
Summary
Abstract
Introduction
Protocol
Representative Results
Discussion
Acknowledgements
Materials
References
Biology
Karakterisering van epitheliale wondgenezing in vivo met behulp van het Cnidarische modelorganisme Clytia hemisphaerica
Dit artikel beschrijft een methode om wonden te creëren in het epitheel van een levende Clytia hemisphaerica medusa en wondgenezing in beeld te brengen met een hoge resolutie in vivo. Daarnaast wordt een techniek gepresenteerd om kleurstoffen en medicijnen te introduceren om signaalprocessen in de epitheelcellen en extracellulaire matrix tijdens wondgenezing te verstoren.
Alle dierlijke organen, van de huid tot ogen tot darmen, zijn bedekt met vellen epitheelcellen waarmee ze homeostase kunnen behouden en tegelijkertijd worden beschermd tegen infecties. Daarom is het niet verwonderlijk dat het vermogen om epitheliale wonden te herstellen van cruciaal belang is voor alle metazoën. Epitheliale wondgenezing bij gewervelde dieren omvat overlappende processen, waaronder ontstekingsreacties, vascularisatie en re-epithelisatie. Regulatie van deze processen omvat complexe interacties tussen epitheelcellen, naburige cellen en de extracellulaire matrix (ECM); de ECM bevat structurele eiwitten, regulerende eiwitten en actieve kleine moleculen. Deze complexiteit, samen met het feit dat de meeste dieren ondoorzichtige weefsels en ontoegankelijke ECM's hebben, maakt wondgenezing moeilijk te bestuderen bij levende dieren. Veel werk aan epitheliale wondgenezing wordt daarom uitgevoerd in weefselkweeksystemen, met een enkel epitheelceltype dat als een monolaag op een kunstmatige matrix is geplateerd. Clytia hemisphaerica (Clytia) biedt een unieke en opwindende aanvulling op deze studies, waardoor epitheliale wondgenezing kan worden bestudeerd in een intact dier met een authentieke ECM. Het ectodermale epitheel van Clytia is een enkele laag van grote plaveiselepitheelcellen, waardoor beeldvorming met hoge resolutie mogelijk is met behulp van differentiële interfererende contrast (DIC) microscopie bij levende dieren. De afwezigheid van migrerende fibroblasten, vasculatuur of ontstekingsreacties maakt het mogelijk om de kritieke gebeurtenissen bij re-epithelisatie in vivo te ontleden. De genezing van verschillende soorten wonden kan worden geanalyseerd, waaronder eencellige microwounds, kleine en grote epitheelwonden en wonden die het keldermembraan beschadigen. Lamellipodia-vorming, samentrekking van de portemonnee, celrekking en collectieve celmigratie kunnen allemaal in dit systeem worden waargenomen. Bovendien kunnen farmacologische middelen via de ECM worden geïntroduceerd om cel:ECM-interacties en cellulaire processen in vivo te wijzigen. Dit werk toont methoden voor het maken van wonden in levende Clytia, het vastleggen van films van genezing en het onderzoeken van genezingsmechanismen door micro-injecterende reagentia in de ECM.
Vellen epitheelcellen bedekken het externe oppervlak van alle metazoën, bekleden interne organen en verdelen het dierlijke lichaam in afzonderlijke compartimenten. Het epitheel scheidt ook het binnenlichaam van de externe omgeving en beschermt het tegen schade en infectie. Vandaar dat de komst van epitheellagen een essentieel onderdeel was van de evolutie van meercellige dieren, en epitheliale lagen worden gezien bij alle dieren, van gewervelde dieren tot de meest basale metazoën1. Het epitheel van sommige organen is een enkele monolaag, zoals in de longluchtzakken, bloedvaten en darm2, evenals in de opperhuid van ongewervelde dieren zoals planaria en cnidarians3. In andere weefsels, zoals de huid4 en het hoornvlies5 van gewervelde dieren, is het epitheel gestratificeerd, wat betekent dat er meerdere epitheelcellagen2 zijn. In alle gevallen wordt de meest basale epitheellaag op het keldermembraan aangebracht, een eiwitplaat die een gespecialiseerd gebied van de extracellulaire matrix (ECM)6,7,8 vormt.
Breuken in het epitheel moeten snel worden gerepareerd om een doorlopende epitheelplaat te creëren. Schade aan het epitheel treedt op tijdens natuurlijke processen, zoals het afstoten van epitheelcellen in de darm,9,10 en als gevolg van ontsteking of fysiek trauma. Wanneer een enkele epitheelcel beschadigd is, moet deze zichzelf herstellen of worden geëlimineerd om de omliggende cellen in staat te stellen zich aan elkaar te hechten en het gat11,12 te sluiten. In wonden groter dan de grootte van een enkele cel moeten epitheelcellen bewegen om elkaar te bereiken en het blad13 te repareren. Dit kan worden bereikt door celverspreiding als de openingen klein zijn of kan de migratie van epitheelcellen uit de marges van een wond vereisen om de wondspleet te dichten; Dit laatste proces wordt re-epithelisatie14,15 genoemd. In embryonale weefsels verspreiden epitheelcellen zich en migreren ze naar dichte wonden of worden ze over de opening getrokken door de samentrekking van actomyosinekabels die zich vormen tussen de cellen aan de wondrand, in een mechanisme dat lijkt op een portemonneestring16. In veel volwassen weefsels omvat re-epithelisatie de migratie van coherente celvellen, waarbij cellen hun verbindingen met naburige cellen behouden14,17,18. In andere weefsels worden cel-celverbindingen ontmanteld en gedragen epitheelcellen zich meer als mesenchymale cellen, die op een gecoördineerde maar onafhankelijke manier in het wondgebied bewegen tijdens re-epithelisatie 14,19,20,21.
Epitheelcelbewegingen worden gereguleerd door complexe interacties tussen de migrerende cellen en tussen de cellen en de ECM. Hoewel er een enorme hoeveelheid experimentele literatuur is over mechanismen van wondactivering van epitheelcellen en daaropvolgende migratie, moet er nog veel worden ontdekt. Het initiële signaal dat epitheelcellen activeert om te migreren als reactie op een wond is bijvoorbeeld niet definitief geïdentificeerd 22, noch is het volledig begrepen hoe actine opnieuw wordt ingezet om lamellipodia te creëren aan de kant van epitheelcellen die zich het dichtst bij de wond bevinden 22,23,24,25,26,27. Collectieve celmigratie vereist dat informatie van cellen aan de wond wordt gedeeld met cellen die distaal zijn van de wond, en de communicatieroute is nog steeds onduidelijk28. Cel:cel juncties en cel:ECM bijlagen moeten worden gedemonteerd en hervormd als cellen in het blad zichzelf herschikken, maar de regulatie van dit proces is slecht begrepen14,29. Vooruitgang boeken op deze en andere gerelateerde vragen is niet alleen belangrijk als een fundamenteel biologisch probleem, maar ook vanwege de klinische betekenis van correcte wondgenezing. Ziekten die het vermogen van epitheelcellen om correct te migreren in gevaar brengen, resulteren in chronische wonden; een voorbeeld is de genetische ziekte epidermolysis bullosa, waarbij genen die betrokken zijn bij de aanhechting van de epitheelcellen aan de ECM worden gemuteerd, wat resulteert in een kwetsbare huid die schilfert en blaren geeft. Re-epithelisatie is ook gecompromitteerd in natuurlijk verouderende weefsels30,31. Een beter begrip is daarom essentieel voor het ontwikkelen van interventies om de wondgenezingsresultaten te verbeteren.
