
Method Article
استخراج الأنابيب الدقيقة المعدلة من خلايا الثدييات لدراسة معقدات بروتين الأنابيب الدقيقة بواسطة المجهر الإلكتروني بالتبريد
In This Article
Summary
نصف هنا بروتوكولا لاستخراج التوبولين الداخلي من خلايا الثدييات، التي يمكن أن تفتقر إلى إنزيمات محددة معدلة للأنابيب الدقيقة أو تحتوي عليها، للحصول على أنابيب دقيقة مخصبة لإجراء تعديل معين. ثم نصف كيف يمكن تزيين الأنابيب الدقيقة المستخرجة ببروتينات مرتبطة بالأنيبيبات الدقيقة المنقاة لإعداد شبكات للمجهر الإلكتروني بالتبريد.
Abstract
تعد الأنابيب الدقيقة جزءا مهما من الهيكل الخلوي وتشارك في التنظيم داخل الخلايا وانقسام الخلايا والهجرة. اعتمادا على تعديلات ما بعد الترجمة ، يمكن أن تشكل الأنابيب الدقيقة مجمعات مع بروتينات متفاعلة مختلفة. غالبا ما تكون مجمعات البروتين الدقيقة هذه متورطة في الأمراض البشرية. إن فهم بنية هذه المجمعات مفيد لتوضيح آليات عملها ويمكن دراستها بواسطة المجهر الإلكتروني بالتبريد (cryo-EM). للحصول على مثل هذه المجمعات للدراسات الهيكلية ، من المهم استخراج الأنابيب الدقيقة التي تحتوي أو تفتقر إلى تعديلات محددة بعد الترجمة. هنا ، نصف بروتوكولا مبسطا لاستخراج التوبولين الداخلي من خلايا الثدييات المعدلة وراثيا ، بما في ذلك بلمرة الأنابيب الدقيقة ، تليها الترسيب باستخدام الطرد المركزي الفائق. يمكن بعد ذلك استخدام التوبولين المستخرج لإعداد شبكات مجهر إلكتروني بالتبريد مع أنابيب دقيقة مرتبطة ببروتين مرتبط بالأنابيب الدقيقة المنقى ذي الأهمية. على سبيل المثال ، نوضح استخلاص الأنابيب الدقيقة التيروزينية بالكامل من خطوط الخلايا المهندسة لتفتقر إلى الإنزيمات الثلاثة المعروفة لإزالة التيوبولين. ثم يتم استخدام هذه الأنابيب الدقيقة لصنع مركب بروتيني مع التيوبولين المرتبط بالأنابيب الدقيقة غير النشطة إنزيميا على شبكات cryo-EM.
Introduction
الأنابيب الدقيقة هي جزء مهم من الهيكل الخلوي. يشاركون في وظائف مختلفة مثل هجرة الخلايا والانقسام ولكنهم يساهمون أيضا في التنظيم داخل الخلايا. للتكيف مع المصائر الوظيفية المختلفة، تتفاعل الأنابيب الدقيقة مع مجموعة متنوعة من البروتينات المرتبطة بالأنيبيبات الدقيقة (MAPs)، والإنزيمات، والبروتينات الأخرى، والتي سنشير إليها مجتمعة باسم «البروتينات المتفاعلة مع الأنابيب الدقيقة». يمكن توجيه ارتباط الأنابيب الدقيقة لهذه البروتينات بتعديلات توبولين مختلفة ، يشار إليها عادة باسم "رمز التوبولين"1. ومن الأمثلة على هذا التفضيل الكينيسين المرتبط بالسنترومير الانقسامي (MCAK)2 ومجال CAP-Gly من الدينين والدينين CAP-Gly من p1503 ، والذي يفضل أن يرتبط بالتوبولين التيروزين ، في حين أن محركات كينيسين البروتين المرتبط بالسنترومير E (CENP-E)4 و kinesin-25 تفضل التوبولين الذي يفتقر إلى التيروزين C-terminal.
بينما يمكن استخدام مجموعة متنوعة من الطرق لدراسة تفاعلات البروتين في الأنابيب الدقيقة ، غالبا ما يستخدم المجهر الإلكتروني بالتبريد (cryo-EM) لدراسة هذه التفاعلات بدقة قريبةمن الذرية 6,7. في السنوات الأخيرة ، كشفت هياكل cryo-EM عن كيفية البروتينات الحركية الكبيرة مثل dynein8،9،10 و kinesin 11 ، + بروتينات TIP مثل EB312,13 و MCAK14 ، والبروتينات الأخرى مثل Tau 15,16 ، وحتى الجزيئات الصغيرة مثل باكليتاكسيل ، بيلوروسيد ، وزامبانوليد 17 التفاعل مع الأنابيب الدقيقة. لدراسة تفاعلات الأنابيب الدقيقة والبروتين ، عادة ما يتم استخراج الأنابيب الدقيقة من دماغ الخنزير18. بعد ذلك ، يتم إجراء معظم الدراسات في المختبر ، بما في ذلك هياكل الأنابيب الدقيقة cryo-EM ، باستخدام توبولين دماغ الخنازير. وبالتالي ، فإن نتائج هذه الدراسات تحجب أهمية الطبيعة غير المتجانسة لتعديلات التوبولين19 بين الأنسجة وأنواع الخلايا. هذا يخلق مشكلة معينة عند التحقيق في البروتين الذي يتطلب أو يفضل تعديلا معينا لربط الأنابيب الدقيقة. يمكن توضيح ذلك باستخدام توبولين التيروزين ، الركيزة ل microtubule detyrosinase MATCAP.
Detyrosination هو تعديل توبولين يفتقر فيه إلى التيروزين الأحماض الأمينية C-terminal من α-tubulin ، والذي يرتبط بوظيفة ميتوتيك والقلب والخلايا العصبية20. في حين أن الأنابيب الدقيقة التيروزينية بالكامل هي الركيزة المثالية ل MATCAP ، فإن هذا غائب إلى حد كبير في الأنابيب الدقيقة المتاحة تجاريا من دماغ الخنازير بسبب وظيفة vasohibins 21,22 و MATCAP 23 detyrosinases في هذا النسيج 22،23،24،25،26. على الرغم من أن هيلا توبولين المتاح تجاريا يحتوي في الغالب على أنابيب دقيقة تيروزينية ، إلا أن نزع التيروزين يمكن أن يحدث ، وبالتالي فإن مصدر التوبولين هذا أقل ملاءمة لإنشاء عينة موحدة لتحليل cryo-EM.
لتحفيز ارتباط MATCAP بالأنابيب الدقيقة وإنشاء عينة متجانسة للتحليل الهيكلي ، سعينا للحصول على مصدر للأنابيب الدقيقة التي يتم تيروزينها بالكامل. تحقيقا لهذه الغاية ، تم إنشاء خط خلية MATCAP ونقص vasohibin ، والذي تم استخدامه لاستخراج الأنابيب الدقيقة التيروزينية بالكامل. استند إجراء الاستخراج إلى بروتوكولات راسخة تستخدم دورات متكررة من البلمرة وإزالة بلمرة الأنابيب الدقيقة لاستخراج التوبولين من أنسجة المخ أو الخلايا18،27،28،29،30 ، مع خطوة بلمرة واحدة فقط والطرد المركزي على وسادة الجلسرين. باستخدام MATCAP كمثال ، نوضح بعد ذلك كيف يمكن استخدام هذه الأنابيب الدقيقة لدراسات cryo-EM. لتحضير شبكات cryo-EM ، يتم وصف بروتوكول تطبيق من خطوتين بتركيز ملح منخفض. تصف الطرق الواردة في هذه الورقة استخراج الأنابيب الدقيقة القابلة للتخصيص بكميات ونقاء كافيين لإجراء تحليل cryo-EM وتوفير بروتوكول مفصل حول كيفية استخدام هذه الأنابيب الدقيقة لإنشاء مجمعات الأنابيب الدقيقة للبروتين على شبكات cryo-EM.
