
Summary
Abstract
Introduction
Protocol
Representative Results
Discussion
Acknowledgements
Materials
References
Biology
Caratterizzazione della guarigione delle ferite epiteliali in vivo utilizzando l'organismo modello cnidariano Clytia hemisphaerica
Questo articolo descrive un metodo per creare ferite nell'epitelio di una Clytia hemisphaerica medusa viva e immagini di guarigione delle ferite ad alta risoluzione in vivo. Inoltre, viene presentata una tecnica per introdurre coloranti e farmaci per perturbare i processi di segnalazione nelle cellule epiteliali e nella matrice extracellulare durante la guarigione delle ferite.
Tutti gli organi animali, dalla pelle agli occhi all'intestino, sono ricoperti da fogli di cellule epiteliali che consentono loro di mantenere l'omeostasi proteggendoli dalle infezioni. Pertanto, non sorprende che la capacità di riparare le ferite epiteliali sia fondamentale per tutti i metazoi. La guarigione delle ferite epiteliali nei vertebrati comporta processi sovrapposti, tra cui risposte infiammatorie, vascolarizzazione e riepitelizzazione. La regolazione di questi processi comporta complesse interazioni tra cellule epiteliali, cellule vicine e matrice extracellulare (ECM); l'ECM contiene proteine strutturali, proteine regolatrici e piccole molecole attive. Questa complessità, insieme al fatto che la maggior parte degli animali ha tessuti opachi e ECM inaccessibili, rende difficile studiare la guarigione delle ferite negli animali vivi. Gran parte del lavoro sulla guarigione delle ferite epiteliali viene quindi eseguito nei sistemi di coltura tissutale, con un singolo tipo di cellula epiteliale placcato come monostrato su una matrice artificiale. Clytia hemisphaerica (Clytia) fornisce un complemento unico ed emozionante a questi studi, consentendo di studiare la guarigione delle ferite epiteliali in un animale intatto con un'autentica ECM. L'epitelio ectodermico di Clytia è un singolo strato di grandi cellule epiteliali squamose, che consente l'imaging ad alta risoluzione utilizzando la microscopia a contrasto differenziale interferente (DIC) negli animali vivi. L'assenza di fibroblasti migratori, risposte vascolarizzanti o infiammatorie consente di sezionare gli eventi critici nella riepitelizzazione in vivo. È possibile analizzare la guarigione di vari tipi di ferite, tra cui microferite unicellulari, ferite epiteliali piccole e grandi e ferite che danneggiano la membrana basale. La formazione di lamellipodi, la contrazione dei cordoni della borsa, l'allungamento cellulare e la migrazione cellulare collettiva possono essere osservati in questo sistema. Inoltre, agenti farmacologici possono essere introdotti attraverso la ECM per modificare le interazioni cellula:ECM e i processi cellulari in vivo. Questo lavoro mostra metodi per creare ferite in Clytia vivo, catturare filmati di guarigione e sondare i meccanismi di guarigione microiniettando reagenti nell'ECM.
Fogli di cellule epiteliali coprono la superficie esterna di tutti i metazoi, rivestono gli organi interni e dividono il corpo animale in compartimenti discreti. L'epitelio separa anche il corpo interno dall'ambiente esterno e lo protegge da danni e infezioni. Quindi, l'avvento degli strati epiteliali è stato una parte essenziale dell'evoluzione degli animali multicellulari e gli strati epiteliali sono visti in tutti gli animali, dai vertebrati aimetazoi più basali1. L'epitelio di alcuni organi è un singolo monostrato, come nelle sacche d'aria polmonari, nei vasi sanguigni e nell'intestino2, così come nell'epidermide di invertebrati come planaria e cnidari3. In altri tessuti, come la pelle4 e la cornea5 dei vertebrati, l'epitelio è stratificato, il che significa che ci sono più strati di cellule epiteliali2. In tutti i casi, lo strato epiteliale più basale è apposto sulla membrana basale, un foglio proteico che forma una regione specializzata della matrice extracellulare (ECM)6,7,8.
Le brecce nell'epitelio devono essere rapidamente riparate per ricreare un foglio epiteliale continuo. Il danno all'epitelio si verifica durante i processi naturali, come lo spargimento di cellule epiteliali nell'intestino,9,10 e come risultato di infiammazione o trauma fisico. Quando una singola cellula epiteliale è danneggiata, deve ripararsi o essere eliminata per consentire alle cellule circostanti di attaccarsi l'una all'altra e chiudere il foro11,12. Nelle ferite più grandi delle dimensioni di una singola cellula, le cellule epiteliali devono muoversi per raggiungersi e riparare il foglio13. Ciò può essere ottenuto diffondendo le cellule se le lacune sono piccole o può richiedere la migrazione delle cellule epiteliali dai margini di una ferita per chiudere la lacuna della ferita; Quest'ultimo processo è chiamato riepitelizzazione14,15. Nei tessuti embrionali, le cellule epiteliali si diffondono e migrano per chiudere le ferite o sono tirate attraverso lo spazio dalla contrazione dei cavi di actomiosina che si formano tra le cellule al margine della ferita, in un meccanismo simile a un cordone della borsa16. In molti tessuti adulti, la riepitelizzazione comporta la migrazione di fogli cellulari coerenti, dove le cellule mantengono le loro giunzioni con le cellule vicine14,17,18. In altri tessuti, le connessioni cellula:cellula vengono smantellate e le cellule epiteliali si comportano più come cellule mesenchimali, muovendosi in modo coordinato ma indipendente nella regione della ferita durante la riepitelizzazione 14,19,20,21.
I movimenti delle cellule epiteliali sono regolati da complesse interazioni tra le cellule in migrazione e tra le cellule e la MEC. Mentre c'è un'enorme quantità di letteratura sperimentale che affronta i meccanismi di attivazione delle ferite delle cellule epiteliali e la successiva migrazione, molto resta ancora da scoprire. Ad esempio, il segnale iniziale che attiva le cellule epiteliali a migrare in risposta a una ferita non è stato definitivamente identificato 22, né è completamente compreso come l'actina venga ridistribuita per creare lamellipodi sul lato delle cellule epiteliali più vicine alla ferita 22,23,24,25,26,27. La migrazione collettiva delle cellule richiede che le informazioni provenienti dalle cellule della ferita siano condivise con le cellule distali della ferita e la via di comunicazione non è ancora chiara28. Le giunzioni cella:cella e gli attacchi cell:ECM devono essere smontati e riformati man mano che le celle del foglio si riorganizzano, ma la regolazione di questo processo è poco compresa14,29. Fare progressi su queste e altre domande correlate non è importante solo come problema biologico fondamentale, ma anche per il significato clinico della corretta guarigione delle ferite. Le malattie che compromettono la capacità delle cellule epiteliali di migrare correttamente provocano ferite croniche; un esempio è la malattia genetica epidermolisi bollosa, dove i geni coinvolti nell'attaccamento delle cellule epiteliali alla ECM sono mutati, con conseguente pelle fragile che si squama e vesciche. La riepitelizzazione è compromessa anche nei tessuti naturalmente invecchiati30,31. Una migliore comprensione è quindi essenziale per lo sviluppo di interventi volti a migliorare i risultati di guarigione delle ferite.
