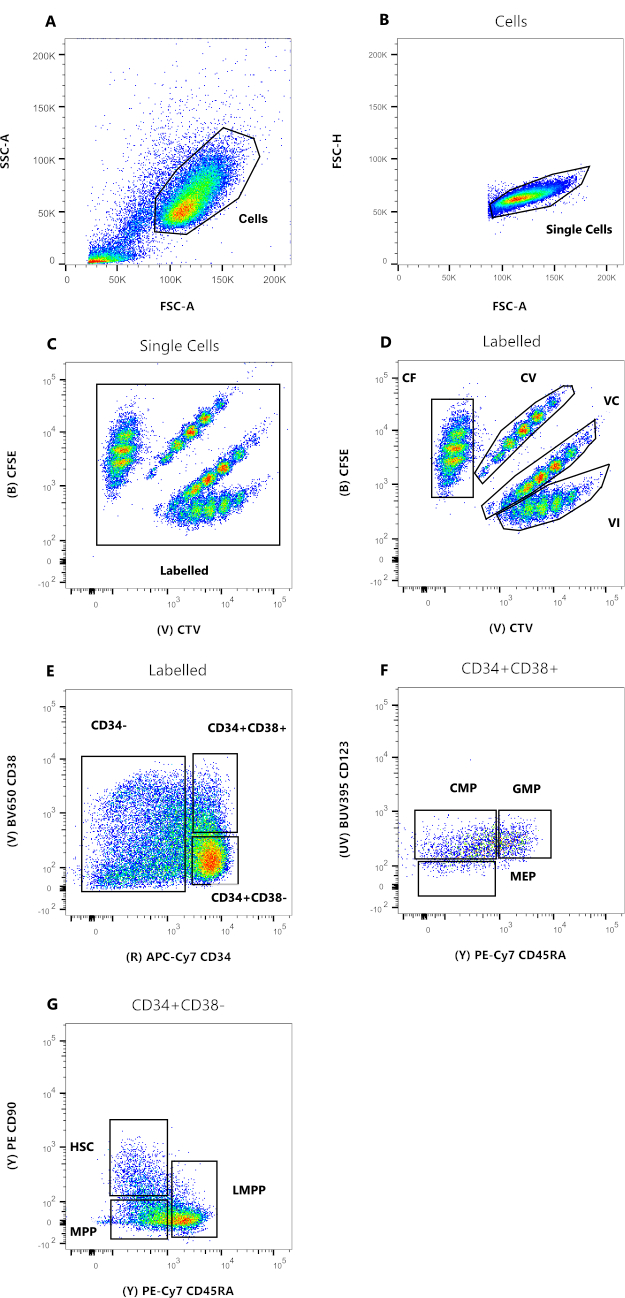
איור 2: אסטרטגיית Gating לניתוח ציטומטריית זרימה. (A) FSC-A לעומת SSC-A, כדי להוציא פסולת ותאים מזהמים. (B) FSC-A לעומת FSC-H, כדי לא לכלול כפילויות וגושי תאים. (C) CTV לעומת CFSE, השער המסומן מאפשר לא לכלול כל אירוע פלואורסצנטי אוטומטי שעלול להשפיע על רזולוציית הנתונים. (D) CTV לעומת CFSE. חשוב מאוד לשער בקפדנות את ארבע האוכלוסיות, בהתבסס על דילולי הצבע של חלוקת התא. (E) CD34 לעומת CD38, כדי להבחין בין מבשרי מחויבות (CD34-), אבות מוגבלים (HPC) (CD34+CD38+), ואבות לא בשלים (CD34+CD38-). (F) CD45RA לעומת CD123, כדי להבחין בין שלושה סוגים של אבות מוגבלים: CMP (CD123+CD45RA-), MEP (CD123-CD45RA-), ו-GMP (CD123+CD45RA+). (G) CD45RA לעומת CD90, מ- CD34+CD38-, כדי לזהות HSCs, LMPPs ו- MPPs. לחץ כאן כדי להציג גרסה גדולה יותר של איור זה.
שלבי הגדרת השיא וההקצאה (איור 3 ואיור 4) הם היבטים מכריעים של הפרוטוקול ומחייבים הגדרה של שערים קפדניים. עבור הגדרת השיא (איור 3), דרושים לפחות 1,000 אירועים לזיהוי אמין. במובן זה, זה יכול להיות מועיל לבודד תאים נוספים במהלך שלב מיון התא עבור בארות "בתפזורת". איור 4 מתאר ארבע דוגמאות של בארות בודדות המכילות משפחות מרובות. איור זה מבהיר את החשיבות של איור 2D ואיור 3, במיוחד לזיהוי של כל משפחה וכל פסגה. איור 4A מדגים דוגמה פשוטה, מאחר שכל התאים בשער CF קרובים מאוד זה לזה וניתן לשייך אותם בקלות לפסגה אחת. איור 4C מראה דוגמה נוספת למשפחה המחולקת באופן חד-משמעי על שתי פסגות מופרדות היטב, כפי שהיא מוצגת בבירור בהיסטוגרמה של איור 4D. איור 4E,G חושף את החשיבות של הקפדה על מספר רב של אירועים; שניהם מציגים מעט אירועים קרובים, אך מחוץ לשערי שילוב הצבעים. אירועים אלה יכולים להיכלל באופן שגוי בשערי VI ו- CF, בהתבסס אך ורק על ניתוח באר יחידה. לבסוף, איור 4F,H מציג שתי דוגמאות שונות של משפחות המפוזרות על פסגות מרובות, עם דוגמה אחת של שתי פסגות עוצמה דומות (איור 4F) ואחת עם שתי פסגות עוצמה לא שוות (איור 4H).
