Engenharia de Assemblóides de Tendão para Sondar Crosstalk Celular em Doenças e Reparos
In This Article
Summary
Aqui, apresentamos um sistema modelo assemblóide para mimetizar o crosstalk celular do tendão entre o tecido central do tendão que suporta carga e um compartimento extrínseco contendo populações celulares ativadas por doença e lesão. Como um importante caso de uso, demonstramos como o sistema pode ser implantado para sondar a ativação de células endoteliais extrínsecas relevantes para a doença.
Abstract
Os tendões permitem a locomoção transferindo forças musculares para os ossos. Eles dependem de um núcleo tendíneo resistente composto por fibras colágenas e populações de células estromais. Esse núcleo de suporte de carga é englobado, nutrido e reparado por uma camada de tecido semelhante a um sinovial que compreende o compartimento extrínseco do tendão. Apesar desse design sofisticado, lesões tendíneas são comuns, e o tratamento clínico ainda depende de fisioterapia e cirurgia. As limitações dos sistemas de modelos experimentais disponíveis retardaram o desenvolvimento de novos tratamentos modificadores da doença e regimes clínicos de prevenção de recaída.
Os estudos in vivo em humanos limitam-se a comparar tendões saudáveis com tecidos doentes ou rotos em estágio terminal amostrados durante a cirurgia de reparo e não permitem o estudo longitudinal da doença tendínea subjacente. Modelos animais in vivo também apresentam limites importantes em relação à complexidade fisiológica opaca, à carga ética sobre os animais e aos grandes custos econômicos associados ao seu uso. Além disso, modelos animais in vivo são pouco adequados para sondagem sistemática de drogas e vias de interação multicelular e multitecidual. Sistemas modelos in vitro mais simples também ficaram aquém. Uma das principais razões é a falha em replicar adequadamente a carga mecânica tridimensional necessária para estudar significativamente as células tendinosas e sua função.
O novo sistema de modelo 3D apresentado aqui alivia alguns desses problemas explorando explantes de núcleo do tendão da cauda murina. É importante ressaltar que esses explantes são facilmente acessíveis em grande número a partir de um único mouse, retêm padrões de carregamento 3D in situ no nível celular e apresentam uma matriz extracelular semelhante à in vivo. Neste protocolo, instruções passo a passo são dadas sobre como aumentar os explantes do núcleo do tendão com hidrogéis de colágeno carregados de células endoteliais derivadas do músculo, fibroblastos derivados do tendão e macrófagos derivados da medula óssea para substituir as populações de células ativadas por doenças e lesões dentro do compartimento extrínseco do tendão. Demonstra-se como os assembloides tendinosos resultantes podem ser desafiados mecanicamente ou através de estímulos microambientais definidos para investigar crosstalk multicelular emergente durante doença e lesão.
Introduction
Em sua função de transferir forças musculares para os ossos para permitir o movimento, os tendões enfrentam alguns dos mais extremos estresses mecânicos que ocorrem no corpo humano 1,2,3. Devido ao envelhecimento das sociedades, ao aumento da prevalência da obesidade e à crescente popularidade de atividades esportivas mecanicamente exigentes, projeta-se que a prevalência de doenças e lesões tendíneas aumente nos paísesdesenvolvidos4,5,6. O desenvolvimento de novos esquemas de tratamento baseados em evidências e modificadores da doença para combater esse aumento tem sido dificultado pelas limitações dos sistemas-modelo atualmente disponíveis 1,7,8.
Idealmente, os modelos de reparo de doenças e lesões permitiriam estudar como o órgão-alvo processa um conjunto definido de parâmetros de entrada (imitando gatilhos de doença, Tabela 1) em parâmetros de saída mensuráveis (representando as características da doença, Tabela 2), controlando os fatores de confusão. Estudos utilizando tais sistemas modelo seriam então capazes de identificar os processos (pato-) fisiológicos subjacentes ao reparo de doenças e lesões e obter conhecimentos que poderiam ser explorados para prevenir ou reduzir as características de doenças e lesões nas clínicas. Aplicando esse princípio aos tendões, um sistema modelo útil deve recapitular partes centrais da resposta in vivo do tendão à doença e à lesão, que englobam as seguintes características: microdano, inflamação, neovascularização, hipercelularidade, turnover acelerado da matriz e descompartimentalização9,10,11,12,13,14,15. Usando essas características como base, os seguintes requisitos para um sistema modelo de reparo de lesões e doenças tendíneas bem-sucedido podem ser inferidos.
Hipotetiza-se que a sobrecarga mecânica seja um fator central na patogênese da lesão tendínea e da doença, sendo, portanto, uma abordagem experimental comumente usada para criarmicrodanos16. A capacidade de carga mecânica controlável é, portanto, um pré-requisito primordial para modelos de reparo de doenças e lesões tendíneas. Idealmente, o sistema do modelo permite três modos principais: carga única de estiramento a dano, carga em fadiga e descarga 8,17,18. Na deformação mecânica, as células residentes no tecido experimentam uma combinação complexa de forças tensionais, forças de cisalhamento (devido ao deslizamento das fibras colágenas ao redor das células) e forças de compressão que ocorrem durante a descarga ou próximo à entese19,20. Os sistemas modelo devem recriar esses padrões de carregamento complexos o mais próximo possível.
Uma maneira alternativa de introduzir microdanos na matriz é aproveitar estressores bioquímicos que mimetizam predisposições sistêmicas para doenças e lesões tendíneas, como citocinas (pró-)inflamatórias, estresse oxidativo ou altas concentrações de glicose 21,22,23. Consequentemente, um microambiente de nicho controlável é vantajoso para um sistema modelo de reparo de doenças e lesões tendíneas.
Um pré-requisito comum para que sistemas-modelo sejam capazes de recapitular inflamação, neovascularização e hipercelularidade é a presença seletiva de populações celulares que conduzem esses processos24. Para processos inflamatórios, essas populações incluem neutrófilos, células T e macrófagos, enquanto células endoteliais e pericitos seriam necessários para estudar a neovascularização 25,26,27,28,29. Os fibroblastos tendíneos não são apenas vitais para o reparo tendíneo, mas, como células proliferativas e migratórias, também parcialmente responsáveis pela hipercelularidade local observada na doença tendínea30,31,32,33,34,35,36.
Além das alterações nas populações celulares residentes, a composição da matriz tendínea encontra-se alterada nas doenças e lesões tendíneas7,37,38,39,40. Para apresentar as pistas microambientais relevantes para a doença, os sistemas-modelo devem ser capazes de integrar uma composição de matriz extracelular compatível com o estágio alvo da doença ou lesão, por exemplo, permitindo combinações proporcionais relevantes de colágeno-1, colágeno-3 e fibronectina celular41.
A compartimentalização de tendões sadios no núcleo do tendão e nos compartimentos extrínsecos (isto é, endotenon, epitenon e paratenon) é central para sua função e frequentemente perturbada em tendões doentes ou lesados 1,42,43,44,45,46,47 . A incorporação da compartimentalização 3D do tendão em sistemas modelo de tendão é, portanto, necessária não apenas para simular mais de perto os processos subjacentes à descompartimentação e recompartimentação, mas também ajuda a estabelecer os gradientes espaço-temporais corretos de citocinas e nutrientes48,49.
Finalmente, a modularidade é outro ativo central dos sistemas modelo, permitindo aos pesquisadores combinar a correta contribuição relativa e interação entre os estressores descritos anteriormente durante os processos investigados 8,17.
Além de selecionar as modalidades de entrada ideais, um passo importante é ser capaz de medir, observar e rastrear mudanças na saída resultante. As propriedades mecânicas do sistema modelo (i.e., comprimento da região dos dedos, módulo elástico linear, deformação máxima à tração, tensão máxima de tração, resistência à fadiga e relaxamento de tensões) são centrais aqui, pois caracterizam a função principal do tendão 50,51,52. Para relacionar essas alterações funcionais às alterações no nível tecidual, é importante permitir métodos que detectem danos à matriz estrutural (colágeno) e rastreiem a proliferação e o recrutamento de populações celulares relevantes para doenças e reparos 30,53,54,55,56,57,58,59,60.
Para estudar o crosstalk célula-célula e célula-matriz emergente, deve-se ser capaz de isolar ou marcar proteínas em quantidades adequadas para quantificação (i.e., ELISA, proteômica, imunohistoquímica, citometria de fluxo)14,21,61,62. A análise da expressão gênica populacional ou, pelo menos, compartimento-específica também deve ser possível (i.e., classificação celular ativada por fluorescência [FACS], sequenciamento de RNA de célula única/RNA em massa e reação em cadeia da polimerase quantitativa em tempo real (RT-qPCR))21,24,27,63. O sistema modelo deve permitir a medição do maior número de parâmetros de saída acima mencionados no mesmo espécime e em vários corpos de prova de uma maneira rápida o suficiente para desbloquear estudos de alto rendimento.
Entre os sistemas-modelo atualmente disponíveis para estudar a reparação de doenças e lesões tendíneas humanas, o próprio corpo humano é, naturalmente, o mais representativo. É também o menos compatível com a intervenção experimental. Enquanto pacientes com lesões agudas do tendão estão abundantemente disponíveis para estudos clínicos, pacientes com tendinopatia precoce (a doença tendínea mais comum) são em grande parte livres de sintomas e muitas vezes não são detectados clinicamente até que alterações mais graves se manifestem14,64,65. Isso torna difícil identificar o momento crítico em que a homeostase tendínea descarrila e os mecanismos por trás desse descarrilamento 16,66,67,68,69. Além disso, a extração de biópsias de tendões saudáveis é eticamente desafiadora, pois pode resultar em danos persistentes. Remanescentes de tendões isquiotibiais da cirurgia de reconstrução do ligamento cruzado anterior são frequentemente usados como controles saudáveis, mas indiscutivelmente diferem em função, propriedades mecânicas, populações celulares e composição da matriz em comparação com os tendões do manguito rotador, Aquiles e patelar, comumente afetados por doença e lesão tendínea 70,71,72,73.
Os modelos animais in vivo são mais acessíveis e tratáveis, mas seu uso impõe uma carga ética significativa aos animais e um custo econômico aos pesquisadores. Além disso, a maioria dos animais modelo populares não desenvolve lesões tendinopáticas espontaneamente (isto é, ratos, camundongos, coelhos) ou não possui os primers e cepas geneticamente modificadas necessárias para rastrear as vias de comunicação multicelular envolvidas (isto é, cavalos, coelhos).
Sistemas modelo 2D in vitro simples estão do outro lado do espectro de complexidade/tratabilidade e permitem melhor o estudo controlado e eficiente em termos de tempo de vias específicas de comunicação intercelular em resposta a um conjunto mais controlável de gatilhos 8,74. No entanto, esses sistemas simplificados geralmente falham em recapitular o carregamento mecânico multidimensional (isto é, tensão, compressão e cisalhamento) que é central para a funcionalidade do tendão. Além disso, as (também) altas rigidezes do plástico de cultura de tecidos tendem a se sobrepor a quaisquer pistas matriciais fornecidas por revestimentos destinados a mimetizar o estado de interesse da doença75,76.
Para superar essa desvantagem, sistemas de modelos 3D cada vez mais sofisticados têm sido desenvolvidos para fornecer uma matriz carregável cuja composição possa ser pelo menos parcialmente compatível com o estado desejado da doença 77,78,79. Ainda assim, esses sistemas não apenas lutam para replicar com precisão as complexas composições de matriz extracelular in vivo e padrões de carga celular, mas geralmente carecem de capacidade de carga a longo prazo e das interfaces compartimentais necessárias para estudar as vias de comunicação intercompartimentais que coordenam o reparo de doenças e lesões tendíneas 48,49,80.
Sistemas modelo de explante de tendão ex vivo têm a vantagem distinta de uma composição de matriz embutida in vivo, que compreende nichos pericelulares, barreiras transcompartimentais, bem como gradientes espaço-temporais de citocinas/nutrientes e recapitula padrões complexos de carga quando esticados8. Como resultado dos limites de difusão de nutrientes dependentes do tamanho, explantes de modelos animais maiores (i.e., cavalos) são difíceis de manter vivos para o estudo a longo prazo de doenças tendíneas e reparo de lesões 81,82,83. Enquanto isso, explantes menores de espécies murinas (i.e., tendão de Aquiles, tendão patelar) são difíceis de fixar e carregar mecanicamente de forma reprodutível. Seu tamanho também restringe a quantidade de material que pode ser coletado para leituras em nível de célula, proteína e gene sem reunir amostras e diminuir o rendimento. Nesse sentido, os fascículos tendinosos da cauda murina oferecem o potencial para desbloquear o estudo de alto rendimento do reparo de doenças e lesões tendíneas, uma vez que estão prontamente disponíveis em grandes quantidades a partir de um único camundongo, preservam a complexa composição da matriz pericelular in vivo e recapitulam os padrões de carga celular. Durante o processo de extração, entretanto, perdem a maior parte de seu compartimento extrínseco e nele continham populações vasculares, imunológicas e de fibroblastos, que hoje são consideradas como condutoras de doenças tendíneas e reparo 8,18.
Para preencher essa lacuna, um sistema modelo combinando as vantagens dos explantes de núcleo derivados do tendão da cauda murina com as vantagens dos sistemas modelo baseados em hidrogel 3D foi desenvolvido. Esse sistema modelo consiste de um hidrogel carregado de células (colágeno-1) lançado ao redor de explantes tendinosos dacauda84,85. Neste artigo, as etapas de fabricação necessárias são fornecidas em detalhes, juntamente com leituras úteis que podem ser obtidas pela co-cultura de explantes de núcleo (compartimento intrínseco) dentro de um hidrogel de colágeno tipo 1 carregado de células endoteliais (compartimento extrínseco).
Protocol
Todos os métodos descritos aqui foram aprovados pelas autoridades responsáveis (números de licença do Cantão de Zurique ZH104-18 e ZH058-21). Uma visão geral é apresentada na Figura 1.
1. Isolamento de componentes do tendão assemblóide de camundongos com 12-15 semanas de idade (i.e., B6/J-Rj)
- Eutanásia dos camundongos por asfixia induzida por gás CO2 . Para maximizar o rendimento, não processe mais de 3 ratos de cada vez e proceda ao isolamento celular imediatamente após a eutanásia.