La migrazione delle cellule epiteliali nella guarigione delle ferite è stata studiata utilizzando sia approcci in vitro che organismi modello. La maggior parte degli studi sulla guarigione delle ferite e sui meccanismi di migrazione cellulare sono stati condotti in coltura tissutale, dove monostrati di un singolo tipo di cellula epiteliale vengono coltivati su un substrato che sostituisce la ECM. I monostrati cellulari vengono graffiati o cresciuti con stencil per creare spazi vuoti di forme e dimensioni specifiche e quindi osservati32,33,34. Il modello in vitro consente una visualizzazione ideale del comportamento cellulare, nonché l'opportunità di modificare le qualità del substrato, di esporre le cellule a farmaci e fattori abiotici e biotici e di trasfettare le cellule con costrutti che esprimono o sopprimono vari geni di interesse. Tuttavia, questo approccio riduzionista potrebbe non riuscire a catturare alcuni dei parametri importanti coinvolti nel comportamento delle cellule epiteliali in un contesto in vivo, compresa la comunicazione tra vari tipi di cellule e gli eventi di segnalazione che si verificano nell'ECM11. I modelli in vivo forniscono il contesto autentico di una ferita, con più tipi di cellule, vie di segnalazione sovrapposte e una complessa ECM35. Uno di questi modelli per gli studi di guarigione delle ferite è il topo19, in cui i recenti progressi hanno permesso ai ricercatori di osservare le cellule epidermiche durante la guarigione di ferite a tutto spessore negli animali vivi36. Tuttavia, il topo e altri sistemi in vivo presentano sfide per studiare la riepitelizzazione. In primo luogo, il grande vantaggio di osservare il comportamento cellulare in un contesto naturale è bilanciato dalla complessità degli eventi temporalmente sovrapposti che si verificano durante la guarigione delle ferite dei vertebrati, tra cui la coagulazione del sangue, il reclutamento di cellule immunitarie e infiammazione, il reclutamento di fibroblasti e la dedifferenziazione cellulare, la rivascolarizzazione e il rimodellamento della ECM. Inoltre, i tessuti opachi rendono difficile l'imaging. I sistemi di larva di Drosophila e epidermide di pesce zebra 37,38 hanno superato alcune di queste difficoltà a causa della loro relativa semplicità39.
Il nostro laboratorio ha recentemente introdotto un nuovo modello per lo studio della guarigione delle ferite epiteliali: la forma medusa (medusa) dell'idrozoo cnidario Clytia hemisphaerica (Clytia)40. Clytia è un organismo modello emergente con un genoma41 completamente sequenziato e annotato, un trascrittoma42 dell'RNAseq a singola cellula e protocolli in atto per la modifica del genoma (mutagenesi e transgenesi)43,44,45. Gli Cnidari sono uno dei più antichi lignaggi esistenti ad avere strati epiteliali, quindi la comprensione della guarigione delle ferite cnidaria fornisce informazioni sui percorsi ancestrali che hanno assicurato l'integrità epiteliale. Per quei percorsi che sono stati conservati in tutto l'albero della vita, Clytia offre un nuovo entusiasmante sistema per studiare la dinamica delle cellule epiteliali e la regolazione funzionale della guarigione delle ferite in vivo.
L'epitelio che copre la superficie superiore della Clytia medusa (exumbrella) è un monostrato di cellule epiteliali squamose trasparenti larghe circa 50 μm per 1-2 μm di spessore (Figura 1). Sono attaccati a un ECM chiamato mesoglea - la "gelatina" della medusa. La mesoglea è simile dal punto di vista compositivo alla ECM riscontrata in altri animali 46,47,48 compresi i vertebrati, ha una membrana basale 40 ed è completamente trasparente. Lo strato epiteliale nella Clytia medusa può essere facilmente graffiato o ferito (vedi sotto). La semplicità e la trasparenza dell'epitelio e della MEC consente l'imaging ad alta risoluzione delle cellule e dei loro movimenti durante la guarigione. Recentemente, Kamran et al. hanno caratterizzato la guarigione di piccole ferite nell'epitelio di Clytia in dettaglio40. È stato dimostrato che la guarigione in Clytia avviene attraverso il crawling cellulare basato sui lamellipodi, la diffusione cellulare e la migrazione cellulare collettiva, così come la chiusura del cordone della borsa che è più tipica dei sistemi embrionali (sebbene osservata in precedenza in strutture animali adulte come la cornea49). La guarigione delle ferite di Clytia è estremamente veloce, come è stato visto in altri sistemi che mancano di una risposta infiammatoria40,50. La guarigione nell'ombrello Clytia dipende completamente dai movimenti delle cellule epiteliali esistenti - nessuna cellula prolifera o migra attraverso la ECM verso il sito della ferita (Filmato supplementare 1). Tutti questi risultati suggeriscono che Clytia è un sistema modello utile per studiare la guarigione delle ferite epiteliali. In effetti, la facilità di imaging delle cellule epiteliali in Clytia durante la guarigione delle ferite ha portato alla scoperta che i lamellipodi delle cellule epiteliali si estendono e si estendono su aree di ECM esposte finché c'è una membrana basale intatta; Se la membrana basale è danneggiata, la guarigione epiteliale passa a un meccanismo a corda della borsa40. Questa è stata la prima dimostrazione di un meccanismo alla base della decisione di chiudere strisciando a base di lamellipodi rispetto alla chiusura dei cordoni della borsa, evidenziando l'importanza delle specifiche interazioni cellula:ECM nella guarigione e nell'osservazione delle cellule nel loro contesto naturale.
Di seguito, vengono descritti i protocolli per la creazione e l'imaging di microferite a singola cellula, piccole ferite che si chiudono principalmente per diffusione cellulare e ferite di grandi dimensioni che richiedono la migrazione cellulare collettiva per chiudersi. Inoltre, viene descritto un protocollo per l'introduzione di piccole molecole nella ECM e nelle cellule epiteliali, consentendo perturbazioni sperimentali di presunte vie regolatorie di guarigione delle ferite.
1. Cultura animale
- Mantenere le colonie di polipi di Clytia su vetrini da microscopio e meduse in acqua di mare artificiale (ASW) a 18 °C in un sistema di zebrafish, con vasche da 2 litri di zebrafish per colonie di polipi e vasche pseudo-kreisel da 5 litri su misura per meduse (Figura supplementare 1)51. ASW è costituito da 4% Instant Ocean in deionizzato (DI) H2O.
- Dai da mangiare agli animali ogni giorno con artemia di 2-3 giorni come descritto51.
NOTA: L'imaging per la guarigione delle ferite è più facile se gli animali non sono stati nutriti di recente, poiché vi è meno detriti rilasciati dall'intestino nel campo visivo. - Raccogliere le meduse dalle colonie di polipi stabilite, se necessario, posizionando le colonie in un becher da 2 litri riempito con 1 litro di ASW durante la notte. Usa meduse femminili di 2-3 settimane per tutti gli esperimenti di guarigione delle ferite. La propagazione della Clytia è stata descritta in dettaglio altrove51.
2. Ferimento
- Creazione di microferite all'interno e tra le cellule (20-500 μm2)
- Creare una pipetta di trasferimento modificata tagliando la punta con le forbici per creare un'apertura più grande (0,5-0,7 cm di diametro).
NOTA: l'apertura della pipetta deve essere sufficientemente ampia da evitare danni all'animale. - Utilizzando la pipetta di trasferimento modificata, posizionare la medusa su un vetrino di depressione con l'ombrello di medusa rivolto verso l'alto, con ASW sufficiente a coprire l'animale.
- Posizionare immediatamente un copricostume sull'animale e sull'immagine (vedere sotto per la descrizione dell'imaging). Il coprislip comprime la mesoglea e il rimbalzo del tessuto compresso crea una forza che spinge le cellule leggermente divaricate52. Questo appare immediatamente come spazi vuoti tra ogni cella e danni all'interno di alcune celle (Figura 1B, B', Figura 2 e Figura 3A-C).
