Dioscin תיווך IgA נפרופתיה הקלה על ידי עיכוב הפעלת תאי B in vivo והפחתת ייצור IgA1 חסר גלקטוז במבחנה
In This Article
Summary
מחקר זה מספק נתונים ניסיוניים לטיפול באימונוגלובולין A נפרופתיה (IgAN) עם Dioscin (DIO), המרכיב הפעיל של Dioscoreae Nipponicae Rhizoma (DNR), ופרדיגמה לחקר ההשפעות של רפואת צמחים והמנגנונים הבסיסיים in vivo ו - in vitro.
Abstract
העלייה של IgA1 חסר גלקטוז במחזור הדם (Gd-IgA1) נגרמת על ידי הפעלה מוגזמת של תאי הפרשה חיוביים IgA בתהליך של תגובות חיסוניות ריריות, המהווה חוליה קריטית בפתוגנזה של נפרופתיה IgA (IgAN). המדבקה של פייר, המקום הבולט שבו לימפוציטים מסוג B הופכים לתאי פלזמה מפרישי IgA, היא המקור העיקרי של IgA. בנוסף, הביטוי הנמוך יותר של ליבה 1β-1,3-גלקטוסילטרנספראז (C1GalT1) והמלווה המולקולרי שלה, מלווה מולקולרי ספציפי C1GalT1 (Cosmc), קשור לגליקוזילציה חריגה של IgA1 בחולי IgAN. הניסיון הקליני שלנו מראה כי רפואת הצמחים של Dioscoreae Nipponicae Rhizoma (DNR) יכולה להקל על פרוטאינוריה והמטוריה ולשפר את תפקוד הכליות בחולי IgAN. Dioscin (DIO) הוא אחד המרכיבים הפעילים העיקריים של DNR, אשר יש פעילויות פרמקולוגיות שונות. מחקר זה בוחן את המנגנון האפשרי של DIO בטיפול IgAN.עכבר מודל IgAN הוקם על ידי השראת חיסון רירית. העכברים חולקו לקבוצות הבקרה, המודל וה-DIO gavage. שקיעת IgA גלומרולרית בעכברים, שינויים פתולוגיים כלייתיים וסמנים של תאי B CD20 וביטוי CXCR5 במדבקה של פייר זוהו על ידי אימונופלואורסנציה ואימונוהיסטוכימיה. לאחר גירוי ליפופוליסכריד (LPS), ההשפעות של DIO על התפשטות תאי DAKIKI, הפרשת IgA ו- Gd-IgA1, C1GalT1 וביטוי Cosmc נחקרו על ידי בדיקת ספירת תאים kit-8 (CCK-8), בדיקת אימונוסורבנט מקושרת אנזימים (ELISA), תגובת שרשרת פולימראז כמותית בזמן אמת (QRT-PCR) וכתמים מערביים (WB). במחקרי in vivo , שקיעת IgA מלווה בהיפרפלזיה מסנגיאלית גלומרולרית וביטוי מוגבר של CD20 ו- CXCR5 במדבקת פייר בעכבר מודל IgAN הוקלה על ידי DIO. מחקרי מבחנה הראו 0.25 מיקרוגרם/מ"ל עד 1.0 מיקרוגרם/מ"ל DIO עיכב את התפשטות תאי DAKIKI המושרה על ידי LPS, IgA ו-Gd-IgA1, וויסות מוגבר של mRNA וביטוי חלבונים של C1GalT1 ו-Cosmc. מחקר זה מדגים כי DIO עשוי להפחית את ייצור Gd-IgA1 על ידי עיכוב הפעלה מוגזמת של תאים מפרישי IgA וויסות ביטוי C1GALT1/Cosmc.
Introduction
IgA Nephropathy (IgAN) הוא הסוג הנפוץ ביותר של glomerulonephritis ראשוני, אשר אין טיפול ספציפי, והוא נשאר גורם משמעותי של מחלת כליות סופנית1. למרות שהפתוגנזה של IgAN עדיין אינה מובנת במלואה, "השערת ריבוי הפגיעות" מקובלת בדרך כלל ונתמכת על ידי גוף גדול של ראיות מחקריות קליניות וניסוייות2. הפתוגנזה של IgAN כרוכה בהפעלת תאי B וייצור IgA1 חסר גלקטוז (Gd-IgA1)3. העלייה במחזור Gd-IgA1 עקב התרבות יתר והפעלה של תאים מפרישי IgA במהלך התגובה החיסונית הרירית היא חוליה קריטית בפתוגנזה של IgAN 4,5,6. כמקום המרכזי להתרבות והפעלה של המרת פנוטיפ לימפוציטים מסוג B לתאים מפרישי IgA, המדבקה של פייר היא המקור העיקרי להפרשת IgA, הקשורה קשר הדוק להתרחשות והתפתחות של IgAN 7,8. בנוסף, התפשטות תאים מפרישי IgA1, כמו גם ביטוי של Core 1β-1,3-galactosyltransferase(C1GalT1) ומלווה מולקולרי ספציפי C1GalT1 (Cosmc), היו קשורים לגליקוזילציה חריגה של IgA1, אשר גורמת לייצור GD-IgA1 בחולי IgAN 6,9.
המחקר הקליני על טיפול IgAN בצמחי מרפא התקדם בשנים האחרונות. נוסחת Yiqi Qingjie היא נוסחה חיונית לטיפול IgAN על ידי המחלקה לנפרולוגיה של בית החולים גואנג'נמן. המחקר הקודם של הקבוצה שלנו מצא כי Gd-IgA1 ירד בסרום של חולי IgAN לאחר הטיפול בנוסחת Yiqi Qingjie. כאחד מעשבי התיבול הנפוצים ביותר בנוסחת Yiqi Qingjie, Dioscoreae Nipponicae Rhizoma (DNR) הוא קנה השורש המיובש של Dioscorea Nipponica Makino, שיש לו תפקידים שונים כגון ויסות חסינות, דיכוי דלקת, הקלה על שיעול ואסתמה10,11. מספר חוקרים טיפלו ב- IgAN עם DNR והשיגו תוצאות טובות 12,13,14. כמרכיב הפעיל העיקרי ב- DNR15, Dioscin (DIO) מוריד חומצת שתן, מעכב פיברוזיס, מעכב תגובה דלקתית ועקה נוגדת חמצון16,17. לכן, DIO עשוי להיות בעל מנגנון פעולה חדשני כדי לעכב את הפרשת התאים של Gd-IgA1 מופרז ולהפעיל השפעות ספציפיות להגנה על הכליות. עם זאת, לא דווח על מחקר על מנגנון הפעולה של DIO לטיפול ב- IgAN.
כדי לחקור את המנגנון הטיפולי הפוטנציאלי של DIO על IgAN ולספק שיטה חדשה לטיפול ב- IgAN, ערכנו ניסויים להשפעות הטיפוליות של DIO על IgAN in vivo ו - in vitro.
Protocol
ועדת האתיקה של בית החולים גואנגנמן אישרה את הניסוי הזה (מספר אישור אתי לניסויים בבעלי חיים: IACUC-GAMH-2023-003).