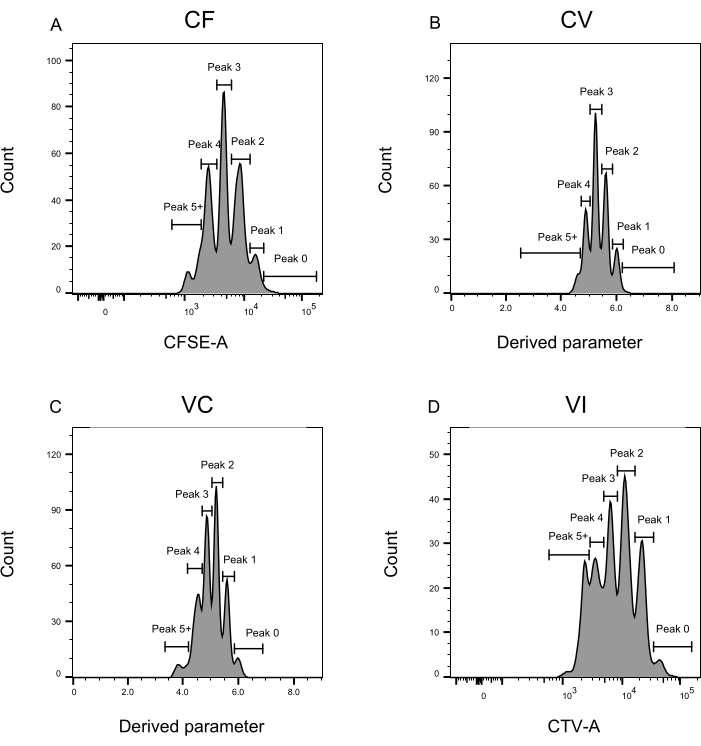
איור 3: הגדרת שיא לניתוח ציטומטריית זרימה. (א-ד) יש להגדיר פסגות הרושמות לפחות 500 אירועים, כדי להבטיח ייצוג טוב לכל שיא בודד. (A) היסטוגרמה לעוצמת CFSE-A. ניתן לזהות מספר פסגות, שכל אחת מהן מתאימה לאוכלוסייה שונה של תאים מתחלקים. (ב,ג) היסטוגרמות לעוצמת הפרמטר הנגזר, המייצגות את תערובת CFSE-CTV, CV (B) ו- VC (C), בהתאמה. (D) היסטוגרמה לעוצמת CTV-A. אנא לחץ כאן כדי להציג גרסה גדולה יותר של איור זה.
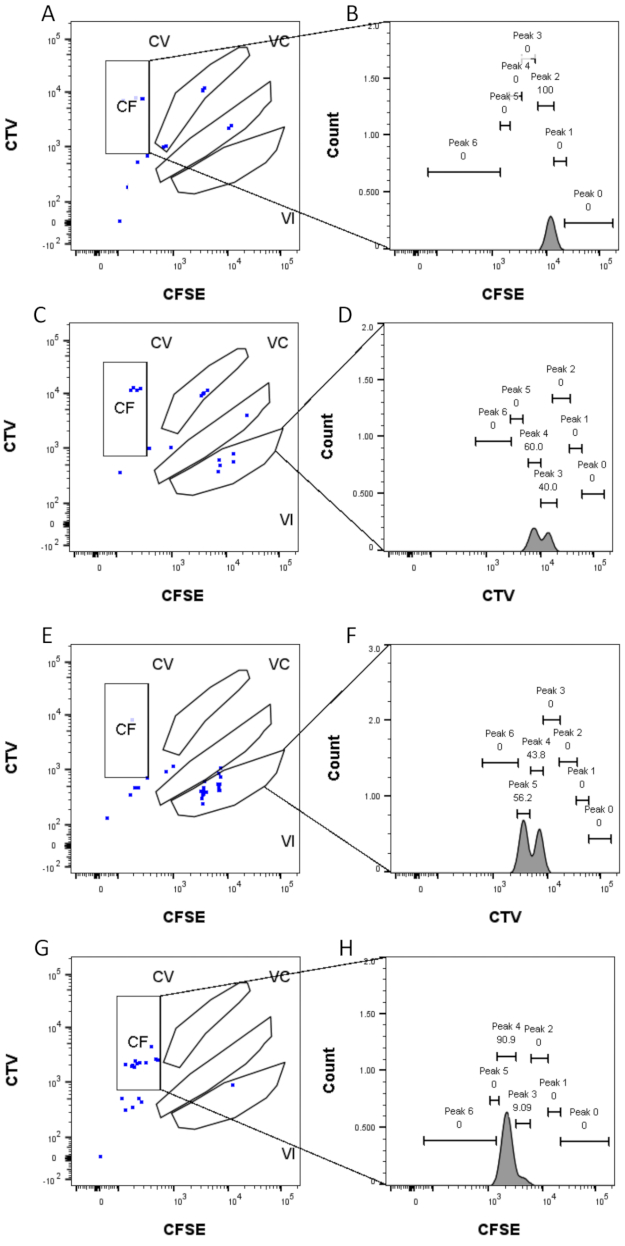
איור 4: מטלת שיא. (A,B) ניתן לזהות רק שיא אחד עבור באר זו, בשער CF. (ג,ד) שתי פסגות בעוצמות כמעט שוות ניתן לזהות בבאר זו, בשער VI. הפסגות נפתרות היטב. (ה,ו) שתי פסגות בעוצמה דומה ניתן לזהות בבאר זו, בשער VI. רק האירועים בשער נלקחו בחשבון, בהתבסס על האסטרטגיה שנקבעה באמצעות בארות הצובר . (ז-ח) שתי פסגות של עוצמה לא שוות ניתן לזהות בבאר זו, בשער CF. אנא לחץ כאן כדי להציג גרסה גדולה יותר של נתון זה.