- Creare una pipetta di trasferimento modificata tagliando la punta con le forbici per creare un'apertura più grande (0,5-0,7 cm di diametro).
- Creazione di piccole ferite epiteliali (0,02-0,125 mm2)
- Utilizzando una pipetta di trasferimento modificata (come sopra), posizionare la medusa su un vetrino di depressione con l'ombrello medusa rivolto verso l'alto.
- Utilizzando una punta per pipetta da 200 μL, grattare delicatamente la superficie della medusa. Graffi delicati possono anche creare strappi nella membrana basale, che sono facilmente evidenti22. Coprire l'animale con un copricostume per l'imaging. In alternativa, il posizionamento del coprislip è talvolta sufficiente per creare piccole ferite epiteliali anche senza graffiare (Figura 1C, C', Figura 2 e Figura 3A-C).
NOTA: Non premere verso il basso quando si graffia la superficie della medusa, poiché ciò danneggia la ECM e crea una superficie irregolare - le cellule epiteliali che migrano su una superficie irregolare sono più difficili da mantenere a fuoco.
- Creazione di grandi ferite epiteliali (0,5-0,9 mm2)
- Fare un ago per microiniezione usando un estrattore di micropipette e un tubo capillare di vetro (punto 5.2). Inserire l'ago di microiniezione vuoto in un supporto per microiniettore fissato a un micromanipolatore. Tagliare la punta dell'ago in modo che l'apertura sia di circa 20-40 μm.
NOTA: Gli aghi tagliati per ferite epiteliali di grandi dimensioni possono essere conservati e riutilizzati per aumentare la coerenza tra gli esperimenti. - Impostare la pressione di mantenimento sul microiniettore su zero e impostare la pressione di espulsione su circa 20 PSI. Impostare il microiniettore per erogare un impulso d'aria di 2 s.
NOTA: potrebbe essere necessario regolare la pressione di espulsione in base al diametro dell'apertura dell'ago (ad esempio, le punte più piccole utilizzeranno una pressione più elevata, mentre le punte più grandi utilizzeranno una pressione inferiore). - Posiziona la medusa con l'ombrello rivolto verso l'alto su uno scivolo di depressione sul palco di un cannocchiale da dissezione, con ASW appena sufficiente a coprire l'animale. Utilizzando il micromanipolatore, regolare la punta dell'ago di microiniezione in modo che sia appena sopra l'acqua. Per fare questo, immergere con attenzione la punta in acqua (l'acqua potrebbe entrare nella punta della pipetta), quindi ritrarla in modo che sia vicina alla superficie epiteliale della medusa.
NOTA: La punta deve essere posizionata su un quadrante della medusa. I canali radiali della medusa dividono la campana della medusa in quattro quadranti distinti. Il targeting di un quadrante si tradurrà in immagini più pulite, poiché le gonadi e i canali radiali sono esclusi dall'area della ferita. - Pulsare l'aria premendo start sull'iniettore. Ripeti l'impulso nello stesso punto da due a quattro volte, a seconda della larghezza della punta. Le punte più grandi richiedono meno impulsi.
NOTA: Dovrebbe essere visibile una rientranza nell'acqua/medusa causata dall'impulso dell'aria. - Coprire l'animale ferito con un copricostume per l'imaging di ferite di grandi dimensioni (Figura 1D,D').
- Seguire i passaggi riportati di seguito (paragrafo 3) per l'imaging della guarigione delle ferite epiteliali.
- Fare un ago per microiniezione usando un estrattore di micropipette e un tubo capillare di vetro (punto 5.2). Inserire l'ago di microiniezione vuoto in un supporto per microiniettore fissato a un micromanipolatore. Tagliare la punta dell'ago in modo che l'apertura sia di circa 20-40 μm.
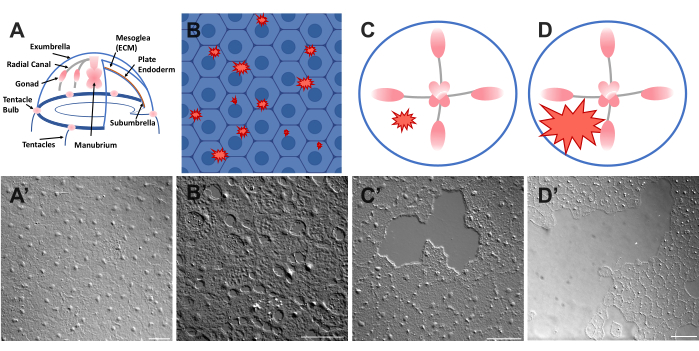
Figura 1: Strato epiteliale dell'esplosione intatto e ferito in Clytia medusa. (A) Grafica a fumetti del corpo di Clytia medusa. (A») Epitelio dell'ombrello medusa intatto visto dall'alto. (B) Vignetta di microferite unicellulari (forme frastagliate rosse) con cellule epiteliali in blu. (B') Microferite unicellulari. (C) Cartone animato di una piccola ferita epiteliale (forma frastagliata rossa). (C') Piccola ferita epiteliale. (D) Cartone animato di una grande ferita epiteliale (forma frastagliata rossa). (') Grande ferita epiteliale. Le immagini sono state tutte ottenute utilizzando la microscopia DIC. Barre della scala in (A'-C'): 50 μm. Barra di scala in (D'): 100 μm. Fare clic qui per visualizzare una versione ingrandita di questa figura.
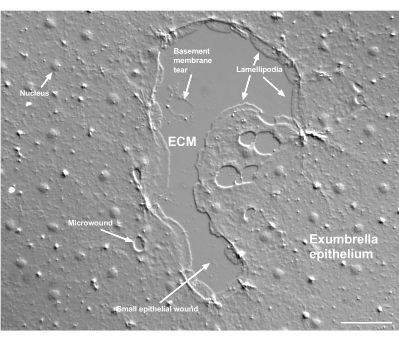
Figura 2: Ferite di dimensioni multiple e una membrana basale danneggiata. Viene mostrata una tipica piccola ferita epiteliale ad ombrello, con etichette che indicano lamellipodi che si formano da cellule marginali. Inoltre, si osservano microferite all'interno e tra le cellule epiteliali. Si noti la piccola lacerazione della membrana basale nella parte superiore della ferita. Il film 4 mostra la guarigione di questa ferita. Barra scala: 50 μm. Fare clic qui per visualizzare una versione ingrandita di questa figura.
3. Imaging della guarigione delle ferite epiteliali
- Assicurarsi che il microscopio sia stato allineato per l'illuminazione Köhler53 e che sia stato impostato correttamente per la microscopia a contrasto differenziale interferente (DIC)54. Le cellule epiteliali sono quasi invisibili con l'ottica standard (Figura 3D,E).
- Regolare la messa a fuoco sull'ombrello. Sebbene si tratti di uno strato sottile, le celle esagonali dovrebbero essere chiare.
NOTA: L'exumbrella e il subumbrella sono separati da una spessa mesoglea sostenuta da fibre verticali. Le celle subombrello si trovano sullo stesso piano focale dei canali radiali. Se inizialmente focalizzato sullo strato del subombrellore, regolare lentamente la messa a fuoco attraverso la mesoglea e le fibre verticali fino a trovare l'ombrello. - Identificare manualmente una ferita da visualizzare. Per ferite di grandi dimensioni, utilizzare un obiettivo 10x. Per ferite più piccole e ferite unicellulari, utilizzare un obiettivo 20x.