- Garantir a morte por indução bilateral de pneumotórax.
- Esterilize a pele do rato com etanol a 80% e mova o rato para uma capa de biossegurança estéril.
- Isolar explantes do núcleo do tendão da cauda.
- Use um bisturi (nº 21) para separar a cauda do rato, cortando-a em sua base.
- Começando pela ponta da cauda, segure-a com a pinça e mexa-a para quebrar a pele. Em seguida, puxe suavemente a pinça para longe da cauda para expor os explantes do núcleo do tendão.
- Coloque os explantes do núcleo do tendão no meio de cultura padrão (DMEM/F12 + 10% FBS + 1% penicilina/estreptomicina + 1% anfotericina + 1% aminoácidos não essenciais) e separe-os da parte da cauda arrancada usando uma lâmina de bisturi fresca (nº 21).
- Repita as etapas 1.4.2. e 1.4.3. até que toda a cauda seja processada e os explantes tendinosos se tornem menores que 25 mm.
- Cortar os explantes isolados do núcleo em pedaços de 25 mm de comprimento utilizando uma lâmina de bisturi fresca (nº 21).
- Meça o diâmetro médio dos explantes do núcleo com um microscópio de luz conectado a um software de análise de imagem através de uma câmera digital acoplável em C.
- Use apontar e clicar para selecionar a ferramenta de medição de linha no lado direito.
- Meça o diâmetro do explante em três locais diferentes e calcule seu diâmetro médio.
- Para facilitar o clampeamento e os ensaios mecânicos posteriores, proceder apenas com explantes de núcleo com diâmetro médio superior a 100 μm.
- A descarga combinada com a exposição às condições padrão de cultura (37 °C, 20% O2, suplementação sérica) altera a expressão gênica dentro de 6 h após o isolamento e resulta em degradação dentro de 7 dias21. Para começar com um estado quase homeostático, produza os assembloides tendinosos e inicie os experimentos imediatamente após o isolamento do núcleo do tendão.
- Dependendo do arranjo experimental, explantes desvitalizados do tendão central são necessários como grupo controle. Para desvitalizar os explantes do núcleo do tendão, congele-os em um pequeno recipiente cheio de nitrogênio líquido por 5 s usando uma pinça e, em seguida, descongele-os por 5 s à temperatura ambiente (TR). Repita este ciclo de congelamento-descongelamento 3 vezes e prossiga com o passo 4 ("Apertando os explantes do núcleo").
CUIDADO: O nitrogênio líquido pode causar queimaduras a frio, asfixia e fragiliza muitos materiais comuns. Use apenas recipientes projetados para líquidos de baixa temperatura e use roupas de proteção (por exemplo, protetor facial, luvas apropriadas, sapatos de tampo fechado).
- Isolar fibroblastos tendíneos.
- Use um bisturi (nº 21) para fazer uma incisão transversal no meio do pé do rato. Faça dois cortes perpendiculares ao pé ao longo das laterais das patas traseiras e até os quadris de cada extremidade desta incisão.
- Use a pinça para fixar o retalho cutâneo no pé e descascar a pele que cobre os músculos da panturrilha. Ao isolar as células endoteliais do mesmo camundongo, remova toda a pele.
- Separe o tendão de Aquiles do osso calcâneo com uma lâmina de bisturi fresca (nº 21). Fixe a extremidade solta do tendão de Aquiles com a pinça e separe a outra extremidade do músculo gastrocnêmio.
- Lave o tendão de Aquiles uma vez em PBS e use o bisturi (nº 21) para remover todo o tecido muscular restante até que reste apenas o tecido do tendão branco. Se as células endoteliais forem isoladas do mesmo rato, deixe o tendão de Aquiles em PBS e continue com o passo 1.6. primeiro.
- Agrupar os tendões de Aquiles de um animal em um tubo plástico de 15 mL contendo 10 mL de meio de digestão do tendão (DMEM/F12 + penicilina a 1%/estreptomicina + anfotericina a 1% + 2 mg/mL de colagenase 1) e digerir por 6-8 h a 37 °C sob agitação lenta e constante usando um agitador orbital de baixa velocidade a 15 rpm.
- Centrifugar a solução digerida do tendão a 500 x g por 5 min no TR, aspirar o sobrenadante e ressuspender em 8 mL de meio de cultura padrão (DMEM/F12 + 10% FBS + 1% penicilina/estreptomicina + 1% anfotericina + 1% aminoácidos não essenciais) e cultura em frasco de cultura T25 em condições padrão de cultura (37 °C, 20% O2) por 7 dias sem troca de meio. Depois disso, troque a mídia uma vez por semana.
- Dividir as células com 80% de confluência em frasco de cultura T150 (1:6). Congelar as células na passagem 2 em 2 mL de meio de congelamento filtrado estéril (70% DMEM/F12 + 20% FBS + 10% DMSO) distribuídos em dois criotubos de 1,5 mL e mantê-los a -80 °C até novo uso. Use tripsina para remover as células do plástico de cultura de tecidos.
- Isolar células endoteliais derivadas do músculo.
- Se os fibroblastos tendinosos não forem isolados do mesmo rato, comece pelos passos 1.5.1 e 1.5.2.
- Use uma tesoura para separar as patas traseiras do corpo, cortando a articulação do quadril.
- Lave as patas traseiras uma vez em PBS frio (~ 4 °C), remova os músculos (quadríceps femoral, extensor longo dos dedos, sóleo e gastrocnêmio) com um bisturi (nº 21) e coloque os músculos em uma placa de Petri no gelo.
- Use uma lâmina de bisturi fresca (No. 21) para picar o tecido muscular em pedaços menores que 1 mm3 , mantendo a placa de Petri no gelo.
- Agrupar o tecido muscular picado de ambas as patas traseiras em um tubo plástico de 50 mL contendo 12,5 mL de meio de digestão muscular (PBS + 2 mg/mL de colagenase IV + 2 mg/mL dispase II + 2 mM de CaCl2).
- Coloque o tubo de plástico num banho-maria a 37 °C durante 10 minutos. Agite vigorosamente a solução e coloque-a de volta por mais 10 min. Repita até que a solução pareça ser homogênea e restem apenas pedaços (brancos) de tendão e fáscia (ca. 4 x 10 min). Enquanto isso, continuar com o isolamento de fibroblastos tendíneos ou o isolamento de macrófagos.
- Adicione 12,5 mL de PBS frio + 10% FBS ao tubo de plástico para interromper a digestão.
- Use um suporte de pipeta acionado por bateria equipado com uma pipeta de 50 mL para aspirar a suspensão do tubo plástico. Equipar o tubo de plástico com um filtro de células de 400 μm e filtrar a suspensão para remover detritos. Repita o processo com um filtro de células de 100 μm.
- Centrifugar a suspensão filtrada a 400 x g por 5 min no RT. Ressuspender em 10 mL de PBS frio + 10% FBS e centrifugar novamente.
- Ressuspender em 8 mL de meio de cultura endotelial (mistura 1:1 de DMEM/F12 e kit endopan 3 + 10 U/mL de heparina + 20% de SFB + 1% de penicilina/estreptomicina + 1% de anfotericina + 30 mg/mL de suplemento de crescimento endotelial) suplementado com puromicina (4 mg/mL) para seleção populacional.
- Semeando as células de um camundongo em um frasco de cultura T25 previamente revestido com 2 mL de uma solução estéril de gelatina a 0,2% por 2 h a 37 °C e, em seguida, seco durante a noite em RT após a remoção do excesso de solução. Preparar os frascos no dia anterior ao isolamento.
- Após 24 h em condições de cultura padrão (37 °C, 20% O2), remover o meio de suplemento de puromicina, lavar as células anexadas uma vez com PBS e cultivá-las em 8 mL de meio de cultura endotelial.
- Passar as células 1:5 em confluência de 80% para frascos revestidos com gelatina e utilizá-las em experimentos até P2. Use uma solução de descolamento celular diferente de tripsina (Tabela de Materiais) para remover as células do plástico de cultura de tecidos e não congelá-las.
- Isolar macrófagos derivados da medula óssea.
- Se os fibroblastos tendinosos ou as células endoteliais não forem isolados do mesmo camundongo, execute as etapas 1.5.1, 1.5.2, 1.6.2 e 1.6.3 primeiro.
- Depois de remover a pele, o tendão e os tecidos musculares, lave os ossos restantes (fêmur e tíbia) uma vez em PBS frio (~4 °C).
- Coloque os ossos em PBS fresco e frio (~4 °C) e use um bisturi (No. 21) para cortar gradualmente as epífises até que a medula óssea seja exposta. Aparece como um ponto vermelho em ambos os lados do osso.
- Equipar uma seringa com uma agulha de injeção de 0,4 mm x 25 mm (G27) e preenchê-la com 10 mL de meio de cultura de macrófagos (DMEM/F12 + 10% FBS + 1% penicilina/estreptomicina + 1% anfotericina + 1% aminoácidos não essenciais).
- Segure um osso após o outro sobre um tubo de plástico de 50 mL, insira a agulha de injeção a cerca de 1 mm de profundidade na medula óssea exposta na parte superior e lave a medula óssea esvaziando a seringa. A medula óssea ruborizada aparece como uma estrutura avermelhada semelhante a um tubo quando suspensa no meio.
- Dissolva a medula óssea pipetando-a suavemente para cima e para baixo repetidamente usando uma ponta de pipeta de 1 mL. Use um suporte de pipeta acionado por bateria equipado com uma pipeta de 50 mL para filtrar a suspensão da célula através de um filtro de células de 100 μm de volta para o tubo de plástico de 50 mL e centrifuga-lo a 350 x g por 5 minutos no RT.
- Retire o sobrenadante, ressuspenda o pellet em 10 mL de tampão de lise de hemácias e centrifugue novamente a 350 x g por 10 min à temperatura ambiente.
- Ressuspender o pellet em 5 mL de meio de cultura de macrófagos (DMEM/F12 + 10% FBS + 1% penicilina/estreptomicina + 1% anfotericina + 1% aminoácidos não essenciais) e semeá-lo em placas de Petri não tratadas com 100 mm de diâmetro (5-8 x 106 células por placa).
- Após 4 h, adicionar 5 mL de meio de cultura de macrófagos suplementado com 40 ng/mL de fator estimulador de colônias de macrófagos (m-CSF) ao meio de cultura celular sem m-CSF (mistura 1:1) para chegar a uma concentração final de 20 ng/mL m-CSF.
- Após 6 dias, use as células em experimentos ou congele-as até novo uso. Use uma solução de descolamento celular diferente de tripsina (Tabela de Materiais) para remover as células do plástico de cultura de tecidos.
Observação : uma vez isoladas, as células não se expandem mais. Os métodos de isolamento celular aqui descritos também funcionam com camundongos e ratos fora da faixa etária indicada.
- Para verificar o fenótipo das populações celulares isoladas com citometria de fluxo, continue com a etapa 6.3.4.
2. Isolamento de colágeno de ratos Wistar ou Sprague-Dawley
- Siga o protocolo de isolamento descrito em detalhes em outra parte86. Ele também funciona com camundongos, embora com um rendimento muito menor.
- Determinar a concentração da solução resultante com um ensaio de hidroxiprolina, avaliar a pureza com SDS-page e conservar a solução a 4 °C até à utilização nas experiências.
3. Produção dos componentes do sistema de cultura
- Imprima em 3D os suportes de grampo, a estação de montagem e os moldes da câmara.
- Carregue o arquivo .stl anexado (Arquivo Suplementar 1) para os suportes de grampo, a estação de montagem e os moldes da câmara no software de fatiamento. Para adaptar os números de objeto conforme necessário, use apontar e clicar para selecionar objetos e copiar e colar para multiplicá-los.
- Pressione Exportar código G (Ctrl-R) para gerar o código G e, em seguida, exportá-lo (Ctrl-G).
- Carregue o código G em uma impressora 3D.
- Use filamentos biocompatíveis não coloridos para o processo de impressão (ou seja, ácido polilático).
- Corte roscas de 3 mm nos orifícios do suporte da braçadeira que irá transportar os parafusos usando um cortador de rosca (Arquivo Suplementar 2 e Arquivo Suplementar 3, furos 1 e 3).
- Coloque pinos de cavilha de aço inoxidável no orifício na parte de trás do suporte da braçadeira (Arquivo Suplementar 2 e Arquivo Suplementar 3, furo 4).
- Esterilize os suportes de fixação e a estação de montagem com luz UV por pelo menos 1 h antes do uso. Não reutilize suportes de braçadeira impressos em 3D.
- Alternativamente, produza os suportes de abraçadeiras e a estação de montagem com polieterimida usando os planos anexados (Arquivo Suplementar 2, Arquivo Suplementar 3 e Arquivo Suplementar 4), que é mais caro, mas permite melhores métodos de esterilização (ou seja, autoclavagem) e uso repetido.
- Fundir as câmaras usando os moldes impressos em 3D.
- Encha os moldes da câmara com silicone.
- Desgaseifique o silicone em uma câmara de vácuo (90 mbar) por 30 min.
- Deixar a solução polimerizar em RT durante a noite ou em uma placa de aquecimento a 70 °C por 1 h, dependendo da resistência ao calor dos filamentos usados para os moldes.
- Retire cuidadosamente as câmaras polimerizadas dos moldes e corte o silicone supérfluo com bisturi (nº 21).
- OPCIONAL: Se os assembloides e, portanto, as câmaras circundantes forem carregados mecanicamente, reforce os orifícios nas câmaras de silicone com tubos ocos feitos de aço inoxidável.
- Usine as abraçadeiras metálicas de aço inoxidável utilizando a planta anexa (Arquivo Suplementar 5).
- Antes de cada uso, lave as abraçadeiras de aço inoxidável, os suportes de pinça de polieterimida, os parafusos e a câmara de silicone.
- Sonicate por 10 min em etanol 80% (EtOH) e água de osmose reversa a 20% (ROW).
- Sonicate por 10 min em EtOH 50% e Isopropanol 50%.
- Enxágue 3x com ROW.
- Sonicate por 10 min em concentrado de limpeza alcalina a 0,5% (ou seja, 3 mL em 600 mL de ROW).
