
Method Article
在小鼠的文室内使用光遗传学的两种不同的实时位置偏好范式
摘要
在这里,我们提出了两个易于遵循的分步协议,用于在小鼠中使用光遗传学的放置偏好范例。使用这两种不同的设置,偏好和回避行为可以在同一设备中以高空间和时间选择性,以简单明了的方式进行可靠的评估。
摘要
了解神经元激活如何导致特定的行为输出是现代神经科学的基础。将啮齿动物中的光遗传学与经过验证的范式中的行为测试相结合,可以实时测量不同神经元在具有高空间和时间选择性时刺激时的行为后果,从而建立神经元激活和行为之间的因果关系。在这里,我们描述了实时位置首选项 (RT-PP) 范例的分步协议,这是经典条件位置首选项 (CPP) 测试的修改版本。RT-PP在三节装置中执行,可用于回答特定神经元群的光遗传学刺激是否有益或厌恶。我们还描述了 RT-PP 协议的替代版本,即所谓的中性隔间首选项 (NCP) 协议,可用于确认厌恶。这两种方法基于源自行为药理学的经典方法论的延伸,以及神经科学领域中光遗传学的最近实施。除了实时测量位置偏好外,这些设置还可以提供有关条件行为的信息。我们提供易于遵循的分步协议以及我们自己的数据示例,并讨论应用这些类型的实验时需要考虑的重要方面。
引言
光遗传学,一种现代神经科学实验工具,光被用来控制神经元活动,近年来在了解特定神经元群体如何影响行为1,2,3方面有了重大进展。光遗传学的突出的空间和时间选择性使得激发或抑制感兴趣的细胞群与行为输出2、3之间建立因果关系。光遗传学的空间选择性通常通过Cre-Lox系统得到保证,其中Cre重组酶的活性导致Lox位点之间存在的任何DNA序列的重组,即所谓的絮状等位基因(由lox位点)4。在光遗传学中使用Cre-Lox系统的目标是在感兴趣的特定神经元中实现编码光遗传学蛋白酶的等位基因的表达,同时使周围的神经元缺乏表达。Opins 是光敏蛋白,在光刺激特定波长时允许电子流动,通过调节下游效应器通路影响神经兴奋性或影响细胞功能。正在不断开发在作用(兴奋、抑制、调节)、机制、光波长激活和动力学特性5中不同的新型蛋白酶变体,以满足特定实验方法的需要。关于兴奋性,使用去极化或超极化蛋白酶决定神经元的活动(分别激励或抑制)在光刺激在一个特定波长传递到大脑3。
选择性促进者活动将Cre重组酶的表达导向感兴趣的神经元。通过实施感兴趣的蛋白酶的絮状等位基因,Cre介导的重组将确保蛋白酶在神经元中选择性地表达,共同表达Cre重组酶3,6。这种使用双转基因来引导空间选择性已被证明在光遗传学方面非常有效。因此,虽然光刺激激活蛋白酶广泛通过与光源(LED或激光)3相连的脑内植入光纤传递,但只有表达Cre重组酶和絮凝酶的神经元才会对这种刺激做出反应。啮齿动物的Cre-Lox系统可以通过仅使用转基因(Cre重组酶和絮凝蛋白结构在转基因动物中编码),仅病毒注射(Cre重组酶和絮凝蛋白的DNA结构都通过病毒载体提供),或两者的组合(例如, Cre重组酶是由一种转基因动物编码的,它被注射携带浮游生物的病毒5。浮花蛋白DNA结构通常与报告基因在框架中克隆,以在组织部分实现Cre介导重组的可视化。虽然光遗传学也可以在大鼠身上进行,但提出的协议已经为小鼠生成了。为简单起见,同时携带Cre重组酶和絮凝蛋白的小鼠将被称为"光遗传学小鼠"。在下面描述的协议中,光遗传学小鼠是由混合转基因(在两个不同的启动子控制下的Cre重组酶)和病毒(使用腺相关病毒AAV,以提供浮游菌蛋白酶DNA结构-在我们的例子ChR2/H134R)方法。获取和维护转基因小鼠线是该方法的重要组成部分。可出于各种目的生产Cre-驱动程序和絮状蛋白酶转基因小鼠,或购买(如果市售),以及一系列携带DNA序列的病毒,这些病毒编码Cre重组酶和不同形式的絮凝蛋白。
