
Method Article
使用果蝇S2细胞进行细胞分裂的活成像
摘要
使用荧光标记的蛋白质和延时显微镜可以实时可视化细胞分裂。使用此处介绍的协议,用户可以分析细胞分裂时序动力学、线粒主轴组件以及染色体的组装和分离。RNA干扰(RNAi)介导的基因敲除后这些事件的缺陷可以进行评估和量化。
摘要
德罗索菲拉S2细胞是研究组织培养中的线粒体的重要工具,以快速、高通量的方式提供对这一基本细胞过程的分子洞察。S2 细胞已证明适用于固定细胞和活细胞成像应用。值得注意的是,活细胞成像可以产生有价值的信息,了解基因的丢失或敲除如何影响细胞分裂过程中关键事件的动力学和动力学,包括线粒主轴组装、染色体组装和分离,以及整个细胞周期计时。在这里,我们利用S2细胞以荧光标记mCherry:α-tubulin标记线粒主轴和GFP:CENP-A(在果蝇中称为"CID"基因),以标记中心,以分析关键线粒基因对时间的影响。细胞分裂,从前期(特别是在核包络分解;NEBD)到阿纳相的发病。此成像协议还允许在整个线粒体中可视化主轴微管和染色体动力学。在这里,我们的目标是提供一个简单而全面的协议,使读者能够轻松地调整S2细胞进行活成像实验。从这些实验中获得的结果应该通过定义细胞分裂中涉及的基因在多个同时和动态事件中的作用来扩大我们对基因的理解。在这个细胞培养系统中所做的观察可以验证和进一步调查在体内使用令人印象深刻的方法在苍蝇的基因方法。
引言
细胞分裂是所有多细胞生物的关键过程,无论是在它们的发育和平衡1。果蝇长期被用作细胞分裂研究的模型,各种组织类型和遗传条件的实验为研究这一过程提供了重要的见解。虽然这些见解中有许多来自固定细胞条件,但细胞分裂是一个动态过程,有许多运动部分,使得活细胞的可视化成为评估RNAi或基因敲除对细胞分裂许多部分的影响的组成部分,包括主轴形成,染色体和分离,和细胞因子。
多年来,许多协议已经开发和使用,以可视化体内的果蝇细胞分裂。各种群体已经培养了技术,以图像分裂的幼虫象形盘和幼虫脑2,3,4,5,6,7,8.这些技术虽然对特定组织的成像有用,但产量有限,往往需要生成和维护基因,以产生荧光成分并改变感兴趣的基因的表达。来自果蝇的培养S2细胞提供了一种更高的通量替代品,用于快速测试细胞分裂中各种基因的影响。此外,S2细胞能够转染各种荧光蛋白,可以快速进行修饰,以确定RNAi敲除对细胞分裂许多成分的影响。荧光标记感兴趣的基因也可以观察到细胞分裂,允许动态表征其功能9。
在这里,我们提供了一个详细的协议,使用我们最近描述的方法,对线粒S2细胞进行实时成像。我们的方法利用具有荧光标记的稳稳转染细胞进行微管和集中体,其表达受铜诱导原动(金属洛异氨酸;pMT)的控制。该方法可用于利用相对简单的荧光显微镜和基本成像软件,在线粒体的所有阶段成像主轴和染色体动力学。可以进一步定制,以满足个人研究需求,瞬态转染和RNAi提供了扩大的可能性,以确定候选基因在线粒体中的作用。由于该协议相对简单,它可用于小规模功能丧失屏幕,以识别基因,进一步研究在体内将是有益的,允许更集中的努力和效率与后续遗传操纵苍蝇。
研究方案
1. 制备S2细胞进行RNAi治疗
-
从交流pMT:GFP:GFP和pMT:mCherry:β-tubulin稳稳转染S2细胞(+80%存活;在24-28°C生长),种子细胞在6孔无菌培养板中,密度为1 x 106细胞/mL,在新鲜、加热、10%的胎儿牛血清(FBS)中补充施耐德的昆虫介质(SIM)。
注:通过遵循果蝇性RNAi筛查中心(DRSC)(https://dgrc.bio.indiana.edu/Protocols?tab=cells)的协议,可以生成可稳定转染的S2细胞。尽管本文使用的特定稳定 S2 线使用 GFP 和 mCherry,但这些荧光光谱以及其他光谱中都存在许多替代品。对这些荧光蛋白的详细讨论超出了本协议的范围,但其性质和潜在的优缺点已在其他地方进行了专业审查。- 使用细胞计数器或血细胞仪,确定结汇细胞库的密度。