Epitheelcelmigratie bij wondgenezing is bestudeerd met behulp van zowel in vitro benaderingen als modelorganismen. De meeste studies naar wondgenezing en mechanismen van celmigratie zijn uitgevoerd in weefselkweek, waarbij monolagen van een enkel epitheelceltype worden gekweekt op een substraat dat de ECM vervangt. Celmonolagen worden bekrast of gekweekt met stencils om openingen van specifieke vormen en maten te creëren en vervolgenswaargenomen 32,33,34. Het in vitro model maakt een ideale visualisatie van celgedrag mogelijk, evenals de mogelijkheid om kwaliteiten van het substraat te veranderen, om cellen bloot te stellen aan geneesmiddelen en abiotische en biotische factoren, en om cellen te transfecteren met constructies die verschillende genen van belang uitdrukken of onderdrukken. Deze reductionistische benadering kan er echter niet in slagen om enkele van de belangrijke parameters die betrokken zijn bij het gedrag van epitheelcellen in een in vivo context vast te leggen, waaronder communicatie tussen verschillende celtypen en signaleringsgebeurtenissen die zich voordoen in de ECM11. In vivo modellen bieden de authentieke context van een wond, met meerdere celtypen, overlappende signaalwegen en een complexe ECM35. Een dergelijk model voor wondgenezingsstudies is de muis19, waarin recente ontwikkelingen onderzoekers in staat hebben gesteld epidermale cellen te observeren tijdens de genezing van wonden van volledige dikte bij levende dieren36. De muis en andere in vivo systemen vormen echter uitdagingen om re-epithelisatie te bestuderen. Ten eerste wordt het grote voordeel van het observeren van celgedrag in een natuurlijke context gecompenseerd door de complexiteit van de tijdelijk overlappende gebeurtenissen die optreden tijdens de genezing van gewervelde wonden, waaronder bloedstolling, rekrutering van immuuncellen en ontsteking, rekrutering van fibroblasten en celdedifferentiatie, re-vascularisatie en remodellering van de ECM. Verder maken ondoorzichtige weefsels beeldvorming moeilijk. De Drosophila-larve en zebravis epidermissystemen 37,38 hebben een aantal van deze problemen overwonnen vanwege hun relatieve eenvoud39.
Ons lab introduceerde onlangs een nieuw model voor het bestuderen van epitheliale wondgenezing: de medusa (kwallen) vorm van de hydrozoaire cnidarian Clytia hemisphaerica (Clytia)40. Clytia is een opkomend modelorganisme met een volledig gesequenced en geannoteerd genoom41, eencellig RNAseq-transcriptoom 42 en protocollen voor genoommodificatie (mutagenese en transgenese)43,44,45. Cnidarians zijn een van de oudste bestaande afstammingslijnen met epitheliale lagen, dus het begrijpen van cnidarian wondgenezing biedt inzicht in de voorouderlijke paden die epitheelintegriteit verzekerden. Voor die paden die bewaard zijn gebleven in de boom des levens, biedt Clytia een opwindend nieuw systeem om de epitheelceldynamica en de functionele regulatie van wondgenezing in vivo te bestuderen.
Het epitheel dat het bovenoppervlak van de Clytia medusa (exparaplu) bedekt, is een monolaag van transparante, plaveiselepitheelcellen die ongeveer 50 μm breed en 1-2 μm dik zijn (figuur 1). Ze zijn bevestigd aan een ECM genaamd de mesoglea - de "gelei" van de kwal. De mesoglea is qua samenstelling vergelijkbaar met de ECM die wordt aangetroffen bij andere dieren 46,47,48 inclusief gewervelde dieren, heeft een keldermembraan 40 en is volledig transparant. De epitheellaag in de Clytia medusa kan gemakkelijk worden bekrast of gewond (zie hieronder). De eenvoud en transparantie van het epitheel en ECM maakt hoge resolutie beeldvorming van de cellen en hun bewegingen tijdens genezing mogelijk. Onlangs hebben Kamran et al. de genezing van kleine wonden in het Clytia-epitheel in detail gekarakteriseerd40. Er werd aangetoond dat genezing in Clytia plaatsvindt door lamellipodia-gebaseerde celkruipen, celverspreiding en collectieve celmigratie, evenals portemonnee-stringsluiting die meer typerend is voor embryonale systemen (hoewel eerder gezien in volwassen dierlijke structuren zoals het hoornvlies49). Clytia wondgenezing is extreem snel, zoals is gezien in andere systemen die een ontstekingsreactie missen40,50. Genezing in de Clytia-exumbrella is volledig afhankelijk van bewegingen van de bestaande epitheelcellen - geen cellen prolifereren of migreren door de ECM naar de wondplaats (aanvullende film 1). Al deze bevindingen suggereren dat Clytia een nuttig modelsysteem is om epitheliale wondgenezing te bestuderen. Inderdaad, het gemak van het afbeelden van epitheelcellen in Clytia tijdens wondgenezing leidde tot de ontdekking dat epitheelcellamellipodia zich uitstrekken en verspreiden over gebieden van blootgestelde ECM zolang er een intact keldermembraan is; Als het keldermembraan beschadigd is, schakelt epitheliale genezing over op een portemonneesnaarmechanisme40. Dit was de eerste demonstratie van een mechanisme dat ten grondslag ligt aan de beslissing om te sluiten door lamellipodia-gebaseerde kruipen versus portemonnee string sluiting, wat het belang benadrukt van specifieke cel: ECM-interacties bij genezing en van het observeren van cellen in hun natuurlijke context.
Hieronder worden protocollen beschreven voor het maken en afbeelden van eencellige microwounds, kleine wonden die voornamelijk sluiten door celverspreiding en grote wonden die collectieve celmigratie vereisen om te sluiten. Verder wordt een protocol beschreven voor de introductie van kleine moleculen in de ECM- en epitheelcellen, waardoor experimentele verstoringen van vermeende regulerende routes van wondgenezing mogelijk zijn.
1. Dierlijke cultuur
- Clytia-poliepkolonies op microscoopglaasjes en medusae in kunstmatig zeewater (ASW) op 18 °C in een zebravissysteem houden, met 2 L zebravistanks voor poliepkolonies en op maat gemaakte 5 L pseudo-kreiseltanks voor medusae (aanvullende figuur 1)51. ASW bestaat uit 4% Instant Ocean in gedeïoniseerd (DI) H2O.
- Voer de dieren dagelijks met 2-3 dagen oude artemia zoals beschreven51.
OPMERKING: Beeldvorming van wondgenezing is gemakkelijker als de dieren niet onlangs zijn gevoerd, omdat er minder vuil uit de darm in het gezichtsveld vrijkomt. - Verzamel babymedusae van de gevestigde poliepkolonies indien nodig door kolonies 's nachts in een bekerglas van 2 L gevuld met 1 L ASW te plaatsen. Gebruik 2-3 weken oude vrouwelijke medusae voor alle wondgenezingsexperimenten. De vermeerdering van Clytia is elders in detail beschreven51.
2. Verwonding
- Het creëren van microwounds in en tussen cellen (20-500 μm2)
- Maak een aangepaste transferpipet door de punt met een schaar af te snijden om een grotere opening te maken (diameter 0,5-0,7 cm).
OPMERKING: De opening in de pipet moet breed genoeg zijn om schade aan het dier te voorkomen. - Plaats met behulp van de aangepaste transferpipet de medusa op een depressieglaasje met de medusa-exumbrella naar boven gericht, met net genoeg ASW om het dier te bedekken.