1. הכנת עכברים להליך הניסוי
- יש לגדל 22 עכברי Balb/c זכרים בדרגת SPF (בני 6-7 שבועות, משקל גוף 20-25 גרם) במתקן בעלי החיים של בית החולים/מרכז המחקר. חלק את בעלי החיים לקבוצות בקרה (n = 8) ומודל (n = 14) באמצעות שיטת טבלת המספרים האקראיים.
- לאחר שבוע אחד של גידול אדפטיבי בכלוב המעבדה, יש להאכיל את קבוצת המודל (קבוצת IgAN) בתמיסת גמא גלובולין (BGG) של 0.1% בקר במים חומציים המכילים 6 mmol / L HCl למשך 9 שבועות על פי פרוטוקול המידול של Zou et al.18.
- הזריקו 0.1 מ"ל של תמיסת BGG 0.1% במי מלח לווריד הזנב במשך 3 ימים רצופים תוך המשך שתיית תמיסת BGG להכנת עכבר ניסויי IgAN מודל18.
- תן לקבוצת הביקורת לשתות בחופשיות 6 mmol / L HCl acidified מים ללא BGG במשך 9 שבועות. להזריק את נפח המלח המתאים לתוך וריד הזנב במשך 3 ימים רצופים.
הערה: קבוצות הבקרה והמודלים הוזנו באותה איכות כמו הזנה רגילה. - לאחר הזרקת וריד הזנב, בחר 2 עכברים בקבוצת הביקורת ו -2 עכברים בקבוצת המודל באופן אקראי ובחן אותם על ידי פרוטאינוריה, מיקרוסקופ אור ואימונופלואורסנציה כדי לקבוע אם המידול היה מוצלח.
הערה: לא מסופק מזון לבעלי החיים, אך הם אינם אסורים במים; רשום את תפוקת השתן. - לאסוף שתן במשך 24 שעות על ידי כלובים מטבוליים צנטריפוגה אותו ב 400 x גרם במשך 5 דקות; יש להשליך משקעי שתן. לאחר דילול פי 10 של הסופרנטנט, יש למדוד את ריכוז הפרוטאינוריה באמצעות ערכת בדיקת חלבון בשתן ולאחר מכן להכפיל לפי גורם הדילול ונפח השתן כדי לקבל 24 שעות של חלבון שתן כולל.
הערה: שיטות מיקרוסקופיה ואימונופלואורסנציה מוצגות בסעיפים 3 ו-4, בהתאמה. - לאחר הכנת המודל בהצלחה, חלקו 12 עכברים בקבוצת המודל ל-6 עכברים, כל אחד בקבוצת המודל (קבוצת IgAN) ובקבוצת DIO gavage (קבוצת DIO), בהתאם לשיטת טבלת המספרים האקראיים.
- תן לקבוצת הביקורת להמשיך לשתות 6 mmol / L HCl מים חומציים ללא BGG, ולקבוצת המודל 0.1% תמיסת BGG המורכבת ממים חומציים המכילים 6 mmol / L HCl. חשב את המינון של ניהול gavage קבוצת DIO על פי נוסחת המרת המינון של מתודולוגיית ניסוי פרמקולוגית (שהומר על פי מסת גוף האדם של 70 ק"ג)19. טבליות Gavage DIO 0.06 גרם/ק"ג פעם ביום במשך 8 שבועות.
- לאחר 8 שבועות של gavage, להרדים את העכברים intraperitoneally עם 0.4% נתרן pentobarbital (60 מ"ג / ק"ג), ולאחר אישור הרדמה נכונה על ידי צביטת הבוהן, לבודד כליות ואת המדבקה של פייר עבור מיקרוסקופ אור הבא וניתוח immunohistochemistry.
הערה: התוכנית עבור מודל in vivo נמצאת באיור משלים 1.
2. ניתוח היסטולוגי
- קטעי פרפין לכליות ומדבקת פייר
- לתקן רקמות כליות בעובי 3 מ"מ או 1 מדבקת פייר עם 4% paraformaldehyde במשך 24 שעות, להתייבש עם אתנול הדרגתי וקסילן. טובלים בשעווה למשך שעתיים, אוטמים ומקפיאים.
- חותכים קטעי כליות בעובי 2 מיקרומטר ובמקטעי טלאי פייר בעובי 4 מיקרומטר ומורחים אותם במים חמים. גרפו את הפרוסות שנפרשו במגלשת זכוכית נקייה ואופים אותן בתנור בטמפרטורה קבועה של 40°C למשך שעה. התחל להכתים לאחר עיבוד מקדים של הדגימה.
הערה: קח את פני העטרה של החלק hilar של הכליה, עם עובי של בלוק רקמות 3 מ"מ.
- מטילים ומכתימים את חלקי הפרפין בטמפרטורת החדר (RT) למשך 10 דקות בתמיסת חומצה תקופתית, תוך הימנעות מאור. יש לשטוף במים מזוקקים ולנגב את הכתמים היבשים בתמיסת הצביעה של שיף למשך 20-30 דקות, תוך הימנעות מאור. יש לשטוף במים מזוקקים עד שהחלקים יהיו אדומים מתחת למיקרוסקופ.
- מניחים את החלקים בתמיסת צביעת המטוקסילין, גרעיני כתמים למשך 3 דקות (גרעינים מוכתמים עמוק מדי יכולים להיות מחולקים על ידי אתנול הידרוכלוריד), ושוטפים במים זורמים עד שהשקופיות חסרות צבע.
- בצע התייבשות שגרתית עם ריכוזים הדרגתיים של אתנול וקסילן, לאטום את החלקים עם מסטיק נייטרלי, ולצפות תחת מיקרוסקופ. PAS-חיובי הוא אדום, והגרעין כחול.
3. ניתוח אימונוהיסטוכימי של מדבקת פייר
- הכינו חלקי פרפין של מדבקת פייר כמתואר בשלב 2.1.
הערה: עובי החלקים עבור Immunohistochemical ו immunofluorescence לאחר מכן הוא 4 מיקרומטר. - Dewax את קטעי פרפין:
- הכניסו את החלקים לקסילן I למשך 5 דקות, לקסילן II למשך 5 דקות ולקסילן III למשך 5 דקות.
- יתר על כן, לשטוף את שקופיות אתנול נטול מים I במשך 5 דקות, אתנול נטול מים II במשך 5 דקות, 85% אלכוהול במשך 5 דקות, 75% אלכוהול במשך 5 דקות. לאחר מכן שטפו את המגלשות במים מזוקקים.
- שליפת אנטיגן
- הכינו תמיסת ציר 50x של נתרן ציטראט ודללו במים מזוקקים עד פי 1 לשימוש. מחממים אותו באוטוקלאבה במשך 2 דקות, ואז מניחים את הפרוסות באוטוקלאב, ומבטיחים שרמת הנוזל עולה על רמת הפרוסות.
- חממו בטמפרטורה גבוהה במשך 5 דקות, ואז אפשרו למגלשות להתקרר באופן טבעי. שטפו את הפרוסות שלוש פעמים בתמיסת PBS במשך 5 דקות כל אחת.