- Sonicate por 10 min em concentrado de limpeza alcalina a 0,5%.
- Deixe em 0,5% de concentrado de limpeza alcalina enquanto agita por 1 h 50 min.
- Enxágue 3x com ROW.
- Sonicate por 10 min em ROW.
- Secar os componentes e autoclavá-los.
4. Fixação dos explantes do núcleo
- Coloque suportes de fixação correspondentes juntamente com uma braçadeira de metal cada na estação de montagem.
- Coloque pedaços de papel autoclavado molhado (4 mm x 25 mm) sobre as braçadeiras de metal e, em seguida, corte o papel ao longo das bordas internas das braçadeiras com um bisturi (nº 21). Corte 2 pedaços de papel adicionais menores (4 mm x 1,5 mm) de outro pedaço de papel e mantenha-os molhados.
- Usando pinças pontiagudas, coloque 8 explantes de núcleo no papel entre as abraçadeiras de metal com seus pontos finais nas abraçadeiras de metal.
- Cubra os pontos finais dos explantes do núcleo com os pedaços de papel menores colhidos (4 mm x 1,5 mm) e, em seguida, coloque grampos de metal sobre eles. Use uma chave de fenda e os pequenos parafusos (M3 x 6 mm) para fixar os explantes do núcleo entre as abraçadeiras metálicas e o suporte da braçadeira.
- Transfira cuidadosamente os explantes do núcleo pinçado para as câmaras de cultura de silicone e preencha essas câmaras com 2 mL de meio de cultura celular padrão (DMEM/F12 + 10% FBS + 1% penicilina/estreptomicina + 1% anfotericina + 1% aminoácidos não essenciais).
- OPCIONAL: Se os conjuntos/câmaras circundantes forem carregados mecanicamente, fixe-os com parafusos adicionais (M3 x 16 mm) no orifício 3 (Arquivo Suplementar 2 e Arquivo Suplementar 3, furo 3).
5. Preparação e fundição do hidrogel de colágeno
- Retirar as células-alvo do plástico de cultura de tecidos com solução de descolamento celular, centrifugar-as a 400 x g por 5 min em RT e ressuspendê-las em 1 mL de meio de cultura padrão.
- Para um assemblóide, são necessários 10 μL de PBS (20x), 1,28 μL de NaOH 1 M (125x), 8,72 μL de água bidestilada (ddH2O, 23x), 80 μL de colágeno-1 (2,5x ou 1,6 mg/mL final) e 100 μL (2x) de meio de cultura padrão (para assembloides core // cell-free) ou suspensão celular. Prepare esses componentes em duas soluções separadas e misture-os apenas imediatamente antes da fundição.
- Solução de reticulação: Junte o PBS, o NaOH, o ddH20 e a suspensão celular de até 12 assemblóides (+10% de margem de segurança) em uma solução de reticulação e mantenha-a em um tubo plástico de 15 mL sobre gelo. Ajustar a concentração da suspensão celular para atingir as seguintes concentrações finais após a mistura das duas soluções: 250.000 células/mL de fibroblastos tendinos, 500.000 células/mL de células endoteliais derivadas do músculo ou 370.000 células/mL de macrófagos derivados da medula óssea.
- Solução de colágeno-1: Agrupar a solução de colágeno-1 necessária para até 12 assembloides (+10% de margem de segurança) em outro tubo plástico de 15 mL e mantê-la no gelo.
- Uma vez que a solução de reticulação e a solução de colágeno-1 estejam prontas sobre gelo, aspirar o meio de cultura celular das câmaras de cultura contendo os explantes de núcleo pinçados.
- Adicione a solução de colagénio-1 à solução de reticulação com uma pipeta de 1000 μL e misture as duas soluções pipetando para cima e para baixo rapidamente sem criar bolhas. Cobrir os explantes individuais do núcleo do tendão com 200 μL da solução mista, pipetando-a nos pomares fornecidos pelas câmaras de silicone.
- Deixe os hidrogéis polimerizarem durante 50 min a 37 °C.
- Preencher cuidadosamente as câmaras de cultura de silicone com 1,5 mL do respectivo meio de co-cultura, pipetando-o nos cantos das câmaras.
- Para co-cultura de núcleo // fibroblastos, preencher DMEM/F12, FBS 10%, aminoácidos não essenciais a 1%, penicilina/estreptomicina a 1%, anfotericina a 1%, ácido L-Ascórbico 200 μM, fator estimulador de colônia de macrófagos a 20 ng/mL.
- Para co-cultura de núcleo // macrófagos, preencher DMEM/F12, FBS 10%, aminoácidos não essenciais a 1%, penicilina/estreptomicina a 1%, anfotericina a 1%, ácido L-Ascórbico 200 μM, fator estimulador de colônia de macrófagos 20 ng/mL.
- Para a co-cultura de células endoteliais core//, preencher a mistura 1:1 de DMEM/F12 e o kit endopan 3 + 10 U/mL de heparina + 20% de SFB + 1% de penicilina/estreptomicina + 1% de anfotericina + 30 mg/mL de suplemento de crescimento endotelial.
- Cultivar os assemblóides nas condições de cultura apropriadas para a hipótese. Para mimetizar um ambiente de nicho semelhante a uma lesão, por exemplo, cultive-os a 37 °C e 20% deO2. Troque o meio de cultura duas vezes em 1 semana. Para evitar infecções, coloque as câmaras em uma grande placa de Petri ou uma caixa estéril antes de colocá-las em uma incubadora.
NOTA: O tempo de cultura depende da hipótese e da configuração da co-cultura. Por exemplo, assemblóides de fibroblastos centrais em um ambiente de nicho semelhante a uma lesão tornam-se mecanicamente instáveis após cerca de 3 semanas.
6. Métodos de leitura disponíveis
- Realizar microscopia de fluorescência, incluindo ensaios de viabilidade e morfologia.
- Em geral, os assembloides podem ser fotografados como montagens completas. Para isso, retire os assembloides das braçadeiras, cortando-as com tesouras próximas às pinças e transferindo-as para uma placa de 12 poços.
- Lave os assembloides uma vez com PBS.
- Se a análise de viabilidade for realizada, corar cada assemblóide com 100 μL de homodímero de etídio 4 x 10−6 M em PBS (EthD-1) por 20 min a 37 °C no escuro.
- Lavar os assemblóides 3x com PBS e, em seguida, fixá-los com 500 μL de formaldeído a 4% cada por 20 min em RT.
CUIDADO: O formaldeído a 4% tem efeitos alergênicos, carcinogênicos e mutagênicos, é tóxico para a reprodução e pode causar toxicidade ao desenvolvimento (reprotoxic) ou danos aos órgãos. Use roupas e luvas de proteção, proteção ocular e máscara ou outra proteção respiratória. - Lave os assemblóides 3x com PBS e continue com o protocolo de coloração de escolha. Uma seleção de colorações foi descrita anteriormente84,85.
OBS: Evitar colorações que utilizem fluoróforos com comprimento de onda de emissão próximo ao da autofluorescência do colágeno (em torno de 480 nm).
- De acordo com as instruções do fabricante, execute o isolamento de RNA compartimento-específico para RT-qPCR ou sequenciamento de RNA em todo o genoma.
- Retire os assemblóides das braçadeiras com tesoura.
- OPCIONAL: Utilizar pinça para separar os explantes do núcleo do compartimento extrínseco de hidrogel.
- Pool 20-24 explantes de núcleo de 20 mm ou hidrogéis de colágeno carregados de 2 células para isolar quantidades suficientes de RNA.
- Use 1 mL de trizol frio e ruptura mecânica (ou seja, esferas metálicas ou moagem criogênica) para destruir a matriz extracelular dos explantes de núcleo agrupados ou os hidrogéis de colágeno agrupados.
CUIDADO: Toxicidade oral, dérmica e por inalação. Causa irritação na pele e nos olhos. Apenas manuseie com luvas e em um armário de segurança química. - Continuar com o isolamento de RNA do lisado celular usando kits de extração de RNA padrão, conforme descrito anteriormente ou conforme descrito nas instruções do fabricante 84,85.
- Citometria de fluxo compartimento-específica.
- Retire os assemblóides das braçadeiras com tesoura.
- OPCIONAL: Utilizar a pinça para separar os explantes do núcleo do compartimento extrínseco do hidrogel.
- Compartimentos de digestão em 1 mL de PBS com colagenase I (3 mg/mL) e dispase II (4 mg/mL) por 4 h a 37 °C sob agitação constante.
- Centrifugar a solução digerida a 500 x g durante 5 min em RT e aspirar o sobrenadante.
- Ressuspender o pellet em 100 μL de tampão FACS (FBS a 1% em PBS) contendo os anticorpos conjugados a fluoróforos de escolha. Uma seleção de anticorpos conjugados a fluoróforos em funcionamento foi descrita anteriormente84,85.
- Incubar a solução corante por 30 min em TR.
- Diluir a solução corante com 1,4 ml de tampão FACS e centrifugar-a durante 5 minutos a 500 x g em RT.
- Ressuspender o pellet em 350 μL de tampão FACS e filtrar a solução através de uma tampa de filtro de malha de nylon de 100 μm antes de analisá-la com o citômetro de fluxo de escolha de acordo com as instruções do fabricante.
- Analise o sobrenadante.
- Substituir o meio de co-cultura por meio de co-cultura sem soro 3 dias antes da coleta do sobrenadante.
- Realizar análise imediata e tardia do sobrenadante enriquecido e não diluído com kits de ensaio imunoenzimático (ELISA) e de descoberta em escala meso (MSD). Para a análise tardia, armazenar o sobrenadante em tubos plásticos de 1,5 mL a -80 °C.
- Avaliar as propriedades mecânicas do assemblóide.
- Use um dispositivo de alongamento feito sob medida para aplicar forças mecânicas e medir propriedades mecânicas22. Os pinos de aço inoxidável e os parafusos de aço inoxidável tornam as abraçadeiras acopláveis a outros dispositivos de alongamento também.
- Como as propriedades mecânicas do assemblóide são largamente determinadas pela do explante de núcleo embutido 18, meça as propriedades mecânicas do explante de núcleo antes de incorporá-lo em um hidrogel para diminuir o risco de destruir o hidrogel recém-fundido no processo de medição.
Representative Results
Isolamento de componentes (Figura 1 e Figura 2)
Antes de utilizar os explantes centrais e as populações celulares em co-cultura assemblóide, esses componentes devem ser verificados ao microscópio (Figura 1). Os explantes do núcleo devem ter um diâmetro uniforme (100-200 μm) e não apresentar torções ou rugas visíveis. As células endoteliais devem apresentar uma forma alongada em contato com outras células, o que não ocorre quando semeadas em densidade muito baixa devido ao baixo rendimento inicial do isolamento. Nesse caso, as células endoteliais assumem uma forma mais arredondada, com extensões citoesqueléticas e proliferam marcadamente mais lentamente. Divida-os 1:5 após 7-10 dias. Os fibroblastos de tendão isolados dos tendões de Aquiles assumem uma morfologia mais arredondada em comparação com seus homólogos humanos dentro de 1-2 passagens (10-14 dias cada) quando foram divididos 1:6. Os macrófagos são muito menores que os fibroblastos ou as células endoteliais e não proliferam após o isolamento. Dependendo do lote, sua forma pode variar de piramidal a redonda.
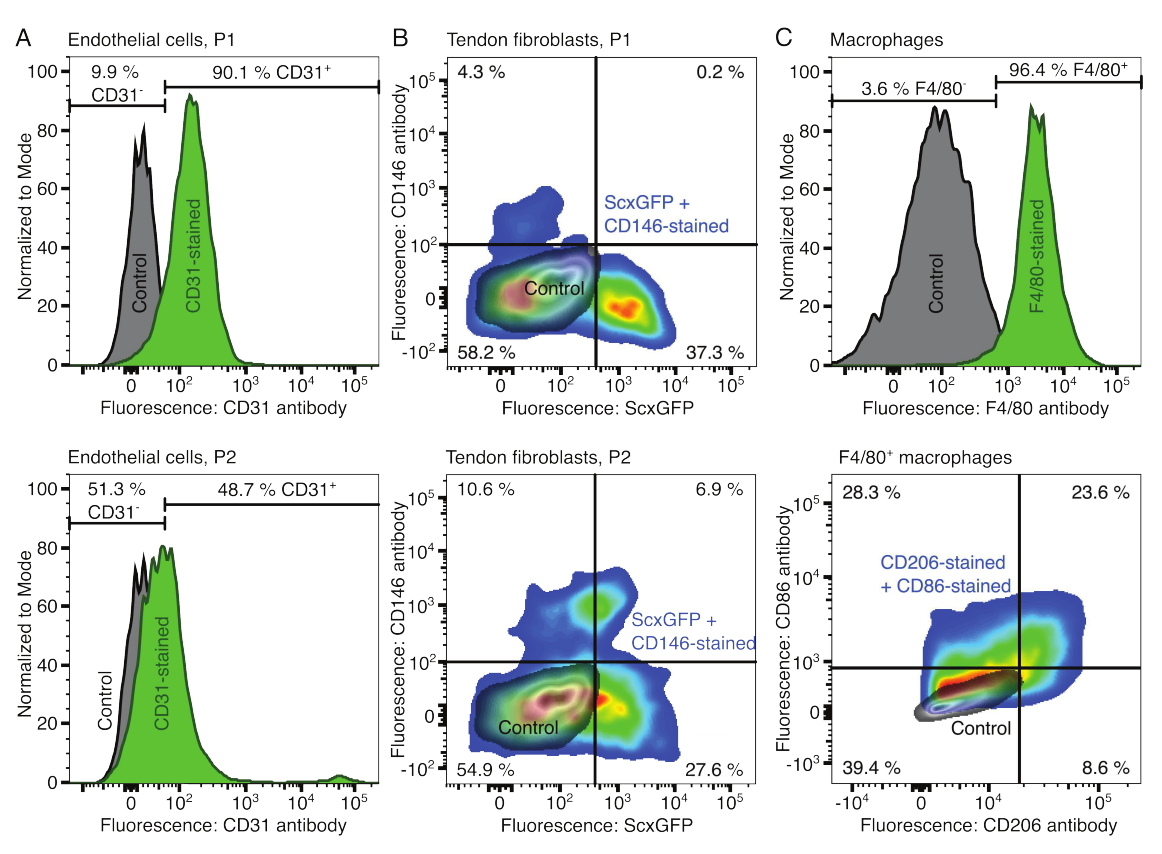
Os fenótipos dos componentes celulares foram verificados por citometria de fluxo. Um anticorpo CD31 conjugado foi utilizado como marcador para células endoteliais (Figura 2A). Definindo o limiar de fluorescência com base em uma amostra controle não corada (cinza), 90,1% das células endoteliais da passagem 1 (P1) e 48,7% da passagem 2 (P2) foram identificadas como CD31-positivas. Uma linhagem de camundongos geneticamente modificada co-expressando o marcador de fibroblastos tendíneos Scleraxis juntamente com uma proteína fluorescente verde (ScxGFP) e um anticorpo CD146 conjugado foi usada para caracterizar os fibroblastos tendíneos (Figura 2B)35,60. Após uma passagem (P1), 37,3% dos fibroblastos eram ScxGFP+CD146-, 0,2% ScxGFP+CD146+, 4,3% ScxGFP-CD146+ e 58% ScxGFP-CD146-. Após duas passagens (P2), a porcentagem de células ScxGFP+CD146- diminuiu para 27,6%, a porcentagem de células ScxGFP+CD146+ aumentou para 6,9%, a porcentagem de células ScxGFP-CD146+ aumentou para 10,6% e a porcentagem de células ScxGFP-CD146- diminuiu para 54,9%. Para identificar e caracterizar os macrófagos, foi utilizado um anticorpo F4/80 associado a um anticorpo CD86 e um anticorpo CD206 (Figura 2C). Após isolamento e cultura, 96,4% das células derivadas da medula óssea foram positivas para F4/80. Dentre essas células F4/80 positivas, 8,6% eram CD206+CD86-, 23,6% CD206+CD86+, 28,3% CD206-CD86+ e 39,4% CD206-CD86-. A velocidade de reticulação do colágeno pode variar de lote para lote e deve ser testada antes do início dos experimentos.