- Plaats een coverslip over het dier en afbeelding onmiddellijk (zie hieronder voor beschrijving van de beeldvorming). De coverslip comprimeert de mesoglea en de rebound van het samengeperste weefsel creëert een kracht die de cellen iets uit elkaar duwt52. Dit verschijnt onmiddellijk als openingen tussen elke cel en schade binnen sommige cellen (figuur 1B, B', figuur 2 en figuur 3A-C).
- Maak een aangepaste transferpipet door de punt met een schaar af te snijden om een grotere opening te maken (diameter 0,5-0,7 cm).
- Het creëren van kleine epitheliale wonden (0,02-0,125 mm2)
- Plaats met behulp van een aangepaste transferpipet (zoals hierboven) de medusa op een depressieglaasje met de medusa-exumbrella naar boven gericht.
- Krab met een pipetpunt van 200 μL voorzichtig aan het oppervlak van de medusa. Zacht krabben kan ook scheuren in het keldermembraan veroorzaken, die gemakkelijk zichtbaar zijn22. Bedek het dier met een dekslip voor beeldvorming. Als alternatief is de plaatsing van de dekplaat soms voldoende om kleine epitheelwonden te creëren, zelfs zonder krassen (figuur 1C, C', figuur 2 en figuur 3A-C).
OPMERKING: Druk niet op het oppervlak van de medusa, omdat dit de ECU beschadigt en een onregelmatig oppervlak creëert - epitheelcellen die op een onregelmatig oppervlak migreren, zijn moeilijker scherp te houden.
- Het creëren van grote epitheliale wonden (0,5-0,9 mm2)
- Maak een microinjectienaald met behulp van een micropipettrekker en glazen capillaire buis (stap 5.2). Plaats de lege microinjectienaald in een micro-injectorhouder die aan een micromanipulator is bevestigd. Snijd de punt van de naald zo af dat de opening ongeveer 20-40 μm is.
OPMERKING: Gesneden naalden voor grote epitheelwonden kunnen worden opgeslagen en hergebruikt om de consistentie tussen experimenten te vergroten. - Stel de hold-druk op de micro-injector in op nul en stel de uitwerpdruk in op ongeveer 20 PSI. Stel de micro-injector in om een luchtpuls van 2 s af te geven.
OPMERKING: De uitwerpdruk moet mogelijk worden aangepast op basis van de diameter van de naaldopening (d.w.z. kleinere uiteinden gebruiken een hogere druk, terwijl grotere uiteinden een lagere druk gebruiken). - Plaats de medusa met de exumbrella naar boven gericht op een depressieglaasje op het podium van een ontleedscoop, met net genoeg ASW om het dier te bedekken. Stel met behulp van de micromanipulator de punt van de microinjectienaald zo in dat deze zich net boven het water bevindt. Om dit te doen, dompelt u de punt voorzichtig in water (water kan de pipetpunt binnendringen) en trekt u deze vervolgens in zodat deze zich dicht bij het epitheeloppervlak van de medusa bevindt.
OPMERKING: De punt moet over één kwadrant van de medusa worden geplaatst. De radiale kanalen van de medusa verdelen de medusaklok in vier verschillende kwadranten. Het richten op een kwadrant zal resulteren in schonere beeldvorming, omdat de geslachtsklieren en de radiale kanalen worden uitgesloten van het wondgebied. - Pulseer lucht door op start op de injector te drukken. Herhaal de puls twee tot vier keer op dezelfde plek, afhankelijk van de breedte van de punt. Grotere tips vereisen minder pulsen.
OPMERKING: Een inkeping in het water/medusa veroorzaakt door de puls van lucht moet zichtbaar zijn. - Bedek het gewonde dier met een dekslip voor het in beeld brengen van grote wonden (figuur 1D,D').
- Volg de onderstaande stappen (rubriek 3) voor beeldvorming van epitheliale wondgenezing.
- Maak een microinjectienaald met behulp van een micropipettrekker en glazen capillaire buis (stap 5.2). Plaats de lege microinjectienaald in een micro-injectorhouder die aan een micromanipulator is bevestigd. Snijd de punt van de naald zo af dat de opening ongeveer 20-40 μm is.
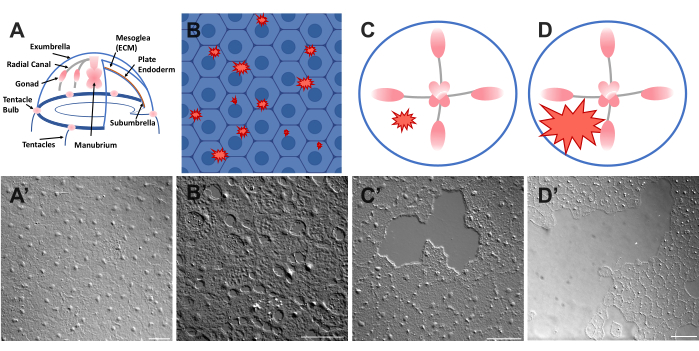
Figuur 1: Intacte en gewonde exparaplu epitheellaag in Clytia medusa. (A) Cartoon afbeelding van de Clytia medusa lichaam. (A') Intact medusa exumbrella epitheel van bovenaf gezien. (B) Cartoon van eencellige microwounds (rode gekartelde vormen) met epitheelcellen in blauw. B') Eencellige microwounds. (C) Cartoon van een kleine epitheliale wond (rode gekartelde vorm). C') Kleine epitheliale wond. (D) Cartoon van een grote epitheliale wond (rode gekartelde vorm). D') Grote epitheliale wond. Beelden werden allemaal verkregen met behulp van DIC-microscopie. Schaalstaven in (A'-C'): 50 μm. Schaalbalk in (D'): 100 μm. Klik hier om een grotere versie van deze figuur te bekijken.
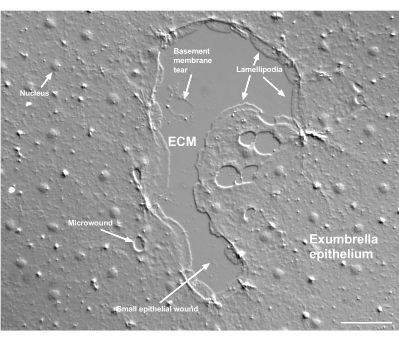
Figuur 2: Meerdere wonden en een beschadigd keldermembraan. Een typische kleine exparalu-epitheelwond wordt getoond, met labels die lamellipodia aangeven die zich vormen uit marginale cellen. Daarnaast worden microwounds in en tussen epitheelcellen gezien. Let op de kleine keldermembraan scheur in het bovenste deel van de wond. Film 4 toont genezing van deze wond. Schaalbalk: 50 μm. Klik hier om een grotere versie van deze figuur te bekijken.
3. Beeldvorming van epitheliale wondgenezing
- Zorg ervoor dat de microscoop is uitgelijnd voor Köhler-verlichting53 en dat deze correct is ingesteld voor dic-microscopie (differentieel interfererend contrast)54. Epitheelcellen zijn bijna onzichtbaar met standaard optica (Figuur 3D,E).
- Pas de focus aan op de exparaplu. Hoewel dit een dunne laag is, moeten zeshoekige cellen helder zijn.
OPMERKING: De exumbrella en subumbrella worden gescheiden door een dikke mesoglea die wordt ondersteund door verticale vezels. De subparaplataire cellen bevinden zich in hetzelfde brandpuntsvlak als de radiale kanalen. Als u in eerste instantie op de subparaplarlaag bent gericht, past u de focus langzaam aan door de mesoglea en verticale vezels totdat u de exparaplu vindt. - Identificeer handmatig een wond om af te beelden. Gebruik voor grote wonden een 10x objectief. Gebruik voor kleinere wonden en eencellige wonden een 20x-objectief.