- 确定在总体积为 4 mL(例如,±4 x 106总细胞)中实现 1 x 106细胞/mL 所需的汇入细胞悬浮液的体积。将此容量的电池库存添加到新鲜 SIM 卡中,使总容量达到 4 mL/孔总体积。通过直接从100μL介质的储存瓶中稀释100μL的细胞(细胞/mL),确定细胞存量(细胞/mL)的密度,并通过血细胞计或自动细胞计数器(如果可用)手动计算1:2稀释。
注:如果使用S2细胞未稳态转染,可以进行具有荧光标记(GFP、mCherry等)的瞬态转染测定,以及DNA标记(CENP-A、Histone H2B等)。此外,从果蝇遗传资源中心获得具有各种线粒主轴和DNA标记的稳稳转染细胞系,可能更适合用户(https://dgrc.bio.indiana.edu/cells/Catalog)的确切实验需求。 - 将新播种的细胞放入24-28°C培养箱中,等待36-48小时。
2. 用dsRNA对感兴趣的基因进行种子细胞的处理
注:下面的示例和结果部分将使用短停止(Shot),一种行为因-微管交联剂作为感兴趣的基因。使用无dsRNA或dsRNA对不相关基因(如β-乳糖酶、lacZ)的模拟治疗作为阴性对照。
- 温暖施耐德的昆虫介质不补充FBS(无血清介质;SFM)在24-28°C培养箱中。
-
将细胞从先前播种的井(从步骤1.1.2)转移到15 mL无菌管中,记录转移的细胞总量。
- 保留少量细胞(±100 μL),用于在细胞计数器或血细胞计中计数。
- 在室温下,在1,000 x g下轻轻沉淀细胞3分钟。
- 当离心细胞时,使用细胞计数器或血细胞计确定每mL的细胞浓度。将这个数字乘以正在离心的总体积(在1.2.2中注明),以获得细胞的总数。
- 从颗粒细胞中吸出上清液。
-
重新悬浮加热SFM中的细胞,以获得3 x 106细胞/mL的浓度。
- 将1 mL的再悬浮细胞转移到6孔板中的新孔中。
- 将10-50μg的dsRNA(在100μL无RNA酶水中稀释)直接添加到细胞中(或感兴趣的靶基因),并旋转板进行混合。在24-28°C培养箱中孵育板1小时,允许直接dsRNA进入细胞。
- 在1小时孵育期间,在24-28°C培养箱中加热10%FBS补充SIM卡。
- 用dsRNA孵育细胞1小时后,在未去除1mL介质的情况下,将2 mL的10%FBS补充SIM直接补充到井中。
- 将细胞放入24-28°C培养箱3-7天。
注:与dsRNA浓度一样,可能需要针对所需的靶基因优化总治疗时间。为了评估RNAi的疗效,使用抗体对靶基因对整个细胞解液进行西体消液。如果初始dsRNA靶序列证明无效,则设计针对靶基因中独特序列的替代方案。
3. 荧光蛋白表达的诱导和细胞制备的成像
- 如果使用pMT质粒(包含可诱导的金属洛他素promotor)生成稳定转染的细胞,则通过用硫酸铜(CuSO4)处理细胞,在最终浓度为500 μM,成像前24-36小时,RNAi治疗后4天。使用构成表达质粒,如pAct不需要铜感应。
-
准备细胞进行成像
- 在 24-28 °C 培养箱中加热 10% FBS 补充 SIM 卡 1 小时。
- 将细胞转移到15 mL无菌管中,记录转移的细胞总量。
- 保留少量细胞(±100 μL),用于在细胞计数器或血细胞计中计数。
- 在室温下,在1,000 x g下轻轻沉淀细胞3分钟。
- 当离心细胞时,使用细胞计数器或血细胞计确定浓度(细胞/mL)。将这个数字乘以正在离心的总体积(在2.2.2中注明),以获得细胞的总数。
- 从颗粒细胞中吸出上清液。
- 在新鲜、加热的10%FBS补充SIM卡中重新悬浮细胞,获得2 x 106细胞/mL的浓度。加入适量的CuSO4以保持500μM浓度。
- 将200-500 μL的再悬浮细胞转移到多孔活细胞室的一口井中,并放置在倒置荧光显微镜上。在成像实验前,让细胞在腔室中沉淀15-30分钟。
注:活细胞室孔可以预涂有多L-莱辛,以增加粘附性。
4. 设置活细胞成像程序
注:本实验中的实时细胞成像是使用倒置成像系统及其相关软件(例如,奥林巴斯IX83与细胞传感器尺寸软件包一起完成的)。细节将因显微镜制造商和软件包而异;因此,下面列出了一般准则和操作。