- חסימה של peroxidase אנדוגני: לסמן את גבולות הרקמה במעגל עם עט immunohistochemical. דגרו על המקטעים בתמיסת מי חמצן 3% למשך 15 דקות ב-RT, מוגנים מפני אור, ושטפו את המקטעים שלוש פעמים בתמיסת PBS למשך 5 דקות בכל פעם.
- חסימת סרום: חסמו את המקטעים על ידי הטלת סרום עיזים 10% על חלקי הרקמה המסומנים למשך 30 דקות ב-RT. ודאו שהחלקים מכוסים באופן שווה בכתם.
- דגירה ראשונית של נוגדנים: נערו בעדינות את התמיסה החוסמת והוסיפו חלק מהנוגדן הראשוני המוכן (CD20 [1:800]; CXCR5 [1:800]) לסעיף. מניחים את החלק שטוח בקופסה רטובה ודגורים לילה על 4 מעלות צלזיוס.
הערה: יש להוסיף כמות קטנה של מים לקופסה הרטובה כדי למנוע אידוי של הנוגדן. - דגירה משנית של נוגדנים: שטפו את המקטעים שלוש פעמים בתמיסת PBS למשך 5 דקות בכל פעם. הסר PBS על ידי ניעור המקטעים יבש, כסה את הרקמה בטיפת נוגדן משני (HRP-label) של הזנים הקשורים של הנוגדן הראשוני, ודגר ב- RT למשך 50 דקות.
- הומוגניזציה של 3,3'-diaminobenzidine (DAB): שטפו את המקטעים בתמיסת PBS שלוש פעמים במשך 5 דקות כל אחד. לאחר ניעור החלקים יבשים, זרוק את התמיסה הכרומוגנית DAB שזה עתה הוכנה על המקטעים. שימו לב לזמן התפתחות הצבע מתחת למיקרוסקופ; החיובי הוא חום-צהוב. יש לשטוף במי ברז כדי לסיים את התפתחות הצבע.
- גרעיני צביעה: יש להכתים מחדש עם המטוקסילין למשך כדקה, לשטוף במי ברז ולאחר מכן לשטוף במשך 10 דקות במי ברז כדי לחזור לכחול.
- התייבשות ואיטום:
- הכניסו את החלקים ל-75% אלכוהול למשך 5 דקות ו-85% אלכוהול למשך 5 דקות. מניחים את החלקים באתנול נטול מים I למשך 5 דקות, אתנול נטול מים II למשך 5 דקות, ואתנול III נטול מים למשך 5 דקות.
- שוטפים את החלקים בקסילן I למשך 5 דקות, מוציאים אותם לייבוש קל ואוטמים את החלקים במסטיק ניטרלי.
- רכישת תמונות: איסוף תמונות על ידי בדיקה מיקרוסקופית וניתוח על ידי תוכנת הילה לניתוח תמונה פנורמית של הרקמה.
הערה: גרעינים המוכתמים בהמטוקסילין הם כחולים, וביטוי חיובי DAB נצפה כחום-צהוב.
4. כליות IgA immunofluorescence
- הכן קטעי פרפין לכליות כמתואר בשלב 2.1.
- קטעי פרפין Dewax:
- הכניסו את החלקים לקסילן I למשך 5 דקות, לקסילן II למשך 5 דקות ולקסילן III למשך 5 דקות. יש לטפל בחלקים באתנול נטול מים I, אתנול נטול מים II, 95% אתנול, 90% אתנול, 80% אתנול, 70% אתנול ו-50% אתנול, כל אחד למשך 5 דקות, ולשטוף במים מזוקקים.
- שליפת פרוטאינאז K: נערו את המקטעים יבשים וציירו עיגול סביב מקטע הרקמה בעזרת עט היסטוכימי. הוסף את תמיסת העבודה proteinase K (יחס של 1:9 בין תמיסת ציר ו- PBS) כדי לכסות את הרקמה ולדגור ב- 37 ° C למשך 30 דקות. שטפו את החלקים שלוש פעמים עם PBS במשך 5 דקות כל אחד.
- חדירה לקרום התא: נערו מעט את המקטעים יבשו ולאחר מכן כסו אותם ב-0.1% טריטון. דגרו במשך 20 דקות ב- RT, ושטפו את המקטעים שלוש פעמים עם PBS במשך 5 דקות כל אחד.
- חסימה: יש להוסיף 10% סרום עיזים כדי לכסות את הרקמה באופן שווה לחסימה ב-RT למשך 30 דקות.
- דגירה ראשונית של נוגדנים: יש להוסיף כמות מתאימה של נוגדן IgA מצומד AF488-conjugate נגד עכבר (1:500) כדי לכסות את הרקמה באופן אחיד ולדגור למשך הלילה בטמפרטורה של 4°C.
- גרעיני צביעה: שטפו את הפרוסות שלוש פעמים עם PBS במשך 5 דקות כל אחת. לאחר הסרת PBS, להוסיף את הכתם 4′,6-diamidino-2-phenylindole (DAPI) טיפה על החלקים לדגור במשך 15 דקות ב RT, מוגן מפני אור.
- שטפו ואטמו את המקטעים: שטפו את המקטעים שלוש פעמים עם PBS למשך 5 דקות, ולאחר מכן אטמו אותם עם אמצעי הרכבה נגד דהייה.
- מיקרוסקופיה וצילום: התבוננו במקטעים תחת מיקרוסקופ פלואורסצנטי וצלמו.
הערה: עבור DAPI, עירור אולטרה סגול הוא אורך גל 330-380 ננומטר, ואורך גל הפליטה הוא 420 ננומטר, אור כחול. אורך גל עירור פלואורסצאין איזותיוציאנט (FITC) הוא 465-495 ננומטר, ואורך גל הפליטה הוא 515-555 ננומטר, אור ירוק.
5. תרבית תאים
- להשיג קו לימפוציטים B אנושי DAKIKI מ ATCC, ארה"ב. תרבית תאי DAKIKI במדיום RPMI-1640 בתוספת 10% FBS ו-1% פניצילין-סטרפטומיצין.
- תרבית את התאים באינקובטור של 37 מעלות צלזיוס, 5% CO2 ותת-תרבית אותם כל 2-3 ימים. השתמש בתאים בשלב הגידול הלוגריתמי עבור כל הניסויים.
- במפגש של 70%-80%, אספו את התאים עם פיפטה סטרילית וצנטריפוגה את התאים ב 140 x גרם למשך 5 דקות. השליכו את הסופרנטנט, השהו מחדש עם המדיום נטול הסרום, ולאחר 24 שעות, השאירו את כל התאים בתקופה שקטה לטיפול הבא.
6. בדיקות ציטוטוקסיות LDH לבדיקת ריכוזים בטוחים של DIO על תאי DAKIKI רגילים
- זרעו תאי DAKIKI בצלחות 96 בארות בצפיפות של 4x105 תאים / באר והקימו קבוצת ביקורת נמוכה, קבוצת ביקורת גבוהה, וריכוזים שונים של DIO (0.25, 0.5, 1.0, 2.0, 4.0, 8.0 מיקרוגרם / מ"ל). דגירה באינקובטור 5% CO2, 37 מעלות צלזיוס למשך 24 שעות לאחר הטיפול המתאים על פי שיטת הקיבוץ.
