
Summary
Abstract
Introduction
Protocol
Representative Results
Discussion
Acknowledgements
Materials
References
Biology
Karaktärisering av epitelial sårläkning in vivo med hjälp av Cnidarian Model Organism Clytia hemisphaerica
Denna artikel beskriver en metod för att skapa sår i epitelet hos en levande Clytia hemisphaerica medusa och avbilda sårläkning med hög upplösning in vivo. Dessutom presenteras en teknik för att introducera färgämnen och läkemedel för att störa signaleringsprocesser i epitelcellerna och extracellulär matris under sårläkning.
Alla djurorgan, från hud till ögon till tarmar, är täckta med ark av epitelceller som gör att de kan upprätthålla homeostas samtidigt som de skyddar dem mot infektion. Därför är det inte förvånande att förmågan att reparera epitelsår är avgörande för alla metazoaner. Epitelial sårläkning hos ryggradsdjur involverar överlappande processer, inklusive inflammatoriska svar, vaskularisering och re-epitelisering. Reglering av dessa processer innefattar komplexa interaktioner mellan epitelceller, angränsande celler och den extracellulära matrisen (ECM); ECM innehåller strukturella proteiner, reglerproteiner och aktiva små molekyler. Denna komplexitet, tillsammans med det faktum att de flesta djur har ogenomskinliga vävnader och otillgängliga ECM, gör sårläkning svår att studera hos levande djur. Mycket arbete med epitelial sårläkning utförs därför i vävnadsodlingssystem, med en enda epitelcelltyp pläterad som ett monolager på en artificiell matris. Clytia hemisphaerica (Clytia) ger ett unikt och spännande komplement till dessa studier, vilket gör att epitelial sårläkning kan studeras i ett intakt djur med en autentisk ECM. Det ektodermala epitelet i Clytia är ett enda lager av stora skivepitelceller, vilket möjliggör högupplöst avbildning med hjälp av differentiell interfererande kontrastmikroskopi (DIC) i levande djur. Frånvaron av migrerande fibroblaster, vaskulatur eller inflammatoriska reaktioner gör det möjligt att dissekera de kritiska händelserna vid re-epitelisering in vivo. Läkning av olika typer av sår kan analyseras, inklusive encelliga mikrosår, små och stora epitelsår och sår som skadar basalmembranet. Lamellipodia-bildning, handväskans strängkontraktion, cellsträckning och kollektiv cellmigration kan alla observeras i detta system. Dessutom kan farmakologiska medel introduceras via ECM för att modifiera cell: ECM-interaktioner och cellulära processer in vivo. Detta arbete visar metoder för att skapa sår i levande Clytia, fånga filmer av läkning och undersöka läkningsmekanismer genom att mikroinjicera reagens i ECM.
Ark av epitelceller täcker den yttre ytan av alla metazoaner, linjer inre organ och delar djurkroppen i diskreta fack. Epitelet separerar också den inre kroppen från den yttre miljön och skyddar den mot skador och infektioner. Därför var tillkomsten av epitelskikt en väsentlig del av utvecklingen av flercelliga djur, och epitelskikt ses hos alla djur från ryggradsdjur till de mest basala metazoanerna1. Epitelet hos vissa organ är ett enda monolager, såsom i lungluftsäckarna, blodkärlen och tarmen2, liksom i epidermis hos ryggradslösa djur som planaria och cnidarians3. I andra vävnader, såsom huden4 och hornhinnan5 hos ryggradsdjur, stratifieras epitelet, vilket innebär att det finns flera epitelcellskikt2. I alla fall är det mest basala epitelskiktet fäst vid basalmembranet, ett proteinark som bildar en specialiserad region av den extracellulära matrisen (ECM)6,7,8.
Brott i epitelet måste snabbt repareras för att återskapa ett kontinuerligt epitelark. Skador på epitelet uppstår under naturliga processer, såsom utsöndring av epitelceller i tarmen,9,10 och som ett resultat av inflammation eller fysiskt trauma. När en enda epitelcell skadas måste den antingen reparera sig själv eller elimineras så att de omgivande cellerna kan fästa vid varandra och stänga hålet11,12. I sår större än storleken på en enda cell måste epitelceller röra sig för att nå varandra och reparera arket13. Detta kan uppnås genom cellspridning om luckorna är små eller kan kräva migrering av epitelceller från sårets kanter för att stänga sårgapet. Denna senare process kallas re-epithelialization14,15. I embryonala vävnader sprids epitelceller och migrerar till nära sår eller dras över gapet genom sammandragning av aktomyosinkablar som bildas mellan cellerna vid sårmarginalen, i en mekanism som liknar en handväskasträng 16. I många vuxna vävnader innebär re-epitelisering migrering av sammanhängande cellark, där celler behåller sina korsningar med angränsande celler14,17,18. I andra vävnader demonteras cell-cellanslutningar och epitelceller beter sig mer som mesenkymala celler och rör sig på ett samordnat men oberoende sätt in i sårområdet under re-epitelisering 14,19,20,21.
Epitelcellrörelser regleras av komplexa interaktioner mellan de migrerande cellerna och mellan cellerna och ECM. Även om det finns en enorm mängd experimentell litteratur som behandlar mekanismer för såraktivering av epitelceller och efterföljande migration, återstår mycket att upptäcka. Till exempel har den initiala signalen som aktiverar epitelceller att migrera som svar på ett sår inte identifierats definitivt 22, och det är inte heller helt förstått hur aktin omfördelas för att skapa lamellipodi på sidan av epitelceller närmast såret 22,23,24,25,26,27. Kollektiv cellmigration kräver att information från celler vid såret delas med celler distala mot såret, och kommunikationsvägen är fortfarande oklar28. Cell:cellkorsningar och cell:ECM-fästen måste demonteras och reformeras när celler i arket ordnar om sig själva, men regleringen av denna process är dåligt förstådd14,29. Att göra framsteg i dessa och andra relaterade frågor är inte bara viktigt som ett grundläggande biologiskt problem utan också på grund av den kliniska betydelsen av korrekt sårläkning. Sjukdomar som äventyrar epitelcellernas förmåga att migrera korrekt resulterar i kroniska sår; Ett exempel är den genetiska sjukdomen epidermolysis bullosa, där gener som är involverade i bindningen av epitelcellerna till ECM muteras, vilket resulterar i ömtålig hud som skalar och blåsor. Re-epitelisering äventyras också i naturligt åldrande vävnader30,31. En bättre förståelse är därför avgörande för att utveckla interventioner för att förbättra sårläkningsresultaten.