-
使用运行倒置荧光显微镜的软件,准备一个程序,用于随着时间的推移对细胞(或细胞)进行成像。
- 通过双击软件桌面图标打开软件。
- 通过单击"文件"创建新实验文件,然后单击"新建实验文件"。
- 首先,插入一个时间推移循环,在哪些图像上拍摄。为此,单击图标栏中的秒表图标(延时循环图标)。在"实验管理器"选项卡中设置 s 中的间隔,然后通过将所需的总实验长度 (s) 除以间隔来设置同一选项卡中的周期数。允许循环在所需的总时间段内重复。
注:通常,图像每 30-60 s 拍摄一次,时间为 3-4 小时。 - 在延时循环层内,插入红外焦点检查以保持目标的聚焦(如果可用)。为此,通过单击带有双箭头图标的正方形(移动 XY 图标)并从下拉菜单中选择 Z 漂移补偿,在延时循环图层中添加 Z漂移补偿(ZDC) 步骤。
注:术语红外对焦检查是指使用红外脉冲在目标与滑动/成像室之间保持恒定距离的系统。许多显微镜都有这样的系统,每个都有自己的专有名称。读者应查阅操作手册或代表,了解具体的命名详情。 - 红外对焦检查后,在程序中添加一个步骤,通过首先插入 z 堆栈步骤,然后指定通道,在 3-5 z 堆栈上获取多通道图像(例如,用于 GFP 的 FITC 和 mCherry 的 TRITC)。为此,通过单击"色轮"图标(多通道组图标),在延时循环图层中添加多通道组图层。接下来,通过单击3 层图标(Z堆栈循环图标)在多通道组图层中添加 Z-Stack 循环图层。在"实验管理器"选项卡中设置所需的步长大小和切片数,并将每个通道的曝光率设置尽可能低,以尽量减少光漂白。
注:LED 的 z 堆栈间隔、曝光时间和传输百分比可能有所不同。在这些实验中,使用三个 z 堆栈,范围为 3 μm,每个堆栈之间的间隔为 1 μm。定义单元格的中心,而不是顶部和底部,往往导致最佳结果。然后,在曝光 50 ms 和 50% 的传输率下收集每个通道的图像,并且没有中性密度 (ND) 滤波器。
5. 使用活细胞成像程序进行图像分割细胞
注:S2细胞不需要CO2,在23-27°C下以最佳方式生长。所有成像均在环境温度下进行,房间内控制良好。
- 使用眼部,找到井的顶部(或底部)角落,并使用 mCherry(β-图布林)通道将目标(40-60x 油浸)聚焦到细胞上。
- 沿井的顶部(或底部)扫描以远离垂直井分线。
注:太靠近井分线成像可能会干扰红外焦点检查。 - 查找 G2 后期或早期 M 相(前相)的细胞(或细胞)。
注:这些细胞可以通过存在两个"类似恒星"的微管结构(中位体)和一个完整的细胞核(由细胞内的折射光的圆形区域表示)来识别,这两种结构都很容易使用β-图布林(红色)进行区分。激励过滤器。在早期的相间选择细胞(由一个或零容易可见的中位体)显著,由于G2/M进展的滞后,可能导致浪费时间。相反,在NEBD之后选择细胞会妨碍对早期主轴装配和染色体动力学的线粒时间的精确计算和成像。此外,S2 细胞通常包含 >2 中心体。虽然这些通常集群成双极主轴12,建议用户避免这些细胞,除非其他需要的实验设计。图片和讨论适当的单元格选择可在代表结果部分找到。 - 单击"实时视图"按钮开始查看软件屏幕中的单元格。使用显微镜的精细对焦旋钮,对感兴趣的细胞(或细胞)进行聚焦。单击"设置对焦"按钮设置红外对焦检查。
- 单击"开始"按钮启动时差成像程序。通过选择所需的通道并调整平均像素强度来清楚地看到感兴趣的单元格(或单元格),根据需要调整直方图。
- 允许程序运行,在15-20分钟后检查细胞,以确保核包络故障(NEBD)发生,这是由圆形黑暗消失决定的,该物质位于细胞中心附近的折射点。
- 继续允许程序运行,间歇性地检查(每 15-20 分钟一次),以确定是否发生了抗性异常。如果总时间段中还有更多时间,此时停止程序并保存该文件。或者,如果端相和细胞因子事件感兴趣,则允许程序运行足够的时间,以便对这些过程进行成像。
-