- על פי ההוראות של ערכת זיהוי ציטוטוקסיות, להוסיף 5 μL של ליזט לכל באר בקבוצת הבקרה גבוהה, ולאחר מכן מניחים את הצלחת באינקובטור 5% CO2, 37 ° C למשך 15 דקות.
- מוציאים את הצלחת, מוסיפים 100 מיקרוליטר של תערובת התגובה לכל באר, דוגרים בחושך במשך 10 דקות ב-RT, ואז מוסיפים 50 מיקרוליטר של תמיסת תגובת העצירה. מדוד את ערך OD ב- 490 ננומטר בקורא המיקרו-לוחות בהקדם האפשרי.
- חשב את שיעור שחרור LDH של ריכוזים שונים של DIO ו- ≤10% כמינון המרבי המנוהל על פי הנוסחה: LDH שיעור שחרור = (באר ניסיונית LDH - LDH שליטה נמוכה) / (שליטה גבוהה LDH - LDH שליטה נמוכה) x 100%.
7. בדיקת CCK-8 לזיהוי ההשפעה של DIO על התפשטות תאי DAKIKI
- בהתבסס על תוצאות הניסויים הקודמים שלנו20, ביססו מודל IgAN באמצעות LPS 40 מיקרוגרם/מ"ל כדי לגרום לתאי DAKIKI.
- לאחר מכן, זרעו4x10 5 תאים/באר בצלחות של 96 בארות והתחלקו לקבוצות הבקרה, המודל והריכוז הנמוך, הבינוני והגבוה של DIO (0.25 מיקרוגרם/מ"ל, 0.5 מיקרוגרם/מ"ל ו-1.0 מיקרוגרם/מ"ל). לדגור את הצלחות באינקובטור 5% CO2, 37 ° C במשך 24 שעות.
- לאחר מכן, הוסף 20 μL של מגיב CCK-8 לכל באר והחזיר את הלוחות לאינקובטור (5% CO2, 37 ° C) למשך שעתיים. לאחר הדגירה, לזהות את OD באורך גל של 450 ננומטר על קורא microplate בהקדם האפשרי.
8. ELISA לגילוי ההשפעה של DIO על הפרשת IgA ו- Gd-IgA1 על ידי תאי DAKIKI
- זרעו תאי DAKIKI בצלחות 6 בארות בצפיפות של 6x106 תאים / באר, וקבצו וטפלו בתאים לפי שלב 7.2. תרבית את התאים במשך 24 שעות, ולאחר מכן צנטריפוגה ב 850 x גרם במשך 10 דקות ב 4 ° C כדי לקבל את supernatant.
- זיהוי ריכוזי IgA ו-Gd-IgA1 בהתאם להוראות הערכה.
9.qRT-PCR לזיהוי ההשפעה של DIO על C1GALT1 ורמות mRNA Cosmc בתאי DAKIKI
- זרעו את תאי ה-DAKIKI בצפיפות של 6x106 תאים/באר בצלחת בעלת 6 בארות, קבצו והתייחסו לתאים כאל CCK8 שהוזכר בשלב 7.2, ודגרו במשך 24 שעות. לחלץ את סך כל הרנ"א מתאי DAKIKI בהתאם להוראות ערכת מיצוי הרנ"א הכוללת.
- לאחר לקיחת 1 מיקרוליטר של RNA מופק מכל קבוצת דגימות ומדידת ריכוזו, יש לתמלל לאחור 1 מיקרוגרם של סך הרנ"א מכל דגימה ל-cDNA בהתאם להוראות הערכה.
- לאחר מכן, בצע הגברה RT-PCR כדי לזהות את הביטוי של כל גן (95 ° C במשך 15 דקות, 95 ° C עבור 10 שניות, ו 60 ° C עבור 30 שניות). חשב את רמת הביטוי של כל גן באמצעות שיטת 2-ΔΔCT עם β-אקטין כהפניה פנימית.
הערה: רצפי הפריימר היו כדלקמן:
C1GALT1: 5'-AAGGTTGACACCCAGCCTAA-3', 5'-CTTTGACGTGTTTGGCCTTT-3';
Cosmc: 5'-GCTCCTTTTTGAAGGGTGTG-3', 5'-TACTGCAGCCCAAAGACTCA-3';
β-אקטין : 5'-TCACCCACACTGTGCCCATCTACGA-3', 5'-CAGCGGAACCGCTCATTGCCAATGG-3'.
10. כתם מערבי לבחינת השפעת DIO על ביטוי חלבוני C1GALT1 וקוסמק בתאי DAKIKI
- זרעו את תאי DAKIKI בצפיפות של 6x106 תאים / באר בצלחת 6 בארות. קבץ אותם והתייחס אליהם כפי שהוזכר בשלב 7.2. לאחר 24 שעות של דגירה, לאסוף כל קבוצה של תאים.
- הוסף כמות מתאימה של תמיסת ליזה תאית (PMSF: מעכב פוספטאז: RIPA lysis solution = 1:1:100) ודגר על קרח במשך 30 דקות. לאחר מכן צנטריפוגה ב 13,500 x גרם במשך 10 דקות ב 4 ° C ולאסוף את supernatant.
- קבע את ריכוז החלבון באמצעות ערכת בדיקת ריכוז חלבון BCA.
- ערבבו את דגימות החלבון עם מאגר טעינה 5x SDS-PAGE ביחס של 4:1 על ידי ערבול, וחממו את הדגימות המעורבות בטמפרטורה של 100°C למשך 5 דקות כדי לנטרל את החלבון.
- כדי לזהות חלבונים בעלי משקל מולקולרי שונה, יש להוסיף את סמן החלבון (5 מיקרוליטר/באר) ואת הדגימות (20 מיקרוגרם/באר) לנתיבים שונים של ג'ל SDS-PAGE 12%, להריץ אלקטרופורזה SDS-PAGE ולהעביר את הג'ל לקרומי PVDF.
- חסום את הממברנות עם חלב 5% ללא שומן למשך שעתיים ב- RT ודגור עם הנוגדנים הראשוניים המתאימים (C1GALT1 [1:1000], Cosmc [1:2000]) למשך 24 שעות. השתמש בנוגדן β-אקטין (1:100000) כבקרה פנימית.
- שטפו את קרום הפוליווינילידן פלואוריד (PVDF) עם 1x Tris-Buffered Saline, 0.1% Tween 20 דטרגנט (TBST) שלוש פעמים (10 דקות לזמן), ולאחר מכן דגרו עם נוגדן IgG נגד ארנב עיזים משני (1: 10000) ב- RT למשך שעתיים.
- שטפו שוב את הממברנה עם TBST (שלוש פעמים למשך 10 דקות כל אחת), ופנקו אותה בכמות מתאימה של תמיסת עבודה כימילומינסנציה משופרת (ECL) (בהתאם להוראות היצרן) לזיהוי רצועת חלבונים.