Epitelcellmigration vid sårläkning har studerats med både in vitro-metoder och modellorganismer. Majoriteten av studierna av sårläkning och mekanismer för cellmigration har utförts i vävnadsodling, där monolager av en enda epitelcelltyp odlas på ett substrat som ersätter ECM. Cellmonolager repas eller odlas med stenciler för att skapa luckor av specifika former och storlekar och observeras sedan32,33,34. In vitro-modellen möjliggör en idealisk visualisering av cellbeteende, liksom möjligheten att ändra substratets egenskaper, att exponera celler för läkemedel och abiotiska och biotiska faktorer och att transfektera celler med konstruktioner som uttrycker eller undertrycker olika gener av intresse. Detta reduktionistiska tillvägagångssätt kan emellertid misslyckas med att fånga några av de viktiga parametrarna som är involverade i epitelcellbeteende i ett in vivo-sammanhang, inklusive kommunikation mellan olika celltyper och signalhändelser som inträffar i ECM11. In vivo-modeller ger det autentiska sammanhanget för ett sår, med flera celltyper, överlappande signalvägar och en komplex ECM35. En sådan modell för sårläkningsstudier är mus19, där de senaste framstegen har gjort det möjligt för forskare att observera epidermala celler under läkning av fullhudssår hos levande djur36. Musen och andra in vivo-system utgör dock utmaningar för att studera re-epithelialisering. För det första balanseras den stora fördelen med att observera cellbeteende i ett naturligt sammanhang av komplexiteten hos de temporärt överlappande händelserna som inträffar under läkning av ryggradsdjur, inklusive blodkoagulering, rekrytering av immunceller och inflammation, rekrytering av fibroblaster och celldedifferentiering, revaskularisering och ombyggnad av ECM. Vidare gör ogenomskinliga vävnader avbildning svår. Drosophila-larven och zebrafiskepidermissystemen 37,38 har övervunnit några av dessa svårigheter på grund av deras relativa enkelhet 39.
Vårt laboratorium introducerade nyligen en ny modell för att studera epitelial sårläkning: medusa (maneter) form av hydrozoan cnidarian Clytia hemisphaerica (Clytia)40. Clytia är en framväxande modellorganism med ett fullständigt sekvenserat och annoterat genom 41, encells-RNAseq-transkriptom42 och protokoll på plats för genommodifiering (mutagenes och transgenes)43,44,45. Cnidarians är en av de äldsta existerande linjerna som har epitellager, så att förstå cnidarian sårläkning ger insikter i förfädernas vägar som säkerställde epitelintegritet. För de vägar som har bevarats genom livets träd erbjuder Clytia ett spännande nytt system för att studera epitelcelldynamik och funktionell reglering av sårläkning in vivo.
Epitelet som täcker den övre ytan av Clytia medusa (exparaply) är ett monolager av transparenta, skivepitelceller som är ungefär 50 μm breda och 1-2 μm tjocka (figur 1). De är fästa vid en ECM som kallas mesoglea - manetens "gelé". Mesoglea är kompositionellt lik ECM som finns hos andra djur 46,47,48 inklusive ryggradsdjur, har ett basalmembran 40 och är helt transparent. Epitelskiktet i Clytia medusa kan lätt repas eller skadas (se nedan). Enkelheten och transparensen i epitelet och ECM möjliggör högupplöst avbildning av cellerna och deras rörelser under läkning. Nyligen karakteriserade Kamran et al. läkning av små sår i Clytia-epitelet i detalj40. Det visades att läkning i Clytia sker genom lamellipodia-baserad cellkrypning, cellspridning och kollektiv cellmigration, liksom handväska strängstängning som är mer typisk för embryonala system (även om det tidigare setts i vuxna djurstrukturer som hornhinnan49). Clytia sårläkning är extremt snabb, vilket har setts i andra system som saknar ett inflammatoriskt svar40,50. Läkning i Clytia-exparaplyet är helt beroende av rörelser hos de befintliga epitelcellerna - inga celler prolifererar eller migrerar genom ECM till sårplatsen (kompletterande film 1). Alla dessa fynd tyder på att Clytia är ett användbart modellsystem för att studera epitelial sårläkning. Faktum är att lättheten att avbilda epitelceller i Clytia under sårläkning ledde till upptäckten att epitelcellslamelipodi sträcker sig och sprids över områden med exponerad ECM så länge det finns ett intakt basalmembran; Om basalmembranet är skadat växlar epitelläkning till en handväska strängmekanism40. Detta var den första demonstrationen av en mekanism som ligger till grund för beslutet att stänga med lamellipodia-baserad krypning kontra handväska strängstängning, vilket belyser vikten av specifika cell: ECM-interaktioner vid läkning och att observera celler i deras naturliga sammanhang.
Nedan beskrivs protokoll för att skapa och avbilda encelliga mikrosår, små sår som stängs främst genom cellspridning och stora sår som kräver kollektiv cellmigration för att stängas. Vidare beskrivs ett protokoll för införande av små molekyler i ECM och epitelceller, vilket möjliggör experimentella störningar av förmodade regulatoriska vägar för sårläkning.
1. Djurkultur
- Underhålla Clytia-polypkolonier på mikroskopglas och medusae i artificiellt havsvatten (ASW) vid 18 °C i ett zebrafisksystem, med 2 L zebrafisktankar för polypkolonier och skräddarsydda 5 L pseudo-kreisel-tankar för medusae (kompletterande figur 1)51. ASW består av 4% Instant Ocean i avjoniserad (DI)H2O.
- Foder djuren dagligen med 2-3 dagar gammal artemia som beskrivs51.
OBS: Sårläkningsavbildning är lättare om djuren inte nyligen har matats, eftersom det finns mindre skräp som släpps ut från tarmen till synfältet. - Samla baby medusae från de etablerade polypkolonierna efter behov genom att placera kolonier i en 2 L bägare fylld med 1 L ASW över natten. Använd 2-3 veckor gamla kvinnliga medusae för alla sårläkningsexperiment. Förökning av Clytia har beskrivits i detalj någon annanstans51.
2. Sår
- Skapa mikrosår inom och mellan celler (20-500 μm2)
- Skapa en modifierad överföringspipett genom att klippa spetsen med sax för att göra en större öppning (0,5-0,7 cm diameter).
OBS: Öppningen i pipetten ska vara tillräckligt bred för att undvika skador på djuret. - Använd den modifierade överföringspipetten och placera medusa på en fördjupningsbild med medusa-exparaplyet uppåt, med tillräckligt med ASW för att täcka djuret.
- Placera omedelbart ett täckglas över djuret och bilden (se nedan för beskrivning av bildbehandling). Täckglaset komprimerar mesoglea, och rebound av den komprimerade vävnaden skapar en kraft som skjuter cellerna något isär52. Detta visas omedelbart som mellanrum mellan varje cell och skador i vissa celler (figur 1B, B ', figur 2 och figur 3A-C).
- Skapa en modifierad överföringspipett genom att klippa spetsen med sax för att göra en större öppning (0,5-0,7 cm diameter).
- Skapa små epitelsår (0,02-0,125 mm2)
- Använd en modifierad överföringspipett (som ovan), placera medusa på en fördjupningsbild med medusa-exparaplyet uppåt.