Protocol
ملاحظة: انظر جدول المواد للحصول على التفاصيل المتعلقة بجميع المواد والمعدات المستخدمة في هذا البروتوكول.
1. ثقافة الخلية
ملاحظة: يجب أن تتم جميع زراعة الخلايا في غطاء تدفق رقائقي معقم.
- لاتباع هذا البروتوكول ، قم أولا بإذابة قارورة من الخلايا المجمدة في حمام مائي بدرجة حرارة 37 درجة مئوية. هنا ، نستخدم خط خلية HCT116 المعدل وراثيا والذي يفتقر إلى الإنزيمات الثلاثة المعروفة لإزالة التيروزين ، VASH1 و VASH2 و MATCAP ، لإنشاء توبولين التيروزين.
- تحضير لوحة Ø 10 سم تحتوي على 10 مل من وسط زراعة الخلايا المناسب.
ملاحظة: في هذا البروتوكول ، تم استخدام DMEM (وسط النسر المعدل من Dulbecco) المكمل ب 10٪ v / v FCS (مصل عجل الجنين) والمضاد الحيوي للبنسلين / الستربتومايسين (وسط الثقافة) لخط خلايا HCT116 المعدل وراثيا. - عد الخلايا ، والبذور ~ 2.5 × 106 خلايا قابلة للحياة (~ 20٪ التقاء) في لوحة 10 سم المعدة. هز اللوحة برفق لتوزيع الخلايا بالتساوي. احتضان الطبق في حاضنة زراعة الخلايا 37 درجة مئوية بالغاز مع 5٪ (v / v) CO2 حتى تصل الخلايا إلى التقاء 80٪ -90٪.
ملاحظة: عادة ما يستغرق هذا 3 أيام لخلايا HCT116 ، ولكن الوقت قد يعتمد على كثافة البذر المحددة وخط الخلية المستخدم. - تخلص من وسط زراعة الخلايا باستخدام ماصة أو شفاط فراغ واغسل الطبق 2 × 5 مل من PBS.
ملاحظة: احرص على عدم توزيع PBS بقوة كبيرة على الطبقة الأحادية للخلية لأن ذلك قد يفصل الخلايا عن اللوحة. - أضف 1-2 مل من التربسين ، واحتضان الخلايا في الحاضنة لمدة 2-5 دقائق لفصل الخلايا.
- أضف 2 مل من وسط الاستزراع إلى الطبق لإخماد التربسين.
- قسم معلق الخلية إلى ثلاثة إلى خمسة أجزاء متساوية وأعد زرعها لتوسيع الخلايا في وسط الاستزراع حتى يتم الحصول على 6-12 لوحة متقاربة 15 سم.
2. الحصاد
- اغسل الخلايا برفق باستخدام 10 مل من PBS (1x) لإزالة أي وسط لزراعة الخلايا.
- افصل الخلايا عن الألواح عن طريق احتضان الخلايا لمدة 5 دقائق في درجة حرارة الغرفة باستخدام 3 مل من برنامج تلفزيوني بارد متجمد مكمل ب 5 مللي متر EDTA (معقمة / مفلترة) ، وبعد ذلك ، باستخدام مكشطة خلية.
- اجمع الخلايا في أنبوب سعة 50 مل على الثلج ، وقم بتدويرها لأسفل (10 دقائق ، 250 × جم).
- لاحظ حجم الخلايا المحصودة باستخدام المقياس الحجمي على الأنبوب سعة 50 mL.
ملاحظة: يمكن أن يكون الحجم المتوقع في أي مكان بين ~ 0.5 مل و 4 مل. خطوة الإيقاف المؤقت: قم بتجميد حبيبات الخلية في LN2 ، وقم بتخزينها في -20 درجة مئوية حتى الاستخدام. لاحظ أنه لا يمكن تخزين حبيبات الخلية إلا لبضعة أسابيع أو أشهر. إذا تم الاحتفاظ بالحبيبات لفترة أطول ، فقد يؤدي ذلك إلى انخفاض المحصول أو عدم وجود أنابيب دقيقة على الإطلاق.
- لاحظ حجم الخلايا المحصودة باستخدام المقياس الحجمي على الأنبوب سعة 50 mL.
3. استخراج الأنابيب الدقيقة
ملاحظة: احتفظ بكل شيء للخطوات 3.1-3.5 على الجليد ؛ يجب أن يبقى كل شيء من الخطوة 3.6 فصاعدا دافئا (30-37 درجة مئوية).
- قم بإعداد 10 مل من محلول تحلل الثلج البارد يحتوي على أنابيب 100 مللي متر / KOH (درجة الحموضة 6.9) ، 2 مللي متر EGTA / KOH ، 1 مللي متر MgCl2 ، 1 mM PMSF ، وقرص واحد مثبط للبروتياز (صغير).
- أعد تعليق حبيبات الخلية المحصودة في محلول تحلل 1: 1 فولت / فولت (إذا كنت تشك في الحجم الدقيق ، خذ كمية أقل من محلول التحلل بدلا من أكثر).
- Lyse الخلايا عن طريق صوتنة: 15 ثانية على ، 45 ثانية قبالة ، أمبير 30 ، أربع دورات (تحديد الظروف تجريبيا ، وتغيير وفقا لصوتيكاتور).
- بعد صوتنة ، وجمع عينة لتحليل SDS-PAGE: 2 ميكرولتر من lysate + 18 ميكرولتر من الماء + 5 ميكرولتر من 5x SDS عينة العازلة.
ملاحظة: تحقق تحت المجهر الضوئي القياسي إذا كانت الخلايا قد تحللت بالكامل بالفعل.
- بعد صوتنة ، وجمع عينة لتحليل SDS-PAGE: 2 ميكرولتر من lysate + 18 ميكرولتر من الماء + 5 ميكرولتر من 5x SDS عينة العازلة.
- ماصة الخلايا المحللة في أنبوب الطرد المركزي. تدور لمدة 1 ساعة عند 100000 × جم عند 4 درجات مئوية في دوار جهاز طرد مركزي فائق لمسح المحلل.
ملاحظة: تأكد من أن جميع الجيوب الموجودة في الدوار جافة ونظيفة لضمان التوازن الصحيح لجهاز الطرد المركزي. - استخدم حقنة لإخراج المحللة التي تم تطهيرها. احرص على عدم إزعاج الحبيبات وكذلك الطبقة العائمة في الأعلى.
- اجمع عينة من المحللة التي تم تطهيرها: 2 ميكرولتر من المحللة + 18 ميكرولتر من الماء + 5 ميكرولتر من 5x SDS عينة المخزن المؤقت.
- شطف بعناية بيليه, وغرف قليلا من بيليه عن طريق تحريك طرف ماصة P10 من خلال بيليه; أضف 200 ميكرولتر من الماء و 50 ميكرولتر من المخزن المؤقت SDS.
- تكملة طاف من الخطوة السابقة مع 1 mM GTP و 20 μM باكليتاكسيل لبلمرة الأنابيب الدقيقة ؛ لحجم 1 مل ، أضف 10 ميكرولتر من 2 mM paclitaxel و 10 μL من 100 mM GTP.
تنبيه: قد يسبب باكليتاكسيل تهيج الجلد ، وتلف خطير في العين ، وتهيج الجهاز التنفسي ، والعيوب الوراثية ، وتلف الطفل الذي لم يولد بعد ، وتلف الأعضاء. التعرض لفترات طويلة أو متكررة يسبب تلف الأعضاء. لا تتنفس أو ترش أو تنفض الغبار عن المادة بأي شكل من الأشكال. ارتداء قفازات النتريل المطاطية لمنع ملامسة الجلد. - احتضان المادة الطافية المكملة ب GTP / paclitaxel لمدة 30 دقيقة عند 37 درجة مئوية للسماح للأنابيب الدقيقة بالتجمع.