- Start een programma dat beelden verzamelt als een film in realtime of dat een reeks afbeeldingen verzamelt met regelmatige tussenpozen. Bewaak de voortgang om ervoor te zorgen dat het wondgebied niet uit het gezichtsveld verdwijnt en dat de cellen van belang in focus blijven.
- Eencellige wonden sluiten binnen een minuut; Stel je daarom hun afsluiting voor met een film.
- Om details van de celdynamiek voor kleine wonden vast te leggen, verzamelt u ongeveer elke 10 s afbeeldingen. Het sluiten van kleine wonden duurt 20-50 minuten, afhankelijk van de grootte.
- Stel de niet-afgesloten dia's niet langer dan 45 minuten voor, omdat verdamping van water uit de glijbaan na verloop van tijd leidt tot dierensterfte en scheuring van de cellen.
- Voor langere observatie, sluit rond de coverslip af met vaseline om verdamping te verminderen.
OPMERKING: Sommige medusa kunnen pulseren op de dia, wat de beeldvorming verstoort. In dit geval dient het monteren van dieren in een 1:10 verdunning van 1% ethyl-3-aminobenzoaat methaansulfonaat (Tricaine), aangepast aan pH 7,5, in ASW als een effectief verdovingsmiddel en heeft het geen duidelijk effect op genezing in een tijdsbestek van 1 uur. De dieren zullen echter sterven als ze enkele uren in Tricaine worden achtergelaten.

Figuur 3: Het creëren van een kleine wond in het overkoepelende epitheel. (A) Voorzichtig krabben van de exumbrella met een pipetpunt van 200 μL om een kleine epitheliale wond te creëren. (B) Het plaatsen van de dekplaat is soms voldoende om kleine epitheelwonden te creëren. (C) Medusa gemonteerd op een depressieglijbaan. (D ) Klein epitheliaal wondbeeld zonder DIC-optica en (E) met DIC-optiek. Schaalstaven: 50 μm Klik hier om een grotere versie van deze figuur te bekijken.
4. Analyse
- Afbeeldingsbestanden voorbereiden
OPMERKING: Om de afbeeldingsbestanden te verwerken, gebruikt u FIJI / ImageJ met bijgewerkte BioFormat-plug-ins.- Stel de schaal in op de juiste pixelverhouding per micron voordat u de afbeeldingsstapel registreert; Analyseer > ingestelde schaal. Dit is nodig voor het extraheren van metingen op ware grootte in downstream-analyses.
- Vaak drijft het dier lichtjes op de microscoopglaas; daarom, om drift in films te elimineren, registreert u de afbeeldingen met behulp van de FIJI-plug-in lineaire stackuitlijning met SIFT. Plug-ins > registratie > lineaire stackuitlijning met SIFT.
- Sla de geregistreerde stack op als een .avi bestand. Bestand > opslaan als > AVI ... Stel in de pop-up de framesnelheid in (geanimeerde figuren hierin zijn ingesteld op 10 fps) en klik op OK. Gebruik deze uitvoer om wondgenezingsanalyses uit te voeren.
- Analyse van het wondgebied
- Gebruik het lassogereedschap in FIJI/ImageJ om de wond te schetsen door de celranden te volgen. Meet het wondgebied dat zojuist is omlijnd met Command+M of CTRL+M.
- Herhaal de wondgebiedmeting om de 10 frames. De metingen van FIJI/ImageJ kunnen vervolgens worden uitgezet met Prism 9 (Figuur 4).
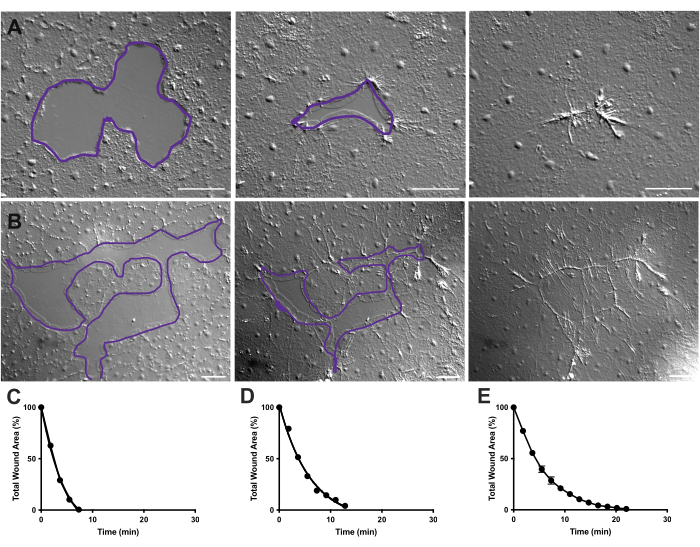
Figuur 4: Analyse van het wondgebied in kleine epitheelwonden. (A ) Voorbeeld van een kleine epitheliale wondgenezing gedurende 10 min. (B) Voorbeeld van een andere epitheliale wondgenezing gedurende 21 min. De paarse contouren in A,B zijn vergelijkbaar met de metingen van wondgebieden met behulp van het lassogereedschap in FIJI/ImageJ. (C) Genormaliseerde vermindering van het wondgebied in de loop van de tijd in A. (D) Genormaliseerde vermindering van het wondgebied in de loop van de tijd in B. (E) Gemiddelde vermindering van het wondgebied in de loop van de tijd voor 14 kleine wonden. n = 14. Foutbalken gecentreerd rond het gemiddelde ± SEM. Schaalbalken: 50 μm Klik hier om een grotere versie van deze figuur te bekijken.
5. Mesogleale injecties
- Injectieschaal maken
- Bereid polydimethylsiloxaan (PDMS) door een PDMS-base en uithardingsmiddel te combineren, in een verhouding van 10 delen base tot 1 deel uithardingsmiddel per gewicht. Roer krachtig om de basis en het uithardingsmiddel volledig te mengen.
- Om bubbels te verwijderen, plaatst u het mengsel gedurende 15 minuten in een vacuümkamer. Giet het mengsel in een petrischaal van 60 mm met microcentrifugebuisdoppen om de mal op zijn plaats te houden. Plaats de mal onmiddellijk op buisdoppen met een helling van 45° en tape op zijn plaats. De mal bestaat uit drie gestapelde, offset glazen dia's die aan elkaar zijn gelijmd om ribbels in de uiteindelijke injectieschaal te creëren.
- Plaats de hele schaal, vorm en het mengsel gedurende 2 uur in een oven op 60 °C om het elastomeer uit te harden. Verwijder de mal voor een voltooide injectieschaal.
- Micropipette trekken
- Ontwerp met behulp van een micro-elektrodetrekker een trekprogramma. Gebruik een programma in één stap met hoge snelheid. De warmte is ongeveer het glazen RAMP testresultaat55,56. Controleer de resulterende micropipetten op lange consistente taps toelopend.
OPMERKING: Gebruik dunwandige glazen borosilicaatcapillairen met een buitendiameter van 1,0 mm, een binnendiameter van 0,75 mm en een lengte van 10 cm.
- Ontwerp met behulp van een micro-elektrodetrekker een trekprogramma. Gebruik een programma in één stap met hoge snelheid. De warmte is ongeveer het glazen RAMP testresultaat55,56. Controleer de resulterende micropipetten op lange consistente taps toelopend.
- Injectie van kleurstoffen en medicijnen
- Maak een microinjectienaald (zoals hierboven).
- Vul de micro-injectienaald met een lange pipetpunt met een overtollig volume kleurstof of medicijn voor injectie in de medusa.