- Avviare un programma che raccoglie immagini come filmato in tempo reale o che raccoglie una serie di immagini a intervalli regolari. Monitorare i progressi per assicurarsi che l'area della ferita non si allontani dal campo visivo e che le cellule di interesse rimangano a fuoco.
- Le ferite unicellulari si chiudono entro un minuto; Pertanto, immagina la loro chiusura con un film.
- Per catturare i dettagli della dinamica cellulare per piccole ferite, raccogliere immagini circa ogni 10 secondi. La chiusura di piccole ferite richiede 20-50 minuti a seconda delle dimensioni.
- Non visualizzare i vetrini non sigillati per più di 45 minuti, poiché l'evaporazione dell'acqua dal vetrino nel tempo porta alla morte degli animali e alla rottura delle cellule.
- Per un'osservazione più lunga, sigillare attorno al coprivetrino con vaselina per ridurre l'evaporazione.
NOTA: Alcune medusa possono pulsare sul vetrino, che interferisce con l'imaging. In questo caso, montare animali in una diluizione 1:10 di 3-amminobenzoato di etile all'1% metansolfonato (tricaina), regolato a pH 7,5, in ASW serve come anestetico efficace e non ha alcun effetto apparente sulla guarigione in un lasso di tempo di 1 ora. Tuttavia, gli animali moriranno se lasciati per diverse ore nella tricaina.

Figura 3: Creazione di una piccola ferita nell'epitelio exumbrellar. (A) Graffiare delicatamente l'ombrello con una punta di pipetta da 200 μL per creare una piccola ferita epiteliale. (B) Il posizionamento del foglietto di copertura è talvolta sufficiente per creare piccole ferite epiteliali. (C) Medusa montata su uno scivolo a depressione. (D ) Piccola immagine della ferita epiteliale senza ottica DIC e (E) con ottica DIC. Barre della scala: 50 μm Fare clic qui per visualizzare una versione ingrandita di questa figura.
4. Analisi
- Preparazione dei file di immagine
NOTA: per elaborare i file immagine, utilizzare FIJI/ImageJ con plug-in BioFormat aggiornati.- Impostare la scala sul corretto rapporto pixel per micron prima di registrare la pila di immagini; Analizza > imposta scala. Ciò è necessario per estrarre le misurazioni delle dimensioni effettive nelle analisi a valle.
- Spesso, l'animale si sposta leggermente sul vetrino del microscopio; pertanto, per eliminare la deriva nei filmati, registrare le immagini utilizzando il plugin FIJI linear stack alignment with SIFT. Plugin > la registrazione > l'allineamento lineare dello stack con SIFT.
- Salvare lo stack registrato come file .avi. File > Salva come > AVI ... Nel pop-up, impostare la frequenza dei fotogrammi (le figure animate qui sono impostate su 10 fps) e fare clic su OK. Utilizzare questo output per eseguire l'analisi della guarigione delle ferite.
- Analisi dell'area della ferita
- Utilizzando lo strumento lazo in FIJI/ImageJ, delineare la ferita tracciando i bordi delle celle. Misurare l'area della ferita appena delineata con Comando+M o CTRL+M.
- Ripetere la misurazione dell'area della ferita ogni 10 fotogrammi. Le misurazioni di FIJI/ImageJ possono quindi essere tracciate utilizzando il prisma 9 (figura 4).
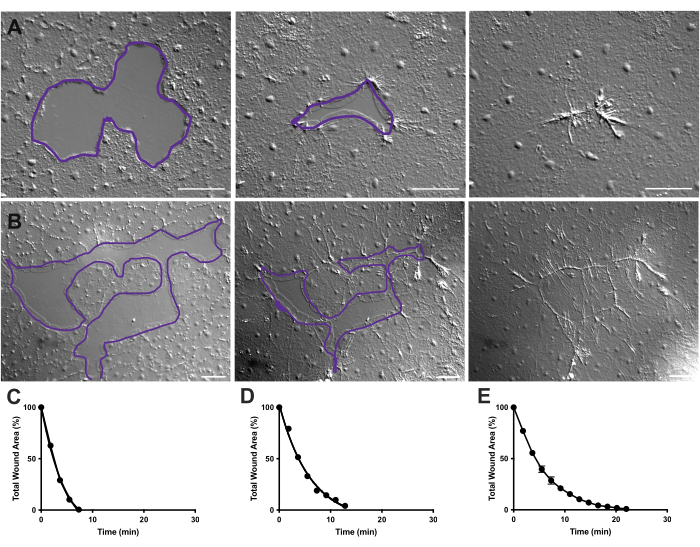
Figura 4: Analisi dell'area della ferita in piccole ferite epiteliali. (A ) Esempio di una piccola ferita epiteliale che guarisce oltre 10 min. (B) Esempio di una diversa guarigione della ferita epiteliale nell'arco di 21 min. I contorni viola in A,B sono paragonabili alle misurazioni delle aree della ferita utilizzando lo strumento lazo in FIJI/ImageJ. (C) Riduzione normalizzata dell'area della ferita nel tempo in A. (D) Riduzione normalizzata dell'area della ferita nel tempo in B. (E) Riduzione media dell'area della ferita nel tempo per 14 piccole ferite. n = 14. Barre di errore incentrate sul SEM ± medio. Barre della scala: 50 μm Fare clic qui per visualizzare una versione più grande di questa figura.
5. Iniezioni mesogleali
- Creazione del piatto di iniezione
- Preparare il polidimetilsilossano (PDMS) combinando una base PDMS e un agente di polimerizzazione, in un rapporto di 10 parti di base a 1 parte di agente di polimerizzazione in peso. Mescolare energicamente per mescolare completamente la base e l'agente indurente.
- Per rimuovere le bolle, mettere la miscela in una camera a vuoto per 15 minuti. Versare la miscela in una capsula di Petri da 60 mm con tappi a tubo microcentrifuga per mantenere lo stampo in posizione. Posizionare immediatamente lo stampo sui tappi dei tubi con un'inclinazione di 45° e del nastro adesivo. Lo stampo è costituito da tre vetrini sfalsati impilati incollati insieme per creare creste nel piatto di iniezione finale.
- Mettere l'intero piatto, lo stampo e il composto in forno a 60 °C per 2 ore per polimerizzare l'elastomero. Rimuovere lo stampo per un piatto di iniezione completato.
- Tiratura di micropipette
- Utilizzando un estrattore a microelettrodi, progettare un programma di trazione. Utilizzare un programma one-step ad alta velocità. Il calore è approssimativamente il risultato del test RAMP in vetro55,56. Controllare le micropipette risultanti per ottenere conicità lunghe e coerenti.
NOTA: Utilizzare capillari in borosilicato di vetro a parete sottile con diametro esterno di 1,0 mm, diametro interno di 0,75 mm e lunghezza di 10 cm.
- Utilizzando un estrattore a microelettrodi, progettare un programma di trazione. Utilizzare un programma one-step ad alta velocità. Il calore è approssimativamente il risultato del test RAMP in vetro55,56. Controllare le micropipette risultanti per ottenere conicità lunghe e coerenti.
- Iniezione di coloranti e farmaci
- Fare un ago per microiniezione (come sopra).
- Riempire l'ago per microiniezione utilizzando una lunga punta di pipetta con un volume eccessivo di colorante o farmaco iniettabile nella medusa.