ייצוג נתונים ובדיקות סטטיסטיות
איור 5 מראה סוגים שונים של ייצוג נתונים של שני ניסויים נפרדים, שניהם בוצעו לאחר 72 שעות של תרבית תאים. HSCs ו-MPPs גודלו בתרבית בשתי תרביות תאים שונות, שאמורות לשנות את תכונות חלוקת התא והתמיינותו. מדיות אלה נקראות "Diff" (בידול)32 ו-"GT"33; הראשון מקדם התמיינות מיאלואידית ואריתרואידית, מכיוון שהוא מכיל אריתרופויאטין (EPO) וגורם מגרה מושבה גרנולו-מונוציטית (GM-CSF), ואילו השני פותח בהקשר של ניסויים קליניים בריפוי גנטי, במטרה לשמר ולהגביר אחוז גבוה של HSPCs. איור 5A הוא מפת חום מייצגת עבור המצב "Diff", המייצג מגוון משפחות תאים, הן בגורלות תאים והן בחלוקות. במפת חום זו, כל שורה מייצגת משפחה בודדת, כל ריבוע תא בודד, והעמודות מקבצות את כל התאים הנמצאים באותו דור (לדוגמה, תאים בדור 2 מחולקים לפחות פעמיים). ניתן להבחין בין משפחות הומוגניות מאוד, המורכבות מסוג תא יחיד ומציגות את אותו מספר חלוקות (למשל, משפחה #63), לבין משפחות הטרוגניות, כולל שלושה סוגי תאים במשך שני דורות (למשל, משפחה #84). מאחר ששיעור ההתאוששות התאית בניתוח זה הוא כ-70%, משפחות שלמות, המוגדרות על-ידי כך שכל התאים שלהן התאוששו בדורות שונים (למשל, משפחה של תא אחד בדור 1 ושני תאים בדור 2), נצפות רק לעתים רחוקות (הצגת האשטאג ליד מספר הזהות שלהן באיור 5A). ישנם הסברים רבים המסבירים את הגילוי החלקי, אשר יכול להיות טכני (בעיית צביעה, אובדן תאים עקב הפרוטוקול) או ביולוגי (מוות תאי ו / או אפופטוזיס). ניתן להתגבר על מגבלות טכניות באמצעות אנלייזר שנועד להפחית את הנפח המת המשויך לדגימה הבודדת, ועל ידי ביצוע צביעת התא ישירות בלוח תרבית התא כדי להפחית את צנרת הנפח. לעומת זאת, שיטות אורתוגונליות לקביעת כמות המוות של תאים (למשל, באמצעות ניסויי הדמיה של תאים חיים) יכולות לעזור להבחין בין הגורמים הטכניים והביולוגיים שתוצאתם גילוי חלקי.
איור 5Bi מראה כיצד לדמיין את ההשפעה של מצב התרבית על הרכב סוג התא, כאילו מישהו ביצע בדיקה בתפזורת. כאן, תנאי Diff מקדם מספר גדול יותר של גורלות, ואחוז גבוה יותר של תאי CD34+ (המוגדרים ככל סוגי התאים למעט CD34-). מרווחי בר-סמך מחושבים בסקריפט באמצעות רצועת אתחול בסיסית, עם 250,000 מערכי נתונים של bootstrapped34. ראוי לציין שכל שאר ההיסטוגרמות באיור 5 מציגות רווחי סמך המחושבים באותו אופן. טבלה 5 משחזרת את כל המידע על מספר המשפחות ומספר התאים בכל דור.
איור 5Bii מייצג באופן גרפי את תפוקת הבדיקות הסטטיסטיות שבוצעו בסקריפט "2_bar_plot". תיאור רשמי של המסגרת הסטטיסטית זמין26. בקצרה, מסגרת זו מאפשרת בדיקת השערות סטטיסטיות תוך הנחה שתאים מאותה משפחה הם תלויים (הנחה שהיא עצמה ניתנת לבדיקה), בניגוד לסטטיסטיקה הקלאסית המחייבת עצמאות בין כל התאים הנצפים. במקרה הספציפי המוצג באיור, הבדיקה הסטטיסטית מאתגרת את ההשערה כי בחירות גורל התא של MPPs, הנמדדות כתדרים של סוגי התאים השונים הקיימים בתרבית, אינן תלויות בתנאי תרבית התא שבהם נעשה שימוש. ראשית, סטטיסטיקת מבחן G משמשת להערכת הפער בין תדרי סוג התא ממדיות תא שונות (לדוגמה ב- Bii, סטטיסטיקה זו מיוצגת באמצעות הסרגל האדום). לאחר מכן, אקראיות של הנתונים מתבצעת באמצעות תמורות, החלפת משפחות שלמות של תאים בין שני תנאי תרבית התא. זאת כדי לשמר את התלות בין תאים הקשורים למשפחה, תוך שמירה על מספר המשפחות בכל קבוצה עקבי עם הנתונים המקוריים. סטטיסטיקת מבחן G מחושבת מתוך ערכת הנתונים האקראית. הערכים הכחולים המיוצגים ב- 5Bii הם סטטיסטיקת מבחן G עבור 250,000 תמורות. לבסוף, ערך p מחושב כדי להעריך את המידה שבה מערך הנתונים המקורי חורג מההתפלגות של הנתונים הקבועים. בדוגמה, הסטטיסטיקה המקורית סוטה במידה רבה מההתפלגות, וכתוצאה מכך ערך p קטן ובכך דוחה את ההשערה שגורל התא של MPPs אינו תלוי בתנאי התרבית.
איור 5C מייצג את אחוז משפחות התאים בכל דור מרבי, כדי לחקור כיצד תנאים שונים משנים את חלוקת התאים לכל משפחת תאים. תרשים נתונים זה מראה כי ב 72 שעות, התאים בתרבית במצב Diff להשלים מספר גדול יותר של חלוקות מאשר תאים במצב GT. מיוצג הוא מספר הדורות המרבי לכל משפחה, כך שמשפחה אחת המציגה תאים בדורות 1 ו -2 נחשבת דור 2. אותה מסגרת סטטיסטית שבה נעשה שימוש באיור 5B יכולה לשמש לבדיקה סטטיסטית של העצמאות בין חלוקת התאים למצב התרבית.