光遗传学与行为测试相结合已被证明是研究不同大脑区域或离散神经元群体,特别是行为类型的作用的宝贵工具。在奖励相关行为方面,光遗传学已经验证了先前在行为药理学和实验心理学领域的发现,也使得某些神经元如何影响行为的时空相关解剖有了一个新的水平。在一些研究中用于评估奖励相关行为的一种方法是称为条件地点偏好 (CPP) 的经典方法的修改版本。经典CPP已经被用来评估滥用药物的奖励或厌恶性质,通过他们的能力诱导巴甫洛夫人与环境的线索7,8。在巴甫洛夫语中,这种药物是一种无条件的刺激,因为它可以引起接近或戒断,如果它是奖励或厌恶,分别。药物与各种中性刺激的连续配对,本身不会引起任何反应,可以导致接近或退出仅仅在呈现以前的中性,但在配对后,所谓的条件刺激9。CPP 分析通常在包含两个相同大小的隔间的装置中执行,但每个隔间都由不同的特征定义,例如地板纹理、墙壁图案和照明(中性刺激)。两个隔间通过走廊或隔间之间的开口连接。在调理期间,受试者(通常是小啮齿动物)接受药物的被动注射,同时限制在两个主要隔间和盐水之一,而限制到另一个隔间。当受试者被允许自由探索整个仪器时,药物的奖励效果随后在无药物会话中进行评估。在以前药物配对的隔间中花费的时间(条件反应)被认为是反映巴甫洛夫学习机制,在药物的有益效果和与其给药(条件刺激)相关的隔间线索之间介导。如果动物花更多的时间在药物配对的隔间,药物已经诱导一个有条件的地方偏好,这意味着它有奖励作用的行为。另一方面,如果药物被视为厌恶,动物将避免药物配对的隔间,并花更多的时间在盐水配对的隔间,表明有条件的地方厌恶(CPA)8,9,10,11。
由于光遗传学可以实现控制神经元活动在"实时",使用的行为范式类似,但不同于,CPP设置允许测量位置偏好在直接神经元激活。因此,视点遗传学驱动的地点偏好分析通常被称为实时位置偏好 (RT-PP) 分析范式。在RT-PP范式中,通过Cre-Lox系统对不同神经元的光遗传学刺激取代了在经典CPP中执行的药物的系统传递,因此RT-PP范式代替测量光遗传学引起的神经元刺激被视为有回报或厌恶。目前的描述将侧重于光遗传学小鼠,但也光遗传学大鼠可以使用类似的协议进行测试。
RT-PP范式中的光遗传学小鼠可以在整个设备中自由移动,并且在整个会话过程中记录行为,而不是像经典 CPP 范式那样一次调节到一个隔间。进入其中一个隔间与颅内光刺激配对。在适当的光刺激参数下,表达兴奋蛋白的神经元将被激活。如果光刺激被认为是有益的,光遗传学小鼠将留在光对隔间,而如果光刺激被认为是厌恶,小鼠将退出隔间,以逃避刺激。这种类型的分析允许评估或有学习:受试者可以通过进入隔间触发光刺激,从而触发神经元激活,并通过退出隔间来停止刺激,类似于工具任务期间的杠杆按下。此外,在后续的会话中,可以评估关联学习机制,在没有刺激的情况下测量每个隔间中的时间。这样,研究人员就可以分离对感兴趣的神经元的刺激的立即回报效应和与之相关的关联记忆形成12。