- Skrapa försiktigt ytan på medusan med en 200 μL pipettspets. Mjuka repor kan också skapa revor i källarmembranet, vilket är lätt uppenbart22. Täck djuret med ett täckglas för avbildning. Alternativt är placeringen av täckglaset ibland tillräckligt för att skapa små epitelsår även utan repor (figur 1C, C', figur 2 och figur 3A-C).
OBS: Tryck inte ner när du skrapar ytan på medusa, eftersom detta skadar ECM och skapar en oregelbunden yta - epitelceller som migrerar på en oregelbunden yta är svårare att hålla i fokus.
- Skapa stora epitelsår (0,5-0,9 mm2)
- Gör en mikroinjektionsnål med hjälp av en mikropipettavdragare och glaskapillärrör (steg 5.2). Placera den tomma mikroinjektionsnålen i en mikroinjektorhållare fäst vid en mikromanipulator. Skär nålspetsen så att öppningen är ca 20-40 μm.
OBS: Skurna nålar för stora epitelsår kan lagras och återanvändas för att öka konsistensen mellan experiment. - Ställ in hålltrycket på mikroinjektorn till noll och ställ in utmatningstrycket på cirka 20 PSI. Ställ in mikroinjektorn för att leverera en 2 s puls av luft.
OBS: Utmatningstrycket kan behöva justeras baserat på nålöppningens diameter (dvs. mindre spetsar använder högre tryck, medan större spetsar använder lägre tryck). - Placera medusan med exparaplyet uppåt på en fördjupningsbild på scenen i ett dissekeringsomfång, med precis tillräckligt med ASW för att täcka djuret. Använd mikromanipulatorn och justera mikroinjektionsnålspetsen så att den ligger precis ovanför vattnet. För att göra detta, doppa försiktigt spetsen i vatten (vatten kan komma in i pipettspetsen) och dra sedan tillbaka den så att den ligger nära medusas epitelyta.
OBS: Spetsen ska placeras över en kvadrant av medusa. Medusas radiella kanaler delar medusaklockan i fyra distinkta kvadranter. Inriktning på en kvadrant kommer att resultera i renare avbildning, eftersom gonaderna och de radiella kanalerna utesluts från sårområdet. - Pulsera luft genom att trycka på start på injektorn. Upprepa pulsen på samma plats två till fyra gånger, beroende på spetsens bredd. Större spetsar kräver färre pulser.
OBS: Ett streck i vattnet/medusa som orsakas av luftpulsen ska vara synligt. - Täck det skadade djuret med ett täckglas för avbildning av stora sår (figur 1D,D').
- Följ stegen nedan (avsnitt 3) för avbildning av epitelial sårläkning.
- Gör en mikroinjektionsnål med hjälp av en mikropipettavdragare och glaskapillärrör (steg 5.2). Placera den tomma mikroinjektionsnålen i en mikroinjektorhållare fäst vid en mikromanipulator. Skär nålspetsen så att öppningen är ca 20-40 μm.
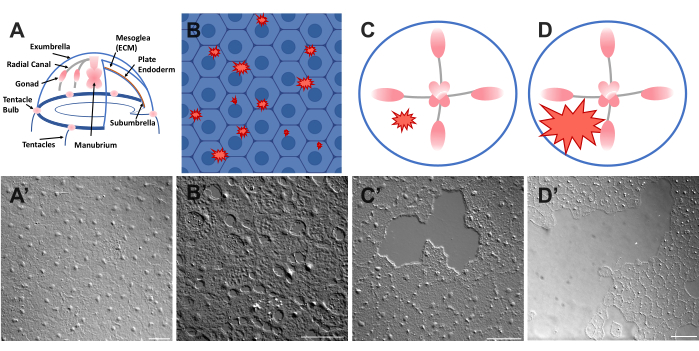
Figur 1: Intakt och sårad exparaplyepitelskikt i Clytia medusa. (A) Tecknad grafik av Clytia medusa-kroppen. (A') Intakt medusa exparaplyepitel sett ovanifrån. (B) Tecknad film av encelliga mikrosår (röda taggiga former) med epitelceller i blått. (B') Encellsmikrosår. (C) Tecknad film av ett litet epitelsår (röd taggig form). (C') Litet epitelsår. (D) Tecknad film av ett stort epitelsår (röd taggig form). (D') Stort epitelsår. Bilder erhölls alla med hjälp av DIC-mikroskopi. Skalstänger i (A'-C'): 50 μm. Skalstapel i (D'): 100 μm. Klicka här för att se en större version av denna figur.
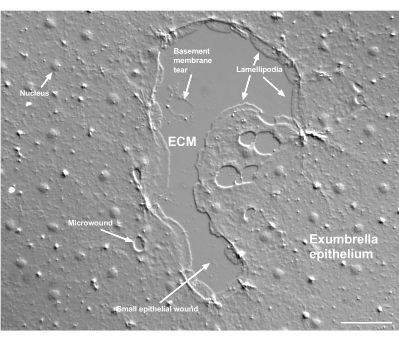
Figur 2: Flera sår och ett skadat basalmembran. Ett typiskt litet exparaplyepitelsår visas, med etiketter som indikerar lamellipodia som bildas från marginella celler. Dessutom ses mikrosår inom och mellan epitelceller. Notera den lilla basalmembranrivningen i den övre delen av såret. Film 4 visar läkning av detta sår. Skalstapel: 50 μm. Klicka här för att se en större version av denna figur.
3. Avbildning av epitelial sårläkning
- Se till att mikroskopet har justerats för Köhler-belysning53 och att det har ställts in korrekt för differentiell störande kontrast (DIC) mikroskopi54. Epitelceller är nästan osynliga med standardoptik (figur 3D, E).
- Justera fokus till exparaplyet. Även om detta är ett tunt lager, bör sexkantiga celler vara tydliga.
OBS: Exparaplyet och subparaplyet är åtskilda av en tjock mesoglea som stöds av vertikala fibrer. Subparaplycellerna är i samma fokalplan som de radiella kanalerna. Om du ursprungligen fokuserar på subparaplyskiktet, justera sedan fokus långsamt genom mesoglea och vertikala fibrer tills du hittar exparaplyet. - Identifiera ett sår till bilden manuellt. För stora sår, använd ett 10x mål. För mindre sår och encellssår, använd ett 20x mål.
- Starta ett program som samlar bilder som en film i realtid eller som samlar in en serie bilder med jämna mellanrum. Övervaka framstegen för att se till att sårområdet inte driver ut ur synfältet och att cellerna av intresse förblir i fokus.
- Encellssår stängs inom en minut; Föreställ dig därför deras stängning med en film.
- För att fånga detaljer om celldynamik för små sår, samla bilder ungefär var 10: e sekund. Stängning av små sår tar 20-50 min beroende på storlek.
- Avfyra inte de oförseglade objektglasen i mer än 45 minuter, eftersom avdunstning av vatten från glaset över tiden leder till djurdöd och bristning av cellerna.