- خلال خطوة الحضانة هذه ، اسمح للدوار وأجهزة الطرد المركزي الفائقة بالتسخين حتى 30 درجة مئوية.
- تحضير العازلة للوسادة: أضف 0.6 مل من الجلسرين إلى 0.4 مل من محلول التحلل ، واستكمل الخليط ب 20 ميكرومتر باكليتاكسيل. سخني المخزن المؤقت للوسادة إلى 37 درجة مئوية.
- أضف 800 ميكرولتر من المخزن المؤقت للوسادة إلى أنبوب الطرد المركزي الفائق. ماصة المحللة المكملة ب GTP / paclitaxel بعناية فوق المخزن المؤقت للوسادة.
ملاحظة: منع خلط المخزن المؤقت للوسادة والمحلول عن طريق السحب برفق شديد. - تدور لمدة 30 دقيقة عند 100000 × جم عند 30 درجة مئوية في دوار الطرد المركزي الفائق. ضع علامة على الحافة المواجهة للخارج لأنبوب الطرد المركزي للتعرف بسهولة على المكان الذي يجب أن تتشكل فيه حبيبات الأنابيب الدقيقة.
- قم بإزالة المخزن المؤقت للوسادة بعناية باستخدام ماصة ، مع الحرص على عدم إزعاج حبيبات الأنابيب الدقيقة.
- اجمع عينة من المخزن المؤقت للوسادة: 2 ميكرولتر + 18 ميكرولتر من الماء + 5 ميكرولتر من 5x SDS عينة عازلة .
- اغسل الحبيبات بعناية 3x باستخدام محلول تحلل دافئ لإزالة الجلسرين. قم بتوزيع المخزن المؤقت الدافئ برفق بجوار الحبيبات (لا تغسل السائل مباشرة فوق الحبيبة) ، وقم بتدوير الأنبوب عدة مرات لإزالة أكبر قدر ممكن من الجلسرين من الحبيبات وجدران الأنبوب ، ثم استنشاق وكرر.
ملاحظة: إذا لم يتم غسل الجلسرين بشكل صحيح ، فسوف تذوب الشبكة بسرعة كبيرة تحت إضاءة الإلكترون. قد يتضح ذلك من خلال حركة الجسيمات العالية ، مما يخلق صورا ضبابية. - قم بإعداد مخزن إعادة التعليق بالمكونات التالية: أنابيب 100 مللي متر / KOH (درجة الحموضة 6.9) ، 2 مللي متر EGTA / KOH ، و 1 مللي متر MgCl2 ، وقم بتسخين المخزن المؤقت إلى 37 درجة مئوية.
- أعد تعليق الحبيبات المغسولة برفق بطرف مقطوع في ~ 50 ميكرولتر من المخزن المؤقت لإعادة التعليق المسخن مسبقا ، واحتفظ بالأنبوب عند 37 درجة مئوية.
- اجمع عينة من جزء الحبيبات المعاد تعليقه: 2 ميكرولتر + 18 ميكرولتر من الماء + 5 ميكرولتر من 5x SDS عينة المخزن المؤقت.
نصيحة: يمنع الطرف المقطوع كسر الأنابيب الدقيقة. قم بإعداد كتلة تسخين معدنية إلى 37 درجة مئوية ، واحتفظ بها في صندوق بوليسترين بحيث يمكن نقل أنابيب العينة بسهولة دون تبريدها إلى درجة حرارة الغرفة.
- اجمع عينة من جزء الحبيبات المعاد تعليقه: 2 ميكرولتر + 18 ميكرولتر من الماء + 5 ميكرولتر من 5x SDS عينة المخزن المؤقت.
4. إعداد شبكة Cryo-EM
- قم بإعداد جهاز الفريزر الغاطس عن طريق تثبيت ورق النشاف. قم بتسخين الفريزر المغطس حتى 30 درجة مئوية ، واضبط الترطيب على 100٪. السماح ~ 30 دقيقة لموازنة درجة الحرارة والرطوبة.
- قم بإعداد إعدادات الفريزر الغاطس لتطبيقين ، وقم بتشغيل البرنامج بأكمله مرة واحدة للتأكد من ضبط الإعدادات بشكل صحيح. تأكد من أن التطبيق الأول له قوة 10 و 2 s وقت لطخة و 0 s وقت انتظار وأن التطبيق الثاني له قوة 10 و 6.5 s وقت لطخة و 10 s وقت انتظار.
- توهج تفريغ شبكات cryo-EM عند 30 مللي أمبير لمدة 60 ثانية.
- قم بتبريد مجموعة حاوية البوليسترين باستخدام LN2 ، وقم بإعداد الإيثان السائل في كوب معدني عن طريق تكثيف غاز الإيثان في كوب معدني بارد.
- قم بإعداد مخزن مؤقت للتخفيف بالمكونات التالية: أنابيب 100 مللي متر / KOH (درجة الحموضة 6.9) ، 2 مللي متر EGTA / KOH ، و 1 مللي متر MgCl2 ، وقم بتسخينه إلى 37 درجة مئوية.
- قم بتخفيف البروتين المتفاعل مع الأنابيب الدقيقة 1: 1 فولت / فولت مع مخزن مخفف قبل تطبيقه مباشرة على الشبكات لضمان تقليل تركيز الملح (استخدمنا تركيز ملح نهائي قدره 50 مللي مول كلوريد الصوديوم). يحفظ المزيج على حرارة 37 درجة مئوية.
- احصل على شبكة مفرغة متوهجة مع ملاقط الغاطس وانقر عليها في الفريزر الغاطس.
- ضع حاوية البوليسترين مع الإيثان السائل في الفريزر المغرق ، وقم بتشغيله خلال البرنامج المعد: قم أولا بتطبيق 3.5 ميكرولتر من محلول الأنابيب الدقيقة على الشبكة ، واترك الفريزر المغرق يلطخ الشبكة ، ثم ضع على الفور 3.5 ميكرولتر من البروتين المخفف حديثا ، وأخيرا ، دع المكبس يمسح ويغرق في تجميد الشبكة في الإيثان السائل.
- انقل الشبكات إلى صندوق تخزين الشبكة ، وقم بتخزينها في ديوار LN2 حتى التصوير.