OPMERKING: Voor Clytia moet dimethylsulfoxide (DMSO) worden bewaard op een verdunning van <1:100 met ASW, omdat hogere DMSO-concentraties de wondgenezing belemmeren. Bij het injecteren van een heldere oplossing kan Fast Green FCF-oplossing (1:100 verdunning van 0,1% Fast Green FCF in ASW) worden toegevoegd om de geïnjecteerde vloeistof te visualiseren. - Plaats met behulp van een aangepaste transferpipet zoals hierboven een medusa met de subparaplu naar boven gericht in een PDMS-injectieschaal met net genoeg ASW om het dier te bedekken (figuur 5C). Plaats de schaal op het podium van een ontleedkijker.
OPMERKING: Het beperken van overtollige ASW voorkomt dat de medusa in het gerecht zwemt en maakt meer succesvolle injecties mogelijk. - Focus op de punt van de micro-injectienaald en breng deze in het water in de buurt van de medusa. Druk met de micromanipulator de naald in de schaal totdat deze buigt en breekt. Deze tipopening is ongeveer 10-20 μm.
OPMERKING: Deze naald kan herhaaldelijk worden gebruikt voor dezelfde kleurstof / medicijninjecties die dag. Het wordt aanbevolen om elke dag een nieuwe tip te gebruiken en voor afzonderlijke kleurstoffen / medicijnen. - Steek met behulp van de micromanipulator de punt van de naald door de subparaplu in de mesoglea zonder de paraplu te doorboren.
OPMERKING: Een kreuken/vouwen van het epitheel zal merkbaar zijn. Zodra de naald in de medusa is ingebracht, stopt het kreuken/vouwen. - Stel op de micro-injector de hold-druk in op nul en de uitwerpdruk op ≤20 PSI. Injecteer in een of twee kwadranten en vul elk met een vlek kleurstof of medicijn ongeveer 1/4 van het gebied van dat kwadrant.
OPMERKING: Afhankelijk van de grootte van de medusa zijn grotere of kleinere volumes geschikt op enkele injectieplaatsen. Overvulling van de medusa resulteert in extreme schade aan het epitheel en zelfs de dood van het dier. - Afhankelijk van welke kleurstof of welk medicijn wordt geïnjecteerd, worden dieren in een bekerglas verse ASW geplaatst om kleurstof of medicijndiffusie en incubatie mogelijk te maken.
- Monteer voor beeldvorming de medusa op een depressieglaasje met behulp van een aangepaste transferpipet, waarbij het dier zo wordt geplaatst dat de exumbrella naar boven is gericht (figuur 5). Dieren kunnen in dit stadium gewond raken om het effect van een geïnjecteerd reagens te testen.
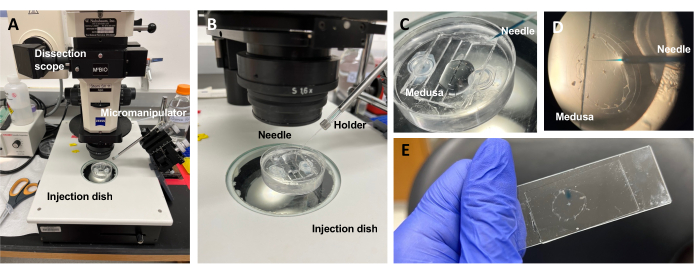
Figuur 5: Injectie-instelling voor het introduceren van kleurstoffen of medicijnen in de ECU. (A) Injectie setup. (B) Close-up van de injectieopstelling met de oriëntatie van de micro-injectienaald (ongeveer 45° hoek ten opzichte van het dier in de schaal). (C) Close-up van de siliconen injectieschaal met de medusa in een kleine hoeveelheid ASW voor injectie. (D) Een microinjectienaald geladen met Fast Green FCF die via de subparaplu de mesoglea van de medusa binnendringt. (E) Na-injectie van Fast Green FCF in een gemonteerde medusa. Klik hier om een grotere versie van deze figuur te bekijken.
Volgens de bovenstaande protocollen werden eencellige microwounds, kleine wonden en grote wonden in beeld gebracht. Geregistreerde stapels afbeeldingsbestanden werden opgeslagen als .avi bestanden.
In film 1 is te zien dat microwounds zich sluiten tussen en in cellen (figuur 1 en figuur 2). Kleine lamellipodia worden waargenomen tijdens de sluiting, gevolgd door samentrekking en genezing. Puin wordt uitgesloten en in het water geloosd. Genezing is voltooid in een minuut of minder.
In film 2 en 3 genezen kleine wonden van verschillende vormen door de vorming van lamellipodia, uitbreiding van lamellipodiale contacten en verspreiding van cellen aan de wondrand, zoals eerder beschreven40 (figuur 1 en figuur 2). Cellen in lagen achter de marginale cellen nemen niet deel aan de genezing van wonden van deze grootte en er is ook geen sprake van collectieve celmigratie. Snelle en progressieve sluiting van epitheliale openingen wordt gevolgd door weefselcontractie langs de nieuw gevormde wondnaad40. De genormaliseerde genezingssnelheid van deze twee wonden, uitgedrukt als een percentage van het oorspronkelijke gebied in de loop van de tijd, wordt weergegeven (figuur 4C, D). Hoewel er enige variabiliteit is in de dynamiek van wondsluiting, maakt het gemiddelde van het percentage gebiedssluiting in de loop van de tijd voor 14 wonden van verschillende vormen, variërend van 0,02-0,125 mm2 , het mogelijk om een gemiddelde curve voor wondgenezing bij onbehandelde dieren vast te stellen (figuur 4E).
Schade aan het keldermembraan is duidelijk te zien wanneer deze optreedt (figuur 2). In film 4 verspreiden cellen aan de rand van een kleine wond waarin zich schade aan het keldermembraan bevindt zich rond het beschadigde gebied en wordt het sluiten van de opening voltooid met een samentrekking van de portemonnee.
Als het weefsel uitgedroogd of te beschadigd is om te herstellen, kunnen celbewegingen stoppen of kan het hele vel cellen barsten (film 5 en film 6). Dit gebeurt meestal na lange perioden van beeldvorming (45 minuten of langer). Als celuitbarstingen vroeg in de beeldvorming optreden, wordt het monster weggegooid.
Zoals te zien is in film 7, genezen grote wonden in verschillende fasen. Ten eerste wordt de rand van de wond glad en regelmatig door samentrekkingen aan de rand, zoals eerder gemeld57. Vervolgens worden lamellipodia gezien om zich te vormen uit de cellen aan de wondrand, waarbij lamellipodia naar voren bewegen om het contact met aangrenzende lamellipodia te maximaliseren. Het volgen van de kernen in cellen aan de wondrand en verschillende lagen achter de marginale cellen laat zien dat grote gaten dicht raken door collectieve celmigratie40. Cellen komen nooit los, maar bewegen samen als een vel.
De introductie van kleurstoffen en farmacologische middelen kan een krachtig hulpmiddel zijn voor het ontleden van biologische mechanismen. Veel stoffen zijn uitgesloten van Clytia (niet getoond), waarschijnlijk vanwege de slijmlaag die het oppervlak van het dier bedekt. Micro-injectie kan echter worden gebruikt om moleculen rechtstreeks in de ECU te introduceren, waardoor de ECM-structuur wordt verstoord of regulerende activiteiten in de ECU worden verstoord. Bovendien kunnen kleurstoffen en andere moleculen epitheelcellen binnendringen vanaf de basale kant. Figuur 6 toont bijvoorbeeld nucleaire kleuring met Hoechst, membraankleuring met FM1-43 en remming van lamellipodiavorming door cytochalasine B nadat deze reagentia in de ECU zijn geïnjecteerd. De introductie van deze moleculen in de ECM en epitheelcellen vóór verwonding maakt experimenten mogelijk die het effect van farmacologische hulpmiddelen op het genezingsproces testen.