通过记录 NEBD (tNEBD)的时间(以分钟)和初始染色体分离时间(taastage开始)以最小值,并从 taastage 开始中减去 tNEBD,以分钟进行 NEBD 到 aastage 发病时间的分析内溴到给定细胞的一个阶段开始时间。
- 为此,请单击帧向上按钮并注意发生 NEBD 和 aastage 的帧,减去这些帧以确定过移帧的总数,并将其乘以成像帧之间的时间间隔。
- 继续扫描预分细胞,以获得给定条件的多个 n。细胞在初始沉降后,可在活细胞室中成像长达12小时。
结果
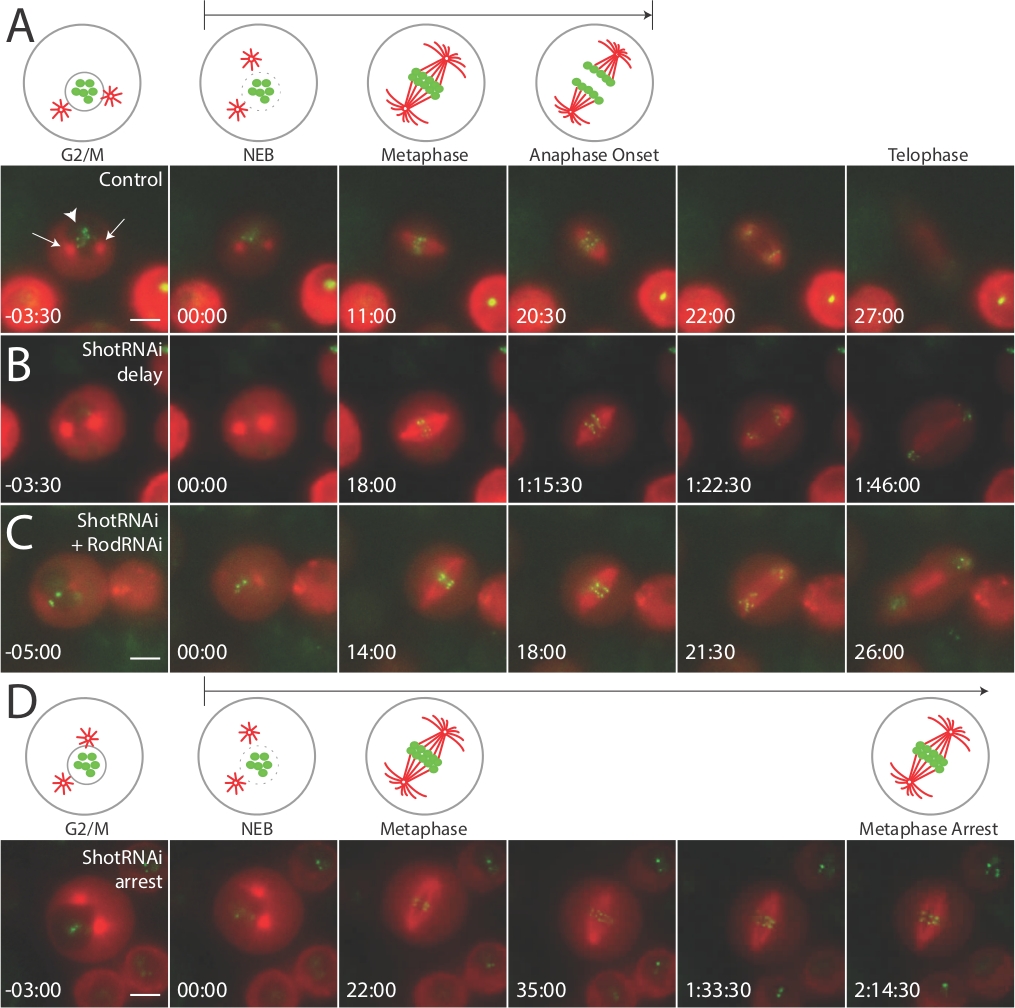
我们上面描述的方法将导致对经历细胞分裂的果蝇S2细胞的识别和成像。即将分裂(即,在进入M相之前)的细胞可以通过存在两个中心体和由折射光指示的完整核和在β-tubulin通道中观察细胞内的较暗点来瞄准(图1)。最左边的面板,红色通道;图 1A中的箭头 。我们估计大约 2-5% 的细胞属于此类别,在成像实验之间,我们通常最多花费 2-3 分钟扫描另一个合格的细胞。NEBD可以通过这个暗点的消失来可视化,导致细胞质的均匀着色(图1,面板标记为"00:00",红色)。在NEBD之后,只需记录这些事件相对于NEBD的时间点,即可测量每个细胞形成主轴、大会染色体和分离染色体的时间。
我们利用这些分部来评估线粒度,特别是NEBD到一个阶段开始,注意到染色体开始分离的点。多数 (+90%)铜诱导细胞,表示两个mCherry:β-图布林和GFP:CID。一小部分细胞(±10%)只表示一个标记,或者既不表示一个标记。在细胞选择过程中避免这些细胞。通常,S2细胞显示NEBD到发期开始时间在20-30分钟之间(图1A,控制)。接下来,我们用dsRNA对短停止(Shot)进行治疗,这是一种我们怀疑可能影响细胞周期动力学的活性素-微管交联蛋白。事实上,在拍摄后,击倒细胞表现出显著的线粒延迟(图1B,ShotRNAi延迟),许多细胞在元相中被抑制,在整个2-3小时的成像实验中从未过渡到Aastage(图1D),肖特纳伊被捕)。我们推断这种延迟/停止现象可能是由于主轴装配检查点(即 M 相检查点)的激活造成的。为了直接检验这个假说,我们共同处理细胞与dsRNA对射击和粗糙交易(Rod),这个检查点13的重要组成部分。这导致抑制逮捕表型,导致NEBD到Aastage时间类似于控制(图1C,ShotRNAi_RodRNAi)。因此,这种活成像协议允许我们得出结论,Shot 是及时 M 相级进展所必需的,并且其丢失导致检查点激活,从而延迟或逮捕线粒细胞。

图1:实时成像揭示S2细胞中线粒检查点激活的细胞周期缺陷。
在所有条件下,对S2细胞进行活细胞成像,通过诱导GFP:CID和mCherry:_Tubulin进行稳稳地共染,如本文所述。(A) 卡通在线粒体中说明重要地标,相应的图像从指示基因型的代表性电影中显示,其时间点相对于 NEBD (t=00:00) 表示。控制细胞通常在20-30分钟内进入抗相。 ShotRNAi处理的细胞显示NEBD-抗相延迟(B),并经常经历元相阻滞,从未在成像实验(D)内进入抗相。与 ShotRNAi 和 RodRNAi(主轴组件的一个组件)共同处理细胞,抑制 Shot 表型,导致类似于控制细胞 (C ) 的抗相发病动力学。a中的箭头表示"类似恒星"的中位结构,大箭头表示原子核。每个刻度条表示 5 微米。此图经10 (https://www.molbiolcell.org/info-for-authors)许可后调整和重新发布。请点击此处查看此图的较大版本。
{kind=link}
视频 1:"控制"S2 单元分裂的延时电影。视频显示"控制"基因型中 S2 细胞分裂的代表性电影。此影片对应于图 1A。此视频经过 10 (https://www.molbiolcell.org/info-for-authors)的许可后改编和重新发布。请点击此处下载此视频。
视频 2: 延迟的"ShotRNAi" S2 细胞分裂的延时电影。视频显示S2细胞分裂的代表性电影在"ShotRNAi"基因型,导致一个型板状线粒延迟。此影片对应于图 1B。请点击此处下载此视频。