איור 5D בוחן את סוג הסימטריה/אסימטריה של החלוקה הראשונה עבור סוגי האבות הקדמונים השונים (HSCs או MPPs). עבור משפחות התאים השלמות בדור 1 – הדור היחיד שבו ניתן לקבוע בוודאות את שני תאי הבת כתאי אחות – ניתן להגדיר ארבעה סוגים שונים של סימטריה/אסימטריה: התווית "Sym Undiff" מתארת משפחות שבהן שתי הבנות שומרות על הפנוטיפ של תא המוצא. "סים דיף" פירושו שלשתי הבנות יש אותו פנוטיפ, והוא שונה מתא המוצא. "Asym Undiff" פירושו שבת אחת שומרת רק על הפנוטיפ של תא המוצא. לבסוף, "Asym Diff" מתאר משפחות שבהן לשתי הבנות יש פנוטיפים שונים, ואף אחד מהם אינו זהה לתא המוצא. כדי להשיג כוח סטטיסטי בהערכת גורלות סימטריים/א-סימטריים אלה, רצוי לבצע את ניתוח המולטי-ג'ן בנקודות זמן מוקדמות, על מנת לצפות ביותר משפחות שצאצאיהן נמצאים בדור 1.
לבסוף, איור 5E מייצג את האחוזים של סוגי תאים כפונקציה של מספר החטיבות, כדי לקבל תובנות על התקדמות דפוס ההתמיינות בין חלוקות. לדוגמה, הנתונים המוצגים באיור מצביעים על כך שתאים מתקדמים למצב CD34, כאשר מעל 50% מהתאים שזוהו בכיתה זו לאחר שלוש חלוקות בלבד. יתר על כן, ניתן להסיק כי MPPs אינם מעדיפים חלוקה של התחדשות עצמית, שכן אחוז קטן של תאים לשמור על הפנוטיפ המקורי. חלק ממסקנות אלה ניתן לבחון באמצעות המסגרת הסטטיסטית שהוצגה בנתונים הקודמים.
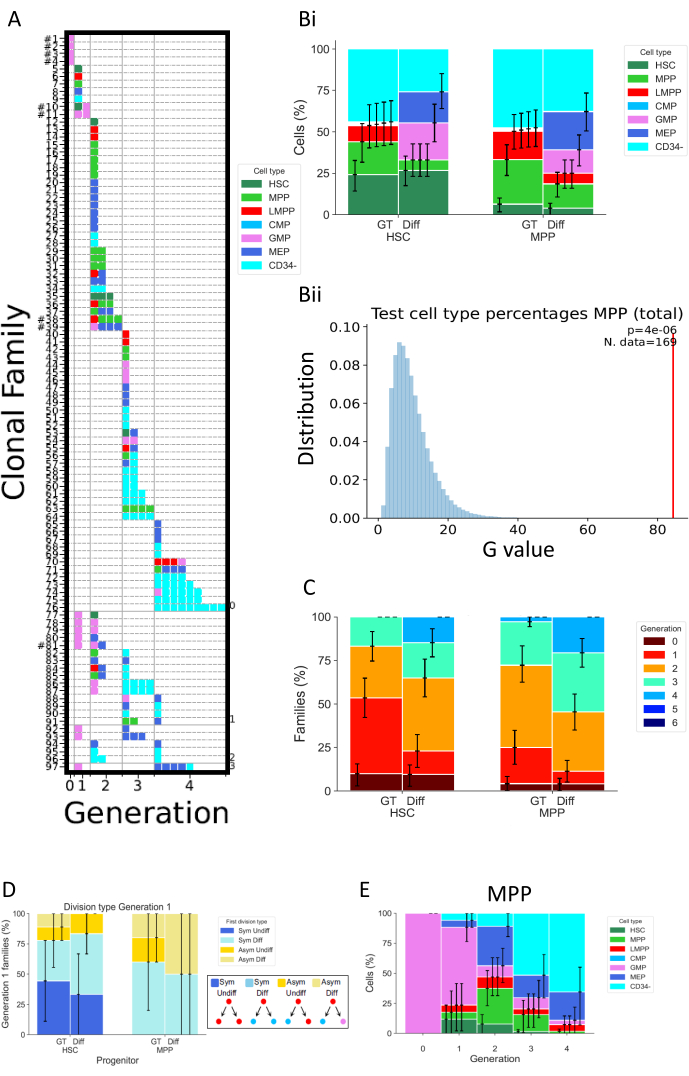
איור 5: דוגמה לייצוג נתונים עבור ניסוי אחד של 72 שעות באמצעות HSPCs מדם טבורי. (A) מפות חום עבור מערך נתונים נבחר (HSC, בתווך "Diff", לאחר 72 שעות של תרבית). החלקות מייצגות את כל התאים הבודדים (ריבועים) על פי קרבתם (שורות), מספר החלוקות שבוצעו (עמודות, הנקראות דור) ופנוטיפ (צבעים). (בי) היסטוגרמה המשווה פרופורציות של סוגי התאים של צאצאי התא של HSCs ו- MPPs, בין המצב GT לבין המצב Diff. (Bii) הגרף מייצג את הבדיקות הסטטיסטיות שבוצעו בסקריפט "2_bar_plot" עבור MPPs בתרבית 72 שעות, בהשוואה בין קוקטיילים ציטוקינים "Diff" ו- "GT". ערך הניסוי מוצג באדום, והערכים נוצרים באמצעות 250,000 תמורות בכחול. ערך p של מבחן G מצוין בפינה הימנית העליונה עם מספר המשפחות ששימשו לבדיקה. (C) היסטוגרמה המשווה את אחוז המשפחות (314 משפחות בסך הכל) בכל דור (מקודד בצבע), עבור HSCs ו- MPPs לכל תנאי תרבית. מרווחי בר-סמך מחושבים באמצעות 250,000 ערכות נתונים עם אתחול. (D) היסטוגרמה המייצגת את סוג הסימטריה/אסימטריה בין גורל תאי הבת למשפחות בעלות שני תאים בדור 1: סים אונדיף (שתי הבנות שומרות על הפנוטיפ של תא המוצא), סים דיף (לשתי הבנות יש אותו פנוטיפ, והוא שונה מתא המוצא), אסים אונדיף (רק בת אחת שומרת על הפנוטיפ של תא המוצא), ו- Asym Diff (לשתי הבנות יש פנוטיפים שונים ואף אחד מהם אינו דומה לתא המוצא). (ה) היסטוגרמות של תרומתם של סוגי תאים המסווגים לדור עבור MPPs שגודלו בתרבית עם קוקטייל "Diff"; n = 204 תאים ו-97 משפחות. אנא לחץ כאן כדי להציג גרסה גדולה יותר של איור זה.