النتائج
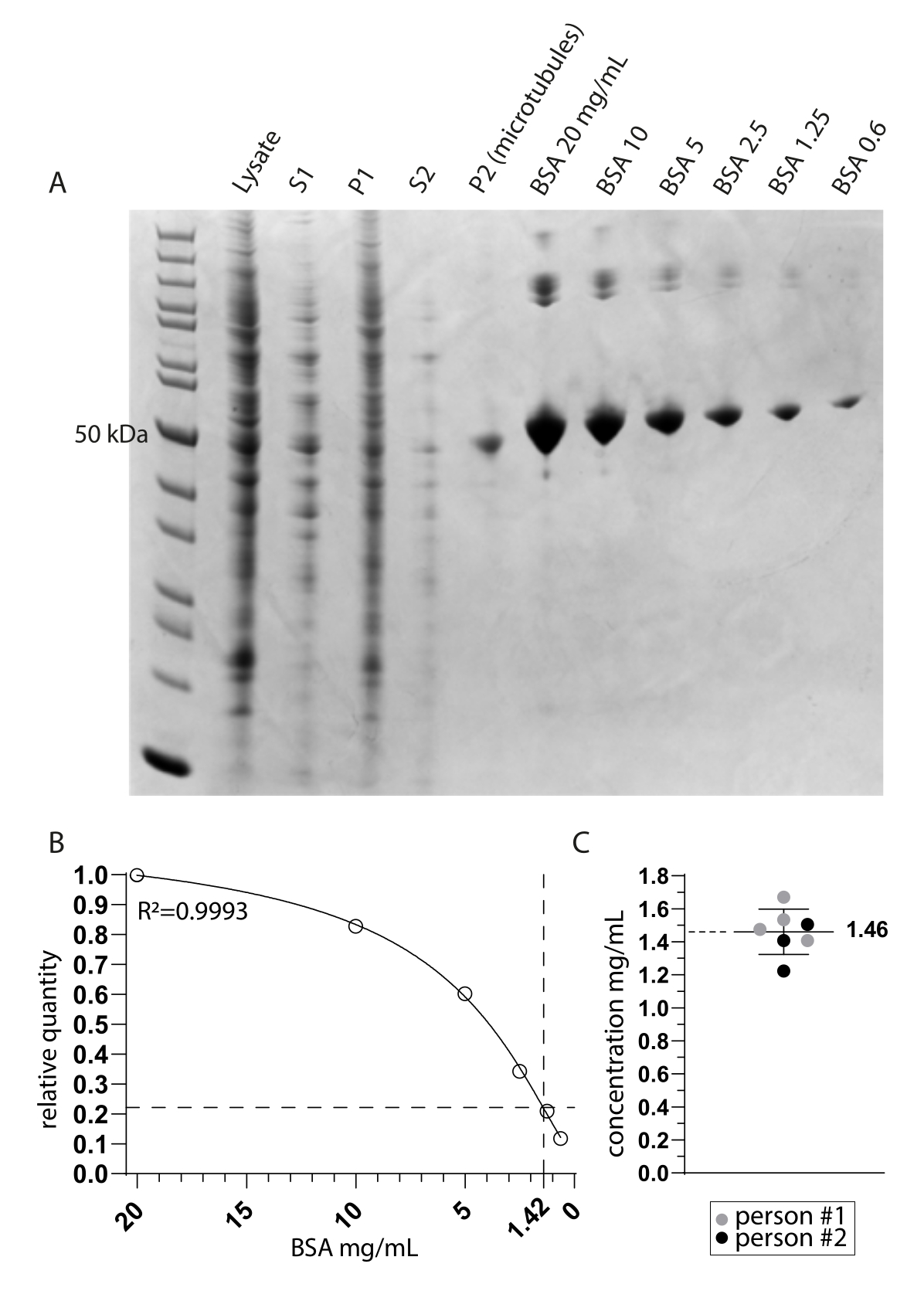
قمنا بالتحقيق في توبولين ديتيروزيناز MATCAP المرتبط بالأنابيب الدقيقة التيروزينية بواسطة cryo-EM. للقيام بذلك ، قمنا باستخراج الأنابيب الدقيقة المطبعية بالكامل من خط خلايا HCT116 المعدل وراثيا والذي يفتقر إلى جميع إنزيمات إزالة التيروزين الثلاثة المعروفة ، VASH1 / 2 و MATCAP. استخدمنا 6-12 طبقا متقاربا بطول 15 سم لاستخراج الأنابيب الدقيقة من حوالي 0.5-4 مل من حبيبات الخلايا (الشكل 1). بعد خطوة الطرد المركزي الثانية (الخطوة 3.11) ، ينتج عن ذلك حبيبات مرئية ولكنها صغيرة وشفافة (الشكل 1I). عادة ما يكون عائد الأنابيب الدقيقة ~ 75 ميكروغرام. إذا كانت الحبيبات غير مرئية ، فقد يشير ذلك إلى وجود مشكلة في إحدى الخطوات السابقة ، مثل درجة حرارة بلمرة الأنابيب الدقيقة غير الصحيحة ، أو مشاكل في جودة GTP أو باكليتاكسيل المستخدم ، أو إضافة الكثير من محلول التحلل ، مما يؤدي إلى تركيز توبولين منخفض جدا للبلمرة. لتقييم جودة وتركيز الأنابيب الدقيقة المستخرجة ، قمنا بتحليل العينات على هلام SDS المصبوغ ب Coomassie (الشكل 2 أ). أشارت هذه التحليلات إلى أن الأنابيب الدقيقة المستخرجة كانت نقية نسبيا. كان تركيز الأنابيب الدقيقة المقحمة المشتقة من القياس الكمي BSA ~ 1.4 مجم / مل. هذا يتفق بشكل جيد مع الرقم المقاس باستخدام مقياس الطيف الضوئي (الشكل 2B ، C).
يمكن استخدام الأنابيب الدقيقة المستخرجة حديثا مباشرة لعمل عينات ل cryo-EM. يجب أن تبدو الأنابيب الدقيقة سليمة وفيرة على الصور المجهرية. لمزيد من تحليل cryo-EM ، من الضروري أن يكون لديك كثافة منخفضة للأنابيب الدقيقة لكل صورة مجهرية لتجنب عبور الأنابيب الدقيقة فوق بعضها البعض (الشكل 3 أ). قد تشير الأنابيب الدقيقة المكسورة أو تلك غير المرئية إلى أن الأنابيب الدقيقة قد أزيلت بلمرة (على سبيل المثال ، بسبب درجة حرارة منخفضة أو التحلل المائي GTP) أو أن معلمات النشاف والتجميد لم يتم ضبطها بشكل صحيح. الخلفية حول الأنابيب الدقيقة كثيفة ، ويفترض أنها تحتوي على توبولين غير مبلمر.
الوزن الجزيئي ل MATCAP هو 53 كيلو دالتون ، وله مجال حفاز كروي أقل بقليل من حجم مونومر توبولين. وبالتالي ، يمكن اكتشاف زخرفة MATCAP على الأنابيب الدقيقة بصريا. أظهرت الأنابيب الدقيقة التي لم تربط MATCAP حواف "ناعمة" ، في حين أن الأنابيب الدقيقة التي تربط MATCAP لها حواف "خشنة" ، تتميز بنقاط كثيفة الإلكترون على سطح الأنابيب الدقيقة (الشكل 3B). يمكن أيضا تمييز الأنابيب الدقيقة المرتبطة ب MATCAP وغير المنضمة إلى MATCAP في فئات 2D المحسوبة ، على الرغم من أنه بسبب الشكل والحجم ، قد يختلف هذا بالنسبة للبروتينات الأخرى المتفاعلة مع الأنابيب الدقيقة (الشكل 3C). لتأكيد أن الكثافة تنتمي بالفعل إلى البروتين محل الاهتمام ، يمكن للمرء الاستفادة من الهياكل المحددة أو المتوقعة تجريبيا. نقترح أيضا إنشاء شبكة تحكم تحتوي على أنابيب دقيقة فقط للمقارنة. يشير هذا إلى ما إذا كانت الأنابيب الدقيقة قد تم بلمرتها واستخلاصها سليمة بتركيز عال بما فيه الكفاية وأن عملية تجميد الغطس قد تم تنفيذها بشكل صحيح. لاحظنا أن وفرة الأنابيب الدقيقة انخفضت في الشبكات مع تطبيق MATCAP الثاني.