- För längre observation, försegla runt täckglaset med vaselin för att minska avdunstningen.
OBS: Vissa medusa kan pulsera på bilden, vilket stör avbildningen. I detta fall fungerar montering av djur i en 1:10 utspädning av 1% etyl-3-aminobensoatmetansulfonat (trikain), justerat till pH 7,5, i ASW som ett effektivt bedövningsmedel och har ingen uppenbar effekt på läkning inom en tidsram på 1 timme. Djuren kommer dock att dö om de lämnas i flera timmar i Tricaine.

Figur 3: Skapa ett litet sår i exparaplyepitelet. (A) Försiktigt skrapande av exparaplyet med en 200 μL pipettspets för att skapa ett litet epitelsår. (B) Att placera täckglaset är ibland tillräckligt för att skapa små epitelsår. (C) Medusa monterad på en fördjupningsglas. (D ) Liten epitelial sårbild utan DIC-optik och (E) med DIC-optik. Skalstänger: 50 μm Klicka här för att se en större version av denna figur.
4. Bedömning
- Förbereda bildfiler
OBS: För att bearbeta bildfilerna, använd FIJI/ImageJ med uppdaterade BioFormat-plugins.- Ställ in skalan till rätt pixel per mikron-förhållande innan du registrerar bildstacken; Analysera > uppsättningsskala. Detta är nödvändigt för att extrahera faktiska storleksmätningar i nedströmsanalyser.
- Ofta driver djuret något på mikroskopglaset; därför, för att eliminera drift i filmer, registrera bilderna med FIJI-plugin linjär stackjustering med SIFT. Plugins > registrering > linjär stackjustering med SIFT.
- Spara den registrerade stacken som en .avi fil. Arkiv > Spara som > AVI ... I popup-fönstret ställer du in bildfrekvensen (animerade figurer här är inställda på 10 fps) och klickar på OK. Använd dessa utdata för att utföra sårläkningsanalys.
- Analys av sårområdet
- Använd lassoverktyget i FIJI/ImageJ och skissera såret genom att spåra cellkanterna. Mät sårområdet som just skisserades med Command + M eller CTRL + M.
- Upprepa sårområdesmätningen var 10: e bildruta. Mätningarna från FIJI/ImageJ kan sedan plottas med Prism 9 (figur 4).
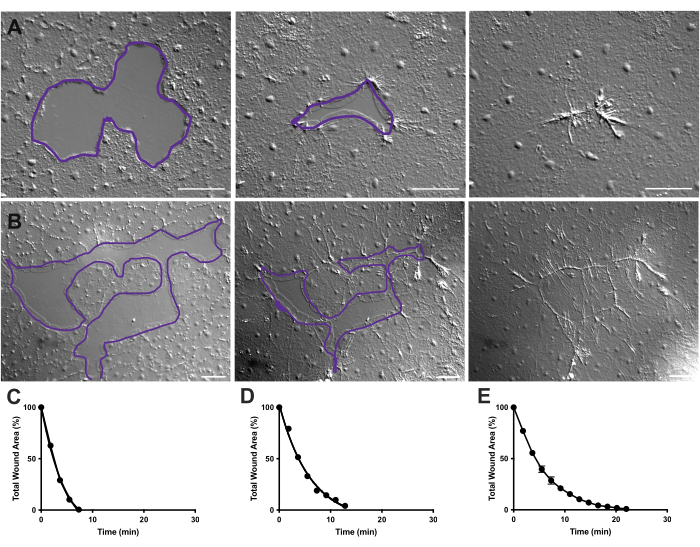
Figur 4: Analys av sårområdet i små epitelsår. (A ) Exempel på en liten epitelial sårläkning under 10 min. (B) Exempel på en annan epitelial sårläkning under 21 min. De lila konturerna i A,B är jämförbara med mätningarna av sårområden med lassoverktyget i FIJI/ImageJ. (C) Normaliserad minskning av sårområdet över tid i A. (D) Normaliserad minskning av sårområdet över tid i B. (E) Genomsnittlig minskning av sårområdet över tid för 14 små sår. n = 14. Felstaplar centrerade kring medelvärdet ± SEM. Skalstänger: 50 μm Klicka här för att se en större version av denna figur.
5. Mesogleala injektioner
- Skapa injektionsfat
- Bered polydimetylsiloxan (PDMS) genom att kombinera en PDMS-bas och härdningsmedel i ett förhållande av 10 viktdelar bas till 1 viktdel härdningsmedel. Rör om kraftigt för att blanda basen och härdningsmedlet helt.
- För att ta bort bubblor, sätt blandningen i en vakuumkammare i 15 minuter. Häll blandningen i en 60 mm petriskål med mikrocentrifugrörslock för att hålla formen på plats. Placera omedelbart formen på rörlocken med 45° lutning och tejpa på plats. Formen är tre staplade, förskjutna glasskivor limmade ihop för att skapa åsar i den slutliga injektionsskålen.
- Lägg hela formen, formen och blandningen i en ugn vid 60 ° C i 2 timmar för att härda elastomeren. Ta bort formen för en färdig injektionsskål.
- Mikropipett drar
- Använd en mikroelektrodavdragare och designa ett dragprogram. Använd ett enstegsprogram med hög hastighet. Värmen är ungefär glasets RAMP-testresultat55,56. Kontrollera resulterande mikropipetter för långa konsekventa avsmalningar.
OBS: Använd tunnväggiga borosilikatkapillärer med en 1,0 mm ytterdiameter, 0,75 mm innerdiameter och 10 cm längd.
- Använd en mikroelektrodavdragare och designa ett dragprogram. Använd ett enstegsprogram med hög hastighet. Värmen är ungefär glasets RAMP-testresultat55,56. Kontrollera resulterande mikropipetter för långa konsekventa avsmalningar.
- Injektion av färgämnen och droger
- Gör en mikroinjektionsnål (som ovan).
- Återfyll mikroinjektionsnålen med en lång pipettspets med en överflödig volym färgämne eller läkemedel för injektion i medusa.
OBS: För Clytia bör dimetylsulfoxid (DMSO) hållas vid en <1:100 utspädning med ASW, eftersom högre DMSO-koncentrationer hindrar sårläkning. Om du injicerar en klar lösning kan Fast Green FCF-lösning (1:100 spädning av 0,1% Fast Green FCF i ASW) tillsättas för att visualisera den injicerade vätskan. - Använd en modifierad överföringspipett enligt ovan, placera en medusa med subparaplyet uppåt i en PDMS-injektionsskål med precis tillräckligt med ASW för att täcka djuret (figur 5C). Placera skålen på scenen i ett dissekeringsomfång.
OBS: Begränsning av överskott av ASW förhindrar medusa från att simma i skålen och möjliggör mer framgångsrika injektioner. - Fokusera på mikroinjektionsnålspetsen och för den vidare i vattnet nära medusa. Tryck in nålen i skålen med mikromanipulatorn tills den böjer sig och går sönder. Denna spetsöppning är ungefär 10-20 μm.