טבלה 5: תיאור מספר המשפחות והתאים שנותחו לכל תנאי ניסוי (תא מקור ותווך תרבית תאים). אנא לחץ כאן כדי להוריד טבלה זו.
קובץ משלים 1: אנא לחץ כאן להורדת קובץ זה.
קובץ משלים 2: לחץ כאן להורדת קובץ זה.
קובץ משלים 3: אנא לחץ כאן להורדת קובץ זה.
קובץ משלים 4: לחץ כאן להורדת קובץ זה.
בדיקת MultiGen היא בדיקה בעלת תפוקה גבוהה, קלה לביצוע ולא יקרה, שסייעה לחקור לימפוציטים 23,24,35 ותאים המטופויטיים מורין26,27. כאן אנו מציגים פיתוח חדש של הגישה המאפשרת לפענח ex vivo את השלב המוקדם של מחויבות HSPC אנושית, ברמת התא הבודד באמצעות תרבית לטווח קצר (איור 6). מערכות תרבית אקס-ויו של תא בודד משמשות בדרך כלל להערכת גורלם ארוך הטווח של HSPCs לתאים בוגרים, אך חלק מהגורלות מתרחשים מוקדם יותר מאחרים36, מה שעלול להטות את הניתוח לכיוון פחות גורלות. בנוסף, מערכות תרבות אלה בדרך כלל מפספסות מידע על מחלוקות במהלך מחויבות גורל. צעדי המחויבות הראשונים הוכחו כמתרחשים כבר בתחילת התרבות, לעתים ללא חלוקה26,37, מה שהופך את התרבות לטווח קצר ואת חלוקת המעקב לחיונית לחקר מחויבות הגורל המוקדם. על ידי מעקב בו זמנית אחר הגורל, החלוקה והקרבה, בדיקה זו מאפשרת להבין את תפקידה של החלוקה הראשונה והחלטת הגורל ב- HSPCs אנושיים. באמצעות הבדיקה ניתן להסיק לאחר כמה חלוקות מתרחש תהליך המחויבות, את האיזון בין התחדשות עצמית לבין בידול עבור אותם אבות מוקדמים, וכיצד תכונות אלה עוברות בירושה לאורך דורות. למיטב ידיעתנו, זהו המבחן היחיד המאפשר מדידות מסוג זה עבור HSPCs אנושיים, ברזולוציה של תא בודד. בנוסף, באמצעות שילובים שונים של צבעי חלוקת תאים, הגדלנו את התפוקה של הניתוח, מה שהופך את הבדיקה הזו לכלי רב ערך ליצירת מערכי נתונים גדולים במהירות. שילובי הצבעים מאפשרים לעקוב אחר מספר משפחות באותן בארות, ולהגדיל את מספר התאים הזמינים לניתוח בתרבית לטווח קצר. מספר השילובים יכול להיות גדול עוד יותר, על ידי הוספת צבעים אחרים (למשל, צבע צהוב) או שינוי היחס בין CFSE ו- CTV. עם זאת, זה מקטין את מספר הפרמטרים האחרים שניתן לנתח.
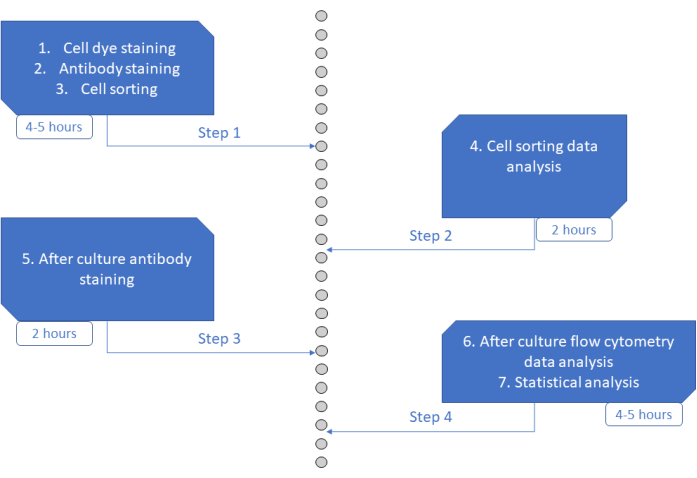
איור 6: ייצוג סכמטי של הפרוטוקול. לחץ כאן כדי להציג גרסה גדולה יותר של איור זה.
כדי לבצע את הניתוח בהצלחה, בשל מספר גדול של בארות ואת מספר מופחת של תאים לנתח, יש צורך להפעיל את ניתוח ציטומטריה זרימה על מנתח מצויד קורא לוחות. הדור החדש של מנתחי ספסל מותאם במיוחד לבדיקה זו, מכיוון שלרובם יש נפח מת קטן יותר כדי להפחית את אחוז אובדן התאים. זה בתורו מבטיח יעילות גבוהה יותר בשחזור כל באר, מה שמביא ליעילות המוערכת בטווח של70% 26. הערכת אובדן התאים במהלך רכישת ציטומטריית הזרימה חיונית לניתוח של כל משפחה בנפרד. לדוגמה, בהנחה שאין מוות תאי וסופרים את מספר החלוקות, ניתן להעריך את מספר התאים לכל משפחה. עם זאת, רצוי לבצע כמה ניסויים מאששים, במיוחד בהערכת מוות תאי בתנאי התרבית הנבדקת ומדידת קצב ההתאוששות באופן ניסיוני באמצעות מספר מוגדר של תאים.