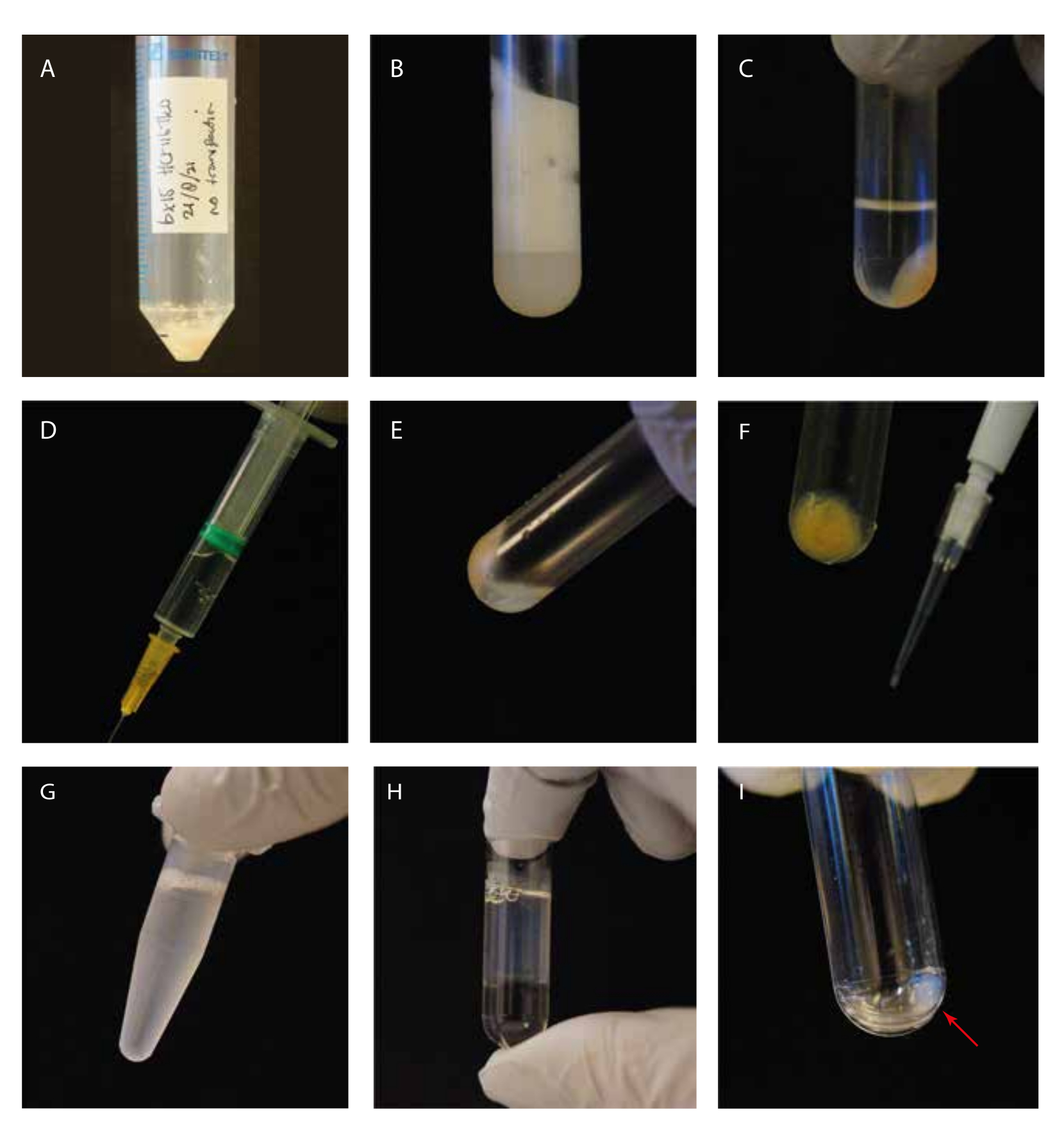
الشكل 1: التوجيه البصري للخطوات التجريبية. (أ) حبيبات الخلية قبل التحلل؛ (ب) الخلايا الصوتية في أنبوب الطرد المركزي الفائق قبل الطرد المركزي ؛ (ج) الخلايا الصوتية في أنبوب الطرد المركزي الفائق بعد الطرد المركزي ؛ (د) حقنة مع المادة الطافية التي تم تطهيرها؛ (ه) الحبيبات المتبقية بعد إزالة المادة الطافية ، بما في ذلك "طبقة عائمة بيضاء" ؛ (و) طرف P10 مع حبيبات حطام خلوية لجل SDS Coomassie ؛ (ز) مادة طاف مكملة ب GTP/باكليتاكسيل قبل الحضانة؛ (ح) طاف محتضن GTP/باكليتاكسيل فوق وسادة من الغليسيرول في أنبوب للطرد المركزي الفائق؛ (I) تنظيف حبيبات الأنابيب الدقيقة بعد خطوة الطرد المركزي الثانية. يرجى النقر هنا لعرض نسخة أكبر من هذا الرقم.
{kind=link}

الشكل 2: نقاء الأنابيب الدقيقة وتحديد التركيز . (أ) جل صفحة SDS مصبوغ بكوماسي يوضح العينات المأخوذة أثناء بروتوكول الاستخراج ومقارنة تركيز BSA. يتوافق S1 و P1 مع المادة الطافية والحبيبات بعد خطوة الطرد المركزي الأولى ، على التوالي. يتوافق S2 و P2 بالمثل مع خطوة الطرد المركزي الثانية. (ب) خط انحدار غير خطي لكميات BSA النسبية المشتقة من A. يشير استيفاء نطاق الأنابيب الدقيقة حوالي 50 كيلو دالتون في ممر P2 (الأنابيب الدقيقة) إلى تركيز نهائي قدره 1.42 مجم / مل. (ج) يشير التحليل الطيفي الضوئي للأنابيب الدقيقة المعاد تعليقها (P2) التي تم قياسها بواسطة شخصين وتصحيحها لمعامل الانقراض المشترك ل TUBA1A و TUBB3 (0.971) إلى متوسط تركيز انحراف معياري قدره 1.46 ملغم / مل ± 0.14 ملغم / مل. يرجى النقر هنا لعرض نسخة أكبر من هذا الرقم.
{kind=link}

الشكل 3: مثال على الصور المجهرية. (أ) أمثلة على الصور المجهرية التي تعرض الأنابيب الدقيقة المرتبطة ب MATCAP. الأنابيب الدقيقة التي يشير إليها الصندوق الأخضر سليمة ومزينة ويمكن استخدامها لتحليل cryo-EM. الأنابيب الدقيقة المشار إليها في المربع البرتقالي هي عبارة عن أنبوب دقيق سليم ومزخرف ولكنه يقع بالقرب من الأنابيب الدقيقة اليسرى منه. لذلك ، من غير المناسب تضمينه في تحليل cryo-EM. الأنابيب الدقيقة في الصندوق الأحمر تعبر وتنكسر. يجب استبعادها من تحليل cryo-EM. (ب) منظر مكبر للأنيبيب الدقيق الأخضر المحاط باللوحة اليسرى. تشير رؤوس الأسهم الحمراء إلى النقاط السوداء التي ظهرت فقط على الأنابيب الدقيقة في الصور المجهرية التي كان لها تطبيق MATCAP ، وبالتالي ، من المحتمل أن تتوافق مع MATCAP المرتبط بالأنابيب الدقيقة. (ج) مثال على فئات 2D لجزيئات الأنابيب الدقيقة التي تم التقاطها من A والتي أظهرت زخرفة عالية ومنخفضة وفئة 2D من مجموعة بيانات مختلفة لم نلاحظ أي زخرفة لها بواسطة MATCAP (اللوحة الموجودة في أقصى اليمين). أشرطة المقياس = (أ) 50 نانومتر ، (ب) 25 نانومتر ، (ج) 10 نانومتر. يرجى النقر هنا لعرض نسخة أكبر من هذا الرقم.