视频 3: "ShotRNAi_RodRNAi"S2细胞分裂的延时电影。视频显示S2细胞分裂的代表性电影在"ShotRNAi_RodRNAi"基因型。此影片对应于图 1C。此视频经过 10 (https://www.molbiolcell.org/info-for-authors)的许可后改编和重新发布。请点击此处下载此视频。
视频4:被捕的"ShotRNAi"S2细胞师的延时电影。视频显示S2细胞分裂的代表性电影在"ShotRNAi"基因型,导致一个型状线性逮捕。此影片对应于图 1D。请点击此处下载此视频。
讨论
确定适当的单元格
成像分离S2细胞的关键是首先找到正确的细胞。时间可以浪费成像细胞,被误认为是准备分裂,但未能在合理的时间范围内这样做。细胞必须被识别,有两个明显和可见的中心体和一个完整的细胞核。中位体必须有微管纤维从它们发出,给他们一个"明星般的"外观。完整的原子核在调整对焦时折射光,也使细胞的近似中心在外观上变暗。原子核还将包含GFP:CID"点",这是另一个显著特征,有助于识别它。应避免使用 >2 中心体的细胞、没有从其中散发的图布林纤维的图布林蓬塔或只有一个中心体的细胞。此外,如果两个中心体是可见的,但一个核不是,NEBD已经发生,并且细胞的分裂太先进,如果需要完整的M相分析,无法成像。一旦找到适当的细胞,启动成像的速度至关重要,因为NEBD发生得很快(通常在3-5分钟内),延迟成像的开始可能导致错失机会。快速聚焦成像软件中的细胞,以便两个中心体都可见,或者如果中心体彼此离平面,则设置两者之间的焦点,这样当在定义中心上方和下方收集 Z 堆栈时,可以捕获每个中心点。聚焦后,立即使用红外对焦检查(如果可用)在舞台和活细胞室表面之间设置偏移,然后开始成像程序。尽管此处介绍的协议是专门为评估 NEBD 到抗相动力学的实验而定制的,但简单的修改可能适合研究替代线动事件的读者。对于 NEBD 到元相和抗相到端相成像,我们建议 10s 的图像捕获间隔,因为这些过程是高度动态的,并且在 S2 单元中快速发生(5-10 分钟)。在S2细胞中,从相向过渡(我们典型的实验焦点)是一个较长的过程(20-30分钟),我们每隔30秒收集图像。最后,端相通细胞因子在S2细胞中发生得相当缓慢,我们建议60秒的间隔为这个大约长达一小时的过程提供足够的分辨率。
在整个成像计划中保持重点
如果红外对焦检查设备不可用,请确保避免碰撞其坐的显微镜或表,因为这可能导致细胞在成像程序期间失去焦点,从而导致模棱两可、无法解释的结果。用 Poly-L-lysine 预涂活细胞室孔有助于细胞粘附,这有助于避免细胞运动,对于无需红外对焦检查的设置特别有用。此外,包括减震平台或空气表在内的设置可进一步帮助避免细胞离开焦点。最后,一些软件程序具有暂停功能,允许用户重新聚焦和恢复程序。
成像多个单元
有助于积累数据的是,通常多个准备分割的单元可以放置在相同的视场中进行成像。这对于细胞具有长M相持续时间或长时间阻尼(例如,诱导SAC激活的条件)的实验特别有用。对于这些细胞,我们通常图像,直到细胞被光漂白(约2-3小时),但逮捕细胞的全部时间应该由研究人员决定。有时,多个预分单元可以位于不同的焦平面上,在这种情况下,添加 z 堆栈以容纳所有单元格可能很有用。添加更多 z 堆栈还有助于提高分辨率。但是,在执行此操作时应小心谨慎,因为它会使细胞受到更多的辐射,并导致更快的光漂白,并导致硬盘存储设备上的文件大小过大。
避免光漂白和光毒性