- צלם את התמונות באמצעות מערכת הדמיה כימילומינסנציה ובצע ניתוח כמותי למחצה של ערכי האפור של חלבונים באמצעות מערכת ניתוח התמונה Image J.
11. ניתוח סטטיסטי
- השתמש ביישום תוכנה מתאים לניתוח הנתונים. בטא את כל הנתונים כאמצעי ± SD (סטיית תקן) והערך דגימות מרובות על ידי מבחן ANOVA חד-כיווני להשוואה בין קבוצות.
הערה: התוכנה הסטטיסטית SPSS 26.0 שימשה לניתוח סטטיסטי. שיטת LSD שימשה להשוואות דו-כיווניות בין קבוצות כאשר השונות הייתה שווה, ושיטת Dunnett T3 שימשה להשוואות דו-כיווניות בין קבוצות כאשר השונות לא הייתה שווה. P<0.05 נחשב כמצביע על הבדל מובהק סטטיסטית.
Representative Results
השפעת DIO על רקמת הכליה במודל עכברי IgAN
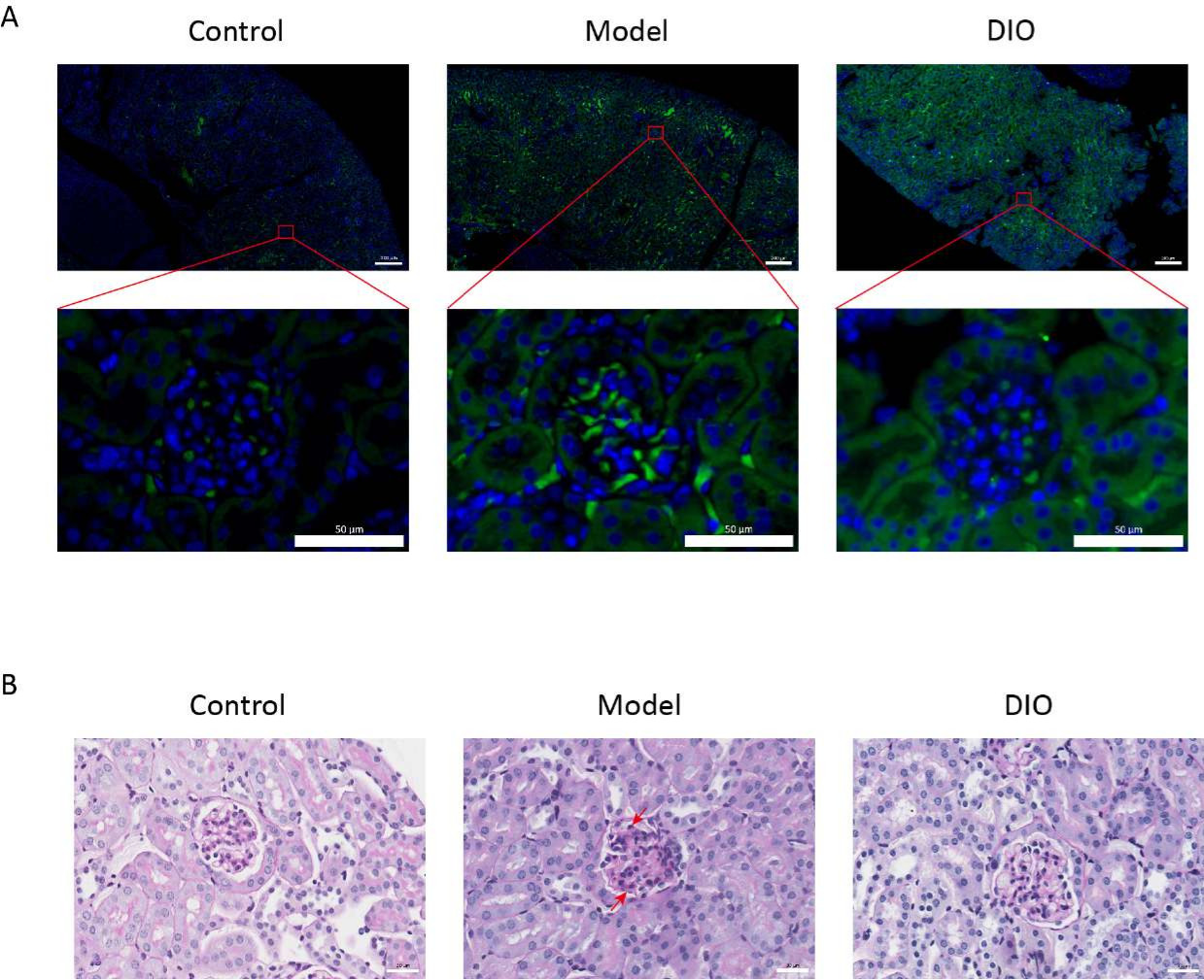
בהשוואה לקבוצת הביקורת, במודל עכברי IgAN המושרה על ידי מערכת החיסון הרירית (קבוצת המודל) הייתה עלייה משמעותית בפרוטאינוריה (איור משלים 2), שקיעת IgA נראתה באזור המסנגיאלי, פלואורסצנטיות התפזרה באופן אחיד באשכולות לאורך כל האזור המסנגיאלי (איור 1A), צביעת PAS של רקמת הכליה הראתה שגשוג תאים מזנגיאליים והיפרפלזיה סטרומלית (איור 1B)אשר צומצם בקבוצת DIO gavage (קבוצת DIO).,
השפעת DIO על לימפוציטים מסוג B במדבקת פייר
המדבקה של פייר היא האתר המוביל להמרה של לימפוציטים מסוג B לתאים מפרישי IgA. לקחנו את המדבקה של פייר כאובייקט המחקר כדי לבחון את ההשפעה של DIO על לימפוציטים מסוג B על ידי זיהוי הביטוי של סמני תאי B CD20 ו- CXCR5. תוצאות אימונוהיסטוכימיות הראו כי הביטוי של CD20 ו- CXCR5 היה גבוה משמעותית בקבוצת המודל בהשוואה לקבוצת הביקורת. DIO יכול לעכב את הביטוי של הסמנים המולקולריים לעיל (איור 2A,B).
טווח הריכוז הבטוח של DIO על תאי DAKIKI
LDH הוא סמן של שלמות קרום הפלזמה ואינדיקטור למוות תאי, עם שיעורי שחרור LDH גבוהים יותר המעידים על נזק חמור יותר לתאים. בדיקת שחרור LDH שימשה לקביעת טווח הריכוז הבטוח של DIO. הריכוז הבטוח המרבי של DIO נקבע על ידי שיעור שחרור LDH מתחת ל -10%. התוצאות (איור 3) לא הראו ציטוטוקסיות משמעותית המושרה על-ידי DIO בריכוזים של 0.25 עד 1.0 מיקרוגרם/מ"ל. לכן, המחקר הבא השתמש 0.25, 0.5, ו 1.0 מיקרוגרם / מ"ל DIO כרמת המינון.