{kind=link}

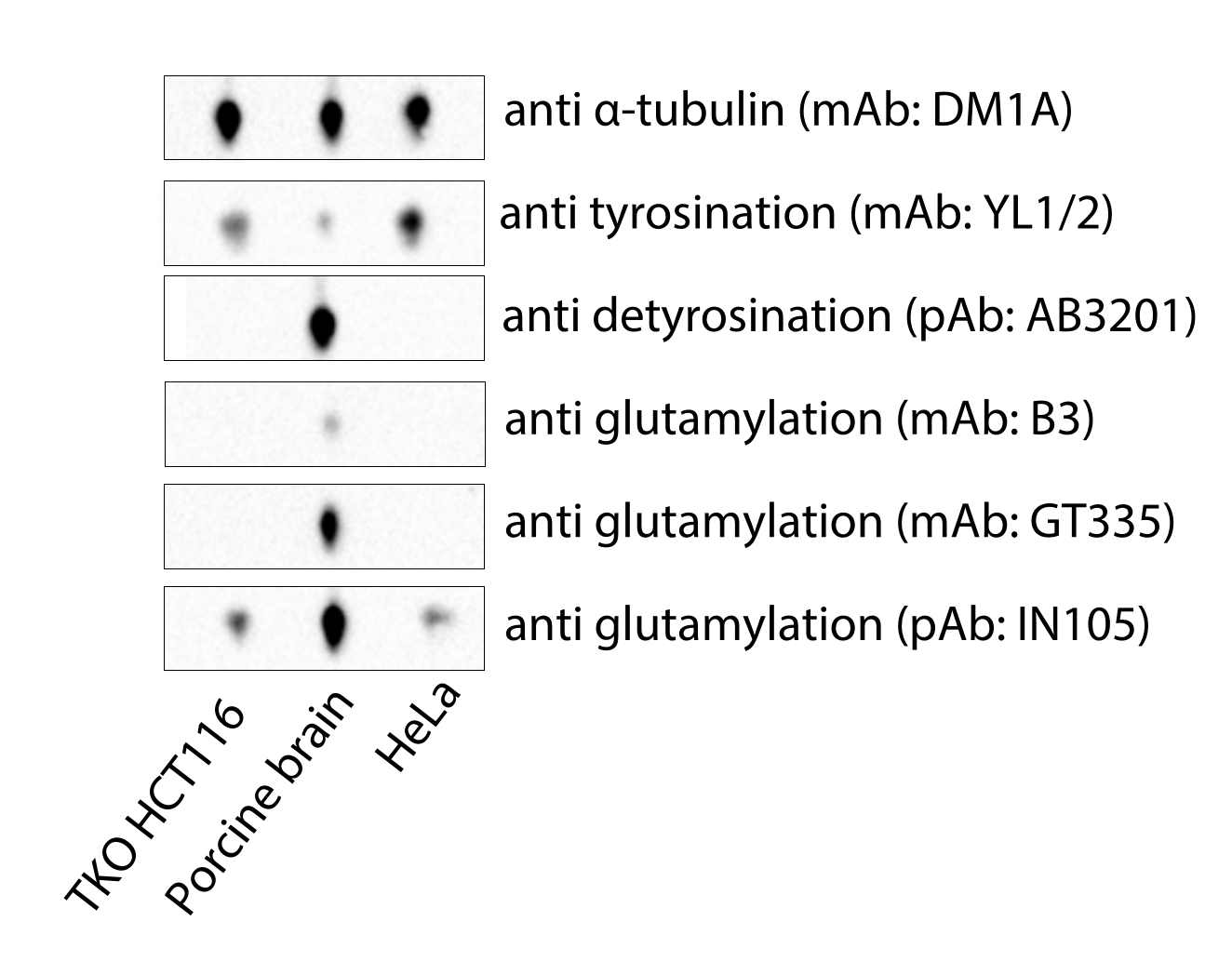
الشكل 4: تحليل اللطخة المناعية للأنابيب الدقيقة المشتقة من خلايا HCT116 التي تعاني من نقص MATCAP ونقص VASH1 / 2 (TKO) ، ودماغ الخنازير التجاري ، وتوبولين HeLa. الاختصارات: TKO = خروج المغلوب الثلاثي. mAb = جسم مضاد وحيد النسيلة ؛ pAb = الأجسام المضادة متعددة النسيلة. يرجى النقر هنا لعرض نسخة أكبر من هذا الرقم.
{kind=link}
Discussion
تصف هذه الطريقة كيفية استخراج التوبولين الداخلي بسرعة من خطوط الخلايا ثم تزيين تلك الأنابيب الدقيقة على شبكات cryo-EM. الأنابيب الدقيقة حساسة لدرجة الحرارة. أنها تبلمر في بيئة باردة والبلمرة في بيئة دافئة31. لذلك ، من الأهمية بمكان تنفيذ دوران الصوتنة والتخليص (الخطوات 1.1-1.5) عند 4 درجات مئوية لإذابة التوبولين. إذا كانت هناك أي عوامل تعمل على استقرار الأنابيب الدقيقة بشكل جيد بحيث لا تتبلمر في هذه الخطوة ، التخلص من هذه الأنابيب الدقيقة وعوامل التثبيت في الحبيبات بعد دوران التخليص الأولي. بعد (إعادة) بلمرة الأنابيب الدقيقة ، من المهم الحفاظ على المحلول الذي يحتوي على الأنابيب الدقيقة المبلمرة دافئا في جميع الأوقات. استخلصنا الأنابيب الدقيقة من خلايا HCT116 ، والتي تعاني من نقص في بروتينات VASH1 و VASH2 و MATCAP. يمكن استخدام خطوط الخلايا الأخرى ، وكذلك الأنسجة ، لاستخراج الأنابيب الدقيقة29 ، على الرغم من أن الملوثات والأنماط المتماثلة للتوبولين والمحصول قد تكون مختلفة تماما عما هو موضح هنا. يمكن أيضا استخدام البلازميدات المفرطة في التعبير التي تحتوي على إنزيمات معدلة لإدخال تعديلات توبولين محددة.
تستخدم البروتوكولات الأخرى18،27،28،29،30 دورات متعددة من البلمرة وإزالة البلمرة من الأنابيب الدقيقة للحصول على أنابيب دقيقة خالية من البروتينات المتفاعلة الأخرى. هنا ، قمنا بتبسيط هذه البروتوكولات وبلمرة الأنابيب الدقيقة مرة واحدة فقط. من الممكن أنه بسبب هذه البلمرة المفردة ، قد تشارك هذه الأنابيب الدقيقة في الرواسب مع البروتينات الأخرى المتفاعلة مع الأنابيب الدقيقة. ومع ذلك ، فقد وجدنا أن هذا البروتوكول يعطي أنابيب دقيقة نقية بما فيه الكفاية لأغراض cryo-EM. إذا كانت هناك حاجة إلى عينة أنقى لفحوصات محددة ، فإن دورات إضافية من البلمرة وإزالة البلمرة يمكن أن تنتج عينة أنقى ، على الرغم من أن هذا قد يكون على حساب محصول الأنابيب الدقيقة. في هذا البروتوكول ، استخدمنا باكليتاكسيل لبلمرة الأنابيب الدقيقة. ومع ذلك ، يمكن أن ينحاز باكليتاكسيل إلى شبكة الأنابيب الدقيقة نحو تطور وارتفاع معين ، مما قد يتداخل مع تقارب الأنابيب الدقيقة للبروتين محل الاهتمام. يمكن استخدام كواشف تثبيت الأنابيب الدقيقة الأخرى إذا كان باكليتاكسيل غير مناسب. ومن الأمثلة على هذه الكواشف جزيئات غير تاكسان مثل peloruside أو متغيرات GTP غير القابلة للتحلل المائي مثل GMPCPP17,32.
للتحقيق الهيكلي للبروتينات التي ترتبط بالأنابيب الدقيقة على شبكات cryo-EM ، يحتاج المرء إلى ربط كمية كافية من البروتين الذي يهم الأنابيب الدقيقة. المشكلة الشائعة هي أن مجمعات البروتين المستقرة في المحلول تنهار على الشبكة. لتشكيل مركب البروتين على الشبكة ، كان من الضروري أولا وضع طبقة من الأنابيب الدقيقة ثم تطبيق البروتين المرتبط بالأنابيب الدقيقة بتركيز ملح منخفض على الشبكة المغلفة بالأنابيب الدقيقة ، وبالتالي تجميع مركب البروتين مباشرة على الشبكة. أبلغ آخرون بالمثل عن بروتوكول33,34 قليل الملح وبروتوكولتطبيق من خطوتين 34,35,36 لتزيين الأنابيب الدقيقة بنجاح. من المحتمل أن يؤدي انخفاض تركيز الملح إلى تحيز مركب البروتين نحو تفاعل أكثر استقرارا بسبب انخفاض الشحنات الكهروستاتيكية. ومع ذلك ، نظرا لانخفاض تركيز الملح ، فإن البروتين محل الاهتمام معرض لخطر الترسيب. لذلك ، يوصى بشدة بالحفاظ على البروتين عند أو حول تركيزات الملح ذات الصلة من الناحية الفسيولوجية حتى قبل وقت قصير من تزجيج الشبكات. من المحتمل أن يمنع بروتوكول التطبيق المكون من خطوتين مركب البروتين من الانهيار أثناء خطوات النشاف أو التجميد. في هذا البروتوكول ، استخدمنا Vitrobot. ومع ذلك ، فإن طرق التزجيج الأسرع (VitroJet) أو استخدام الشبكات الخالية من البقع (Puffalot) أو الأجهزة التي لها كلتا الخاصيتين (الحرباء) يمكن أن تتغلب على التطبيق المكون من خطوتين ، ولكن هذه ليست متاحة حاليا على نطاق واسع للاختبار.