我们的方法描述了暴露时间、成像间隔、z 堆栈和 LED 光源传输百分比的特定设置,这些设置通常适用于我们的实验设置和预期结果。由于显微镜和实验目标可能不同,可能对我们的系统工作良好,可能会导致其他系统过早光漂白或光毒性。光损伤可能导致主轴运动不足、微管分裂、微管动力学缺陷以及主轴检查点激活时间过长。避免此类陷阱的一种潜在方法是限制暴露时间。大多数现代相机现在具有宽动态范围,直方图可以调整,即使在非常低的曝光下也能可视化结构。另一种技术是增加成像间隔(例如,图像之间的几分钟),以便减少细胞在总收集间隔内暴露在光下的次数。这在长时间(数小时)的成像中特别有利,而且还有助于减小此类实验的文件大小。限制 Z 堆栈的数量也有助于减少光线照射,使用 2 个甚至仅 1 个而不是 3。此外,调整光的发射百分比(对于 LED 光源),或使用中性密度 (ND) 滤波器(对于卤素灯和 LED 光源)将降低光强度,并结合更长的曝光时间可以保持可见性.通过选择具有已分离的中位体的细胞,整体暴露可以受到限制,因为这些细胞通常在开始成像后快速(一两分钟内)进入线粒体。也可以限制在NEBD之前允许细胞成像的时间。我们的实验室通常将此时间设置为 10 分钟,但用户可以轻松施加更保守的限制。此外,我们建议的一个更"侵入性"的措施是针对 SAC 成分(如 Rod 或 Mad2)的 dsRNA 进行治疗。如果逮捕表型是由于非特异性细胞损伤(例如,有缺陷的微管动力学),与原始实验中感兴趣的基因的真诚效果相比,这种处理不太可能抑制逮捕。
未来方向
此处描述的方法可用于相对简单的表观显微镜,以快速成像活细胞分裂,并可轻松调整以适应研究人员的特定实验设计和目标。S2和其他果蝇细胞已经发布了几种很好的培养方法,RNAi敲除方法,瞬态转染,荧光显微镜。16,17.我们的协议有几个优点.(1) 使用双稳定细胞系 (GFP:CID, mCherry: α-图布林) 同时标记两个关键线粒结构,避免瞬态转染的麻烦和潜在并发症,并确保几乎所有细胞都会表达荧光标记作为成像的潜在候选者。(2) 对线粒体的适应增加了已公布的协议,用于检查细胞骨骼动力学,在活动细胞,非分裂细胞,并延伸到实时检查线形事件。尽管我们相信我们的技术是一个强大的技术,增加了其他人的剧目,总是有可以作出改进。我们发现难以成像的细胞分裂是一个特定的领域,就是中心分裂。这发生在NEBD之前,很难确定当一个中心体准备分成两个。使用荧光细胞周期标记(Cyclin A 和 Cyclin B)可以帮助解决此问题,但代价是可用于可视化细胞分裂成分的通道。我们尝试可视化中心体分割的最佳策略是向成像程序添加多点采集(在 XY 平面中定义到图像的不同点),但这可能会导致大型文件,并且可能要求(取决于点数)增加图像收集之间的间隔,降低结果影片的时间分辨率。另一种解决方案可能是在所需的细胞周期阶段使用针对细胞周期调节器的药物同步细胞,随后在成像前进行冲洗,尽管这些方法在S2细胞中可能并不可靠。
此处介绍的协议为未来在活细胞成像中的应用奠定了基础。通过改进的成像和软件技术,使用更高容量的多腔板可以真正实现高吞吐量调整。这种创新将使大型RNAi屏幕,如那些已经实现的固定制剂12,18,更可操作的活细胞格式。小分子药物屏幕也可以被设想为识别针对细胞分裂过程的新型化合物的手段。改进荧光团和光学滤光片还可能导致多个线粒体组分(不仅仅是我们描述的两个)成像,从而能够对特定的线粒稳压器及其与DNA和/或主轴的相互作用进行成像。例如,用荧光检测器生成细胞,在DNA损伤后或凋亡期间表达,将是识别参与这些过程的新基因的有用工具。类似的方法可以用来检查细胞周期动力学使用荧光标记环素19的表达。
披露声明
作者没有什么可申报的。
致谢
这项工作由美国国家卫生研究院(R01 GM108756)资助。我们感谢加里·卡彭(加州大学伯克利分校)慷慨地为我们提供了一个GFP:CID S2细胞系股票,我们的GFP:CID/mCherry:_Tubulin线产生10。
材料
| Name | Company | Catalog Number | Comments |
| Bright-Line Hemacytometer | Sigma-Aldrich | Z359629-1EA | for cell counting |
| cellSens imaging software | Olympus | ||
| CELLSTAR Cell Culture Flask, 50 mL, 25 CM2, PS, Red Filter Screw Cap, Clear, Sterile, 10 PCS/BAG | Greiner Bio-One | 690175 | |