Aspecto assemblóide (Figura 3)
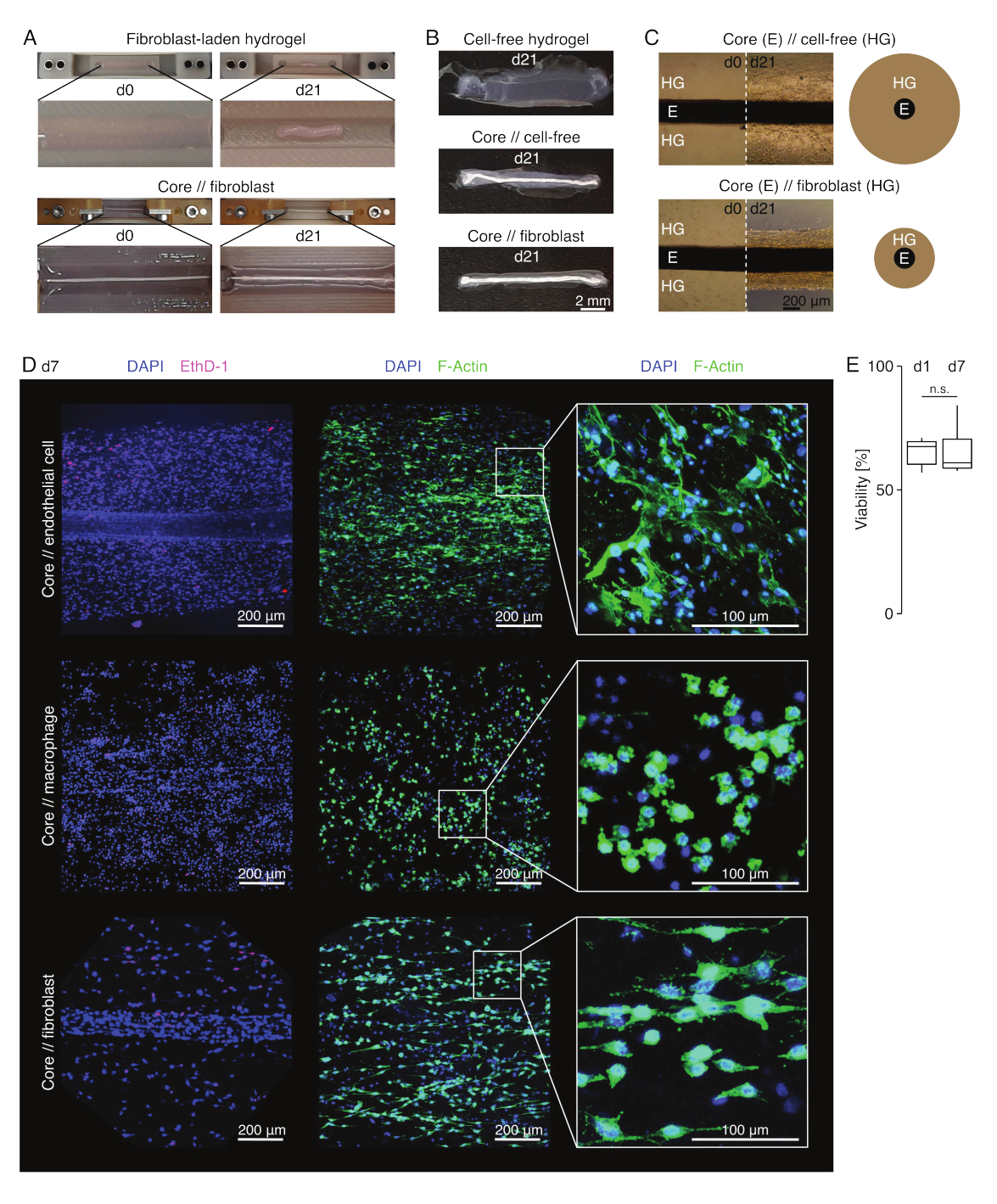
Em condições de cultura semelhantes a lesões (36 °C, 20% deO2), o explante do núcleo permaneceu mecanicamente esticável, não mudou de aparência e permaneceu visualmente distinguível e fisicamente separável do hidrogel circundante por pelo menos 21 dias (Figura 3A,B). O hidrogel circundante foi compactado ao longo do tempo, com a velocidade de compactação dependendo da população celular semeada nele. Os fibroblastos derivados do tendão de Aquiles contraíram seu hidrogel circundante mais rapidamente e o fizeram radialmente quando em um hidrogel fundido ao redor de um explante de núcleo e em todas as direções quando não (Figura 3B,C). Inicialmente, hidrogéis livres de células colocados ao redor de um núcleo também foram compactados. Esta contração foi provavelmente causada pela migração de células do explante central, indicando uma interface intercompartimental dinâmica. Como os hidrogéis livres de células sem um explante de núcleo embutido não compactaram detectavelmente, a contribuição da retração induzida pela perda de água parece ser desprezível (Figura 3B e Arquivo Suplementar 6).
A falta de compactação do hidrogel pode, portanto, ser usada para detectar erros na montagem do assemblóide (ou seja, baixas concentrações de células) e deve ser verificada antes de continuar com métodos de leitura mais caros. Ao estabelecer este método, erros comuns ao reduzir a concentração celular incluíram células moribundas no hidrogel extrínseco porque foram deixadas por muito tempo na solução de reticulação relativamente dura (pH alto, baixa temperatura) e explantes de núcleo de secagem porque o tempo entre a aspiração média e a injeção de hidrogel foi muito longo, ou porque o explante do núcleo foi fixado muito alto para ser incorporado no colágeno.
Microscopia de fluorescência confocal: Análise de viabilidade e morfologia (Figura 3)
Uma vez removidas das pinças com tesoura (Figura 3B), as assembloides podem ser fixadas, coradas e imageadas com um microscópio confocal como um todo, sem seccionamento. Neste contexto, os assemblóides core // endothelial cell, core // macrophage core // fibroblastos foram corados com DAPI (NucBlue) e Ethidium Homodimer (EthD-1) para analisar a viabilidade e DAPI e F-actina para analisar a morfologia e a dispersão celular no hidrogel de colágeno 3D (Figura 3D). A viabilidade dos assemblóides de células endoteliais core // (Figura 3E) foi quantificada e encontrada geralmente é menor após a montagem do assembloid do que previamente relatada para os assembloides core // macrófago e core // fibroblastos84. No entanto, a viabilidade manteve-se estável durante a cultura assemblóide até pelo menos o 7º dia.
Microdano induzido mecanicamente e medição das propriedades mecânicas (Figura 4)
Os parafusos e pinos fixados aos suportes de pinça permitem a fixação dos conjuntos pinçados a dispositivos de alongamento uniaxiais. O dispositivo de alongamento sob medida utilizado é equipado com célula de carga de 10 N e já foi descrito em publicações anteriores (Figura 4A)22. Todas as amostras foram pré-condicionadas com cinco ciclos de alongamento a 1% de deformação antes das medidas.
O registro da curva tensão-deformação completa dos explantes ou assemblóides do núcleo (Figura 4B) permitiria quantificar o módulo elástico linear (α), a tensão máxima (β) e a deformação máxima (у). No entanto, também danifica irreversivelmente o explante do núcleo ou o assemblóide, o que impossibilita avaliar o desenvolvimento longitudinal da tensão máxima (β) e da deformação máxima (у) para as mesmas amostras (Figura 4B). Aqui, o módulo de elasticidade linear foi utilizado como medida da capacidade da amostra em suportar forças, uma vez que essa medida requer o alongamento da amostra até apenas 2% de deformação, o que já foi demonstrado anteriormente não causar reduções permanentes no módulo de elasticidade linear18. Em particular, assemblóides de células endoteliais centrais // foram expostos ao procedimento de pinçamento a 2% de deformação (aproximadamente o final da região elástica linear) ou 6% de deformação (aproximadamente a deformação máxima). O microdano resultante foi avaliado medindo-se o módulo de elasticidade linear antes e após o procedimento (Figura 4C).
De acordo com experimentos previamente conduzidos explorando explantes de núcleo monocultivados, assemblóides de células endoteliais do núcleo // mantiveram seu módulo de elasticidade linear por pelo menos 14 dias quando cultivados em condições de nicho quase homeostático (29 °C, 3% O2) e expostos a cepas não superiores a 2%18,21. Em relação à estimulação mecânica basal, o estiramento estático aplicado através das pinças pareceu mimetizar suficientemente os níveis de deformação nativa experimentados pelas unidades centrais do tendão in vivo para prevenir processos catabólicos geralmente associados à descarga da matriz87. De fato, o declínio progressivo e estatisticamente significativo do módulo de elasticidade linear observado em assemblóides de células endoteliais centrais expostas a 6% de deformação pode ser atribuído ao descarregamento da matriz decorrente de microdanos à matriz induzidos mecanicamente.
Ao realizar esses experimentos, é importante evitar a secagem do assemblóide. Aqui, eles foram envoltos em papel autoclavado e molhado, mas outros métodos também poderiam ser viáveis dependendo de sua compatibilidade com o dispositivo de alongamento usado. Como o atrito entre as abraçadeiras metálicas e o explante do núcleo é limitado, adicione pequenos pedaços de papel entre o metal e o explante do núcleo durante a fixação para evitar o deslizamento e monitore de perto o processo de alongamento para detectar e excluir explantes e assembloides de núcleo deslizados.
Análise do transcriptoma compartimento-específico e do secretoma assemblóide-específico (Figura 5 e Figura 6)
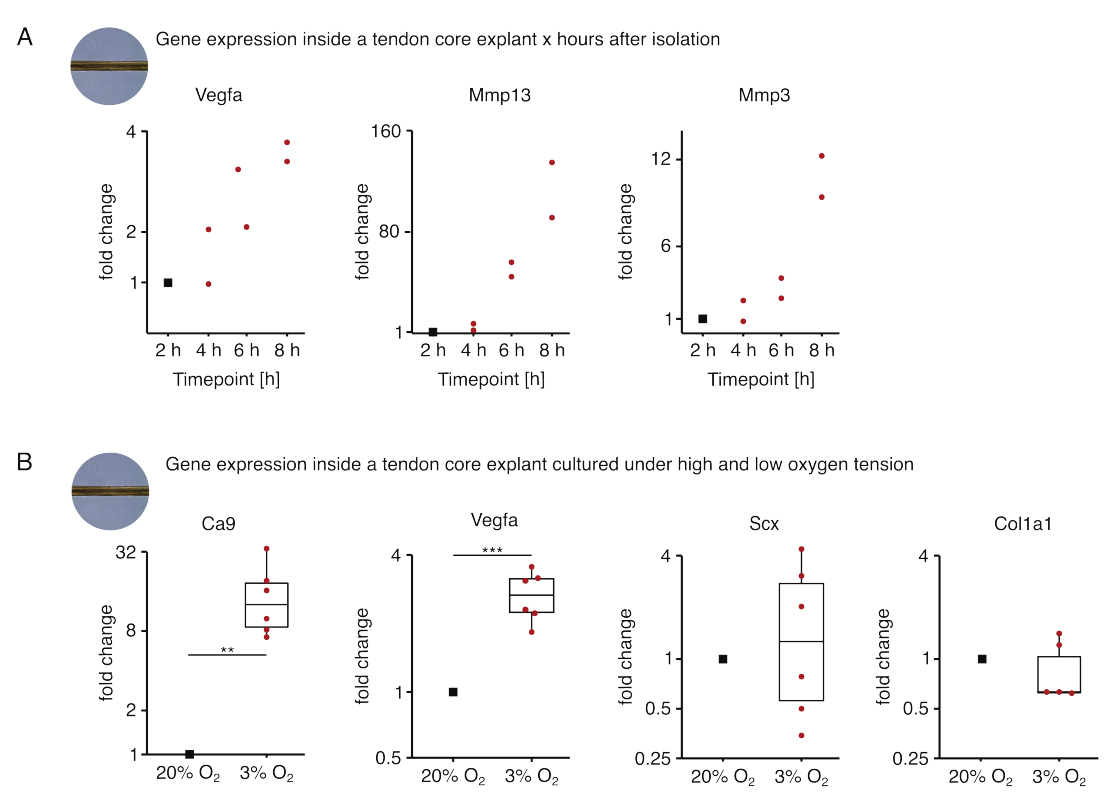
No primeiro conjunto de experimentos de monocultura de núcleo aqui apresentados, a estabilidade da expressão do gene central após o isolamento de explantes foi avaliada para dissociar o isolamento dos efeitos experimentais (Figura 5A). Embora um maior número de réplicas seja necessário para conclusões precisas, a expressão de Vegfa e Mmps aumentou fortemente em explantes centrais recém-isolados dentro de horas após o isolamento do explante, quando cultivados em condições de nicho semelhante a lesões (37 °C, 20% O2).