OBS: Denna nål kan användas upprepade gånger för samma färgämne / läkemedelsinjektioner den dagen. Det rekommenderas att använda ett nytt tips varje dag och för separata färgämnen / droger. - Använd mikromanipulatorn och sätt in nålspetsen genom subparaplyet i mesoglea utan att punktera exparaplyet.
OBS: En veckning / vikning av epitelet kommer att märkas. När nålen sätts in i medusan upphör veckningen/vikningen. - På mikroinjektorn ställer du in hålltrycket på noll och utkastningstrycket på ≤20 PSI. Injicera i en eller två kvadranter, fyll var och en med en fläck av färgämne eller läkemedel ungefär 1/4 av området för den kvadranten.
OBS: Beroende på medusans storlek är större eller mindre volymer lämpliga i enstaka injektionsställen. Överfyllning av medusa resulterar i extrem skada på epitelet och till och med djurets död. - Beroende på vilket färgämne eller läkemedel som injiceras placeras djuren i en bägare med färsk ASW för att möjliggöra färg- eller läkemedelsdiffusion och inkubation.
- För avbildning, montera medusa på en fördjupningsglas med en modifierad överföringspipett, placera djuret så att exparaplyet är vänt uppåt (figur 5). Djur kan skadas i detta skede för att testa effekten av ett injicerat reagens.
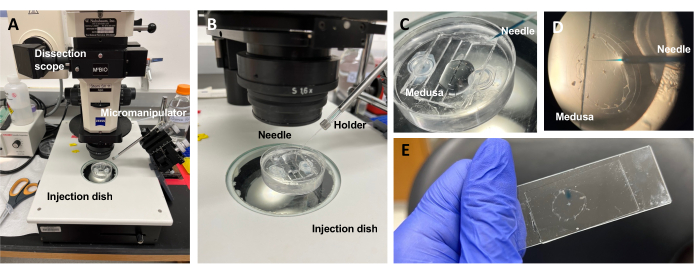
Figur 5: Injektionsinställning för införande av färgämnen eller läkemedel till ECM. (A) Uppställning för injektion. (B) Närbild av injektionsuppställningen som visar mikroinjektionsnålens orientering (ungefär 45° vinkel i förhållande till djuret i skålen). (C) Närbild av silikoninjektionsskålen med medusa i en liten mängd ASW för injektion. (D) En mikroinjektionsnål laddad med Fast Green FCF som kommer in i medusans mesoglea genom subparaplyet. (E) Efter injektion av Fast Green FCF i en monterad medusa. Klicka här för att se en större version av denna figur.
Efter protokollen ovan avbildades encelliga mikrosår, små sår och stora sår. Registrerade högar med bildfiler sparades som .avi filer.
I film 1 kan mikrosår ses stänga mellan och inuti celler (figur 1 och figur 2). Små lamellipodi observeras under stängning, följt av sammandragning och läkning. Skräp utesluts och släpps ut i vattnet. Läkning är klar på en minut eller mindre.
I film 2 och 3 läker små sår av olika former genom bildandet av lamellipodia, förlängning av lamellipodiala kontakter och spridning av celler vid sårmarginalen, som tidigare beskrivits40 (figur 1 och figur 2). Celler i nivåer bakom marginalcellerna deltar inte i läkning av sår av denna storlek och det finns inte heller kollektiv cellmigration. Snabb och progressiv stängning av epitelluckor följs av vävnadskontraktion längs den nybildade sårsömmen40. Den normaliserade läkningshastigheten för dessa två sår, uttryckt i procent av det ursprungliga området över tid, visas (figur 4C,D). Även om det finns en viss variation i dynamiken i sårförslutning, möjliggör medelvärdet av den procentuella stängningen över tid för 14 sår av olika former som sträcker sig från 0,02-0,125 mm2 upprättandet av en genomsnittlig kurva för sårläkning hos obehandlade djur (figur 4E).
Skador på basalmembranet kan tydligt ses när det inträffar (figur 2). I film 4 sprids celler i marginalen av ett litet sår där det finns skador på basalmembranet runt det skadade området, och gapstängningen kompletteras med en handväska strängkontraktion.
Om vävnaden är uttorkad eller för skadad för att repareras kan cellrörelserna stoppas, eller hela cellarket kan spricka (film 5 och film 6). Detta händer vanligtvis efter långa perioder av bildbehandling (45 minuter eller längre). Om cellsprängning inträffar tidigt i avbildningen kasseras provet.
Som visas i film 7 läker stora sår i flera steg. Först blir sårets kant slät och regelbunden på grund av sammandragningar vid marginalen, som tidigare rapporterats57. Därefter ses lamellipodi bildas från cellerna vid sårmarginalen, med lamellipodia som rör sig framåt för att maximera kontakten med intilliggande lamellipodia. Spårning av kärnorna i celler vid sårmarginalen och flera nivåer bakom marginalcellerna visar att stora luckor stängs av kollektiv cellmigration40. Celler lossnar aldrig utan rör sig tillsammans som ett ark.
Införandet av färgämnen och farmakologiska medel kan vara ett kraftfullt verktyg för att dissekera biologiska mekanismer. Många ämnen är uteslutna från Clytia (visas inte), troligen på grund av slemskiktet som täcker djurets yta. Mikroinjektion kan dock användas för att direkt införa molekyler i ECM, störa ECM-strukturen eller störa reglerande aktiviteter i ECM. Dessutom kan färgämnen och andra molekyler komma in i epitelceller från basalsidan. Till exempel visar figur 6 kärnfärgning med Hoechst, membranfärgning med FM1-43 och hämning av lamellipodibildning av cytochalasin B efter att dessa reagens mikroinjicerats i ECM. Införandet av dessa molekyler till ECM och epitelceller före sårning möjliggör experiment som testar effekten av farmakologiska verktyg på läkningsprocessen.
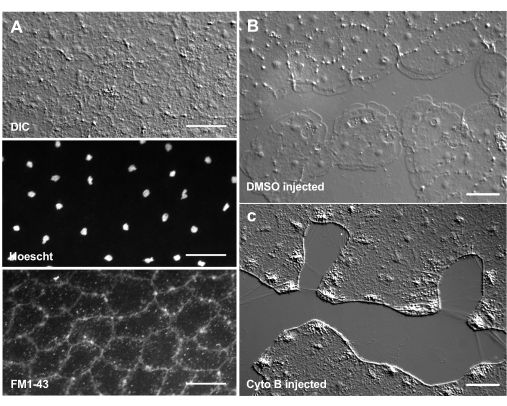
Figur 6: Epitelceller i medusa efter mikroinjektion av färgämnen eller farmakologiska medel. (A) Epitelceller visas i övre panelen 5 minuter efter injektion med 20 μM Hoechst (kärnor) och 50 μM FM1-43 (membran). (B,C) Sårläkning efter injektion med 1:1 000 DMSO-kontroll (B) eller 100 μM cytochalasin B (C). Sår gjordes 15 min efter injektionen. Bilder togs 5 min efter skadan. Bildningen av lamellipodi hämmas av cytochalasin B. De uppenbara "fibrerna" som ofta ses mellan celler i sårområdet tros vara resultatet av spänning som sträcker basalmembranet - de fläckar inte med falloidin (visas inte). Skalstänger: 50 μm. Klicka här för att se en större version av denna figur.