אחד השלבים המכריעים של פרוטוקול זה הוא הקצאת השיא. כאמור, התפלגות שיא באיכות טובה תלויה מאוד בבידוד של פסגות צרות מאוד במיון תאים. עם זאת, עדיין קשה להקצות את המספר הנכון של חטיבות בהתבסס באופן ייחודי על ההתפלגות. מכיוון שמיון תאים וניתוח ציטומטריית זרימה מבוצעים בשתי מכונות שונות, לא ניתן להשוות ישירות את עוצמת כל אות, כך שיהיה קשה לדעת אם השיא הראשון שנצפה בקצה הימני של ההיסטוגרמה הוא שיא 0 או שיא 1. בהקשר זה, פתרונות מעטים אפשריים; דרך אחת היא לבצע ניסוי אורתוגונלי כדי למדוד במדויק את מספר החלוקות המבוצעות על ידי תאים אלה (למשל, דימות תאים חיים). אפשרות נוספת היא פשוט לספור את מספר התאים בבאר תחת מיקרוסקופ שדה בהיר הפוך, לפני ביצוע ניתוח ציטומטריית זרימה. זה יסיק מספר ממוצע של חלוקות (בהנחה שאין מוות תאי). לבסוף, פתרון פוסט-הוק להקצאת שיא הוא איתור מספר חריג של "משפחות בלתי אפשריות"; משפחות אלה מורכבות ממספר גדול מהאפשרי של תאים בכל דור (למשל, חמישה תאים בדור 2, או שני תאים בדור 1 ותא אחד בדור 2). האפשרות להחריג משפחות בלתי אפשריות מקודדת בשלב הניתוח הסטטיסטי, ומסמנת את המשפחה הבלתי אפשרית. אם ההתרחשות של שגיאות אלה גבוהה מדי, סביר להניח כי הקצאת השיא צריכה להיות מתוקנת.
בפרוטוקול זה, כללנו כמה דוגמאות של ייצוג וניתוח נתונים עבור הבדיקה, שכן זה הפך להיות צעד חיוני ביצירה ופרשנות של מערכי נתונים גדולים38. הדוגמה הראשונה היא מפת החום המציגה את מכלול התאים המנותחים, המאורגנים לפי משפחה. זהו כלי יעיל לחקור את המאפיינים הכלליים של הנתונים ואת המסקנות האפשריות: האם משפחות מורכבות מסוגי תאים מרובים או שהן נוטות להיות הומוגניות בהרכבן? האם משפחות מפוזרות על פני דורות רבים, או שהן מתחלקות לרוב באותו מספר פעמים? ניתוח גישוש זה צריך להיות משלים עם חלקות ספציפיות יותר ובדיקות סטטיסטיות. ניתן להשתמש בו כדי להעריך כמותית מחויבות גורל סימטרית וא-סימטרית, דיפרנציאציה ללא חלוקה, האיזון בין התחדשות עצמית להתבדלות, ומספר החלוקות עבור גורל מחויבות נתון. זה בסיסי, במהלך תכנון הניסוי, להגדיר את אורך תרבית התא בהתאם לסוג השאלה שנשאלה; לדוגמה, בשתי השאלות הראשונות (איזון סימטרי/א-סימטרי ודיפרנציאציה ללא חלוקה), תכנון צעדי תרבות קצרים מאוד מאפשר בידוד של מספר רב של משפחות שביצעו רק חלוקה אחת או כלל לא ביצעו חלוקה כלל26. לעומת זאת, ניסויים ארוכים יותר מאפשרים לחקור את מספר החלוקות הדרושות למחויבות תא מסוים, שכן הם דוגמים משפחות בשלבים שונים של התמיינות. עם זאת, שיטה זו אינה מיועדת לתרביות ארוכות טווח (2-3 שבועות), מכיוון שדילול צבע התא אינו מסוגל לעקוב במדויק אחר יותר משבע או שמונה חטיבות22. כתוצאה מכך, כלי זה מותאם בעיקר לחקר המחויבות המוקדמת של אבות hematopoietic, והוא לא נועד להסיק מסקנות מוצקות על תכונות התמיינות לטווח ארוך של תאים אלה.
המסגרת הסטטיסטית פותחה במיוחד לניתוח נתונים מסוג זה ומבוססת על הרעיון של תמורות26. זה היה הכרחי בגלל התצפית של תלות משפחתית על התפלגות סוג התא ועל מספר החטיבות שבוצעו. במילים אחרות, תאים שהם חלק מאותה משפחה נוטים יותר להציג פנוטיפים דומים ולהתחלק באותו מספר פעמים. בעוד ניתוח מעמיק הוא מעבר להיקף של עבודה זו, קבוצה מסופקת של בדיקות סטטיסטיות צריך להיות מספיק בעת הערכת תנאים שונים.
לסיכום, פרוטוקול זה מהווה כלי רב ערך להערכת הדינמיקה התאית של תאי גזע המטופויטיים ותאי אב ex vivo, בצורה מהירה וזולה. בשל גמישותו ורבגוניותו לגבי נקודת זמן, תנאי תרבית וסוג HSPCs שנותחו, הוא מאפשר לבדוק מגוון תנאי ניסוי. כניסוי מבוסס ציטומטריית זרימה, ניתן ליישם אותו ברוב המעבדות, והוא אינו דורש ידע מוקדם נרחב, מה שהופך אותו למועמד טוב לבדיקות סקר וניסויי פיילוט.