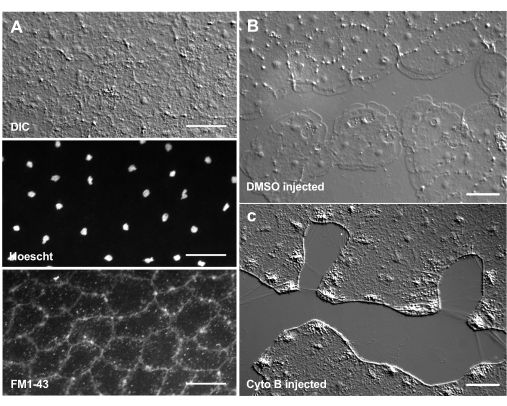
Figuur 6: Epitheelcellen van de medusa na micro-injectie van kleurstoffen of farmacologische middelen. (A) Epitheelcellen getoond in het bovenste paneel 5 min na injectie met 20 μM Hoechst (kernen) en 50 μM FM1-43 (membranen). (B,C) Wondgenezing na injectie met 1:1.000 DMSO controle (B) of 100 μM Cytochalasine B (C). Wonden werden 15 minuten na injectie gemaakt. De foto's werden 5 minuten na de verwonding gemaakt. De vorming van lamellipodia wordt geremd door cytochalasine B. De schijnbare "vezels" die vaak tussen cellen in het wondgebied worden gezien, worden verondersteld het resultaat te zijn van spanning die het keldermembraan uitrekt - ze vlekken niet met falloidine (niet getoond). Schaalstaven: 50 μm. Klik hier om een grotere versie van deze figuur te bekijken.
Film 1: Time-lapse film van eencellige microwound genezing. Verstreken tijd: 20 s. Frame rate: 10 fps. Schaalbalk: 50 μm. Klik hier om deze film te downloaden.
Film 2: Time-lapse film van een kleine epitheliale wondgenezing. Verstreken tijd: 9 min 54 s. Frame rate: 10 fps. Schaalbalk: 50 μm. Klik hier om deze film te downloaden.
Film 3: Time-lapse film van een kleine epitheliale wondgenezing. Deze wond is groter en onregelmatiger gevormd dan de wond in Film 2. Verstreken tijd: 20 min 54 s. Frame rate: 10 fps. Schaalbalk: 50 μm. Klik hier om deze film te downloaden.
Film 4: Time-lapse film van een kleine wond en een microwound genezing met een kelder membraan scheur. Lamellipodia verspreidden zich rond de keldermembraanscheur, hoewel ze over de rest van de ECU kunnen gaan. Zodra het gebied van de wond met de schade aan het keldermembraan is omringd, trekt een samentrekking van de portemonnee cellen over het gebied. Verstreken tijd: 19 min 4 s. Frame rate: 10 fps. Schaalbalk: 50 μm. Klik hier om deze film te downloaden.
Film 5: Cellen die sterven in een kleine epitheelwond. Celdood is waarschijnlijk te wijten aan uitdroging van het dier. Verstreken tijd: 4 min 24 s. Frame rate: 10 fps. Schaalbalk: 100 μm. Klik hier om deze film te downloaden.
Film 6: Een kleine epitheelwond slaagt er niet in om de genezing te voltooien. Verstreken tijd: 42 min 32 s. Frame rate: 10 fps. Schaalbalk: 50 μm. Klik hier om deze film te downloaden.
Film 7: Grote epitheliale wondgenezing. Verstreken tijd: 25 min 29 s. Frame rate: 10 fps. Schaalbalk: 100 μm. Klik hier om deze film te downloaden.
Aanvullende figuur 1: Clytia-tankdimensieschema's. 3D visualisatie van de op maat gemaakte Clytia tanks. (A) Voor- en achteraanzicht. (B) Zijaanzicht. De uitsparing in het groen afgebeelde stuk is bedekt met nylon gaas. Water komt de tank direct over het gaas binnen, veegt over het gaas en creëert een cirkelvormige stroom. Water verlaat het systeem via het gat in het eindstuk dat in blauw is weergegeven. Klik hier om dit bestand te downloaden.
Aanvullende film 1: Acellulaire extracellulaire matrix in Clytia. Z-stack van Clytia genomen met behulp van confocale microscopie. De stack richt zich in eerste instantie op de exumbrella en scant vervolgens elke 10 μm door de ECM naar de plaat endoderm en subparaplu. Afbeeldingen met DIC (links) en Hoechst nucleaire kleuring (rechts) tonen het gebrek aan cellen in de ECM. Schaalbalk: 100 μm. Klik hier om dit bestand te downloaden.
Hier wordt de methodologie gepresenteerd voor het in vivo afbeelden van wonden in Clytia, een relatief nieuw ongewerveld modelorganisme40,43,58. Er zijn verschillende factoren die dit systeem tot een uniek en krachtig onderzoeksinstrument maken, anders dan andere modellen die worden gebruikt om wondgenezing en re-epithelisatie te bestuderen. Ten eerste wordt het monolaagsepitheel bevestigd aan een transparante ECM, waardoor het lijkt op in vitro weefselkweektests (figuur 1, figuur 2, figuur 3, figuur 4). Net als in vitro assays kunnen cellen met hoge resolutie in beeld worden gebracht. In tegenstelling tot bij weefselkweek is er echter een authentieke cellulaire omgeving en ECM, zodat wondgenezing kan worden gezien in de context van de complexe signaleringsgebeurtenissen die optreden bij een levend gewond dier. Ten tweede mist Clytia ontstekingsreacties, migrerende fibroblasten, vasculatuur en bloed. Hierdoor kan het re-epithelisatieproces in vivo worden bestudeerd in afwezigheid van de overlappende gebeurtenissen die optreden bij complexere volwassen dieren tijdens wondgenezing59. Ten derde is de ECU acellulair (aanvullende film 1) en groot, waardoor deze gemakkelijk toegankelijk is met een micro-injectienaald (figuur 5 en figuur 6). Met behulp van deze aanpak kunnen onderzoekers het effect testen van farmacologische reagentia die de ECM-structuur verstoren of signaleren op wondgenezing in vivo. Reagentia kunnen ook in epitheelcellen worden ingebracht en hun effecten op in vivo wondgenezing kunnen worden beoordeeld. Ten vierde zijn er protocollen die bestaan voor het creëren van mutanten en transgene dieren in het Clytia-systeem42,43,44,45. In vivo wondgenezing kan daarom worden waargenomen bij dieren met een verhoogde/verminderde expressie van genen van belang.
Er zijn verschillende kritieke stappen in deze techniek. Ten eerste, zoals weergegeven in figuur 3, is het noodzakelijk om een microscoop te gebruiken die correct is geconfigureerd voor DIC-microscopie, omdat de platte, transparante epitheelcellen bijna onzichtbaar zijn met standaard lichtmicroscopie. Het is ook belangrijk om de vaardigheid te ontwikkelen om dieren voorzichtig te verwonden, zodat het epitheel wordt beschadigd zonder de ECU te gutsen. Een vergelijkbare zachte aanraking is nodig voor het micro-injecteren van materialen in de ECU, omdat uitgebreide schade aan het dier tijdens de injectie een volgende analyse van wondgenezing in gevaar kan brengen. Hoewel er een leercurve is voor deze technieken, hebben zelfs beginnende studenten ze snel onder de knie in het Malamy-lab. Inderdaad, deze protocollen zijn gebruikt om celmigratie aan te tonen in niet-gegradueerde laboratoriumcursussen aan de Universiteit van Chicago.