A neovascularização é uma característica central da doença e do reparo tendíneo que poderia, em parte, ser conduzida por células endoteliais ativadas por fatores pró-angiogênicos (isto é, fator de crescimento endotelial vascular, Vegfa) secretados pelo núcleo do tendão sob hipóxia88. Examinando a primeira etapa desse potencial crosstalk (Figura 5B), a expressão tanto do Vegfa quanto do marcador de hipóxia anidrase carbônica 9 (Ca9) mostrou-se aumentada estatisticamente significativa em explantes monocultivados sob baixa tensão de oxigênio (3% O2) em contraste com aqueles monocultivados sob alta tensão de oxigênio (20% O2). Enquanto isso, a menor tensão de oxigênio parece não causar alterações na expressão de marcadores de fibroblastos tendíneos, como Scleraxis (Scx) e colágeno-1 (Col1a1). Juntos, esses resultados identificam as células residentes no núcleo como contribuintes plausíveis para a sinalização pró-angiogênica em um nicho hipóxico.
Em seguida, a ativação das células endoteliais pela sinalização central pró-angiogênica foi avaliada em co-cultura núcleo // células endoteliais assembloides sob alta (20% O2) e baixa (3% O2) tensão de oxigênio. Felizmente, a composição modular dos assemblóides permite a análise compartimento-específica do transcriptoma após a cultura, separando fisicamente o explante do núcleo do hidrogel extrínseco de colágeno (Figura 6A). No explante central (Figura 6D), a expressão de Vegfa foi novamente confirmada como aumentando sob baixa tensão de oxigênio, embora o efeito sobre outros marcadores hipóxicos, como Fgf2 , tenha sido menos claro e exija números de réplicas mais altos para conclusões precisas. Além disso, a expressão de marcadores pró-inflamatórios, como o Tnf-α , e de degradação da matriz extracelular, como Mmp3 , estavam diminuídos no núcleo sob baixa tensão de oxigênio. No hidrogel extrínseco inicialmente semeado com células endoteliais (Figura 6E), a presença de um explante vivo do núcleo (aC) diminuiu a expressão de Vegfa sob baixa tensão de oxigênio, mas não sob alta tensão de oxigênio. Além disso, a presença de um explante central desvitalizado (dC) sob baixa tensão de oxigênio também não diminuiu a expressão de Vegfa . Sob baixa tensão de oxigênio, a expressão de Tnf-α no hidrogel extrínseco foi comparável em torno de aC/dC, mas aumentou sob alta tensão de oxigênio ao redor de explantes de núcleo vivo. A expressão de Fgf2 foi diminuída em todas as condições em comparação com o hidrogel extrínseco carregado de células endoteliais cultivado ao redor de um explante central desvitalizado sob alta tensão de oxigênio, mas a maioria sob baixa tensão de oxigênio. A expressão de Mmp3 foi maior ao redor de explantes de núcleo vivo sob alta tensão de oxigênio e menor ao redor de explantes de núcleo desvitalizados sob baixa tensão de oxigênio. Em geral, as células endoteliais co-cultivadas parecem responsivas tanto ao explante ativo do núcleo, que é capaz de iniciar crosstalk quanto a variações nos níveis de oxigênio. Uma análise mais abrangente do transcriptoma facilitaria a elucidação de suas respectivas contribuições.
A modularidade do sistema assemblóide permite a integração de células geneticamente modificadas contendo genes fluorescentes repórteres. Aqui, células endoteliais isoladas de camundongos Pdgfb-iCreER mG89 foram semeadas no compartimento hidrogel. Essas células co-expressam a subunidade b do fator de crescimento derivado de plaquetas marcador de células endoteliais (Pdgfb) ao lado da proteína fluorescente verde reforçada (EGFP), o que faz com que as células endoteliais que expressam Pdgfb pareçam verdes sob a microscopia (Figura 6C). Usando este método, confirmou-se que a presença de células endoteliais expressas por Pdgfb foi mantida por 7 dias em cultura (37 °C) e pareceu ser independente da tensão de oxigênio (20% O2 comparado a 3% O2).
Para análise do secretoma de assemblóides, o meio de cultura utilizado respectivamente para core // cell-free e core // fibroblasto, core // macrófago ou core // co-cultura de células endoteliais foi substituído por seu equivalente livre de soro três dias antes de aspirar e congelar o sobrenadante agora enriquecido com o secretoma (Figura 6A). Esse tempo de enriquecimento foi suficiente para detectar citocinas como o fator de crescimento endotelial vascular (VEGF) com um ensaio de DME, como mostrado aqui para explantes centrais e assembloides de fibroblastos do núcleo // cultivados em condições de nicho semelhante a lesão (Figura 6B).
Considerações importantes ao analisar os secretomas e transcriptomas de explantes centrais e assemblóides dizem respeito ao uso de controles adequados. Explantes de núcleo recém-isolados têm valor limitado, pois especialmente sua expressão de Vegfa e Mmps aumenta fortemente dentro de horas após o isolamento (Figura 5A). Explantes combinados no tempo cercados por um hidrogel inicialmente livre de células são mais adequados como controles para a expressão gênica do compartimento central. Para o hidrogel extrínseco, hidrogéis carregados de células cultivados sem um explante de núcleo são controles inferiores em comparação com hidrogéis carregados de células cultivados ao redor de explantes de núcleo desvitalizados (Arquivo Suplementar 7), principalmente porque compactam em formas arredondadas em vez de hidrogéis alongados, o que altera muito a morfologia celular (Figura 3A).

Figura 1: Isolamento e montagem de componentes assemblóides para modelar crosstalk in vivo . Explantes tendíneos foram extraídos da cauda de camundongos, cortados e grampeados. Músculos da perna de camundongo (isto é, quadríceps femoral (QF), gastrocnêmio (G) e tibial anterior (TA)) foram digeridos para isolar células endoteliais que foram então cultivadas em plástico de cultura de tecidos. Os tendões de Aquiles (TA) também foram digeridos para isolar os fibroblastos tendíneos, os quais foram então cultivados em cultura de tecidos plásticos. A medula óssea da tíbia e do fêmur foi liberada para fora dos ossos. Em seguida, os monócitos isolados foram cultivados em plástico de cultura de tecidos e diferenciados em macrófagos virgens. As imagens de microscopia óptica (10x) mostram o aparecimento de explantes centrais, células endoteliais, fibroblastos tendinosos e macrófagos imediatamente antes de sua integração em assemblóides. Durante a montagem, as células cultivadas em plástico foram colocadas em suspensão e, em seguida, semeadas em solução de colágeno-1 (1,6 mg/mL). Em seguida, a mistura célula-hidrogel foi lançada ao redor do explante do núcleo pinçado e polimerizada por 50 min a 37 °C antes da adição do meio de cultura. As condições de cultura foram controladas através das pinças (tensão mecânica) e dos ajustes da incubadora (concentração de oxigênio, temperatura). Clique aqui para ver uma versão maior desta figura.
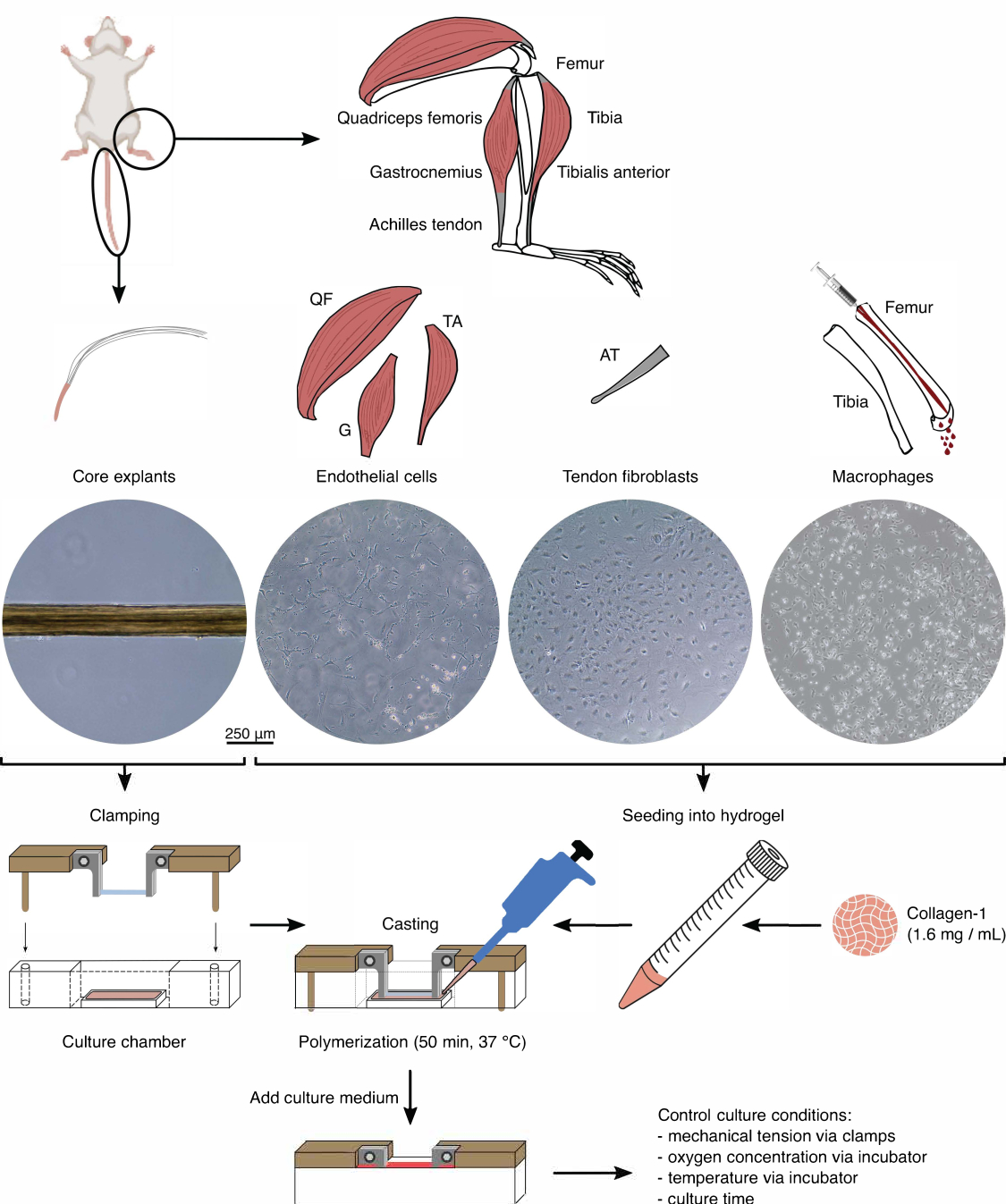{kind=link}

Figura 2: Caracterização dos componentes do assemblóide celular. (A) Análise representativa por citometria de fluxo de células endoteliais derivadas do músculo após uma passagem (P1, fileira superior) e duas passagens (P2, fileira inferior). As contagens de células não coradas (cinza) e CD31 (verde) foram normalizadas para modal. As porcentagens são dadas para o grupo corado com CD31. (B) Análise representativa por citometria de fluxo de fibroblastos derivados do tendão de Aquiles após uma passagem (P1, fileira superior) e duas passagens (P2, fileira inferior). Os eixos relatam intensidades de fluorescência de células não coradas (cinza) e células que expressam ScxGFP e coradas com anticorpos CD146 (cores do arco-íris). (C) Análise representativa por citometria de fluxo de macrófagos derivados da medula óssea após cultura. Na linha superior, as contagens de células não coradas (cinza) e F4/80 (verde) foram normalizadas para modal. As porcentagens são dadas para o grupo corado com F4/80. O gráfico na linha inferior relata intensidades de fluorescência de células não coradas (cinza) e o subconjunto F4/80+ de células coradas com anticorpos CD206 e CD86 (cores do arco-íris). Clique aqui para ver uma versão maior desta figura.
{kind=link}

Figura 3: Imagem e aparência das assembleias. (A) Fotografias representativas tiradas nos dias 0 (d0) e 21 (d21) de cultura (37 °C, 20% O2) mostram uma contração multidimensional de um hidrogel contendo fibroblastos extrínsecos sem um explante de núcleo embutido e forte compactação radial de um hidrogel contendo fibroblastos extrínsecos ao redor de um explante de núcleo. (B) Fotografias representativas tiradas no dia 21 (d21) de cultura (37 °C, 20% O2) mostram diferenças na velocidade de compactação entre hidrogéis livres de células, hidrogéis livres de células fundidos em torno de um explante de núcleo e hidrogéis carregados de fibroblastos de tendão fundidos em torno de um explante de núcleo. (C) As imagens representativas de microscopia óptica (10x) obtidas no dia 0 (d0) e no dia 21 (d21) de cultura (37 °C, 20% O2) indicam mudanças longitudinais na presença de populações celulares e na velocidade de compactação do hidrogel de colágeno (HG) ao redor do explante do núcleo (E) em co-cultura de núcleo // livre de células e núcleo // conjunto de fibroblastos assemblóides. A representação esquemática mostra as diferenças na compactação de hidrogel entre núcleo // assemblóide livre de células e núcleo // co-cultura de assemblóide de fibroblastos. (D) Imagens representativas de microscopia confocal obtidas no dia 7 (d7) de co-cultura núcleo // célula endotelial, núcleo // macrófago e núcleo // núcleo (37 °C, 20 % O2). Imagens na fileira da esquerda mostram assemblóides com núcleos celulares corados em azul (DAPI) e células mortas coradas em rosa (Ethidium homodimer-1). As outras duas fileiras representam assemblóides com núcleos celulares corados em azul (DAPI) e filamentos de actina em verde (F-actina). (E) Boxplots mostrando a viabilidade quantificada de núcleos // assembloides de células endoteliais no dia 1 (d1) e no dia 7 (d7) de co-cultivo. N = 5. As dobradiças superior e inferior correspondem ao primeiro e terceiro quartis(percentis 25e 75) e a média à mediana. Os bigodes se estendem da dobradiça superior/inferior até o maior/menor valor não superior a 1,5 vezes o intervalo interquartil. Valores de p: n.s.p > 0,05. Clique aqui para ver uma versão maior desta figura.