השפעות DIO על התפשטות תאי DAKIKI
תוצאות הניסוי (איור 4) הראו כי בהשוואה לקבוצת המודל (קבוצת LPS-stimulated), DIO עיכב את התפשטות תאי DAKIKI המושרה על ידי LPS באופן תלוי ריכוז. DIO בריכוזים של 0.5 ו-1.0 מיקרוגרם/מ"ל עיכב באופן משמעותי את התפשטות תאי DAKIKI הנגרמת על ידי LPS (P < 0.01).
השפעות DIO על תפקוד ההפרשה של תאי DAKIKI
רמות Gd-IgA1 קשורות קשר הדוק לתהליך הפתולוגי של IgAN, וסך IgA נבדק יחד כאינדיקטור לתפקוד ההפרשה התאית. בדיקת ELISA שימשה לזיהוי תוכן IgA ו- Gd-IgA1 בסופרנאטנט של תרבית תאי DAKIKI. התוצאות הראו (איור 5A,B) שתאי DAKIKI שעוררו על-ידי LPS הפרישו יותר IgA בהשוואה לקבוצת הביקורת (P < 0.01). לשם השוואה, DIO עיכב באופן משמעותי תאי DAKIKI מלהפריש IgA (P < 0.01) באופן תלוי ריכוז. בהשוואה לקבוצת הביקורת, תאי DAKIKI שעוררו על ידי LPS הפרישו יותר Gd-IgA1 עם נטייה סטטיסטית (P < 0.10), ו-DIO עיכב הפרשת Gd-IgA1 מתאי DAKIKI מעוררי LPS באופן תלוי ריכוז (P < 0.05 ו-P < 0.01), ביניהם DIO ב-1.0 מיקרוגרם/מ"ל עיכב משמעותית את הפרשת Gd-IgA1 בשיעור עיכוב של 25%.
המנגנון של DIO מעכב הפרשת Gd-IgA1 על ידי תאי DAKIKI
כדי להמשיך ולחקור את המנגנון האפשרי של DIO המעכב הפרשת יתר של Gd-IgA1 על ידי תאי DAKIKI, רמות ה-glycosylated transferase C1GALT1 והחלבון המלווה Cosmc mRNA בתאי DAKIKI זוהו על-ידי qRT-PCR, והתוצאות הראו (איור 6A,B) שביטוי ה-mRNA היחסי של C1GALT1 ו-Cosmc היה מווסת כלפי מטה בתאי DAKIKI בקבוצת המודל בהשוואה לקבוצת הביקורת (P < 0.01). DIO ויסת את ביטוי ה-mRNA היחסי של C1GALT1 ו-Cosmc בדרגות שונות בהשוואה לקבוצת המודלים, כאשר DIO 1.0 מיקרוגרם /מ"ל שיפר משמעותית את ביטוי ה-mRNA היחסי של C1GALT1 ו-Cosmc (P <-0.05).
במקביל, שיטת WB שימשה לזיהוי ההשפעה של DIO על ביטוי החלבון של C1GALT1 ו- Cosmc בתאי DAKIKI. בהשוואה לקבוצת הביקורת, ביטוי החלבון של C1GALT1 ו- Cosmc בתאי DAKIKI בקבוצת המודל ירד כמובן (P < 0.05). בהשוואה לקבוצת המודל, ביטוי החלבון של C1GALT1 ו- Cosmc לאחר התערבות DIO היה מווסת יותר. ביטוי החלבונים של C1GALT1 ו-Cosmc היה מווסת באופן משמעותי על-ידי DIO בריכוז של 1.0 מיקרוגרם/מ"ל (P < 0.05) (איור 7A-C).

איור 1: היסטופתולוגיה של הכליות. (A) מיקרוסקופ אימונופלואורסצנטי. חלקי כליות של עכברים בכל קבוצה הוכתמו באנטי-IgA (ירוק) ו-DAPI (כחול). סרגל קנה המידה של התמונה לעיל = 200 מיקרומטר. סרגל קנה המידה של התמונה למטה = 50 מיקרומטר. n = 6 לכל קבוצה. (B) תמונות מייצגות של צביעת PAS של רקמת כליה מעכברים בקבוצות הבקרה, המודל וה-DIO. סרגל קנה מידה = 30 מיקרומטר. החץ כלפי מטה מראה את התאים המסנגיאליים והחץ כלפי מעלה מראה את הסטרומה. סרגל קנה מידה = 30 מיקרומטר. n = 6 לכל קבוצה. אנא לחץ כאן כדי להציג גרסה גדולה יותר של איור זה.
{kind=link}

איור 2: השפעת DIO על סמני לימפוציטים מסוג B. (A) הביטוי של CD20 במדבקת פייר. סרגל קנה מידה = 200 מיקרומטר. n = 6 לכל קבוצה. (B) הביטוי של CXCR5 בתיקון פייר. סרגלי קנה המידה נמצאים בפינה הימנית התחתונה של התמונה. סרגל קנה מידה = 200 מיקרומטר. n = 6 לכל קבוצה. אנא לחץ כאן כדי להציג גרסה גדולה יותר של איור זה.
{kind=link}

איור 3. בדיקת הריכוז הבטוח של DIO על תאי DAKIKI. ערכים סטטיסטיים מבוטאים כממוצע ± SD משלושה ניסויים עצמאיים. אנא לחץ כאן כדי להציג גרסה גדולה יותר של איור זה.
{kind=link}

איור 4. ריכוזים שונים של DIO משפיעים על התפשטות תאי DAKIKI. הנתונים בוטאו כממוצע ±SD. בהשוואה לקבוצת הביקורת, **P < 0.01; בהשוואה לקבוצת הדגמים, #P < 0.05' ##P < 0.01; תוצאות כל הניסויים חזרו על עצמן שלוש פעמים. אנא לחץ כאן כדי להציג גרסה גדולה יותר של איור זה.
{kind=link}

איור 5. DIO מעכב הפרשת IgA ו-Gd-IgA1 על ידי תאי DAKIKI. (A) שיטת ELISA זיהתה את הביטוי של IgA בכל קבוצה. (B) שיטת ELISA זיהתה את הביטוי של Gd-IgA1 בכל קבוצה. הנתונים בוטאו כממוצע ± SD. בהשוואה לקבוצת הביקורת, **P < 0.01; בהשוואה לקבוצת הדגמים, #P < 0.05, ##P < 0.01; כל תוצאות הניסוי חזרו על עצמן שלוש פעמים. אנא לחץ כאן כדי להציג גרסה גדולה יותר של איור זה.
{kind=link}

איור 6. המנגנון של DIO מעכב הפרשת יתר של Gd-IgA1 על ידי תאי DAKIKI. (A) QRT-PCR זיהה את ביטוי ה-mRNA של C1GALT1. (B) QRT-PCR זיהה את ביטוי ה-mRNA של Cosmc. הנתונים בוטאו כממוצע ± SD. בהשוואה לקבוצת הביקורת, **P<0.01; בהשוואה לקבוצת הדגמים, #P < 0.05, ##P < 0.01; כל תוצאות הניסוי חזרו על עצמן שלוש פעמים. אנא לחץ כאן כדי להציג גרסה גדולה יותר של איור זה.