Voor een optimale beeldvorming is het belangrijk dat het dier niet beweegt en het gekozen wondgebied niet uit het gezichtsveld verdwijnt. Als dieren pulseren, is de behandeling met Tricaïne zoals beschreven zeer effectief. Voor driften is het vaak nodig om het monster handmatig te verplaatsen. Deze bewegingen kunnen uit de uiteindelijke film worden verwijderd met behulp van de registratiefunctie in FIJI / ImageJ.
Een beperking met dit systeem is dat het niet mogelijk is om identieke wonden te maken, omdat wonden variëren in zowel vorm als grootte met behulp van de hier beschreven methoden. Daarom kan het moeilijk zijn om de exacte snelheid van wondsluiting of celmigratie te kwantificeren. Positionele markers zoals koolstofkorrels kleven aan de blootgestelde ECM in een gewond dier en kunnen worden gebruikt om de snelheid van collectieve celmigratie in grote wonden te meten (niet weergegeven). Voor analyse van kleine wondsluitingen, zelfs met variabele wondgrootte en -vorm, is er een beperkt bereik van sluitingspercentages bij wonden van deze grootte (figuur 4). Het is daarom mogelijk om de effecten van promotive of repressieve farmacologische reagentia kwantitatief te detecteren.
Hoewel dit werk de karakterisering van wondgenezing beschrijft met behulp van alleen DIC-microscopie, kunnen dezelfde benaderingen worden gebruikt om genezing in beeld te brengen met behulp van fluorescentie of confocale microscopie. Om hierbij te helpen, zijn er protocollen om transgene dieren te genereren waarin verschillende cellulaire en extracellulaire eiwitten fluorescerend worden gelabeld. Gelijktijdige beeldvorming met DIC en fluorescentie, gecombineerd met verstoring van wondgenezing met behulp van farmacologische middelen of mutante lijnen, zal een krachtige benadering zijn om mechanismen te begrijpen die ten grondslag liggen aan het wondgenezingsproces in het epitheel.
Niets te onthullen.
E.E.L.L. wordt ondersteund door een subsidie van de National Science Foundation PRFB 2011010. We willen Tsuyoshi Momose en Evelyn Houliston bedanken voor hun hulp bij het opzetten van onze Clytia-kolonies, Jean-Baptiste Reynier voor het verzamelen van de microwound-genezingsbeelden, Harry Kyriazes voor de bouw van de pseudo-kreiseltanks en Elizabeth Baldo voor het onderhoud van de Clytia-habitat. Figuur 1B is gemaakt met BioRender.com.
| Name | Company | Catalog Number | Comments |
| 20500 ACE EKE Microscope Fiber Optic Light Source | Kramer Scientific Corporation | ||
| AxioCam 506 mono | ZEISS | 426557-0000-000-MA285 | |
| Capillary tubes | World Precision Instruments | TW1004 | |
| Cytochalasin B | Abcam | ab143482 | |
| Depression slides | Amscope | BS-C12 | |
| DMR with DIC options and fluorescence halogen lamp | Leica | ||
| Ethyl 3-aminobenzoate methanesulfonate | Sigma Aldrich | E10521-10G | |
| Fast Green FCF | Thermo Scientific | A16520-06 | |
| FM1-43 | Biotium | 70022 | Excitation/Emission: 480/598 nm |
| Hoechst 33342 | Thermo Scientific | 62249 | Excitation/Emission: 361/497 nm |
| imageJ | NIH | ||
| Microloader tips (0.5-10 μL /2-20 μL) | Eppendorf | 930001007 | |
| Micromanipulator | World Precision Instruments | 3301R / M3301L | |
| Microscope Cover Glass (22X40-1.5) | Fisherbrand | 12-544-BP | |
| Petri Dish (60 mm x 15 mm) | Fisherbrand | FB085713A | |
| PicoNozzle v2 | World Precision Instruments | 5430-ALL | |
| Pipette puller | Sutter Instrument Co | P-97 | |
| Pneumatic PicoPump | World Precision Instruments | PV820 | |
| Polycarbonate vacuum, desiccator | Bel-art | F42025-0000 | |
| Prism 9 | GraphPad | ||
| STEMI Sv11 Dissection scope | ZEISS | STEMI SV11 | |
| SYLGARD 184 | Dow Silicones | 1024001 | |
| Transfer pipettes | Fisherbrand | 13-711-7M | |
| Z-Hab mini system | Pentair | ||
| ZEN Microscopy software | Zeiss |
- Tyler, S. Epithelium-the primary building block for metazoan complexity. Integrative and Comparative Biology. 43 (1), 55-63 (2003).
- Kurn, H., Daly, D. T. Histology, Epithelial Cell. StatPearls. , (2022).
- Schempp, C., Emde, M., Wölfle, U. Dermatology in the Darwin anniversary. Part 1: Evolution of the integument. Journal of the German Society of Dermatology. 7 (9), 750-757 (2009).
- Lopez-Ojeda, W., Pandey, A., Alhajj, M., Oakley, A. M. Anatomy, Skin (Integument). StatPearls. , (2022).
- Bukowiecki, A., Hos, D., Cursiefen, C., Eming, S. A. Wound-healing studies in cornea and skin: parallels, differences and opportunities. International Journal of Molecular Sciences. 18 (6), 1257 (2017).
- Frantz, C., Stewart, K. M., Weaver, V. M. The extracellular matrix at a glance. Journal of Cell Science. 123 (24), 4195-4200 (2010).
- Hynes, R. O. The evolution of metazoan extracellular matrix. The Journal of Cell Biology. 196 (6), 671-679 (2012).
- Fidler, A. L., et al. Collagen IV and basement membrane at the evolutionary dawn of metazoan tissues. eLife. 6, 24176 (2017).
- Bullen, T. F., et al. Characterization of epithelial cell shedding from human small intestine. Laboratory Investigation; a Journal of Technical Methods and Pathology. 86 (10), 1052-1063 (2006).
- Watson, A. J. M., et al. Epithelial barrier function in vivo is sustained despite gaps in epithelial layers. Gastroenterology. 129 (3), 902-912 (2005).
- Sonnemann, K. J., Bement, W. M. Wound repair: toward understanding and integration of single-cell and multicellular wound responses. Annual Review of Cell and Developmental Biology. 27, 237-263 (2011).
- Abreu-Blanco, M. T., Verboon, J. M., Parkhurst, S. M. Single cell wound repair. BioArchitecture. 1 (3), 114-121 (2011).
- Fenteany, G., Janmey, P. A., Stossel, T. P. Signaling pathways and cell mechanics involved in wound closure by epithelial cell sheets. Current Biology. 10 (14), 831-838 (2000).
- Pastar, I., et al. Epithelialization in wound healing: a comprehensive review. Advances in Wound Care. 3 (7), 445-464 (2014).
- Rousselle, P., Braye, F., Dayan, G. Re-epithelialization of adult skin wounds: Cellular mechanisms and therapeutic strategies. Advanced Drug Delivery Reviews. 146, 344-365 (2019).
- Bement, W. M., Forscher, P., Mooseker, M. S. A novel cytoskeletal structure involved in purse string wound closure and cell polarity maintenance. The Journal of Cell Biology. 121 (3), 565-578 (1993).
- Vedula, S. R. K., Ravasio, A., Lim, C. T., Ladoux, B. Collective Cell migration: a mechanistic perspective. Physiology. 28 (6), 370-379 (2013).