| CELLSTAR Cell Culture Multiwell Plate, 6 well, PS, Clear, TC, Lid with condensation rings, sterile, single packed | Greiner Bio-One | 657160 | |
| Centrifuge 5804 R | eppendorf | Cat. 022623508 | |
| Copper(II) sulfate pentahydrate, minimum 98% | Sigma-Aldrich | C3036-250G | |
| Corning Fetal Bovine Serum | Fisher Scientific | MT35015CV | |
| Effectene Transfection Reagent 1 mL | Qiagen | 301425 | for transient transfection |
| IX-83 Inverted Epifluorescent Microscope | Olympus | ||
| LabTek Chambered Slide Insert | Applied Scientific Instruments | I-3016 | |
| MEGAscript T7 Transcription Kit | Thermo Fisher Scientific | AM1334 | For dsRNA production |
| MOXI Z Mini Automated Cell Counter Kit | ORFLO | MXZ001 | for cell counting |
| MS-2000 XY Flat-Top Automated Stage and Controller | Applied Scientific Instruments | ||
| Nunc Lab-Tek II Chambered Coverglass (no 1.5 borosilicate glass) 8-well | Thermo Fisher Scientific | 155409 | |
| Orca-Flash 4.0 LT Camera | Hammamatsu Photonics K.K. | C11440-42U | |
| pMT/V5-His A Drosophila Expression Vector | Thermo Fisher Scientific | V412020 | |
| Poly-L-Lysine | Cultrex | 3438-100-01 | |
| Purifier Logic+ Class II, Type A2 Biosafety Cabinets | Labconco | 302310000 | |
| Schneider's insect medium | Sigma-Aldrich | S0146-100ML | |
| Spectra Tub Centrifuge Tubes | VWR | 470224-998 | |
| Uner Counter BOD Incubator | Sheldon manufacturing (VWR) | 89409-346 | |
| X-CITE 120 LED | ExcelitasTechnologies | Led light source | |
| Z -Drift Compensator (ZDC) | Olympus | infrared focus check |
参考文献
- Ragkousi, K., Gibson, M. C. Cell division and the maintenance of epithelial order. Journal of Cell Biology. 207 (2), 181-188 (2014).