NOTA: Per Clytia, il dimetilsolfossido (DMSO) deve essere mantenuto ad una diluizione di <1:100 con ASW, poiché concentrazioni più elevate di DMSO impediscono la guarigione delle ferite. Se si inietta una soluzione limpida, è possibile aggiungere la soluzione Fast Green FCF (diluizione 1:100 dello 0,1% Fast Green FCF in ASW) per visualizzare il liquido iniettato. - Utilizzando una pipetta di trasferimento modificata come sopra, posizionare una medusa con il sottoombrello rivolto verso l'alto in un piatto di iniezione PDMS con ASW appena sufficiente a coprire l'animale (Figura 5C). Posiziona il piatto sul palcoscenico di un cannocchiale da dissezione.
NOTA: Limitare l'eccesso di ASW impedisce alla medusa di nuotare nel piatto e consente iniezioni più efficaci. - Concentrarsi sulla punta dell'ago di microiniezione e farla avanzare nell'acqua vicino alla medusa. Con il micromanipolatore, premere l'ago nel piatto fino a quando non si piega e si rompe. Questa apertura della punta è di circa 10-20 μm.
NOTA: Questo ago può essere utilizzato ripetutamente per le stesse iniezioni di colorante/farmaco quel giorno. Si consiglia di utilizzare una punta fresca ogni giorno e per coloranti / farmaci separati. - Utilizzando il micromanipolatore, inserire la punta dell'ago attraverso il sottoombrello nella mesoglea senza perforare l'ombrello.
NOTA: Una piegatura / piegatura dell'epitelio sarà evidente. Una volta inserito l'ago nella medusa, la piegatura cessa. - Sul microiniettore, impostare la pressione di mantenimento a zero e la pressione di espulsione a ≤20 PSI. Iniettare in uno o due quadranti, riempiendo ciascuno con un punto di colorante o droga circa 1/4 dell'area di quel quadrante.
NOTA: A seconda delle dimensioni della medusa, volumi più grandi o più piccoli sono appropriati in singoli punti di iniezione. Il riempimento eccessivo della medusa provoca danni estremi all'epitelio e persino la morte dell'animale. - A seconda del colorante o del farmaco che viene iniettato, gli animali vengono posti in un becher di ASW fresco per consentire la diffusione e l'incubazione del colorante o del farmaco.
- Per l'imaging, montare la medusa su un vetrino di depressione usando una pipetta di trasferimento modificata, posizionando l'animale in modo che l'ombrello sia rivolto verso l'alto (Figura 5). Gli animali possono essere feriti in questa fase per testare l'effetto di un reagente iniettato.
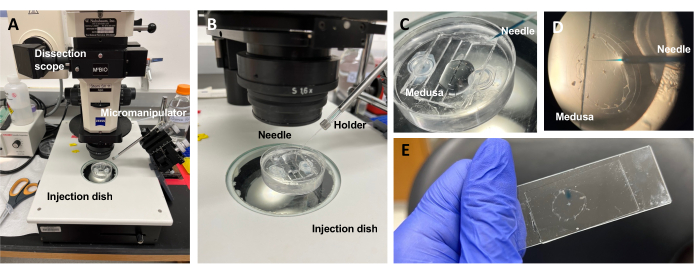
Figura 5: Configurazione dell'iniezione per l'introduzione di coloranti o farmaci nell'ECM. (A) Configurazione dell'iniezione. (B) Primo piano della configurazione dell'iniezione che mostra l'orientamento dell'ago di microiniezione (angolo di circa 45° rispetto all'animale nel piatto). (C) Primo piano del piatto di iniezione di silicone con la medusa in una piccola quantità di ASW per iniezione. (D) Un ago di microiniezione caricato con Fast Green FCF che entra nella mesoglea della medusa attraverso il subombrello. (E) Post-iniezione di Fast Green FCF in una medusa montata. Fare clic qui per visualizzare una versione ingrandita di questa figura.
Seguendo i protocolli di cui sopra, sono state visualizzate microferite a singola cellula, piccole ferite e grandi ferite. Le pile registrate di file di immagine sono state salvate come file .avi.
Nel filmato 1, le microferite possono essere viste chiudersi tra e all'interno delle cellule (Figura 1 e Figura 2). Piccoli lamellipodi si osservano durante la chiusura, seguiti da contrazione e guarigione. I detriti vengono esclusi e rilasciati nell'acqua. La guarigione è completata in un minuto o meno.
Nei filmati 2 e 3, piccole ferite di forme diverse guariscono attraverso la formazione di lamellipodi, l'estensione dei contatti lamellipodiali e la diffusione delle cellule al margine della ferita, come descritto in precedenza40 (Figura 1 e Figura 2). Le cellule in livelli dietro le cellule marginali non partecipano alla guarigione di ferite di queste dimensioni né c'è migrazione cellulare collettiva. La chiusura rapida e progressiva delle lacune epiteliali è seguita dalla contrazione dei tessuti lungo la cucitura della ferita appena formata40. Viene mostrato il tasso normalizzato di guarigione di queste due ferite, espresso come percentuale dell'area originale nel tempo (Figura 4C, D). Mentre c'è una certa variabilità nella dinamica della chiusura della ferita, la media della percentuale di chiusura dell'area nel tempo per 14 ferite di varie forme che vanno da 0,02 a 0,125 mm2 consente di stabilire una curva media per la guarigione delle ferite negli animali non trattati (Figura 4E).
Il danno alla membrana basale può essere visto chiaramente quando si verifica (Figura 2). Nel filmato 4, le cellule al margine di una piccola ferita in cui vi è un danno alla membrana basale si diffondono intorno all'area danneggiata e la chiusura della fessura è completata con una contrazione del cordone della borsa.
Se il tessuto è disidratato o troppo danneggiato per essere riparato, i movimenti cellulari possono fermarsi o l'intero foglio di cellule può scoppiare (Movie 5 e Movie 6). Questo di solito accade dopo lunghi periodi di imaging (45 minuti o più). Se l'esplosione cellulare si verifica all'inizio dell'imaging, il campione viene scartato.
Come mostrato nel film 7, le grandi ferite guariscono in più fasi. In primo luogo, il bordo della ferita diventa liscio e regolare a causa delle contrazioni al margine, come precedentemente riportato57. Quindi, i lamellipodi sono visti formarsi dalle cellule al margine della ferita, con lamellipodia che si sposta in avanti per massimizzare il contatto con i lamellipodi adiacenti. Il monitoraggio dei nuclei nelle cellule al margine della ferita e diversi livelli dietro le cellule marginali mostra che grandi lacune si chiudono per migrazione cellulare collettiva40. Le cellule non si staccano mai, ma si muovono insieme come un foglio.
L'introduzione di coloranti e agenti farmacologici può essere un potente strumento per sezionare i meccanismi biologici. Molte sostanze sono escluse da Clytia (non mostrate), probabilmente a causa dello strato di muco che ricopre la superficie dell'animale. Tuttavia, la microiniezione può essere utilizzata per introdurre direttamente molecole nella ECM, interrompendo la struttura della ECM o perturbando le attività regolatorie nella ECM. Inoltre, coloranti e altre molecole sono in grado di entrare nelle cellule epiteliali dal lato basale. Ad esempio, la Figura 6 mostra la colorazione nucleare con Hoechst, la colorazione a membrana con FM1-43 e l'inibizione della formazione di lamellipodi da parte della citocalasina B dopo che questi reagenti sono stati microiniettati nella ECM. L'introduzione di queste molecole nella ECM e nelle cellule epiteliali prima del ferimento consente esperimenti che testano l'effetto degli strumenti farmacologici sul processo di guarigione.