המחברים מצהירים כי אין ניגוד עניינים רלוונטי לעבודה זו. למממנים לא היה כל תפקיד בעיצוב המחקר, באיסוף הנתונים ובפרשנותם, או בהחלטה להגיש את העבודה לפרסום.
ברצוננו להודות לחברי מכון קירי על עזרתם בהקמת ניסויי ציטומטריית הזרימה. אנו רוצים גם להכיר בתרומתם של חברי הצוות האחרים, במהלך דיונים מרובים. אנו מודים לד"ר ג'וליה מרצ'ינגו ולפרופ' פיל הודג'קין (מכון וולטר אנד אלייזה הול למחקר רפואי) על שיתוף הפרוטוקול שלהם של ריבוב צבעי חלוקת תאים על לימפוציטים. אנו מודים לביובנק הדם הטבורי של בית החולים סנט לואיס על אספקת המשאבים הביולוגיים הדרושים לפיתוח פרוטוקול זה. המחקר נתמך על ידי מענק ATIP-Avenir מ-CNRS ומקרן Bettencourt-Schueller (ל-L.P.), מענקים מ-Labex CelTisPhyBio (ANR-10-LBX-0038) (ל-L.P. ו-A.D.), Idex Paris-Science-Lettres Program (ANR-10-IDEX-0001-02 PSL) (ל-L.P.), Canceropole INCA Emergence (2021-1-EMERG-54b-ICR-1, ל-L.P.), ומענק ITMO MIIC (21CM044, ל-L.P). בנוסף למימון ממועצת המחקר האירופית (ERC) במסגרת תוכנית המחקר והחדשנות Horizon 2020 של האיחוד האירופי ERC StG 758170-Microbar (ל- L.P.), A.D. נתמכה על ידי מלגה מקרן דה פראנס.
| Name | Company | Catalog Number | Comments |
| 1.5 mL polypropylene microcentrifuge tubes | vWR | 87003-294 | |
| 15-mL polypropylene tubes | vWR | 734-0451 | |
| 50-mL polypropylene tubes | vWR | 734-0448 | |
| 96-well U-bottom culture plate | Falcon | 353077 | |
| Anti-human Lin APC | Thermo Fisher | 22-7776-72 | Dilution 1/40 |
| ARIA III | BD | Can be replaced with any FACS sorter able to sort individual cells in 96-wells plate | |
| Carboxyfluorescein succinimidyl ester (CFSE) | Life Technologies | C34570 | |
| Cell Trace Violet (CTV) | Life Technologies | C34571 | |
| Compensation beads | BD | 552843 | |
| Dulbecco’s modified Eagle medium (DMEM) | Life Technologies | 11320033 | |
| Ethylenediaminetetraacetic acid (EDTA) | Thermo Scientific | J62948-36 | Prepare a solution 0.5 M, in sterilised water |
| FC block Fc1.3216 | BD | 564220 | Dilution 1/50 |
| Fetal Bovine Serum (FBS) | Dutscher | S1900-500C | Batch S00CH |
| FlowJo v10.8.1 | BD | ||
| Mouse anti-human CD10 PerCP-5.5, clone HI10a | Biolegend | 312216 | Dilution 1/20 |
| Mouse anti-human CD123 BUV395, clone 7G3 | BD | 564195 | Dilution 1/15 |
| Mouse anti-human CD34 APC-Cy7, clone 581 | Biolegend | 343513 | Dilution 1/40 |
| Mouse anti-human CD38 BV650, clone HB7 | Biolegend | 356620 | Dilution 1/40 |
| Mouse anti-human CD45RA AF700, clone HI100 | BD | 560673 | Dilution 1/20 |
| Mouse anti-human CD45RA PE-Cy7, clone HI100 | BD | 560675 | Dilution 1/20 |
| Mouse anti-human CD90 PE, clone 5E10 | Biolegend | 328110 | Dilution 1/20 |
| Phosphate Buffered Saline (PBS) 1X | Life Technologies | 10010001 | |
| Python | |||
| R | |||
| Sterile 12x75 mm conical polypropylene tubes | Falcon | ||
| ZE5 | Biorad | Can be replaced with any flow cytometry analyzer equipped with a plate reader | |
| Laboratory prepared | |||
| Cell culture media | Depends from the specific experiment. Prepare fresh daily and store at +4 °C until use | ||
| DMEM + 10% FBS | Can be stored in sterile conditions, at +4 °C up to 1 year. To prepare 500 mL, add 50 mL of FBS to 450 mL DMEM | ||
| PBS 1X + EDTA 0.1% | Can be stored in sterile conditions, at room temperature, up to 1 year. To prepare 500 mL, add 3.42 mL of EDTA 0.5 M to 500 mL PBS 1X | ||
| Staining buffer | Can be stored in sterile conditions, at +4 °C up to 1 year. To prepare 500 mL, add 2 mL of EDTA 0.5 M and 1 mL FBS to 500 mL PBS 1X |
- Ginhoux, F., Yalin, A., Dutertre, C. A., Amit, I. Single-cell immunology: Past, present, and future. Immunity. 55 (3), 393-404 (2022).
- Ke, M., Elshenawy, B., Sheldon, H., Arora, A., Buffa, F. M. Single cell RNA-sequencing: A powerful yet still challenging technology to study cellular heterogeneity. Bioessays. 44 (11), 2200084 (2022).
- Regev, A., et al. The human cell atlas. Elife. 6, 27041 (2017).
- Laurenti, E., Göttgens, B. From haematopoietic stem cells to complex differentiation landscapes. Nature. 553 (7689), 418-426 (2018).
- Haas, S., Trumpp, A., Milsom, M. D. Causes and consequences of hematopoietic stem cell heterogeneity. Cell Stem Cell. 22 (5), 627-638 (2018).
- Loughran, S. J., Haas, S., Wilkinson, A. C., Klein, A. M., Brand, M. Lineage commitment of hematopoietic stem cells and progenitors: insights from recent single cell and lineage tracing technologies. Experimental Hematology. 88, 1-6 (2020).