- Aldaz, S., Escudero, L. M., Freeman, M. Live imaging of Drosophila imaginal disc development. Proceedings of the National Academy of Science U. S. A. 107 (32), 14217-14222 (2010).
- Cabernard, C., Doe, C. Q. Live imaging of neuroblast lineages within intact larval brains in Drosophila. Cold Spring Harbor Protocols. 2013 (10), 970-977 (2013).
- Lerit, D. A., Plevock, K. M., Rusan, N. M. Live imaging of Drosophila larval neuroblasts. Journal of Visualized Experiments. (89), (2014).
- Morris, L. X., Spradling, A. C. Long-term live imaging provides new insight into stem cell regulation and germline-soma coordination in the Drosophila ovary. Development. 138 (11), 2207-2215 (2011).
- Prasad, M., Jang, A. C., Starz-Gaiano, M., Melani, M., Montell, D. J. A protocol for culturing Drosophila melanogaster stage 9 egg chambers for live imaging. Nature Protocols. 2 (10), 2467-2473 (2007).
- Restrepo, S., Zartman, J. J., Basler, K. Cultivation and Live Imaging of Drosophila Imaginal Discs. Methods in Molecular Biology. 1478, 203-213 (2016).
- Tsao, C. K., Ku, H. Y., Lee, Y. M., Huang, Y. F., Sun, Y. H. Long Term Ex Vivo Culture and Live Imaging of Drosophila Larval Imaginal Discs. PLoS One. 11 (9), e0163744 (2016).
- Rogers, S. L., Rogers, G. C. Culture of Drosophila S2 cells and their use for RNAi-mediated loss-of-function studies and immunofluorescence microscopy. Nature Protocols. 3 (4), 606-611 (2008).
- Dewey, E. B., Johnston, C. A. Diverse mitotic functions of the cytoskeletal cross-linking protein Shortstop suggest a role in Dynein/Dynactin activity. Molecular Biology of the Cell. 28 (19), 2555-2568 (2017).
- Rodriguez, E. A., et al. The Growing and Glowing Toolbox of Fluorescent and Photoactive Proteins. Trends in Biochemical Sciences. 42 (2), 111-129 (2017).
- Kwon, M., et al. Mechanisms to suppress multipolar divisions in cancer cells with extra centrosomes. Genes and Development. 22 (16), 2189-2203 (2008).
- Basto, R., Gomes, R., Karess, R. E. Rough deal and Zw10 are required for the metaphase checkpoint in Drosophila. Nature Cell Biology. 2 (12), 939-943 (2000).
- Currie, J. D., Rogers, S. L. Using the Drosophila melanogaster D17-c3 cell culture system to study cell motility. Nature Protocols. 6 (10), 1632-1641 (2011).
- Lu, W., Del Castillo, U., Gelfand, I. V. Organelle transport in cultured Drosophila cells: S2 cell line and primary neurons. Journal of Visualized Experiments. (81), e50838 (2013).
- Yang, J., Reth, M. Drosophila S2 Schneider cells: a useful tool for rebuilding and redesigning approaches in synthetic biology. Methods in Molecular Biology. 813, 331-341 (2012).
- Zhou, R., Mohr, S., Hannon, G. J., Perrimon, N. Inducing RNAi in Drosophila cells by soaking with dsRNA. Cold Spring Harbor Protocols. 2014 (5), (2014).
- Goshima, G., et al. Genes required for mitotic spindle assembly in Drosophila S2 cells. Science. 316 (5823), 417-421 (2007).
- Sakaue-Sawano, A., et al. Visualizing spatiotemporal dynamics of multicellular cell-cycle progression. Cell. 132 (3), 487-498 (2008).
转载和许可
请求许可使用此 JoVE 文章的文本或图形
请求许可This article has been published
Video Coming Soon
版权所属 © 2025 MyJoVE 公司版权所有,本公司不涉及任何医疗业务和医疗服务。