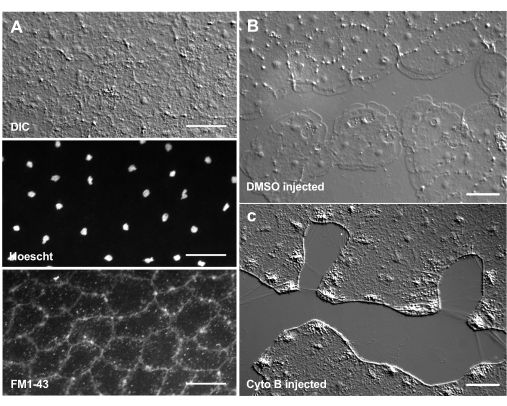
Figura 6: Cellule epiteliali della medusa dopo microiniezione di coloranti o agenti farmacologici. (A) Cellule epiteliali mostrate nel pannello superiore 5 minuti dopo l'iniezione con 20 μM di Hoechst (nuclei) e 50 μM FM1-43 (membrane). (B,C) Guarigione della ferita dopo iniezione con 1:1.000 DMSO di controllo (B) o 100 μM di citocalasina B (C). Le ferite sono state fatte 15 minuti dopo l'iniezione. Le immagini sono state scattate 5 minuti dopo il ferimento. La formazione di lamellipodi è inibita dalla citocalasina B. Si ritiene che le apparenti "fibre" spesso viste tra le cellule nell'area della ferita siano il risultato della tensione che allunga la membrana basale - non si macchiano di falloidina (non mostrata). Barre della scala: 50 μm. Fare clic qui per visualizzare una versione ingrandita di questa figura.
Filmato 1: Filmato time-lapse della guarigione delle microferite a cellula singola. Tempo trascorso: 20 s. Frequenza fotogrammi: 10 fps. Barra scala: 50 μm. Clicca qui per scaricare questo film.
Filmato 2: Filmato time-lapse di una piccola ferita epiteliale che guarisce. Tempo trascorso: 9 min 54 s. Frequenza fotogrammi: 10 fps. Barra scala: 50 μm. Clicca qui per scaricare questo film.
Filmato 3: Filmato time-lapse di una piccola ferita epiteliale che guarisce. Questa ferita è più grande e di forma più irregolare rispetto alla ferita nel film 2. Tempo trascorso: 20 min 54 s. Frequenza fotogrammi: 10 fps. Barra scala: 50 μm. Clicca qui per scaricare questo film.
Filmato 4: Filmato time-lapse di una piccola ferita e una microferita che guarisce con una rottura della membrana basale. I lamellipodi si diffondono intorno alla lacerazione della membrana basale, sebbene possano avanzare sul resto della ECM. Una volta che la regione della ferita con il danno alla membrana basale è circondata, una contrazione del cordone della borsa tira le cellule sulla regione. Tempo trascorso: 19 min 4 s. Frequenza fotogrammi: 10 fps. Barra scala: 50 μm. Clicca qui per scaricare questo film.
Filmato 5: Cellule che muoiono in una piccola ferita epiteliale. La morte cellulare è probabilmente dovuta alla disidratazione dell'animale. Tempo trascorso: 4 min 24 s. Frequenza fotogrammi: 10 fps. Barra scala: 100 μm. Clicca qui per scaricare questo film.
Filmato 6: Una piccola ferita epiteliale non riesce a completare la guarigione. Tempo trascorso: 42 min 32 s. Frequenza fotogrammi: 10 fps. Barra scala: 50 μm. Clicca qui per scaricare questo film.
Filmato 7: Grande guarigione delle ferite epiteliali. Tempo trascorso: 25 min 29 s. Frequenza fotogrammi: 10 fps. Barra scala: 100 μm. Clicca qui per scaricare questo film.
Figura supplementare 1: Schemi delle dimensioni del serbatoio Clytia. Visualizzazione 3D dei serbatoi Clytia su misura. (A) Vista anteriore e posteriore. (B) Vista laterale. Il ritaglio nel pezzo mostrato in verde è rivestito con rete di nylon. L'acqua entra nel serbatoio direttamente sulla rete, spazza la rete e crea una corrente circolare. L'acqua esce dal sistema attraverso il foro nel pezzo terminale mostrato in blu. Clicca qui per scaricare questo file.
Filmato supplementare 1: Matrice extracellulare acellulare in Clytia. Z-stack di Clytia prelevato con microscopia confocale. Lo stack inizialmente si concentra sull'exumbrella e poi esegue scansioni ogni 10 μm attraverso l'ECM fino all'endoderma della piastra e al subumbrella. Le immagini che utilizzano DIC (a sinistra) e la colorazione nucleare di Hoechst (a destra) dimostrano la mancanza di cellule nell'ECM. Barra scala: 100 μm. Clicca qui per scaricare questo file.
Qui, viene presentata la metodologia per l'imaging delle ferite in vivo in Clytia, un organismo modello di invertebrati relativamente nuovo40,43,58. Ci sono diversi fattori che rendono questo sistema uno strumento di ricerca unico e potente, distinto da altri modelli utilizzati per studiare la guarigione e la riepitelizzazione delle ferite. In primo luogo, l'epitelio monostrato è attaccato a una ECM trasparente, quindi simile ai saggi di coltura tissutale in vitro (Figura 1, Figura 2, Figura 3, Figura 4). Come nei saggi in vitro, le cellule possono essere visualizzate ad alta risoluzione. Tuttavia, a differenza della coltura tissutale, esiste un ambiente cellulare autentico e ECM, in modo che la guarigione delle ferite possa essere vista nel contesto dei complessi eventi di segnalazione che si verificano in un animale vivo ferito. In secondo luogo, la clizia manca di risposte infiammatorie, fibroblasti migratori, vascolarizzazione e sangue. Ciò consente di studiare in vivo il processo di riepitelizzazione in assenza degli eventi sovrapposti che si verificano in animali adulti più complessi durante la guarigione delle ferite59. In terzo luogo, l'ECM è acellulare (Filmato supplementare 1) e di grandi dimensioni, consentendo un facile accesso con un ago per microiniezione (Figura 5 e Figura 6). Utilizzando questo approccio, i ricercatori possono testare l'effetto dei reagenti farmacologici che perturbano la struttura o la segnalazione della ECM sulla guarigione delle ferite in vivo. I reagenti possono anche essere introdotti nelle cellule epiteliali e possono essere valutati i loro effetti sulla guarigione delle ferite in vivo. In quarto luogo, esistono protocolli per la creazione di mutanti e animali transgenici nel sistema Clytia42,43,44,45. La guarigione delle ferite in vivo può quindi essere osservata in animali con aumentata/diminuita espressione di geni di interesse.
Ci sono diversi passaggi critici in questa tecnica. Innanzitutto, come mostrato nella Figura 3, è necessario utilizzare un microscopio configurato correttamente per la microscopia DIC poiché le cellule epiteliali piatte e trasparenti sono quasi invisibili con la microscopia ottica standard. È anche importante sviluppare l'abilità di ferire delicatamente gli animali in modo che l'epitelio sia danneggiato senza scavare l'ECM. Un tocco altrettanto delicato è necessario per microiniettare materiali nell'ECM, poiché danni estesi all'animale durante l'iniezione potrebbero compromettere una successiva analisi della guarigione delle ferite. Mentre c'è una curva di apprendimento per queste tecniche, anche gli studenti principianti le hanno padroneggiate rapidamente nel laboratorio Malamy. In effetti, questi protocolli sono stati utilizzati per dimostrare la migrazione cellulare nei corsi di laboratorio universitari presso l'Università di Chicago.