{kind=link}

Figura 4: Estimulação mecânica dos assemblóides e medição das propriedades mecânicas dos assemblóides. (A) Representação gráfica do dispositivo de alongamento feito sob medida, compreendendo as plataformas do suporte da braçadeira, um sensor de força e um motor de passo. A imagem fotográfica mostra um assemblóide montado no dispositivo de alongamento com grampos. A tampa de um tubo plástico de 15 mL (Ø: 17 mm) utilizado para a balança. (B) Gráfico representando curvas de tensão/deformação representativas para explantes de núcleo (azul claro) e assemblóides (vermelho claro). O módulo elástico linear (α), a tensão máxima (β) e a deformação máxima (у) podem ser extraídos dos dados para caracterizar mecanicamente o explante do núcleo ou assemblóide. (C) Gráfico mostrando o módulo elástico linear (Emod) de assemblóides de células endoteliais de núcleo // co-cultivados (29 °C, 3% O2) durante um curso de tempo de 14 dias após serem pinçados (linha sólida), pinçados e esticados até 2% L0 deformação (linha pontilhada), ou pinçados e esticados até 6% L0 deformação (linha tracejada) no início do experimento. N = 5. Os pontos de dados foram normalizados para o módulo de elasticidade linear inicial antes do alongamento e são todos apresentados como média (±sem). Valores de p: *p < 0,05, **p < 0,01. Clique aqui para ver uma versão maior desta figura.
{kind=link}

Figura 5: Alterações no transcriptoma central após isolamento e cultivo sob diferentes condições de nicho. (A) Gráfico de dispersão mostrando as mudanças nas dobras na expressão dos genes Vegfa, Mmp13 e Mmp3 em explantes de núcleo murino monocultivados (37 °C, 20% O2) 2 h, 4 h, 6 h e 8 h após isolá-los da cauda. As alterações de dobra nos respectivos momentos foram normalizadas para a expressão gênica 2 horas após o isolamento. N = 2. (B) Boxplots mostrando as alterações de dobras na expressão dos genes Ca9, Vegfa, Scx e Col1a1 em explantes centrais monocultivados sob baixa tensão de oxigênio (3% O2) normalizados e comparados àqueles monocultivados sob alta tensão de oxigênio (20% O2). N = 5-6. As dobradiças superior e inferior dos boxplots correspondem ao primeiro e terceiro quartis(percentis 25e 75) e o médio à mediana. Os bigodes se estendem da dobradiça superior/inferior até o maior/menor valor não superior a 1,5 vezes o intervalo interquartil. Os pontos de dados usados para normalização são representados como pontos pretos e pontos de dados individuais como pontos vermelhos. Valores de p: **p < 0,01, ***p < 0,001. Clique aqui para ver uma versão maior desta figura.
{kind=link}

Figura 6: Análise do secretoma assemblóide-específico e transcriptoma compartimento-específico. (A) Fotografia representativa mostrando o assemblóide no dia 7 (d7), quando amostras de secretoma e transcriptoma foram coletadas, e representação do fluxo de trabalho subjacente. (B) Concentração de VEGF (pg/mL) no sobrenadante dos assemblóides de núcleo // livre de células e núcleo // fibroblastos após 7 dias de co-cultivo (37 °C, 20% O2) representados como boxplots. N = 6. (C) Imagens representativas de microscopia confocal de assemblóides de células endoteliais centrais // após 7 dias de co-cultura (37 °C) sob alta tensão de oxigênio (20% O2) e baixa tensão de oxigênio (3% O2). Os núcleos celulares são corados em azul (DAPI), e as células endoteliais incorporadas co-expressam proteína fluorescente verde aumentada (EGFP) ao lado da subunidade b do fator de crescimento derivado de plaquetas marcador de células endoteliais (Pdgfb). A linha pontilhada indica a interface compartimental entre o explante central (E) e o hidrogel carregado de células endoteliais (HG). (D) Gráfico de dispersão mostrando as alterações nas dobras na expressão dos genes Vegfa, Tnf-α, Fgf2 e Mmp3 no compartimento central de assembloides de células endoteliais centrais // co-cultivadas sob baixa tensão de oxigênio (3% O2) normalizadas e comparadas àquelas cultivadas sob alta tensão de oxigênio (20% O2). N = 2. (E) Gráfico de dispersão mostrando as alterações de dobras na expressão dos genes Vegfa, Tnf-α, Fgf2 e Mmp3 no compartimento extrínseco de assemblóides de células endoteliais com núcleo vivo (aC) ou núcleo desvitalizado (dC) co-cultivados sob alta tensão de oxigênio (20% O2) e baixa tensão de oxigênio (3% O2). As alterações de dobra nas respectivas condições foram normalizadas para o compartimento extrínseco de um núcleo // assemblóide de células endoteliais com um núcleo desvitalizado (dC) co-cultivado sob alta tensão de oxigênio (20%O2). N = 3-4. Em B, as dobradiças superior e inferior dos boxplots correspondem ao primeiro e terceiro quartis(percentis 25e 75) e o médio à mediana. Os bigodes estendem-se da dobradiça superior / inferior ao maior / menor valor não mais do que 1,5 vezes o intervalo interquartil. Outliers são representados como pontos pretos. Valores de p: *p < 0,05. Em D e E, os pontos de dados usados para normalização são representados como pontos pretos, e pontos de dados individuais são representados como pontos vermelhos. Clique aqui para ver uma versão maior desta figura.
{kind=link}
Tabela 1: Requisitos de entrada para sistemas modelo de doença e lesão tendínea. Uma lista de gatilhos primários de doenças tendinosas e direcionadores secundários correspondeu a uma seleção de parâmetros de entrada cuja tratabilidade é central para modelar a doença e a lesão tendíneas. Clique aqui para baixar esta tabela.
Tabela 2: Requisitos de saída para sistemas modelo de doença e lesão tendínea. Uma seleção de características da doença tendínea correspondeu a uma seleção de parâmetros de saída cuja quantificabilidade é central para a interpretação do comportamento do modelo de doença e lesão do tendão. Clique aqui para baixar esta tabela.
Arquivo Suplementar 1: arquivo .stl para os suportes de grampos, a estação de montagem e os moldes da câmara. Clique aqui para baixar este arquivo.
Arquivo Suplementar 2: Plano do titular da braçadeira de direito. Clique aqui para baixar este arquivo.
Arquivo Suplementar 3: Plano de suporte de braçadeira esquerda. Clique aqui para baixar este arquivo.
Arquivo Suplementar 4: Plano da plataforma de montagem Clique aqui para baixar este arquivo.
Arquivo Suplementar 5: Plano de grampos metálicos. Clique aqui para baixar este arquivo.
Arquivo Suplementar 6: Imagem mostrando encolhimento de hidrogel livre de células. Clique aqui para baixar este arquivo.
Arquivo Suplementar 7: Imagem mostrando um explante de núcleo desvitalizado. Clique aqui para baixar este arquivo.
Discussion
De modo geral, o sistema de modelos assemblóides aqui apresentado tem várias etapas críticas a destacar. Primeiro, o sistema de modelos é tão bom quanto a qualidade de seus componentes. É vital verificar o explante do núcleo e as populações de células a serem semeadas sob o microscópio antes de iniciar o processo de montagem. Da mesma forma, é importante verificar o fenótipo das populações celulares isoladas pelo menos uma vez com citometria de fluxo. Especialmente quando um novo lote de colágeno-1 é usado pela primeira vez, é vantajoso verificar a velocidade de reticulação em uma execução experimental antes de incorporar células nele. A montagem assemblóide requer muito manuseio manual, o que aumenta o risco de infecções. Para minimizar o risco de infecções, trabalhe em uma capela de biossegurança estéril com fluxo de ar laminar, troque luvas com frequência e descontamine as luvas, bem como o espaço de trabalho com etanol 80%. Por razões semelhantes, não use os suportes de braçadeira impressos em 3D mais de uma vez. Antes do processo de incorporação propriamente dito, é importante manter todos os componentes do hidrogel (solução de reticulação, solução de colágeno-1) no gelo para evitar ligações cruzadas prematuras. Consequentemente, deve-se trabalhar rapidamente uma vez que as células são adicionadas à solução de reticulação para limitar a morte celular devido ao pH elevado e à baixa temperatura da solução de reticulação. Para evitar a morte celular relacionada à secagem no explante do núcleo, aspirar o meio que cobre os explantes do núcleo pinçado imediatamente antes de misturar a solução de reticulação com a solução de colágeno-1. Para garantir a colocação central do explante do núcleo dentro do hidrogel, o ideal é lançar o hidrogel em torno de um explante de núcleo fixo que seja levemente tensionado. Para isso, use o pino de cavilha e o parafuso M3 x 16 mm para fixar os suportes de fixação a uma placa (impressa em 3D) com furos nos comprimentos apropriados. Após o tempo de polimerização de 50 min, o explante do núcleo embutido pode ser destensionado novamente, dependendo das condições de cultura desejadas. A quantidade de tensão que o assemblóide experimenta durante o cultivo tem um profundo impacto nos resultados experimentais e deve ser mantida uniforme entre amostras e condições21.
No entanto, o grande impacto do (des)carregamento mecânico nos resultados experimentais é uma das principais vantagens do modelo assemblóide sobre a maioria das alternativas de engenharia tecidual, especialmente porque a composição da matriz mantida do explante central também deve recriar os complexos padrões de carregamento in vivo no nível celular90. Enquanto na prática apenas a medida do módulo de elasticidade linear, a tensão máxima de tração e a tensão máxima de tração dos assemblóides foram demonstradas até o momento, protocolos para medidas de força de fadiga e relaxamento de tensão foram descritos para explantes de tendão central em outros lugares e devem ser aplicáveis aos assemblóides91,92. Além dos padrões de carregamento in vivo, a modularidade multinível do assembloid é provavelmente sua maior vantagem. Graças às câmaras de cultura individuais, um conjunto controlável de condições de nicho pode ser definido para cada amostra separadamente (ou seja, temperatura, tensão de oxigênio, concentração de glicose, suplementação, estimuladores, inibidores e estiramento estático com uma placa). Em seguida, a rigidez da matriz e a composição da matriz do compartimento extrínseco são personalizáveis através da composição do hidrogel e permitiriam, por exemplo, estudar o impacto de um microambiente tecidual cada vez mais doente incorporando mais colágeno-3 e fibronectina celular 93,94,95. As populações celulares avaliadas no compartimento extrínseco são facilmente adaptáveis selecionando quais células semear, mas também podem ser modificadas no explante do núcleo do tendão aproveitando linhagens celulares geneticamente modificadas estabelecidas e linhagens de camundongos (i.e., depleção de células ScxLin)96. A diferente matriz e composição celular dos dois compartimentos fornece ainda uma estrutura 3D compartimentada única, que é outra característica do tendão central 1,30,46.
Ao usar esse sistema, é importante considerar as consequências da modularidade do sistema para a granularidade dos parâmetros de resultado. Enquanto a proliferação e o recrutamento celular podem ser avaliados para cada compartimento separadamente, as propriedades mecânicas, os componentes do secretoma e os produtos de degradação são atualmente mensuráveis apenas para o assemblóide completo. Em relação ao rendimento, uma pessoa devidamente treinada pode preparar até 50 assembloides em um dia de trabalho regular, sendo o principal gargalo o procedimento de fixação. Embora alguns dos métodos de leitura sejam mutuamente exclusivos, é possível avaliar propriedades mecânicas e componentes do secretoma repetidamente na mesma amostra, bem como a composição da população celular (citometria de fluxo), transcriptoma celular (RT-qPCR, sequenciamento de RNA) ou matriz e distribuição celular (imunocitoquímica/microscopia de fluorescência) nos endpoints. Em publicações anteriores, esses métodos foram empregados para caracterizar extensivamente interações intercelulares e compartimentais cruzadas em assemblóides de núcleo // fibroblasto e núcleo // macrófagos expostos a um nicho semelhante a uma lesão84,85. Neste trabalho, a capacidade do sistema modelo assemblóide de sondar a interação compartimental cruzada entre o núcleo e as células endoteliais extrínsecas sob diferentes estímulos microambientais foi explorada.
A modularidade do sistema modelo permite o refinamento futuro do método, o que é necessário para superar as seguintes limitações da iteração de projeto atual. A análise por citometria de fluxo apresentada neste trabalho e os dados de sequenciamento de RNA unicelular publicados recentemente revelaram que os tenócitos residentes no núcleo do tendão e as populações derivadas do tendão de Aquiles são mais heterogêneas do que se supunha anteriormente24,34,59,84,97. Além disso, o comportamento migratório de populações celulares inicialmente residentes no núcleo ou no hidrogel obscurece a compartimentalização do assemblóide durante o cultivo. Ambos os fatores juntos tornam desafiador atribuir diferenças transcriptômicas a tipos celulares específicos e separar processos baseados em proliferação e migração. Essa limitação poderia ser superada refinando a população de entrada com a classificação celular ativada por fluorescência (FACS) com base na composição celular de tendões saudáveis ou doentes caracterizados em estudos in vivo recentes, melhorando a leitura implementando o sequenciamento de RNA de célula única e integrando marcadores de proliferação como uma coloração de EdU (5-etinil-2'-desoxiuridina) durante a microscopia.
Os assemblóides aqui apresentados também compartilham uma fraqueza com a maioria dos sistemas in vitro atualmente disponíveis que simulam órgãos doentes desconectados do resto do corpo98,99. No entanto, a plataforma baseada em câmara de cultura usada aqui posiciona bem o sistema modelo para integração em uma plataforma de múltiplos órgãos, onde assembloides imitando diferentes órgãos são conectados e interações interórgãos podem ser estudadas.