يمكن أن تتأثر الدقة النهائية لكثافة cryo-EM المعاد بناؤها بعدد من العوامل ، بما في ذلك حركة البروتين المرتبط بالأنابيب الدقيقة بالنسبة إلى الأنابيب الدقيقة ومستوى الزخرفة الذي يمكن تحقيقه. من المحتمل أن تكون زخرفة الأنابيب الدقيقة العالية مفيدة للقرار النهائي الذي تم الحصول عليه في إعادة بناء كثافة 3D. يمكن أن يكون هذا محدودا بعدة عوامل ، مثل أعلى تركيز للبروتين يتم الحصول عليه أثناء تنقية البروتين المرتبط بالأنابيب الدقيقة ، وأقل تركيز ملح يمكن أن يتحمله البروتين المتفاعل مع الأنابيب الدقيقة دون تجميع ، وطريقة الربط للبروتين المتفاعل مع الأنابيب الدقيقة (على سبيل المثال ، يمكن أن يمتد البروتين لأكثر من ثنائي توبولين واحد ، مما يعيق نسبة الارتباط 1: 1). على الرغم من أن دقة إعادة بناء cryo-EM قد تتعرض للخطر بسبب الأنابيب الدقيقة ذات الزخرفة القليلة ، إلا أن التحليل الحسابي يمكن أن يتحايل على الكثير من المشكلات ، كما يتضح من بنية معقدة من الأنابيب الدقيقة التي تم الإبلاغ عنها مؤخرا والتي تم تزيينها بشكل ضئيل للغاية8.
يقدم البروتوكول الذي وصفناه هنا طريقة سريعة ومنخفضة التكلفة للحصول على الأنابيب الدقيقة المناسبة لأغراض cryo-EM. على عكس توبولين دماغ الخنازير المتاح تجاريا ، فإن الأنابيب الدقيقة المشتقة من خلايا HCT116 التي تعاني من نقص MATCAP ونقص الفاسوهيبين هي تيروزين بالكامل (الشكل 4). توبولين HeLa التجاري ، وهو كاشف باهظ الثمن ، من حيث المبدأ ، هو تيروزين موحد نسبيا ويحتوي على القليل من التعديلات الأخرى4 مثل الغلوتاميل ، ولكن قد تختلف الدفعات ، ولا يمكن تحقيق التعديل إلا في المختبر. تتمثل إحدى مزايا استخراج الأنابيب الدقيقة من خطوط الخلايا المصممة خصيصا في المرونة التي يتمتع بها المرء للإفراط في التعبير عن الإنزيمات المعدلة للتوبولين أو حذفها ، مثل التيروزيناز التوبولين ، لإنشاء مجموعة أكثر تجانسا من الأنابيب الدقيقة. يمكن أن يفيد ذلك زخرفة وتوحيد عينة cryo-EM وسيفيد في النهاية سهولة وجودة خرائط كثافة cryo-EM والهياكل الجزيئية المشتقة من هذه العينة.
Disclosures
ليس لدى المؤلفين أي تضارب في المصالح للكشف عنه.
Acknowledgements
نشكر جميع أعضاء مجموعات Sixma و Brummelkamp و Perrakis على مناقشاتهم العلمية المثمرة وعلى توفير بيئة عمل ممتعة ، وعلى وجه التحديد ، نشكر Jan Sakoltchik ("الشخص 2") للمساعدة في تحديد تركيز البروتين الموضح في الشكل 3C. نود أيضا أن نشكر مرفق NKI cryo-EM والمركز الهولندي للتنظير الإلكتروني النانوي (NeCEN) في جامعة ليدن على دعمهم. تم دعم هذا العمل من خلال منحة NWO Vici 016.Vici.170.033 الممنوحة ل T.R.B.. A.P. و T.R.B. هما محققان في Oncode ويتلقيان تمويلا من NWO ENW (OCENW. M20.324). تلقت L.L. تمويلا من صندوق العلوم النمساوي (FWF JB4448-B). تم دعم هذا البحث بمنحة مؤسسية من جمعية السرطان الهولندية ووزارة الصحة والرعاية الاجتماعية والرياضة الهولندية.
Materials
| Name | Company | Catalog Number | Comments |
| Material | |||
| 0.05% trypsin-EDTA | Gibco | 25300-054 | Cell culture |
| 10 cm plate | Falcon | 353003 | Cell culture |
| 15 cm plate | Thermo FisherScientific | 168381 | Cell culture |
| 50 mL tubes | Sarstedt | 62.547255 | Cell culture |
| 300 mesh quantifoil holey carbon copper grid R1.2/1.3 | Quantifoil Micro Tools | N1-C14nCu30-01 | Cryo-EM grid preparation |
| Cell scrapers | Falcon | 353085 | Cell culture |
| DMEM | Gibco | 41966-029 | Cell culture |
| EDTA | Merck | 108418 | Cell culture |
| EGTA | Sigma Aldrich | E3899 | Microtubule extraction |
| Ethane gas | Cryo-EM grid preparation | ||
| FCS | Serana | s-FBS-EU-015 | Cell culture |
| Glycerol | VWR | 24.397.296 | Microtubule extraction |
| GTP | Fisher Scientific | G8877-1G | Microtubule extraction |
| HCT116 VASH1 VASH2 MATCAP KO cells | self made | Wild type HCT116 cells RRID: CVCL_0291 | Cell culture |
| KOH | Merck | 1.05033 | Microtubule extraction |
| MgCl2 | Merck | 105833 | Microtubule extraction |
| Microtubule binding protein | self made | Cryo-EM grid preparation | |
| Needle | BD microlance | 300600 | Microtubule extraction |
| Paclitaxel | Santa Cruz Biotechnology | sc-212517 | caution toxic, microtubule extraction |
| PBS | Fisher Scientific | BP399 | Cell culture |
| Penicillin and streptomycin | Sigma Aldrich | P0781-100mL | Cell culture |
| PIPES | Merck | P8203 | Microtubule extraction |
| PMSF (in EtOH) | Roche | 16837091001 | Microtubule extraction |
| SDS sample buffer | self made | Quality assessment | |
| Syringe | BD plastipak | 309658 | Microtubule extraction |
| Ultra protease tables mini | Fisher Scientific | NC0975224 | Microtubule extraction |
| Whatman blotting paper | Whatman | 47000-100 | Cryo-EM grid preparation |
| Equipment | |||
| Flow hood | cell culture | ||
| GloQube | Quorum | Cryo-EM grid preparation | |
| Grid storage box | SWISSCI | 41018 | Cryo-EM grid storage |
| Heating block, electric or metal | to warm the buffers | ||
| Incubator, cell culture | NUAIR | cell culture | |
| LN2 dewar | Cryo-EM grid storage | ||
| Plunge-tweezers | Electron Microscopy Sciences | 0508-L5-PS | Cryo-EM grid preparation, hole drilled in top to fit the vitrobot |
| Polystyrene box | to keep the buffers warm | ||
| Sonicator | Qsonica | Q700 | Microtubule extraction |
| Standard light microscope | Olympus | CKX 41 | Quality assessment |
| TLA 100.3 rotor | Beckman Coulter | Microtubule extraction | |
| TLA 120.2 rotor | Beckman Coulter | Microtubule extraction | |
| Tubes for TLA 100.3 rotor | Beckman Coulter | 326819 | Microtubule extraction |
| Tubes for TLA 120.2 rotor | Beckman Coulter | 347356 | Microtubule extraction |
| Ultracentrifuge | Beckman Coulter | Optima MAX-XP | Microtubule extraction |
| Vitrobot | FEI, ThermoFischer Scientific | mark IV | Cryo-EM grid preparation |
| Vitrobot polystyrene container assembly with metal ethane cup | ThermoFisher Scientific | 200703 | Cryo-EM grid preparation |
| Water bath | cell culture |
References
- Janke, C., Magiera, M. M. The tubulin code and its role in controlling microtubule properties and functions. Nature Reviews Molecular Cell Biology. 21 (6), 307-326 (2020).