Per un'imaging ottimale, è importante che l'animale non si muova e che l'area della ferita scelta non si allontani dal campo visivo. Se gli animali pulsano, il trattamento con Tricaina come descritto è molto efficace. Per la deriva, è spesso necessario riposizionare manualmente il campione. Questi movimenti possono essere eliminati dal filmato finale utilizzando la funzione di registrazione in FIJI/ImageJ.
Una limitazione di questo sistema è che non è possibile creare ferite identiche, poiché le ferite variano sia nella forma che nelle dimensioni utilizzando i metodi descritti qui. Pertanto, può essere difficile quantificare il tasso esatto di chiusura della ferita o migrazione cellulare. I marcatori posizionali come i grani di carbonio si attaccano all'ECM esposto in un animale ferito e possono essere utilizzati per misurare il tasso di migrazione cellulare collettiva in grandi ferite (non mostrato). Per l'analisi della chiusura di piccole ferite, anche con dimensioni e forma della ferita variabili, esiste una gamma limitata di tassi di chiusura tra ferite di queste dimensioni (Figura 4). È quindi possibile rilevare quantitativamente gli effetti dei reagenti farmacologici promozionali o repressivi.
Mentre questo lavoro descrive la caratterizzazione della guarigione delle ferite usando solo la microscopia DIC, gli stessi approcci possono essere utilizzati per la guarigione delle immagini utilizzando la fluorescenza o la microscopia confocale. Per aiutare in questo, sono in atto protocolli per generare animali transgenici in cui varie proteine cellulari ed extracellulari sono marcate in modo fluorescente. L'imaging concomitante con DIC e fluorescenza, combinato con la perturbazione della guarigione delle ferite utilizzando agenti farmacologici o linee mutanti, sarà un potente approccio per comprendere i meccanismi che sono alla base del processo di guarigione delle ferite nell'epitelio.
Nulla da rivelare.
E.E.L.L. è sostenuto da una sovvenzione della National Science Foundation PRFB 2011010. Vorremmo ringraziare Tsuyoshi Momose ed Evelyn Houliston per averci aiutato a stabilire le nostre colonie di Clytia, Jean-Baptiste Reynier per la raccolta delle immagini di guarigione delle microferite, Harry Kyriazes per la costruzione delle vasche pseudo-kreisel ed Elizabeth Baldo per il mantenimento dell'habitat di Clytia. La Figura 1B è stata creata con BioRender.com.
| Name | Company | Catalog Number | Comments |
| 20500 ACE EKE Microscope Fiber Optic Light Source | Kramer Scientific Corporation | ||
| AxioCam 506 mono | ZEISS | 426557-0000-000-MA285 | |
| Capillary tubes | World Precision Instruments | TW1004 | |
| Cytochalasin B | Abcam | ab143482 | |
| Depression slides | Amscope | BS-C12 | |
| DMR with DIC options and fluorescence halogen lamp | Leica | ||
| Ethyl 3-aminobenzoate methanesulfonate | Sigma Aldrich | E10521-10G | |
| Fast Green FCF | Thermo Scientific | A16520-06 | |
| FM1-43 | Biotium | 70022 | Excitation/Emission: 480/598 nm |
| Hoechst 33342 | Thermo Scientific | 62249 | Excitation/Emission: 361/497 nm |
| imageJ | NIH | ||
| Microloader tips (0.5-10 μL /2-20 μL) | Eppendorf | 930001007 | |
| Micromanipulator | World Precision Instruments | 3301R / M3301L | |
| Microscope Cover Glass (22X40-1.5) | Fisherbrand | 12-544-BP | |
| Petri Dish (60 mm x 15 mm) | Fisherbrand | FB085713A | |
| PicoNozzle v2 | World Precision Instruments | 5430-ALL | |
| Pipette puller | Sutter Instrument Co | P-97 | |
| Pneumatic PicoPump | World Precision Instruments | PV820 | |
| Polycarbonate vacuum, desiccator | Bel-art | F42025-0000 | |
| Prism 9 | GraphPad | ||
| STEMI Sv11 Dissection scope | ZEISS | STEMI SV11 | |
| SYLGARD 184 | Dow Silicones | 1024001 | |
| Transfer pipettes | Fisherbrand | 13-711-7M | |
| Z-Hab mini system | Pentair | ||
| ZEN Microscopy software | Zeiss |
- Tyler, S. Epithelium-the primary building block for metazoan complexity. Integrative and Comparative Biology. 43 (1), 55-63 (2003).
- Kurn, H., Daly, D. T. Histology, Epithelial Cell. StatPearls. , (2022).
- Schempp, C., Emde, M., Wölfle, U. Dermatology in the Darwin anniversary. Part 1: Evolution of the integument. Journal of the German Society of Dermatology. 7 (9), 750-757 (2009).
- Lopez-Ojeda, W., Pandey, A., Alhajj, M., Oakley, A. M. Anatomy, Skin (Integument). StatPearls. , (2022).
- Bukowiecki, A., Hos, D., Cursiefen, C., Eming, S. A. Wound-healing studies in cornea and skin: parallels, differences and opportunities. International Journal of Molecular Sciences. 18 (6), 1257 (2017).
- Frantz, C., Stewart, K. M., Weaver, V. M. The extracellular matrix at a glance. Journal of Cell Science. 123 (24), 4195-4200 (2010).
- Hynes, R. O. The evolution of metazoan extracellular matrix. The Journal of Cell Biology. 196 (6), 671-679 (2012).
- Fidler, A. L., et al. Collagen IV and basement membrane at the evolutionary dawn of metazoan tissues. eLife. 6, 24176 (2017).
- Bullen, T. F., et al. Characterization of epithelial cell shedding from human small intestine. Laboratory Investigation; a Journal of Technical Methods and Pathology. 86 (10), 1052-1063 (2006).
- Watson, A. J. M., et al. Epithelial barrier function in vivo is sustained despite gaps in epithelial layers. Gastroenterology. 129 (3), 902-912 (2005).
- Sonnemann, K. J., Bement, W. M. Wound repair: toward understanding and integration of single-cell and multicellular wound responses. Annual Review of Cell and Developmental Biology. 27, 237-263 (2011).
- Abreu-Blanco, M. T., Verboon, J. M., Parkhurst, S. M. Single cell wound repair. BioArchitecture. 1 (3), 114-121 (2011).
- Fenteany, G., Janmey, P. A., Stossel, T. P. Signaling pathways and cell mechanics involved in wound closure by epithelial cell sheets. Current Biology. 10 (14), 831-838 (2000).
- Pastar, I., et al. Epithelialization in wound healing: a comprehensive review. Advances in Wound Care. 3 (7), 445-464 (2014).
- Rousselle, P., Braye, F., Dayan, G. Re-epithelialization of adult skin wounds: Cellular mechanisms and therapeutic strategies. Advanced Drug Delivery Reviews. 146, 344-365 (2019).
- Bement, W. M., Forscher, P., Mooseker, M. S. A novel cytoskeletal structure involved in purse string wound closure and cell polarity maintenance. The Journal of Cell Biology. 121 (3), 565-578 (1993).
- Vedula, S. R. K., Ravasio, A., Lim, C. T., Ladoux, B. Collective Cell migration: a mechanistic perspective. Physiology. 28 (6), 370-379 (2013).