Em sua essência, o sistema modelo é baseado em tendões de roedores posicionais, o que resulta em seu próprio conjunto único de desvantagens. Primeiro, a traduzibilidade dos resultados é prejudicada por camundongos selvagens que não se desenvolvem ou sofrem de doenças tendíneas 8,100,101. A integração de tecidos e células de humanos ou linhagens de camundongos recém-desenvolvidas que exibem aspectos de doença tendínea poderia aliviar esse problema102. A mudança para um assemblóide baseado em humanos é particularmente interessante, pois permitiria estudos com tecidos derivados de pacientes de tendões doentes diferentes (ou seja, tendinite, tendinose ou peritendinite) e até mesmo doadores resistentes ao tratamento que poderiam desbloquear programas de tratamento mais personalizados. Em segundo lugar, os explantes do tendão da cauda murina não lidam particularmente bem com microdanos induzidos por sobrecarga, o que limita a aplicabilidade do sistema modelo para o estudo de danos agudos ao tendão.
Por todas essas razões, os explantes // assemblóides de hidrogel estão em uma posição privilegiada para estudar a biologia do núcleo do tendão, interações estrutura-função da matriz e interações intercompartimentais entre populações celulares específicas em resposta a microdanos induzidos por nicho. Os insights coletados a partir desses estudos de alto rendimento podem dar direção à pesquisa in vivo e ao desenvolvimento de tratamentos.
Disclosures
Os autores não têm nada a revelar.
Acknowledgements
Este trabalho foi financiado pelo ETH Grant 1-005733
Materials
| Name | Company | Catalog Number | Comments |
| 0.4 mm x 25 mm injection needle (G27) | Sterican | 9186174 | |
| 3D printing filament: Clear polylactic acid prusament | Prusa | NA | |
| 4% formaldehyde | Roti-Histofix | P087.4 | |
| Accutase cell detachment solution | Sigma-Aldrich | A6964-100ML | |
| Amphotericin | VWR | L0009-100 | |
| Attachable digital C-mount camera: Moticam 2 | Motic | NA | |
| Bolt screw M3 x 16 mm, stainless steel | RS PRO | 1871235 | |
| Bolt screw M3 x 6 mm, stainless steel | RS PRO | 1871207 | |
| CaCl2 | Sigma-Aldrich | C5670 | |
| CD146 antibody: PE anti-mouse | BioLegend | 134703 | |
| CD206 antibody: Alexa Fluor 488 anti-mouse | BioLegend | 141709 | |
| CD31 antibody: Alexa Fluor 488 anti-mouse | BioLegend | 102413 | |
| CD86 antibody: PE anti-mouse | BioLegend | 105007 | |
| Collagenase I | Thermo Fisher Scientific | 17100017 | |
| Collagenase IV | Gibco | 17104-019 | |
| Dialyzed Fetal Bovine Serum (FBS) | Sigma-Aldrich | F0392-100ML | |
| Dimethyl sulfoxide (DMSO) | Sigma-Aldrich | 7000183 | |
| Dispase II | Sigma-Aldrich | D4693-1G | |
| DMEM/F12 | Sigma | 7002211 | |
| Dowel Pin, 3 mm x 16 mm, stainless steel | Accu | HDP-3-16-A1 | |
| Dragon Skin 10 Slow/1 silicone | KauPO | 09301-004-000001 | |
| Endopan 3 Kit | Pan-Biotech | P04-0010K | |
| Endothelial cell growth supplement | Lonza | CC-3162 | |
| Eppendorf safe-lock plastic tubes (1.5 mL) | Eppendorf | 30121023 | |
| Ethidium homodimer, EthD-1, 2 mM stock in DMSO | Sigma-Aldrich | 46043-1MG-F | |
| F4/80 antibody: Apc/fire 750 anti-mouse | BioLegend | 123151 | |
| Falcon plastic tube (15 mL) | Corning | 352096 | |
| Falcon plastic tube (50 mL) | Corning | 352070 | |
| Flow cytometer: LSR II Fortessa | BD Bioscience | 23-11617-02 | |
| Gelatin | Invitrogen | D12054 | |
| Hellmanex III alkaline cleaning concentrate | Sigma | Z805939-1EA | |
| Heparin | Sigma-Aldrich | H3149-10KU | |
| Hydroxyproline assay | Sigma-Aldrich | MAK008 | |
| Image analysis software: Motic Images Plus 3.0 ML | Motic | NA | |
| L-Ascorbic Acid Phosphate Magnesium Salt n-Hydrate | Wako Chemicals | 013-19641 | |
| LSE Low Speed Orbital Shaker | Corning | 6780-FP | |
| MEM non-essential amino acids | Sigma | 7002231 | |
| Mouse macrophage-stimulating factor (m-CSF) | PeproTech | 315-02-50ug | |
| MSD assay | Mesoscale Discovery | various | |
| NucBlue | Thermo Fisher Scientific | R37605 | |
| Nylon mesh strainer cap, 100 µm | Corning | 734-2761 | |
| Original Prusa i3 MK3S 3D printer | Prusa | i3 MK3S | |
| Penicillin-Streptomycin | Sigma-Aldrich | P4333 | |
| Phosphate-buffered saline (PBS), ph 7.4, sterile, 10 L | Gibco | 10010001 | |
| Puromycin | Gibco | A1113803 | |
| RBC lysis buffer | VWR | 786-650 | |
| recombinant m-CSF | PeproTech | 315-02 | |
| RNA extraction kit: Rneasy plus Micro | Qiagen | 74034 | |
| Slicing software: PrusaSlicer | Prusa | NA | Version 2.6.0 or higher |
| Sterile Cell Strainer 100 µm | Fisherbrand | 22363549 | |
| Surgical scalpel blade No. 21 | Swann-Morton | 307 | |
| Trizol reagent | Thermo Fisher Scientific | 15596018 | |
| Trypsin-EDTA (0.5 %) | Gibco | 15400054 |
References
- Snedeker, J. G., Foolen, J. Tendon injury and repair - A perspective on the basic mechanisms of tendon disease and future clinical therapy. Acta Biomaterialia. 63, 18-36 (2017).
- Wang, J. H. C. Mechanobiology of tendon. Journal of Biomechanics. 39 (9), 1563-1582 (2006).
- Kirkendall, D. T., Garrett, W. E. Function and biomechanics of tendons. Scandinavian Journal of Medicine and Science in Sports. 7 (2), 62-66 (1997).
- Götmark, F., Cafaro, P., O'Sullivan, J. Aging human populations: Good for us, good for the earth. Trends in Ecology and Evolution. 33 (11), 851-862 (2018).
- Maffulli, N., Wong, J., Almekinders, L. C. Types and epidemiology of tendinopathy. Clinics in Sports Medicine. 22 (4), 675-692 (2003).
- Renström, P. A. F. H., Woo, S. L. -. Y. Tendinopathy: A major medical problem in sport. Tendinopathy in Athletes. , (2007).
- Screen, H. R. C., Birk, D. E., Kadler, K. E., Ramirez, F., Young, M. Tendon functional extracellular matrix. Journal of Orthopaedic Research. 33 (6), 793-799 (2016).
- Wunderli, S. L., Blache, U., Snedeker, J. G., Wunderli, S. L., Blache, U., Tendon, J. G. S. Tendon explant models for physiologically relevant in vitro study of tissue biology - a perspective. Connective Tissue Research. 61 (3-4), 262-277 (2020).
- Magnusson, S. P., Langberg, H., Kjaer, M. The pathogenesis of tendinopathy: balancing the response to loading. Nature Reviews Rheumatology. 6 (5), 262-268 (2010).
- Heinemeier, K. M., Schjerling, P., Øhlenschlæger, T. F., Eismark, C., Olsen, J., Kjær, M. Carbon-14 bomb pulse dating shows that tendinopathy is preceded by years of abnormally high collagen turnover. FASEB Journal. 32 (9), 4763-4775 (2018).
- Andersson, G., Backman, L. J., Scott, A., Lorentzon, R., Forsgren, S., Danielson, P. Substance P accelerates hypercellularity and angiogenesis in tendon tissue and enhances paratendinitis in response to Achilles tendon overuse in a tendinopathy model. British Journal of Sports Medicine. 45 (13), 1017-1022 (2011).
- Rolf, C. G., Fu, B. S. C., Pau, A., Wang, W., Chan, B. Increased cell proliferation and associated expression of PDGFRβ causing hypercellularity in patellar tendinosis. Rheumatology. 40 (3), 256-261 (2001).
- Riley, G. The pathogenesis of tendinopathy. A molecular perspective. Rheumatology. 43 (2), 131-142 (2004).
- Jarvinen, M., Jozsa, L., Kannus, P., Järvinen, T. L., Kvist, M., Leadbetter, W. Histopathological findings in chronic tendon disorders. Scandinavian Journal of Medicine & Science in Sports. 7 (2), 86-95 (1997).
- Soslowsky, L. J., et al. Overuse activity injures the supraspinatus tendon in an animal model: A histologic and biomechanical study. Journal of Shoulder and Elbow Surgery. 9 (2), 79-84 (2000).
- Tran, P. H. T., et al. Early development of tendinopathy in humans: Sequence of pathological changes in structure and tissue turnover signaling. FASEB Journal. 34 (1), 776-788 (2020).
- Theodossiou, S. K., Schiele, N. R. Models of tendon development and injury. BMC Biomedical Engineering. 1 (1), 1-24 (2019).
- Stauber, T., Blache, U., Snedeker, J. G. Tendon tissue microdamage and the limits of intrinsic repair. Matrix Biology. 85-86, 68-79 (2020).
- Wang, T., et al. In vitro loading models for tendon mechanobiology. Journal of Orthopaedic Research. 36 (2), 566-575 (2018).
- Fang, F., Sawhney, A. S., Lake, S. P. Different regions of bovine deep digital flexor tendon exhibit distinct elastic, but not viscous, mechanical properties under both compression and shear loading. Journal of Biomechanics. 47 (12), 2869-2877 (2014).
- Wunderli, S. L., et al. Tendon response to matrix unloading is determined by the patho-physiological niche. Matrix Biology. 89, 11-26 (2020).
- Wunderli, S. L., et al. Minimal mechanical load and tissue culture conditions preserve native cell phenotype and morphology in tendon - A novel ex vivo mouse explant model. Journal of Orthopaedic Research. 36 (5), 1383-1390 (2017).
- Arnoczky, S. P., Lavagnino, M., Egerbacher, M., Caballero, O., Gardner, K. Matrix metalloproteinase inhibitors prevent a decrease in the mechanical properties of stress-deprived tendons. The American Journal of Sports Medicine. 35 (5), 763-769 (2007).
- de Micheli, A. J., et al. Single-cell transcriptomic analysis identifies extensive heterogeneity in the cellular composition of mouse Achilles tendons. American Journal of Physiology - Cell Physiology. 319 (5), C885-C894 (2020).
- Arvind, V., Huang, A. H. Reparative and maladaptive inflammation in tendon healing. Frontiers in Bioengineering and Biotechnology. 9 (July), 1-16 (2021).
- Marsolais, D., Côté, C. H., Frenette, J. Neutrophils and macrophages accumulate sequentially following Achilles tendon injury. Journal of Orthopaedic Research. 19 (6), 1203-1209 (2001).
- Garcia-Melchor, E., et al. Novel self-amplificatory loop between T cells and tenocytes as a driver of chronicity in tendon disease. Annals of the Rheumatic Diseases. 80 (8), 1075-1085 (2021).
- Stolk, M., Klatte-Schulz, F., Schmock, A., Minkwitz, S., Wildemann, B., Seifert, M. New insights into tenocyte-immune cell interplay in an in vitro model of inflammation. Scientific Reports. 7 (1), 9801 (2017).
- Tempfer, H., Traweger, A. Tendon vasculature in health and disease. Frontiers in Physiology. 6, 330 (2015).
- Mienaltowski, M. J., Adams, S. M., Birk, D. E. Regional differences in stem cell/progenitor cell populations from the mouse achilles tendon. Tissue Engineering - Part A. 19 (1-2), 199-210 (2013).
- Sakabe, T., et al. Transcription factor scleraxis vitally contributes to progenitor lineage direction in wound healing of adult tendon in mice. Journal of Biological Chemistry. 293, (2018).
- Dyment, N. A., Hagiwara, Y., Matthews, B. G., Li, Y., Kalajzic, I., Rowe, D. W. Lineage tracing of resident tendon progenitor cells during growth and natural healing. PLoS One. 9 (4), e96113 (2014).
- Harvey, T., Flamenco, S., Fan, C. M. A Tppp3 + Pdgfra + tendon stem cell population contributes to regeneration and reveals a shared role for PDGF signalling in regeneration and fibrosis. Nature Cell Biology. 21 (12), 1490-1503 (2019).
- Zhang, J., et al. Characterization of the structure, vascularity, and stem/progenitor cell populations in porcine Achilles tendon (PAT). Cell and Tissue Research. 384 (2), 367-387 (2021).
- Tarafder, S., et al. Tendon stem/progenitor cells regulate inflammation in tendon healing via JNK and STAT3 signaling. FASEB Journal. 31 (9), 3991-3998 (2017).
- Lee, C. H., et al. Harnessing endogenous stem/progenitor cells for tendon regeneration Find the latest version Harnessing endogenous stem/progenitor cells for tendon regeneration. J Clin Invest. 125 (7), 2690-2701 (2015).
- Lui, P. P. Y., Chan, L. S., Cheuk, Y. C., Lee, Y. W., Chan, K. M. Expression of bone morphogenetic protein-2 in the chondrogenic and ossifying sites of calcific tendinopathy and traumatic tendon injury rat models. Journal of Orthopaedic Surgery and Research. 4, 27 (2009).
- Takeuchi, E., et al. Localization and expression of osteopontin in the rotator cuff tendons in patients with calcifying tendinitis. Virchows Archiv. 438 (6), 612-617 (2001).