- Peris, L., et al. Motor-dependent microtubule disassembly driven by tubulin tyrosination. Journal of Cell Biology. 185 (7), 1159-1166 (2009).
- McKenney, R. J., Huynh, W., Tanenbaum, M. E., Bhabha, G., Vale, R. D. Activation of cytoplasmic dynein motility by dynactin-cargo adapter complexes. Science. 345 (6194), 337-341 (2014).
- Barisic, M., et al. Microtubule detyrosination guides chromosomes during mitosis. Science. 348 (6236), 799-803 (2015).
- Sirajuddin, M., Rice, L. M., Vale, R. D. Regulation of microtubule motors by tubulin isotypes and post-translational modifications. Nature Cell Biology. 16 (4), 335-344 (2014).
- Nogales, E., Kellogg, E. H. Challenges and opportunities in the high-resolution cryo-EM visualization of microtubules and their binding partners. Current Opinion in Structural Biology. 46, 65-70 (2017).
- Manka, S. W., Moores, C. A. Microtubule structure by cryo-EM: Snapshots of dynamic instability. Essays in Biochemistry. 62 (6), 737-751 (2018).
- Chaaban, S., Carter, A. P. Structure of dynein-dynactin on microtubules shows tandem adaptor binding. Nature. 610 (7930), 212-216 (2022).
- Lacey, S. E., He, S., Scheres, S. H., Carter, A. P. Cryo-EM of dynein microtubule-binding domains shows how an axonemal dynein distorts the microtubule. eLife. 8, 47145 (2019).
- Walton, T., Wu, H., Brown, A. Structure of a microtubule-bound axonemal dynein. Nature Communications. 12, 477 (2021).
- Sindelar, C. V., Downing, K. H. An atomic-level mechanism for activation of the kinesin molecular motors. Proceedings of the National Academy of Sciences of the United States of America. 107 (9), 4111-4116 (2010).
- Zhang, R., Alushin, G. M., Brown, A., Nogales, E. Mechanistic origin of microtubule dynamic instability and its modulation by EB proteins. Cell. 162 (4), 849-859 (2015).
- Maurer, S. P., Fourniol, F. J., Bohner, G., Moores, C. A., Surrey, T. EBs Recognize a nucleotide-dependent structural cap at growing microtubule ends. Cell. 149 (2), 371-382 (2012).
- Benoit, M. P. M. H., Asenjo, A. B., Sosa, H. Cryo-EM reveals the structural basis of microtubule depolymerization by kinesin-13s. Nature Communications. 9, 1662 (2018).
- Kellogg, E. H., et al. Near-atomic model of microtubule-tau interactions. Science. 360 (6394), 1242-1246 (2018).
- Brotzakis, Z. F., et al. A structural ensemble of a Tau-microtubule complex reveals regulatory Tau phosphorylation and acetylation mechanisms. ACS Central Science. 7 (12), 1986-1995 (2021).
- Kellogg, E. H., et al. Insights into the distinct mechanisms of action of taxane and non-taxane microtubule stabilizers from cryo-EM structures. Journal of Molecular Biology. 429 (5), 633-646 (2017).
- Vallee, R. B. Reversible assembly purification of microtubules without assembly-promoting agents and further purification of tubulin, microtubule-associated proteins, and MAP fragments. Methods in Enzymology. 134, 89-104 (1986).
- Wloga, D., Joachimiak, E., Louka, P., Gaertig, J. Posttranslational modifications of tubulin and cilia. Cold Spring Harbor Perspectives in Biology. 9 (6), 028159 (2017).
- Nieuwenhuis, J., Brummelkamp, T. R. The tubulin detyrosination cycle: Function and enzymes. Trends in Cell Biology. 29 (1), 80-92 (2019).
- Nieuwenhuis, J., et al. Vasohibins encode tubulin detyrosinating activity. Science. 358 (6369), 1453-1456 (2017).
- Aillaud, C., et al. Vasohibins/SVBP are tubulin carboxypeptidases (TCPs) that regulate neuron differentiation. Science. 358 (6369), 1448-1453 (2017).
- Landskron, L., et al. Posttranslational modification of microtubules by the MATCAP detyrosinase. Science. 376 (6595), (2022).
- Erck, C., et al. A vital role of tubulin-tyrosine-ligase for neuronal organization. Proceedings of the National Academy of Sciences of the United States of America. 102 (22), 7853-7858 (2005).
- Pagnamenta, A. T., et al. Defective tubulin detyrosination causes structural brain abnormalities with cognitive deficiency in humans and mice. Human Molecular Genetics. 28 (20), 3391-3405 (2019).
- Peris, L., et al. Tubulin tyrosination regulates synaptic function and is disrupted in Alzheimer's disease. Brain. 145 (7), 2486-2506 (2022).
- Souphron, J., et al. Purification of tubulin with controlled post-translational modifications by polymerization-depolymerization cycles. Nature Protocols. 14 (5), 1634-1660 (2019).
- Gell, C., et al. Purification of tubulin from porcine brain. Methods in Molecular Biology. 777, 15-28 (2011).
- Bodakuntla, S., Jijumon, A. S., Janke, C., Magiera, M. M. Purification of tubulin with controlled posttranslational modifications and isotypes from limited sources by polymerization-depolymerization cycles. Journal of Visualized Experiments. (165), e61826 (2020).
- Castoldi, M., Popov, A. V. Purification of brain tubulin through two cycles of polymerization-depolymerization in a high-molarity buffer. Protein Expression and Purification. 32 (1), 83-88 (2003).
- Li, G., Moore, J. K. Microtubule dynamics at low temperature: evidence that tubulin recycling limits assembly. Molecular Biology of the Cell. 31 (11), 1154-1166 (2020).
- Hyman, A. A., Salser, S., Drechsel, D. N., Unwin, N., Mitchison, T. J. Role of GTP hydrolysis in microtubule dynamics: information from a slowly hydrolyzable analogue. GMPCPP. Molecular Biology of the Cell. 3 (10), 1155-1167 (1992).
- Sindelar, C. V., Downing, K. H. The beginning of kinesin's force-generating cycle visualized at 9-Å resolution. Journal of Cell Biology. 177 (3), 377-385 (2007).
- Kellogg, E. H., et al. Near-atomic cryo-EM structure of PRC1 bound to the microtubule. Proceedings of the National Academy of Sciences of the United States of America. 113 (34), 9430-9439 (2016).
- Maurer, S. P., Bieling, P., Cope, J., Hoenger, A., Surrey, T. GTPγS microtubules mimic the growing microtubule end structure recognized by end-binding proteins (EBs). Proceedings of the National Academy of Sciences of the United States of America. 108 (10), 3988-3993 (2011).
- Manka, S. W., Moores, C. A. Pseudo-repeats in doublecortin make distinct mechanistic contributions to microtubule regulation. EMBO Reports. 21 (12), 51534 (2020).
Reprints and Permissions
Request permission to reuse the text or figures of this JoVE article
Request PermissionExplore More Articles
This article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. All rights reserved