- Li, L., He, Y., Zhao, M., Jiang, J. Collective cell migration: Implications for wound healing and cancer invasion. Burns & Trauma. 1 (1), 21-26 (2015).
- Bornes, L., Windoffer, R., Leube, R. E., Morgner, J., van Rheenen, J. Scratch-induced partial skin wounds re-epithelialize by sheets of independently migrating keratinocytes. Life Science Alliance. 4 (1), 202000765 (2021).
- Theveneau, E., Mayor, R. Collective cell migration of epithelial and mesenchymal cells. Cellular and Molecular Life Sciences. 70 (19), 3481-3492 (2013).
- Haensel, D., Dai, X. Epithelial-to-mesenchymal transition in cutaneous wound healing: where we are and where we are heading. Developmental Dynamics. 247 (3), 473-480 (2018).
- Cordeiro, J. V., Jacinto, A. The role of transcription-independent damage signals in the initiation of epithelial wound healing. Nature Reviews. Molecular Cell Biology. 14 (4), 249-262 (2013).
- Abreu-Blanco, M. T., Watts, J. J., Verboon, J. M., Parkhurst, S. M. Cytoskeleton responses in wound repair. Cellular and Molecular Life Sciences. 69 (15), 2469-2483 (2012).
- Klarlund, J. K., Block, E. R. Free edges in epithelia as cues for motility. Cell Adhesion & Migration. 5 (2), 106-110 (2011).
- Enyedi, B., Niethammer, P. Mechanisms of epithelial wound detection. Trends in Cell Biology. 25 (7), 398-407 (2015).
- Niethammer, P. The early wound signals. Current Opinion in Genetics & Development. 40, 17-22 (2016).
- Jacinto, A., Martinez-Arias, A., Martin, P. Mechanisms of epithelial fusion and repair. Nature Cell Biology. 3 (5), 117-123 (2001).
- Mayor, R., Etienne-Manneville, S. The front and rear of collective cell migration. Nature reviews. Molecular Cell Biology. 17 (2), 97-109 (2016).
- Gupta, S., Yap, A. S. How adherens junctions move cells during collective migration. Faculty Reviews. 10, 56 (2021).
- Blair, M. J., Jones, J. D., Woessner, A. E., Quinn, K. P. Skin structure-function relationships and the wound healing response to intrinsic aging. Advances in Wound Care. 9 (3), 127-143 (2020).
- Falanga, V., et al. Chronic wounds. Nature Reviews. Disease Primers. 8 (1), 50 (2022).
- Liang, C. -. C., Park, A. Y., Guan, J. -. L. In vitro scratch assay: a convenient and inexpensive method for analysis of cell migration in vitro. Nature Protocols. 2 (2), 329-333 (2007).
- Jonkman, J. E. N., et al. An introduction to the wound healing assay using live-cell microscopy. Cell Adhesion & Migration. 8 (5), 440-451 (2014).
- Poujade, M., et al. Collective migration of an epithelial monolayer in response to a model wound. Proceedings of the National Academy of Sciences. 104 (41), 15988-15993 (2007).
- Masson-Meyers, D. S., et al. Experimental models and methods for cutaneous wound healing assessment. International Journal of Experimental Pathology. 101 (1-2), 21-37 (2020).
- Park, S., et al. Tissue-scale coordination of cellular behaviour promotes epidermal wound repair in live mice. Nature Cell Biology. 19 (2), 155-163 (2017).
- Tsai, C. -. R., Wang, Y., Galko, M. J. Crawling wounded: molecular genetic insights into wound healing from Drosophila larvae. The International Journal of Developmental Biology. 62 (6-7-8), 479-489 (2018).
- Richardson, R., et al. Adult zebrafish as a model system for cutaneous wound-healing research. The Journal of Investigative Dermatology. 133 (6), 1655-1665 (2013).
- Erickson, J. R., Echeverri, K. Learning from regeneration research organisms: The circuitous road to scar free wound healing. Developmental Biology. 433 (2), 144-154 (2018).
- Kamran, Z., et al. In vivo imaging of epithelial wound healing in the cnidarian Clytia hemisphaerica demonstrates early evolution of purse string and cell crawling closure mechanisms. BMC Developmental Biology. 17 (1), 17 (2017).
- Chari, T., et al. Whole-animal multiplexed single-cell RNA-seq reveals transcriptional shifts across Clytia medusa cell types. Science Advances. 7 (48), (2021).
- Weissbourd, B., et al. A genetically tractable jellyfish model for systems and evolutionary neuroscience. Cell. 184 (24), 5854-5868 (2021).
- Momose, T., et al. High doses of CRISPR/Cas9 ribonucleoprotein efficiently induce gene knockout with low mosaicism in the hydrozoan Clytia hemisphaerica through microhomology-mediated deletion. Scientific Reports. 8 (1), 11734 (2018).
- Houliston, E., Leclère, L., Munro, C., Copley, R. R., Momose, T. Past, present and future of Clytia hemisphaerica as a laboratory jellyfish. Current Topics in Developmental Biology. 147, 121-151 (2022).
- Schmid, V., et al. The extracellular matrix (mesoglea) of hydrozoan jellyfish and its ability to support cell adhesion and spreading. Hydrobiologia. 216 (1), 3-10 (1991).
- Day, R. M., Lenhoff, H. M. Hydra mesoglea: a model for investigating epithelial cell-basement membrane interactions. Science. 211 (4479), 291-294 (1981).
- Zhang, X., et al. The collagens of hydra provide insight into the evolution of metazoan extracellular matrices. The Journal of Biological Chemistry. 282 (9), 6792-6802 (2007).
- Danjo, Y., Gipson, I. K. Actin 'purse string' filaments are anchored by E-cadherin-mediated adherens junctions at the leading edge of the epithelial wound, providing coordinated cell movement. Journal of Cell Science. 111 (22), 3323-3332 (1998).
- Arenas Gómez, C. M., Sabin, K. Z., Echeverri, K. Wound healing across the animal kingdom: Crosstalk between the immune system and the extracellular matrix. Developmental Dynamics. 249 (7), 834-846 (2020).
- Lechable, M., et al. An improved whole life cycle culture protocol for the hydrozoan genetic model Clytia hemisphaerica. Biology Open. 9 (11), (2020).
- Casares, L., et al. Hydraulic fracture during epithelial stretching. Nature Materials. 14 (3), 343-351 (2015).
- Wayne, R. Chapter 4 - Bright-Field Microscopy. Light and Video Microscopy (Third Edition). , 95-116 (2019).
- Murphy, D. B., Davidson, M. W. . Fundamentals of Light Microscopy and Electronic Imaging: Second Edition. , (2012).
- . Micropipette Techniques for Electrophysiology Available from: https://www.sutter.com/micropipette/cookbook.html (2022)
- Brown, A. L., Johnson, B. E., Goodman, M. B. Making patch-pipettes and sharp electrodes with a programmable puller). Journal of Visualized Experiments. (20), e939 (2008).
- Klarlund, J. K. Dual modes of motility at the leading edge of migrating epithelial cell sheets. Proceedings of the National Academy of Sciences. 109 (39), 15799-15804 (2012).
- Houliston, E., Momose, T., Manuel, M. Clytia hemisphaerica: a jellyfish cousin joins the laboratory. Trends in Genetics. 26 (4), 159-167 (2010).
- Rodrigues, M., Kosaric, N., Bonham, C. A., Gurtner, G. C. Wound healing: a cellular perspective. Physiological Reviews. 99 (1), 665-706 (2019).
ABOUT JoVE
Copyright © 2024 MyJoVE Corporation. All rights reserved