Film 1: Time-lapse-film av encellsläkning av mikrosår. Förfluten tid: 20 s. Bildhastighet: 10 fps. Skalstång: 50 μm. Klicka här för att ladda ner den här filmen.
Film 2: Time-lapse-film av en liten epitelial sårläkning. Förfluten tid: 9 min 54 s. Bildhastighet: 10 fps. Skalstång: 50 μm. Klicka här för att ladda ner den här filmen.
Film 3: Time-lapse-film av en liten epitelial sårläkning. Detta sår är större och mer oregelbundet format än såret i film 2. Förfluten tid: 20 min 54 s. Bildhastighet: 10 fps. Skalstång: 50 μm. Klicka här för att ladda ner den här filmen.
Film 4: Time-lapse film av ett litet sår och ett mikrosår som läker med en källarmembranbristning. Lamellipodia sprids runt basalmembranets rivning, även om de kan avancera över resten av ECM. När sårets område med källarmembranskadorna omges drar en handväska strängkontraktion celler över regionen. Förfluten tid: 19 min 4 s. Bildhastighet: 10 fps. Skalstång: 50 μm. Klicka här för att ladda ner den här filmen.
Film 5: Celler dör i ett litet epitelsår. Celldöd beror sannolikt på uttorkning av djuret. Förfluten tid: 4 min 24 s. Bildhastighet: 10 fps. Skalstapel: 100 μm. Klicka här för att ladda ner den här filmen.
Film 6: Ett litet epitelsår misslyckas med att slutföra läkning. Förfluten tid: 42 min 32 s. Bildhastighet: 10 fps. Skalstång: 50 μm. Klicka här för att ladda ner den här filmen.
Film 7: Stor epitelial sårläkning. Förfluten tid: 25 min 29 s. Bildhastighet: 10 fps. Skalstapel: 100 μm. Klicka här för att ladda ner den här filmen.
Kompletterande figur 1: Scheman för Clytia-tankens dimension. 3D-visualisering av de skräddarsydda Clytia-tankarna. (A) Fram- och bakifrån. (B) Vy från sidan. Utskärningen i stycket som visas i grönt är täckt med nylonnät. Vatten kommer in i tanken direkt över nätet, sveper över nätet och skapar en cirkulär ström. Vatten lämnar systemet genom hålet i ändstycket som visas i blått. Klicka här för att ladda ner den här filen.
Kompletterande film 1: Acellulär extracellulär matris i Clytia. Z-stack av Clytia tas med hjälp av konfokalmikroskopi. Stacken fokuserar initialt på exparaplyet och skannar sedan var 10: e μm genom ECM till plattans endoderm och subparaply. Bilder med DIC (vänster) och Hoechst nukleär färgning (höger) visar bristen på celler i ECM. Skalstång: 100 μm. Klicka här för att ladda ner den här filen.
Här presenteras metodiken för avbildning av sår in vivo i Clytia, en relativt ny modellorganism hos ryggradslösa djur40,43,58. Det finns flera faktorer som gör detta system till ett unikt och kraftfullt forskningsverktyg, som skiljer sig från andra modeller som används för att studera sårläkning och återepitelisering. Först är monolagerepitelet fäst vid en transparent ECM, vilket liknar vävnadsodlingsanalyser in vitro (figur 1, figur 2, figur 3, figur 4). Liksom i in vitro-analyser kan celler avbildas med hög upplösning. Men till skillnad från vävnadsodling finns det en autentisk cellulär miljö och ECM, så att sårläkning kan ses i samband med de komplexa signalhändelserna som inträffar i ett levande skadat djur. För det andra saknar Clytia inflammatoriska svar, migrerande fibroblaster, vaskulatur och blod. Detta gör att re-epiteliseringsprocessen kan studeras in vivo i frånvaro av de överlappande händelser som inträffar hos mer komplexa vuxna djur under sårläkning59. För det tredje är ECM acellulär (kompletterande film 1) och stor, vilket möjliggör enkel åtkomst med en mikroinjektionsnål (figur 5 och figur 6). Med hjälp av detta tillvägagångssätt kan forskare testa effekten av farmakologiska reagens som stör ECM-strukturen eller signaleringen på sårläkning in vivo. Reagenser kan också införas i epitelceller, och deras effekter på sårläkning in vivo kan bedömas. För det fjärde finns det protokoll som finns för att skapa mutanter och transgena djur i Clytia-systemet42,43,44,45. In vivo sårläkning kan därför observeras hos djur med ökat/minskat uttryck av gener av intresse.
Det finns flera kritiska steg i denna teknik. För det första, som visas i figur 3, är det nödvändigt att använda ett mikroskop som är korrekt konfigurerat för DIC-mikroskopi eftersom de platta, transparenta epitelcellerna är nästan osynliga med standard ljusmikroskopi. Det är också viktigt att utveckla förmågan att såra djur försiktigt så att epitelet skadas utan att mejsla ECM. En liknande mild beröring är nödvändig för mikroinjicering av material i ECM, eftersom omfattande skador på djuret under injektionen kan äventyra en efterföljande analys av sårläkning. Även om det finns en inlärningskurva för dessa tekniker, har även nybörjare behärskat dem snabbt i Malamy-labbet. Faktum är att dessa protokoll har använts för att demonstrera cellmigration i grundutbildningskurser vid University of Chicago.
För optimal avbildning är det viktigt att djuret inte rör sig och det valda sårområdet inte driver ut ur synfältet. Om djuren pulserar är behandling med trikain enligt beskrivningen mycket effektiv. För drifting är det ofta nödvändigt att manuellt flytta provet. Dessa rörelser kan elimineras från den slutliga filmen med hjälp av registreringsfunktionen i FIJI/ImageJ.
En begränsning med detta system är att det inte är möjligt att skapa identiska sår, eftersom sår varierar i både form och storlek med de metoder som beskrivs här. Därför kan det vara svårt att kvantifiera den exakta hastigheten för sårförslutning eller cellmigration. Positionsmarkörer som kolkorn fastnar på den exponerade ECM i ett sårat djur och kan användas för att mäta hastigheten för kollektiv cellmigration i stora sår (visas inte). För analys av små sårförslutningar, även med varierande sårstorlek och form, finns det ett begränsat intervall av förslutningshastigheter bland sår av denna storlek (figur 4). Det är därför möjligt att kvantitativt påvisa effekterna av främjande eller repressiva farmakologiska reagenser.