- Perié, L., Duffy, K. R. Retracing the in vivo haematopoietic tree using single-cell methods. FEBS Letters. 590 (22), 4068-4083 (2016).
- Yu, V. W. C., et al. Epigenetic memory underlies cell-autonomous heterogeneous behavior of hematopoietic stem cells. Cell. 167 (5), 1310-1322 (2016).
- Ganuza, M., et al. Lifelong haematopoiesis is established by hundreds of precursors throughout mammalian ontogeny. Nature Cell Biology. 19 (10), 1153-1163 (2017).
- Naik, S. H., Schumacher, T. N., Perié, L. Cellular barcoding: A technical appraisal. Experimental Hematology. 42 (8), 598-608 (2014).
- Quek, L., et al. Genetically distinct leukemic stem cells in human CD34 − acute myeloid leukemia are arrested at a hemopoietic precursor-like stage. The Journal of Experimental Medicine. 213 (8), 1513-1535 (2016).
- Karamitros, D., et al. Single-cell analysis reveals the continuum of human lympho-myeloid progenitor cells. Nature Immunology. 19 (1), 85-97 (2018).
- Boitano, A. E., et al. Aryl hydrocarbon receptor antagonists promote the expansion of human hematopoietic stem cells. Science. 329 (5997), 1345-1348 (2010).
- Delaney, C., et al. Notch-mediated expansion of human cord blood progenitor cells capable of rapid myeloid reconstitution. Nature Medicine. 16 (2), 232-236 (2010).
- Fares, I., et al. Cord blood expansion. Pyrimidoindole derivatives are agonists of human hematopoietic stem cell self-renewal. Science. 345 (6203), 1509-1512 (2014).
- Guo, B., Huang, X., Lee, M. R., Lee, S. A., Broxmeyer, H. E. Antagonism of PPAR-γ 3 signaling expands human hematopoietic stem and progenitor cells by enhancing glycolysis. Nature Medicine. 24 (3), 360-367 (2018).
- Vannini, N., et al. The NAD-booster nicotinamide riboside potently stimulates hematopoiesis through increased mitochondrial clearance. Cell Stem Cell. 24 (3), 405-418 (2019).
- Gupta, R., et al. Nov/CCN3 enhances cord blood engraftment by rapidly recruiting latent human stem cell activity. Cell Stem Cell. 26 (4), 527-541 (2020).
- Horwitz, M. E., et al. Omidubicel vs standard myeloablative umbilical cord blood transplantation: results of a phase 3 randomized study. Blood. 138 (16), 1429-1440 (2021).
- Weinreb, C., Rodriguez-Fraticelli, A., Camargo, F. D., Klein, A. M. Lineage tracing on transcriptional landscapes links state to fate during differentiation. Science. 367 (6479), 3381 (2020).
- Loeffler, D., Schroeder, T. Understanding cell fate control by continuous single-cell quantification. Blood. 133 (13), 1406-1414 (2019).
- Tario, J. D., et al. Optimized staining and proliferation modeling methods for cell division monitoring using cell tracking dyes. Journal of Visualized Experiments. (70), e4287 (2012).
- Marchingo, J. M., et al. T-cell stimuli independently sum to regulate an inherited clonal division fate. Nature Communications. 7, 13540 (2016).
- Horton, M. B., et al. Multiplexed division tracking dyes for proliferation-based clonal lineage tracing. Journal of Immunology. 201 (3), 1097-1103 (2018).
- Lehmann, E. L., Romano, J. P., Casella, G. . Testing statistical hypotheses. , 784 (2005).
- Tak, T., et al. HSPCs display within-family homogeneity in differentiation and proliferation despite population heterogeneity. Elife. 10, 360624 (2021).
- Sommerkamp, P., et al. Mouse multipotent progenitor 5 cells are located at the interphase between hematopoietic stem and progenitor cells. Blood. 137 (23), 3218-3224 (2021).
- Kato, K., Radbruch, A. Isolation and characterization of CD34+ hematopoietic stem cells from human peripheral blood by high-gradient magnetic cell sorting. Cytometry. 14 (4), 384-392 (1993).
- Miltenyi, S., Müller, W., Weichel, W., Radbruch, A. High gradient magnetic cell separation with MACS. Cytometry. 11 (2), 231-238 (1990).
- Doulatov, S., et al. Revised map of the human progenitor hierarchy shows the origin of macrophages and dendritic cells in early lymphoid development. Nature Immunology. 11 (7), 585-593 (2010).
- Goardon, N., et al. Coexistence of LMPP-like and GMP-like leukemia stem cells in acute myeloid leukemia. Cancer Cell. 19 (1), 138-152 (2011).
- Laurenti, E., et al. CDK6 levels regulate quiescence exit in human hematopoietic stem cells. Cell Stem Cell. 16 (3), 302-313 (2015).
- Aiuti, A., et al. Lentiviral hematopoietic stem cell gene therapy in patients with Wiskott-Aldrich syndrome. Science. 341 (6148), 1233151 (2013).
- Davison, A. C., Hinkley, D. V. . Bootstrap Methods and their Application. , (1997).
- Horton, M. B., et al. Lineage tracing reveals B cell antibody class switching is stochastic, cell-autonomous, and tuneable. Immunity. 55 (10), 1843-1855 (2022).
- Notta, F., et al. Distinct routes of lineage development reshape the human blood hierarchy across ontogeny. Science. 351 (6269), 2116 (2016).
- Grinenko, T., et al. Hematopoietic stem cells can differentiate into restricted myeloid progenitors before cell division in mice. Nature Communications. 9 (1), 1898 (2018).
- Saeys, Y., Van Gassen, S., Lambrecht, B. N. Computational flow cytometry: Helping to make sense of high-dimensional immunology data. Nature Reviews Immunology. 16 (7), 449-462 (2016).
Tags
ABOUT JoVE
Copyright © 2024 MyJoVE Corporation. All rights reserved