{kind=link}

איור 7. DIO משפיע על ביטוי חלבונים של C1GALT1 ו- Cosmc בתאי DAKIKI. (A) WB אימת את הוויסות של ביטוי חלבונים של C1GALT1 ו- Cosmc על ידי DIO. (B) ניתוח כמותי למחצה של ביטוי C1GALT1 בוצע באמצעות תמונה J. (C) ניתוח כמותי למחצה של ביטוי Cosmc באמצעות תמונה J. הנתונים בוטאו כממוצע ±SD. בהשוואה לקבוצת הביקורת,*P < 0.05; בהשוואה לקבוצת המודלים, #P < 0.05, ##P < 0.01, כל תוצאות הניסוי חזרו על עצמן שלוש פעמים. אנא לחץ כאן כדי להציג גרסה גדולה יותר של איור זה.
{kind=link}
תרשים משלים 1. הסכימה עבור מודל in vivo. אנא לחץ כאן כדי להוריד איור זה.
תרשים משלים 2. שינויים בפרוטאינוריה. הנתונים בוטאו כממוצע ± SD; n = 6 לכל קבוצה. אנא לחץ כאן כדי להוריד נתון זה.
{kind=link}
Discussion
התכונה הפתולוגית האופיינית של IgAN היא התצהיר של מתחמי חיסון המכילים IgA1 ו- GD-IgA1 באזור mesangial של glomerulus21,22. הפחתת היווצרות של קומפלקסים חיסוניים יכולה להפחית את הפגיעה הכלייתית ולהקל על הסימפטומים הקליניים של IgAN. בניסוי in vivo חקרנו את ההשפעות הטיפוליות של DIO על IgAN, ומצאנו כי DIO יכול להפחית שקיעת IgA בכליה של עכברי מודל IgAN. הוכח כי הצטברות של תאים מפרישי IgA בכליה קשורה לפתוגנזה של IgAN23. כאתר חשוב של התרבות והפעלה של לימפוציטים מסוג B, המדבקה של פייר היא מקור חשוב לתאים מפרישי IgA, ולכן בחנו את הביטוי של סמני לימפוציטים מסוג B (CD20, CXCR5) במדבקה של פייר ומצאנו כי DIO יכול לעכב את הביטוי של לימפוציטים מסוג B במדבקת פייר של עכברי IgAN. תוצאות ניסוי אלה יכולות לספק בסיס ליישום DIO בטיפול ב- IgAN.
ביצענו את הניסויים הבאים במבחנה כדי להמשיך לחקור את מנגנון הפעולה של DIO על IgAN. ראשית, הוכח קודם לכן כי DAKIKI, קו תאי B אימורטלי EBV המפריש IgA1, שחלקו GD-IgA124, אידיאלי למחקר חוץ גופי של מנגנון הפעולה של התרופה על IgAN. בחרנו בתאי DAKIKI כדי לחקור את המנגנון המולקולרי של DIO בטיפול ב-IgAN. בנוסף, התגובה החיסונית הדלקתית הרירית ממלאת תפקיד אינטגרלי בפתוגנזה של IgAN. כפי שהוזכר לעיל, אנו משתמשים ב- LPS כדי לעורר תאי DAKIKI, אשר יכולים לשחרר גורמים פרו-דלקתיים ולתווך תגובות דלקתיות, אשר יכולים לחקות טוב יותר את מנגנון התגובה החיסונית הרירית ב- IgAN. המודל התאי במבחנה עשוי לסייע בחקר האפשרות והמנגנון של תרופות אחרות לטיפול ב- IgAN. התוצאות הראו כי DIO עיכב את התפשטות תאי DAKIKI שעוררו על ידי LPS באופן תלוי ריכוז. DIO יכול לעכב את הפרשת IgA ו- Gd-IgA1 בתאי DAKIKI הנגרמת על ידי גירוי LPS ולווסת את הביטוי של mRNA וחלבון של C1GalT1 והמלווה שלו Cosmc בתאי DAKIKI, מה שמרמז על כך ש- DIO יכול להפחית את הפרשת Gd-IgA1 על ידי ויסות ביטוי C1GALT1/Cosmc ובכך לעכב את הפעלת היתר של תאי DAKIKI.
יש לציין שלבים מרכזיים במהלך הליכי הניסוי. הריכוז של Gd-IgA1 בסופרנאטנט של תא DAKIKI אינו בטווח הגילוי של ערכת ELISA (1.56~100 ננוגרם/מ"ל), והסופרנאטנט שנאסף חייב להיות צנטריפוגה על ידי צינור אולטרה-סינון כדי לקבל את ה-Gd-IgA1 המרוכז. כמו כן, יש לוודא שנפח הסופרנאטנט החל מכל קבוצה זהה והנפח הסופי של התרכיז המתקבל לאחר אולטרה-סינון זהה.
במחקר זה, השתמשנו בשיטות in vitro ו - in vivo בו זמנית, אשר יכולות לתמוך זו בזו באופן הדדי בהשפעות פרמקולוגיות ולספק דוגמה לחקר ההשפעות והמנגנונים שלהן של רפואת צמחים. יש דברים שאפשר לשפר בפרוטוקול הזה. ראשית, לא זיהינו ריכוזי דם בקבוצת ה-DIO gavage של העכברים; לכן, ריכוז DIO שווה ערך לריכוזי דם אינו משמש בניסויי מבחנה . שנית, רק מונומר DIO, המרכיב הפעיל של DNR, נחקר; ההשפעות של רכיבים אחרים של DNR על IgAN עדיין זקוקות למחקר נוסף.
לסיכום, מחקר זה מספק בסיס ניסיוני לטיפול IgAN עם DIO, החומר הפעיל של DNR. מחקר זה ביסס מודל פתולוגי תאי של IgAN על ידי חיקוי התגובה החיסונית הרירית של IgAN הן במבחנה והן in vivo. זה נותן רעיון חדש ללימוד רפואה סינית מסורתית כדי למנוע ולטפל IgAN.
Disclosures
המחברים מצהירים כי אין להם אינטרסים כלכליים מתחרים.
Acknowledgements
עבודה זו נתמכה על ידי הקרן הלאומית למדעי הטבע של סין (81973675).