Medan detta arbete beskriver karakteriseringen av sårläkning med endast DIC-mikroskopi, kan samma tillvägagångssätt användas för bildläkning med fluorescens eller konfokalmikroskopi. För att underlätta detta finns protokoll för att generera transgena djur där olika cellulära och extracellulära proteiner är fluorescerande märkta. Samtidig avbildning med DIC och fluorescens, i kombination med störning av sårläkning med hjälp av farmakologiska medel eller mutantlinjer, kommer att vara ett kraftfullt tillvägagångssätt för att förstå mekanismer som ligger till grund för sårläkningsprocessen i epitelet.
Inget att avslöja.
E.E.L.L. stöds av ett bidrag från National Science Foundation PRFB 2011010. Vi vill tacka Tsuyoshi Momose och Evelyn Houliston för att hjälpa oss att etablera våra Clytia-kolonier, Jean-Baptiste Reynier för insamling av mikrosårläkningsbilder, Harry Kyriazes för byggandet av pseudo-kreisel-tankarna och Elizabeth Baldo för att upprätthålla Clytia-livsmiljön. Figur 1B skapades med BioRender.com.
| Name | Company | Catalog Number | Comments |
| 20500 ACE EKE Microscope Fiber Optic Light Source | Kramer Scientific Corporation | ||
| AxioCam 506 mono | ZEISS | 426557-0000-000-MA285 | |
| Capillary tubes | World Precision Instruments | TW1004 | |
| Cytochalasin B | Abcam | ab143482 | |
| Depression slides | Amscope | BS-C12 | |
| DMR with DIC options and fluorescence halogen lamp | Leica | ||
| Ethyl 3-aminobenzoate methanesulfonate | Sigma Aldrich | E10521-10G | |
| Fast Green FCF | Thermo Scientific | A16520-06 | |
| FM1-43 | Biotium | 70022 | Excitation/Emission: 480/598 nm |
| Hoechst 33342 | Thermo Scientific | 62249 | Excitation/Emission: 361/497 nm |
| imageJ | NIH | ||
| Microloader tips (0.5-10 μL /2-20 μL) | Eppendorf | 930001007 | |
| Micromanipulator | World Precision Instruments | 3301R / M3301L | |
| Microscope Cover Glass (22X40-1.5) | Fisherbrand | 12-544-BP | |
| Petri Dish (60 mm x 15 mm) | Fisherbrand | FB085713A | |
| PicoNozzle v2 | World Precision Instruments | 5430-ALL | |
| Pipette puller | Sutter Instrument Co | P-97 | |
| Pneumatic PicoPump | World Precision Instruments | PV820 | |
| Polycarbonate vacuum, desiccator | Bel-art | F42025-0000 | |
| Prism 9 | GraphPad | ||
| STEMI Sv11 Dissection scope | ZEISS | STEMI SV11 | |
| SYLGARD 184 | Dow Silicones | 1024001 | |
| Transfer pipettes | Fisherbrand | 13-711-7M | |
| Z-Hab mini system | Pentair | ||
| ZEN Microscopy software | Zeiss |
- Tyler, S. Epithelium-the primary building block for metazoan complexity. Integrative and Comparative Biology. 43 (1), 55-63 (2003).
- Kurn, H., Daly, D. T. Histology, Epithelial Cell. StatPearls. , (2022).
- Schempp, C., Emde, M., Wölfle, U. Dermatology in the Darwin anniversary. Part 1: Evolution of the integument. Journal of the German Society of Dermatology. 7 (9), 750-757 (2009).
- Lopez-Ojeda, W., Pandey, A., Alhajj, M., Oakley, A. M. Anatomy, Skin (Integument). StatPearls. , (2022).
- Bukowiecki, A., Hos, D., Cursiefen, C., Eming, S. A. Wound-healing studies in cornea and skin: parallels, differences and opportunities. International Journal of Molecular Sciences. 18 (6), 1257 (2017).
- Frantz, C., Stewart, K. M., Weaver, V. M. The extracellular matrix at a glance. Journal of Cell Science. 123 (24), 4195-4200 (2010).
- Hynes, R. O. The evolution of metazoan extracellular matrix. The Journal of Cell Biology. 196 (6), 671-679 (2012).
- Fidler, A. L., et al. Collagen IV and basement membrane at the evolutionary dawn of metazoan tissues. eLife. 6, 24176 (2017).
- Bullen, T. F., et al. Characterization of epithelial cell shedding from human small intestine. Laboratory Investigation; a Journal of Technical Methods and Pathology. 86 (10), 1052-1063 (2006).
- Watson, A. J. M., et al. Epithelial barrier function in vivo is sustained despite gaps in epithelial layers. Gastroenterology. 129 (3), 902-912 (2005).
- Sonnemann, K. J., Bement, W. M. Wound repair: toward understanding and integration of single-cell and multicellular wound responses. Annual Review of Cell and Developmental Biology. 27, 237-263 (2011).
- Abreu-Blanco, M. T., Verboon, J. M., Parkhurst, S. M. Single cell wound repair. BioArchitecture. 1 (3), 114-121 (2011).
- Fenteany, G., Janmey, P. A., Stossel, T. P. Signaling pathways and cell mechanics involved in wound closure by epithelial cell sheets. Current Biology. 10 (14), 831-838 (2000).
- Pastar, I., et al. Epithelialization in wound healing: a comprehensive review. Advances in Wound Care. 3 (7), 445-464 (2014).
- Rousselle, P., Braye, F., Dayan, G. Re-epithelialization of adult skin wounds: Cellular mechanisms and therapeutic strategies. Advanced Drug Delivery Reviews. 146, 344-365 (2019).
- Bement, W. M., Forscher, P., Mooseker, M. S. A novel cytoskeletal structure involved in purse string wound closure and cell polarity maintenance. The Journal of Cell Biology. 121 (3), 565-578 (1993).
- Vedula, S. R. K., Ravasio, A., Lim, C. T., Ladoux, B. Collective Cell migration: a mechanistic perspective. Physiology. 28 (6), 370-379 (2013).
- Li, L., He, Y., Zhao, M., Jiang, J. Collective cell migration: Implications for wound healing and cancer invasion. Burns & Trauma. 1 (1), 21-26 (2015).
- Bornes, L., Windoffer, R., Leube, R. E., Morgner, J., van Rheenen, J. Scratch-induced partial skin wounds re-epithelialize by sheets of independently migrating keratinocytes. Life Science Alliance. 4 (1), 202000765 (2021).
- Theveneau, E., Mayor, R. Collective cell migration of epithelial and mesenchymal cells. Cellular and Molecular Life Sciences. 70 (19), 3481-3492 (2013).
- Haensel, D., Dai, X. Epithelial-to-mesenchymal transition in cutaneous wound healing: where we are and where we are heading. Developmental Dynamics. 247 (3), 473-480 (2018).