- Li, L., He, Y., Zhao, M., Jiang, J. Collective cell migration: Implications for wound healing and cancer invasion. Burns & Trauma. 1 (1), 21-26 (2015).
- Bornes, L., Windoffer, R., Leube, R. E., Morgner, J., van Rheenen, J. Scratch-induced partial skin wounds re-epithelialize by sheets of independently migrating keratinocytes. Life Science Alliance. 4 (1), 202000765 (2021).
- Theveneau, E., Mayor, R. Collective cell migration of epithelial and mesenchymal cells. Cellular and Molecular Life Sciences. 70 (19), 3481-3492 (2013).
- Haensel, D., Dai, X. Epithelial-to-mesenchymal transition in cutaneous wound healing: where we are and where we are heading. Developmental Dynamics. 247 (3), 473-480 (2018).
- Cordeiro, J. V., Jacinto, A. The role of transcription-independent damage signals in the initiation of epithelial wound healing. Nature Reviews. Molecular Cell Biology. 14 (4), 249-262 (2013).
- Abreu-Blanco, M. T., Watts, J. J., Verboon, J. M., Parkhurst, S. M. Cytoskeleton responses in wound repair. Cellular and Molecular Life Sciences. 69 (15), 2469-2483 (2012).
- Klarlund, J. K., Block, E. R. Free edges in epithelia as cues for motility. Cell Adhesion & Migration. 5 (2), 106-110 (2011).
- Enyedi, B., Niethammer, P. Mechanisms of epithelial wound detection. Trends in Cell Biology. 25 (7), 398-407 (2015).
- Niethammer, P. The early wound signals. Current Opinion in Genetics & Development. 40, 17-22 (2016).
- Jacinto, A., Martinez-Arias, A., Martin, P. Mechanisms of epithelial fusion and repair. Nature Cell Biology. 3 (5), 117-123 (2001).
- Mayor, R., Etienne-Manneville, S. The front and rear of collective cell migration. Nature reviews. Molecular Cell Biology. 17 (2), 97-109 (2016).
- Gupta, S., Yap, A. S. How adherens junctions move cells during collective migration. Faculty Reviews. 10, 56 (2021).
- Blair, M. J., Jones, J. D., Woessner, A. E., Quinn, K. P. Skin structure-function relationships and the wound healing response to intrinsic aging. Advances in Wound Care. 9 (3), 127-143 (2020).
- Falanga, V., et al. Chronic wounds. Nature Reviews. Disease Primers. 8 (1), 50 (2022).
- Liang, C. -. C., Park, A. Y., Guan, J. -. L. In vitro scratch assay: a convenient and inexpensive method for analysis of cell migration in vitro. Nature Protocols. 2 (2), 329-333 (2007).
- Jonkman, J. E. N., et al. An introduction to the wound healing assay using live-cell microscopy. Cell Adhesion & Migration. 8 (5), 440-451 (2014).
- Poujade, M., et al. Collective migration of an epithelial monolayer in response to a model wound. Proceedings of the National Academy of Sciences. 104 (41), 15988-15993 (2007).
- Masson-Meyers, D. S., et al. Experimental models and methods for cutaneous wound healing assessment. International Journal of Experimental Pathology. 101 (1-2), 21-37 (2020).
- Park, S., et al. Tissue-scale coordination of cellular behaviour promotes epidermal wound repair in live mice. Nature Cell Biology. 19 (2), 155-163 (2017).
- Tsai, C. -. R., Wang, Y., Galko, M. J. Crawling wounded: molecular genetic insights into wound healing from Drosophila larvae. The International Journal of Developmental Biology. 62 (6-7-8), 479-489 (2018).
- Richardson, R., et al. Adult zebrafish as a model system for cutaneous wound-healing research. The Journal of Investigative Dermatology. 133 (6), 1655-1665 (2013).
- Erickson, J. R., Echeverri, K. Learning from regeneration research organisms: The circuitous road to scar free wound healing. Developmental Biology. 433 (2), 144-154 (2018).
- Kamran, Z., et al. In vivo imaging of epithelial wound healing in the cnidarian Clytia hemisphaerica demonstrates early evolution of purse string and cell crawling closure mechanisms. BMC Developmental Biology. 17 (1), 17 (2017).
- Chari, T., et al. Whole-animal multiplexed single-cell RNA-seq reveals transcriptional shifts across Clytia medusa cell types. Science Advances. 7 (48), (2021).
- Weissbourd, B., et al. A genetically tractable jellyfish model for systems and evolutionary neuroscience. Cell. 184 (24), 5854-5868 (2021).
- Momose, T., et al. High doses of CRISPR/Cas9 ribonucleoprotein efficiently induce gene knockout with low mosaicism in the hydrozoan Clytia hemisphaerica through microhomology-mediated deletion. Scientific Reports. 8 (1), 11734 (2018).
- Houliston, E., Leclère, L., Munro, C., Copley, R. R., Momose, T. Past, present and future of Clytia hemisphaerica as a laboratory jellyfish. Current Topics in Developmental Biology. 147, 121-151 (2022).
- Schmid, V., et al. The extracellular matrix (mesoglea) of hydrozoan jellyfish and its ability to support cell adhesion and spreading. Hydrobiologia. 216 (1), 3-10 (1991).
- Day, R. M., Lenhoff, H. M. Hydra mesoglea: a model for investigating epithelial cell-basement membrane interactions. Science. 211 (4479), 291-294 (1981).
- Zhang, X., et al. The collagens of hydra provide insight into the evolution of metazoan extracellular matrices. The Journal of Biological Chemistry. 282 (9), 6792-6802 (2007).
- Danjo, Y., Gipson, I. K. Actin 'purse string' filaments are anchored by E-cadherin-mediated adherens junctions at the leading edge of the epithelial wound, providing coordinated cell movement. Journal of Cell Science. 111 (22), 3323-3332 (1998).
- Arenas Gómez, C. M., Sabin, K. Z., Echeverri, K. Wound healing across the animal kingdom: Crosstalk between the immune system and the extracellular matrix. Developmental Dynamics. 249 (7), 834-846 (2020).
- Lechable, M., et al. An improved whole life cycle culture protocol for the hydrozoan genetic model Clytia hemisphaerica. Biology Open. 9 (11), (2020).
- Casares, L., et al. Hydraulic fracture during epithelial stretching. Nature Materials. 14 (3), 343-351 (2015).
- Wayne, R. Chapter 4 - Bright-Field Microscopy. Light and Video Microscopy (Third Edition). , 95-116 (2019).
- Murphy, D. B., Davidson, M. W. . Fundamentals of Light Microscopy and Electronic Imaging: Second Edition. , (2012).
- . Micropipette Techniques for Electrophysiology Available from: https://www.sutter.com/micropipette/cookbook.html (2022)
- Brown, A. L., Johnson, B. E., Goodman, M. B. Making patch-pipettes and sharp electrodes with a programmable puller). Journal of Visualized Experiments. (20), e939 (2008).
- Klarlund, J. K. Dual modes of motility at the leading edge of migrating epithelial cell sheets. Proceedings of the National Academy of Sciences. 109 (39), 15799-15804 (2012).
- Houliston, E., Momose, T., Manuel, M. Clytia hemisphaerica: a jellyfish cousin joins the laboratory. Trends in Genetics. 26 (4), 159-167 (2010).
- Rodrigues, M., Kosaric, N., Bonham, C. A., Gurtner, G. C. Wound healing: a cellular perspective. Physiological Reviews. 99 (1), 665-706 (2019).
ABOUT JoVE
Copyright © 2024 MyJoVE Corporation. All rights reserved