Materials
| Name | Company | Catalog Number | Comments |
| Anti-CD20/MS4A1 Antibody | Boster Biotechnology Company | A03780-3 | |
| Antifade mounting medium | Beyotime, Shanghai, China | P0128S | |
| Balb/c mice | Beijing Weitong Lihua Laboratory Animal Technology Co., Ltd. | 110322220101424000 | |
| blocking serum | Solarbio, Beijing, China | SL038 | |
| Bovine gamma globulin | ShangHai YuanYe Biotechnology Company | S12031 | |
| C1GALT1 polyclonal antibody | Proteintech Group, Inc,USA | 27569-1-AP | |
| Citrate antigen retrieval solution(50×) | Phygene Biotechnology Company | PH0422 | |
| COSMC polyclonal antibody | Proteintech Group, Inc,USA | 19254-1-AP | |
| Cytotoxiciy detection kit | Roche Company | 4744926001 | |
| Dako REAL EnVision detection system, Peroxidase/DAB+ | Dako | K5007 | |
| DAPI | Invitrogen | D1306 | |
| Dioscin | National Institute For Food and Drug Control | 111707-201703 | |
| DIO tablets | Chengdu No 1 Pharmaceutical Co. Ltd. | H51023866 | |
| ECL working solution | Merck Biotechnology, Inc | WBKLS0100 | |
| Enhanced cell counting kit-8 | Beyotime, Shanghai, China | C0043 | |
| Fasking one-step removal of gene cDNA first-strand synthesis premix | TIANGEN,Beijing, China | KR118-02 | |
| Glycogen Periodic acid Schiff (PAS) stain kit | BaSO Biotechnology Company | BA4080A | |
| Goat anti-mouse IgA-AF488 | SouthernBiotech | 1040-30 | |
| Goat anti-rabbit IgG antibody (H+L), HRP conjugated | BeiJing Bioss Biotechnology Company | BS-0295G-HRP | |
| Human Gd-IgA1 ELISA kit | IBL | 27600 | |
| Human IgA ELISA kit | MultiSciences (LiankeBio) | 70-EK174-96 | |
| Pierce BCA protein assay kit | Thermo Scientific | 23227 | |
| PMSF solution | Beyotime, Shanghai, China | ST507 | |
| Proteinase K | Phygene Biotechnology Company | PH1521 | |
| Rabbit anti-CXCR5 polyclonal antibody | BeiJing Bioss Biotechnology Company | bs-23570R | |
| RIPA lysis buffer | Beyotime, Shanghai, China | P0013B | |
| RNAsimple total RNA extraction kit | TIANGEN,Beijing, China | DP419 | |
| RPMI Medium 1640 | Solarbio, Beijing, China | 31800 | |
| Super-Bradford protein assay kit | CWBIO, Beijing, China | CW0013 | |
| Triton X-100 | Beyotime, Shanghai, China | ST795 | |
| β-Actin Rabbit mAb | Abclonal, Wuhan, China | AC026 |
References
- Knoppova, B., et al. The origin and activities of IgA1-containing immune complexes in IgA nephropathy. Frontiers in Immunology. 7, 117 (2016).
- Suzuki, H., et al. The pathophysiology of IgA nephropathy. Journal of The American Society of Nephrology. 22 (10), 1795-1803 (2011).
- He, L., et al. Synthetic double-stranded RNA poly(I:C) aggravates IgA nephropathy by triggering IgA class switching recombination through the TLR3-BAFF axis. American Journal of Nephrology. 42 (3), 185-197 (2015).
- Zhao, N., et al. The level of galactose-deficient IgA1 in the sera of patients with IgA nephropathy is associated with disease progression. Kidney International. 82 (7), 790-796 (2012).
- Xing, Y., et al. C1GALT1 expression is associated with galactosylation of IgA1 in peripheral B lymphocyte in immunoglobulin a nephropathy. BMC Nephrology. 21 (1), 18 (2020).
- Qin, W., et al. External suppression causes the low expression of the Cosmc gene in IgA nephropathy. Nephrology Dialysis Transplantation. 23 (5), 1608-1614 (2008).
- Sakai, F., et al. Lactobacillus gasseri SBT2055 induces TGF-β expression in dendritic cells and activates TLR2 signal to produce IgA in the small intestine. PLoS One. 9 (8), 105370 (2014).
- Gutzeit, C., Magri, G., Cerutti, A. Intestinal IgA production and its role in host-microbe interaction. Immunological Reviews. 260 (1), 76-85 (2014).
- Serino, G., et al. In a retrospective international study, circulating miR-148b and let-7b were found to be serum markers for detecting primary IgA nephropathy. Kidney International. 89 (3), 683-692 (2016).
- Lu, F., et al. Therapeutic effect of Rhizoma Dioscoreae Nipponicae on gouty arthritis based on the SDF-1/CXCR 4 and p38 MAPK pathway: an in vivo and in vitro study. Phytotherapy research: PTR. 28 (2), 280-288 (2014).
- Wang, W., Xu, L., Zhou, L., Wan, S., Jiang, L. A Network pharmacology approach to reveal the underlying mechanisms of Rhizoma Dioscoreae Nipponicae in the treatment of asthma. Evidence-Based Complementary and Alternative Medicine: eCAM. 2022, 4749613 (2022).
- Tian, W. W., Wei, Y. Professor TONG Xiaolin used the experience of Dioscoreae Nipponicae. Jilin Journal of Chinese Medicine. 40 (05), 589-592 (2020).
- Rao, X. R., Bai, Y. W. Das Xiwen's experience in treating IgA nephropathy. Beijing Journal of Traditional Chinese Medicine. 9, 691-693 (2008).
- Si, Y., Zhang, Y. A data mining study on the pattern of medication use in the treatment of IgA nephropathy by Professor Zhang Yu. Journal of Chinese Physician. 20 (01), 109-111 (2018).
- Jiang, H., et al. Optimization of the enzymatic extraction technology of Diosgenin from Dioscorea nipponica. Chinese Traditional Patent Medicine. 39 (03), 621-624 (2017).
- Qi, M., et al. Dioscin alleviates lipopolysaccharide-induced inflammatory kidney injury via the microRNA let-7i/TLR4/MyD88 signaling pathway. Pharmacological Research. 111, 509-522 (2016).
- Yang, L., et al. Recent advances in the pharmacological activities of Dioscin. BioMed Research International. 2019, 5763602 (2019).
- Nal Zou, J., et al. Toll-like receptor 4 signaling pathway in the protective effect of Pioglitazone on experimental immunoglobulin A nephropathy. Chinese Medical Journal. 130 (8), 906-913 (2017).
- Xu, S. Y., Bian, R. L., Chen, X. Pharmacological experiments methodology. Chinese Pharmacological Bulletin. 1, 19 (1992).
- Shen, J. C., Ren, Y., Rao, X. R., You, Y., Li, S. Network pharmacology, molecular docking, and in vitro experiments to explore the molecular mechanism of Dioscorea Nipponica Makion in the treatment of IgA nephropathy. World Journal of Integrated Traditional and Western Medicine. 16 (12), 2246-2254 (2021).
- Mestecky, J., et al. IgA nephropathy: molecular mechanisms of the disease. Annual Review of Pathology. 8, 217-240 (2013).
- Novak, J., et al. IgA1-containing immune complexes in IgA nephropathy differentially affect proliferation of mesangial cells. Kidney International. 67 (2), 504-513 (2005).
- Nihei, Y., et al. Identification of IgA autoantibodies targeting mesangial cells redefines the pathogenesis of IgA nephropathy. Science Advances. 9 (12), (2023).
- Raska, M., et al. Identification and characterization of CMP-NeuAc: GalNAc-IgA1 alpha2,6-sialyltransferase in IgA1-producing cells. Journal of Molecular Biology. 369 (1), 69-78 (2007).
Reprints and Permissions
Request permission to reuse the text or figures of this JoVE article
Request PermissionThis article has been published
Video Coming Soon
ABOUT JoVE
Copyright © 2025 MyJoVE Corporation. All rights reserved