- Cordeiro, J. V., Jacinto, A. The role of transcription-independent damage signals in the initiation of epithelial wound healing. Nature Reviews. Molecular Cell Biology. 14 (4), 249-262 (2013).
- Abreu-Blanco, M. T., Watts, J. J., Verboon, J. M., Parkhurst, S. M. Cytoskeleton responses in wound repair. Cellular and Molecular Life Sciences. 69 (15), 2469-2483 (2012).
- Klarlund, J. K., Block, E. R. Free edges in epithelia as cues for motility. Cell Adhesion & Migration. 5 (2), 106-110 (2011).
- Enyedi, B., Niethammer, P. Mechanisms of epithelial wound detection. Trends in Cell Biology. 25 (7), 398-407 (2015).
- Niethammer, P. The early wound signals. Current Opinion in Genetics & Development. 40, 17-22 (2016).
- Jacinto, A., Martinez-Arias, A., Martin, P. Mechanisms of epithelial fusion and repair. Nature Cell Biology. 3 (5), 117-123 (2001).
- Mayor, R., Etienne-Manneville, S. The front and rear of collective cell migration. Nature reviews. Molecular Cell Biology. 17 (2), 97-109 (2016).
- Gupta, S., Yap, A. S. How adherens junctions move cells during collective migration. Faculty Reviews. 10, 56 (2021).
- Blair, M. J., Jones, J. D., Woessner, A. E., Quinn, K. P. Skin structure-function relationships and the wound healing response to intrinsic aging. Advances in Wound Care. 9 (3), 127-143 (2020).
- Falanga, V., et al. Chronic wounds. Nature Reviews. Disease Primers. 8 (1), 50 (2022).
- Liang, C. -. C., Park, A. Y., Guan, J. -. L. In vitro scratch assay: a convenient and inexpensive method for analysis of cell migration in vitro. Nature Protocols. 2 (2), 329-333 (2007).
- Jonkman, J. E. N., et al. An introduction to the wound healing assay using live-cell microscopy. Cell Adhesion & Migration. 8 (5), 440-451 (2014).
- Poujade, M., et al. Collective migration of an epithelial monolayer in response to a model wound. Proceedings of the National Academy of Sciences. 104 (41), 15988-15993 (2007).
- Masson-Meyers, D. S., et al. Experimental models and methods for cutaneous wound healing assessment. International Journal of Experimental Pathology. 101 (1-2), 21-37 (2020).
- Park, S., et al. Tissue-scale coordination of cellular behaviour promotes epidermal wound repair in live mice. Nature Cell Biology. 19 (2), 155-163 (2017).
- Tsai, C. -. R., Wang, Y., Galko, M. J. Crawling wounded: molecular genetic insights into wound healing from Drosophila larvae. The International Journal of Developmental Biology. 62 (6-7-8), 479-489 (2018).
- Richardson, R., et al. Adult zebrafish as a model system for cutaneous wound-healing research. The Journal of Investigative Dermatology. 133 (6), 1655-1665 (2013).
- Erickson, J. R., Echeverri, K. Learning from regeneration research organisms: The circuitous road to scar free wound healing. Developmental Biology. 433 (2), 144-154 (2018).
- Kamran, Z., et al. In vivo imaging of epithelial wound healing in the cnidarian Clytia hemisphaerica demonstrates early evolution of purse string and cell crawling closure mechanisms. BMC Developmental Biology. 17 (1), 17 (2017).
- Chari, T., et al. Whole-animal multiplexed single-cell RNA-seq reveals transcriptional shifts across Clytia medusa cell types. Science Advances. 7 (48), (2021).
- Weissbourd, B., et al. A genetically tractable jellyfish model for systems and evolutionary neuroscience. Cell. 184 (24), 5854-5868 (2021).
- Momose, T., et al. High doses of CRISPR/Cas9 ribonucleoprotein efficiently induce gene knockout with low mosaicism in the hydrozoan Clytia hemisphaerica through microhomology-mediated deletion. Scientific Reports. 8 (1), 11734 (2018).
- Houliston, E., Leclère, L., Munro, C., Copley, R. R., Momose, T. Past, present and future of Clytia hemisphaerica as a laboratory jellyfish. Current Topics in Developmental Biology. 147, 121-151 (2022).
- Schmid, V., et al. The extracellular matrix (mesoglea) of hydrozoan jellyfish and its ability to support cell adhesion and spreading. Hydrobiologia. 216 (1), 3-10 (1991).
- Day, R. M., Lenhoff, H. M. Hydra mesoglea: a model for investigating epithelial cell-basement membrane interactions. Science. 211 (4479), 291-294 (1981).
- Zhang, X., et al. The collagens of hydra provide insight into the evolution of metazoan extracellular matrices. The Journal of Biological Chemistry. 282 (9), 6792-6802 (2007).
- Danjo, Y., Gipson, I. K. Actin 'purse string' filaments are anchored by E-cadherin-mediated adherens junctions at the leading edge of the epithelial wound, providing coordinated cell movement. Journal of Cell Science. 111 (22), 3323-3332 (1998).
- Arenas Gómez, C. M., Sabin, K. Z., Echeverri, K. Wound healing across the animal kingdom: Crosstalk between the immune system and the extracellular matrix. Developmental Dynamics. 249 (7), 834-846 (2020).
- Lechable, M., et al. An improved whole life cycle culture protocol for the hydrozoan genetic model Clytia hemisphaerica. Biology Open. 9 (11), (2020).
- Casares, L., et al. Hydraulic fracture during epithelial stretching. Nature Materials. 14 (3), 343-351 (2015).
- Wayne, R. Chapter 4 - Bright-Field Microscopy. Light and Video Microscopy (Third Edition). , 95-116 (2019).
- Murphy, D. B., Davidson, M. W. . Fundamentals of Light Microscopy and Electronic Imaging: Second Edition. , (2012).
- . Micropipette Techniques for Electrophysiology Available from: https://www.sutter.com/micropipette/cookbook.html (2022)
- Brown, A. L., Johnson, B. E., Goodman, M. B. Making patch-pipettes and sharp electrodes with a programmable puller). Journal of Visualized Experiments. (20), e939 (2008).
- Klarlund, J. K. Dual modes of motility at the leading edge of migrating epithelial cell sheets. Proceedings of the National Academy of Sciences. 109 (39), 15799-15804 (2012).
- Houliston, E., Momose, T., Manuel, M. Clytia hemisphaerica: a jellyfish cousin joins the laboratory. Trends in Genetics. 26 (4), 159-167 (2010).
- Rodrigues, M., Kosaric, N., Bonham, C. A., Gurtner, G. C. Wound healing: a cellular perspective. Physiological Reviews. 99 (1), 665-706 (2019).
ABOUT JoVE
Copyright © 2024 MyJoVE Corporation. All rights reserved