- Kadler, K. E., Hill, A., Canty-Laird, E. G. Collagen fibrillogenesis: fibronectin, integrins, and minor collagens as organizers and nucleators. Current Opinion in Cell Biology. 20 (5), 495-501 (2008).
- Millar, N. L., et al. MicroRNA29a regulates IL-33-mediated tissue remodelling in tendon disease. Nature Communications. 6, 6774 (2015).
- Riley, G. P., Harrall, R. L., Constant, C. R., Chard, M. D., Cawston, T. E., Hazleman, B. L. Tendon degeneration and chronic shoulder pain: changes in the collagen composition of the human rotator cuff tendons in rotator cuff tendinitis. Annals of the Rheumatic Diseases. 53 (6), 359-366 (1994).
- Thorpe, C. T., Peffers, M. J., Simpson, D., Halliwell, E., Screen, H. R. C., Clegg, P. D. Anatomical heterogeneity of tendon: Fascicular and interfascicular tendon compartments have distinct proteomic composition. Scientific Reports. 6, 20455 (2016).
- Thorpe, C. T., et al. Distribution of proteins within different compartments of tendon varies according to tendon type. Journal of Anatomy. 229 (3), 450-458 (2016).
- Choi, H., et al. Heterogeneity of proteome dynamics between connective tissue phases of adult Tendon. eLife. 9, e55262 (2020).
- Spiesz, E. M., et al. Tendon extracellular matrix damage, degradation and inflammation in response to in vitro overload exercise. Journal of Orthopaedic Research. 33 (6), 889-897 (2015).
- Dyment, N. A., et al. The paratenon contributes to scleraxis-expressing cells during patellar tendon healing. PLoS One. 8 (3), e59944 (2013).
- Cadby, J. A., Buehler, E., Godbout, C., Van Weeren, P. R., Snedeker, J. G. Differences between the cell populations from the peritenon and the tendon core with regard to their potential implication in tendon repair. PLoS One. 9 (3), e92474 (2014).
- Huh, D., Hamilton, G. A., Ingber, D. E. From three-dimensional cell culture to organs-on-chips. Trends in Cell Biology. 21 (12), 745-754 (2011).
- Duval, K., et al. Modeling physiological events in 2D vs. 3D cell culture. Physiology. 32 (4), 266 (2017).
- Wang, J. H. C. Mechanobiology of tendon. Journal of Biomechanics. 39 (9), 1563-1582 (2006).
- Derwin, K. A., Soslowsky, L. J. A quantitative investigation of structure-function relationships in a tendon fascicle model. Journal of Biomechanical Engineering. 121 (6), 598-604 (1999).
- Herod, T. W., Veres, S. P. Development of overuse tendinopathy: A new descriptive model for the initiation of tendon damage during cyclic loading. Journal of Orthopaedic Research. 36 (1), 467-476 (2017).
- Andersen, M. B., Pingel, J., Kjær, M., Langberg, H. Interleukin-6: A growth factor stimulating collagen synthesis in human tendon. Journal of Applied Physiology. 110 (6), 1549-1554 (2011).
- Langberg, H., Rosendal, L., Kjær, M. Training-induced changes in peritendinous type I collagen turnover determined by microdialysis in humans. Journal of Physiology. 534 (1), 297-302 (2001).
- Zitnay, J. L., et al. Molecular level detection and localization of mechanical damage in collagen enabled by collagen hybridizing peptides. Nature Communications. 8, 14913 (2017).
- Hwang, J., et al. Molecular assessment of collagen denaturation in decellularized tissues using a collagen hybridizing peptide. Acta Biomaterialia. 53, 268-278 (2016).
- Lin, A. H., Zitnay, J. L., Li, Y., Yu, S. M., Weiss, J. A. Microplate assay for denatured collagen using collagen hybridizing peptides. Journal of Orthopaedic Research. 37 (2), 431-438 (2019).
- Blomgran, P., Blomgran, R., Ernerudh, J., Aspenberg, P. A possible link between loading, inflammation and healing: Immune cell populations during tendon healing in the rat. Scientific Reports. 6, 29824 (2016).
- Harvey, T., Flamenco, S., Fan, C. -. M. A Tppp3+Pdgfra+ tendon stem cell population contributes to regeneration and reveals a shared role for PDGF signalling in regeneration and fibrosis. Nature Cell Biology. 21 (12), 1490-1503 (2019).
- Pryce, B. A., Brent, A. E., Murchison, N. D., Tabin, C. J., Schweitzer, R. Generation of transgenic tendon reporters, ScxGFP and ScxAP, using regulatory elements of the scleraxis gene. Developmental Dynamics. 236 (6), 1677-1682 (2007).
- Godinho, M. S. C., Thorpe, C. T., Greenwald, S. E., Screen, H. R. C. Elastin is localised to the interfascicular matrix of energy storing tendons and becomes increasingly disorganised with ageing. Scientific Reports. 7 (1), 9713 (2017).
- Smith, M. M., et al. Modulation of aggrecan and ADAMTS expression in ovine tendinopathy induced by altered strain. Arthritis and Rheumatism. 58 (4), 1055-1066 (2008).
- Asundi, K. R., Rempel, D. M. MMP-1, IL-1β, and COX-2 mRNA expression is modulated by static load in rabbit flexor tendons. Annals of Biomedical Engineering. 36 (2), 237-243 (2008).
- Millar, N. L., et al. Inflammation is present in early human tendinopathy. American Journal of Sports Medicine. 38 (10), 2085-2091 (2010).
- Dakin, S. G., et al. Inflammation activation and resolution in human tendon disease. Science Translational Medicine. 7 (311), 311ra173 (2015).
- Jelinsky, S. A., Rodeo, S. A., Li, J., Gulotta, L. V., Archambault, J. M., Seeherman, H. J. Regulation of gene expression in human tendinopathy. BMC Musculoskeletal Disorders. 12, 86 (2011).
- Kannus, P., Józsa, L. Histopathological changes preceding spontaneous rupture of a tendon. A controlled study of 891 patients. The Journal of Bone and Joint Surgery. American. 73 (10), 1507-1525 (1991).
- Comin, J., et al. The prevalence and clinical significance of sonographic tendon abnormalities in asymptomatic ballet dancers: A 24-month longitudinal study. British Journal of Sports Medicine. 47 (2), 89-92 (2013).
- Schubert, T. E. O., Weidler, C., Lerch, K., Hofstädter, F., Straub, R. H. Achilles tendinosis is associated with sprouting of substance P positive nerve fibres. Annals of the Rheumatic Diseases. 64 (7), 1083-1086 (2005).
- Quigley, A. S., Bancelin, S., Deska-Gauthier, D., Légaré, F., Kreplak, L., Veres, S. P. In tendons, differing physiological requirements lead to functionally distinct nanostructures. Scientific Reports. 8 (1), 4409 (2018).
- Herod, T. W., Chambers, N. C., Veres, S. P. Collagen fibrils in functionally distinct tendons have differing structural responses to tendon rupture and fatigue loading. Acta Biomaterialia. 42, 296-307 (2016).
- Shepherd, J. H., Riley, G. P., Screen, H. R. C. Early stage fatigue damage occurs in bovine tendon fascicles in the absence of changes in mechanics at either the gross or micro-structural level. Journal of the Mechanical Behavior of Biomedical Materials. 38, 163-172 (2014).
- Birch, H. L. Tendon matrix composition and turnover in relation to functional requirements. International Journal of Experimental Pathology. 88 (4), 241-248 (2007).
- Kapałczyńska, M., et al. 2D and 3D cell cultures - a comparison of different types of cancer cell cultures. Archives of Medical Science. 14 (4), 910-919 (2018).
- Hussien, A. A., Niederoest, B., Bollhalder, M., Goedecke, N., Snedeker, J. G. The stiffness-sensitive transcriptome of human tendon stromal cells. Advanced Healthcare Materials. 12 (7), 2101216 (2023).
- Heo, S. J., et al. Aberrant chromatin reorganization in cells from diseased fibrous connective tissue in response to altered chemomechanical cues. Nature Biomedical Engineering. 7 (2), 177-191 (2023).
- Liu, C. -. F., Aschbacher-Smith, L., Barthelery, N. J., Dyment, N., Butler, D., Wylie, C. What we should know before using tissue engineering techniques to repair injured tendons: A developmental biology perspective. Tissue Engineering Part B: Reviews. 17 (3), 165-176 (2011).
- Ruiz-Alonso, S., Lafuente-Merchan, M., Ciriza, J., Saenz-del-Burgo, L., Pedraz, J. L. Tendon tissue engineering: Cells, growth factors, scaffolds and production techniques. Journal of Controlled Release. 333, 448-486 (2021).
- Rinoldi, C., et al. Tendon tissue engineering: Effects of mechanical and biochemical stimulation on stem cell alignment on cell-laden hydrogel yarns. Advanced Healthcare Materials. 8 (7), e1801218 (2019).
- Walia, B., Huang, A. H. Tendon stem progenitor cells: Understanding the biology to inform therapeutic strategies for tendon repair. Journal of Orthopaedic Research. 37 (6), 1270-1280 (2018).
- Thorpe, C. T., Riley, G. P., Birch, H. L., Clegg, P. D., Screen, H. R. C. Fascicles from energy-storing tendons show an age-specific response to cyclic fatigue loading. Journal of The Royal Society Interface. 11 (92), 20131058-20131058 (2014).
- Thorpe, C. T., Riley, G. P., Birch, H. L., Clegg, P. D., Screen, H. R. C. Fascicles and the interfascicular matrix show decreased fatigue life with ageing in energy storing tendons. Acta Biomaterialia. 56, 58-64 (2017).
- Youngstrom, D. W., Rajpar, I., Kaplan, D. L., Barrett, J. G. A bioreactor system for in vitro tendon differentiation and tendon tissue engineering. Journal of Orthopaedic Research. 33 (6), 911-918 (2015).
- Stauber, T., et al. Extrinsic macrophages protect while tendon progenitors degrade: Insights from a tissue engineered model of tendon compartmental crosstalk. Advanced Healthcare Materials. 10 (20), e2100741 (2021).
- Stauber, T., Moschini, G., Hussien, A. A., Jaeger, P. K., De Bock, K., Snedeker, J. G. IL-6 signaling exacerbates hallmarks of chronic tendon disease by stimulating progenitor proliferation & migration to damage. eLife. 12, RP87092 (2023).
- Doillon, C. J., Mantovani, D., Rajan, N., Habermehl, J., Cote, M. Preparation of ready-to-use storable and reconstituted type I collagen from rat tail tendon for tissue engineering applications. Nature Protocols. 1 (6), 2753-2758 (2007).
- Blache, U., et al. Inhibition of ERK 1/2 kinases prevents tendon matrix breakdown. Scientific Reports. 11 (1), 6838 (2021).
- Liu, X., et al. The role of vascular endothelial growth factor in tendon healing. Frontiers in Physiology. 12, 766080 (2021).
- Claxton, S., Kostourou, V., Jadeja, S., Chambon, P., Hodivala-dilke, K., Fruttiger, M. Efficient, inducible Cre-recombinase activation in vascular endothelium. Genesis. 46 (2), 74-80 (2008).
- Passini, F. S., et al. Shear-stress sensing by PIEZO1 regulates tendon stiffness in rodents and influences jumping performance in humans. Nature Biomedical Engineering. 5 (12), 1457-1471 (2021).
- Lee, A. H., Szczesny, S. E., Santare, M. H., Elliott, D. M. Investigating mechanisms of tendon damage by measuring multi-scale recovery following tensile loading. Acta Biomaterialia. 57, 363-372 (2017).
- Lee, A. H., Elliott, D. M. Multi-scale loading and damage mechanisms of plantaris and rat tail tendons. Journal of Orthopaedic Research. 37 (8), 1827-1837 (2019).
- Williams, I. F., Heaton, A., McCullagh, K. G. Cell morphology and collagen types in equine tendon scar. Research in Veterinary Science. 28 (3), 302-310 (1980).
- Maffulli, N., Ewen, S. W. B., Waterston, S. W., Reaper, J., Barrass, V. Tenocytes from ruptured and tendinopathic achilles tendons produce greater quantities of type III collagen than tenocytes from normal achilles tendons: An in vitro model of human tendon healing. American Journal of Sports Medicine. 28 (4), 499-505 (2000).
- Pajala, A., Melkko, J., Leppilahti, J., Ohtonen, P., Soini, Y., Risteli, J. Tenascin-C and type I and III collagen expression in total Achilles tendon rupture. An immunohistochemical study. Histology and Histopathology. 24 (10), 1207-1211 (2009).
- Best, K. T., et al. Scleraxis-lineage cell depletion improves tendon healing and disrupts adult tendon homeostasis. eLife. 10, e62203 (2021).
- Lehner, C., et al. Tenophages: A novel macrophage-like tendon cell population expressing CX3CL1 and CX3CR1. DMM Disease Models and Mechanisms. 12 (12), dmm041384 (2019).
- Ajalik, R. E., et al. Human organ-on-a-chip microphysiological systems to model musculoskeletal pathologies and accelerate therapeutic discovery. Frontiers in Bioengineering and Biotechnology. 10, 846230 (2022).
- Ingber, D. E. Human organs-on-chips for disease modelling, drug development and personalized medicine. Nature Reviews Genetics. 23 (8), 467-491 (2022).
- Brehm, M. A., Shultz, L. D., Luban, J., Greiner, D. L. Overcoming current limitations in humanized mouse research. The Journal of Infectious Diseases. 208 Suppl (Suppl 2), 125-130 (2013).
- Nunan, R., Harding, K. G., Martin, P. Clinical challenges of chronic wounds: searching for an optimal animal model to recapitulate their complexity. Disease Models & Mechanisms. 7 (11), 1205-1213 (2014).
- Costa-Almeida, R., Calejo, I., Reis, R. L., Gomes, M. E. Crosstalk between adipose stem cells and tendon cells reveals a temporal regulation of tenogenesis by matrix deposition and remodeling. Journal of Cellular Physiology. 233 (7), 5383-5395 (2018).
Reprints and Permissions
Request permission to reuse the text or figures of this JoVE article
Request PermissionExplore More Articles
This article has been published
Video Coming Soon
ABOUT JoVE
Copyright © 2025 MyJoVE Corporation. All rights reserved