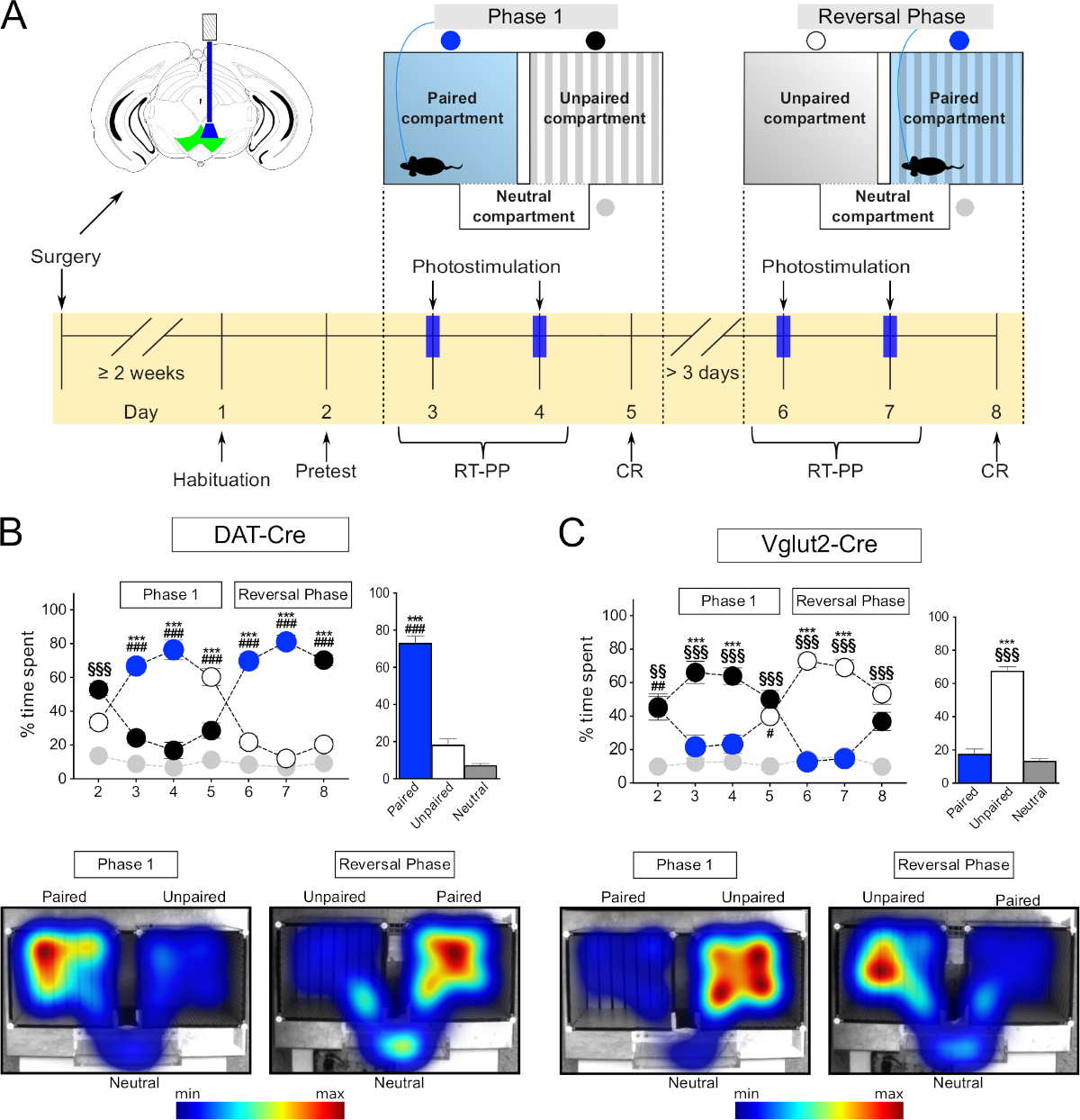
在目前的研究中,我们描述了两个分步设置协议,用于光遗传学驱动的自由移动小鼠的放置偏好行为。第一个协议描述了三节装置内的RT-PP,并基于Root及其13号同事和其他作者最近提出的协议进行了概述。实验由两个阶段组成,包括几个每日会话(如图1A所示)。每个会话都设计用于不同的目的,并且与隔间耦合刺激的参数也会相应地更改。第一个会话"预测试"用于评估主体对任一隔间的初始偏好。当连接到配线时,允许受试者在没有刺激的情况下自由探索设备15分钟。如果对任意一个隔间的初始首选项超过 80%,则鼠标将从分析中排除,因为初始侧偏置可能会扭曲分析。在"预测试"之后,"第 1 阶段"开始。第一部分包括连续两个,每天,30分钟的"RT-PP"会话。在"第1阶段",光遗传学小鼠通过配线连接到激光源,并放置在竞技场中自由探索它。老鼠在进入其中一个主要隔间时接受颅内激光刺激。可以执行试验实验,以确定哪个隔间将被指定为激光配对,哪个隔间未配对。如果刺激被证明是有益的,激光将在"预测试"期间耦合到最不喜欢的隔间,如果刺激是厌恶的,则最优先的隔间。因此,提出的RT-PP协议遵循一种偏置设计,即激光刺激不是随机分配给两个主要隔间中的任何一个(无偏设计),而是为了避免鼠标的任何初始偏好而选择。进入另一个主隔间或连接两个主隔间的中性隔间不会引起颅内光刺激,因此不会轻对。这些会话允许实时评估特定神经元群体的刺激的有益或厌恶特性。在"第一阶段"的最后一天,15分钟的会话,没有任何刺激发生。本课程旨在讨论由刺激与接收环境之间的关联学习产生的有条件的反应("CR")。在"第1阶段"之后至少三天,"反转阶段"发生,其结构与"第1阶段"相同,但以前未配对的主隔间现在与光刺激配对。与"第一阶段"的情况一样,两个刺激会议之后是"CR"会话。"反转阶段"用于确认小鼠的行为取决于光遗传学刺激,与随机参数无关。RT-PP 实验的每个会话都必须在跟踪软件中单独编程。当前协议描述了特定软件中的此类设置,但可以使用任何其他能够向光源发送晶体管-晶体管-逻辑 (TTL) 调制信号的跟踪软件。
第二个协议描述了一种称为"中性隔间首选项 (NCP)"范式的新型设置。RT-PP 的此修改协议利用了连接走廊的较小尺寸和透明度,由于其窄而透明的组成,鼠标自然避免了这种结构。通过将两个主隔间与光刺激配对,并且仅使走廊没有光刺激,NCP 设置可用于测试光遗传学刺激是否会迫使小鼠在走廊中花费更多时间以避免接收光遗传学刺激。通过将两个光对隔间中所花费的时间与走廊中所花费的时间进行比较,可以验证光遗传引起的厌恶。NCP 实验包括连续两次每日会话,其中光遗传学小鼠接受刺激(每次 30 分钟)以实时测量偏好,以及一次无激光会话(15 分钟),以评估与 RT-PP 中类似的条件响应协议。
下面提供的RT-PP和NCP协议最近在我们的实验室中进行了验证,研究位于腹腔内分区域(VTA)的不同类型的神经元如何参与与奖励相关的行为的各个方面12。在这里,为了举例说明RT-PP和NCP协议的实施,多巴胺运输机(DAT)-Cre19和环状谷氨酸转运器2(VGLUT2)-Cre20转基因小鼠被立体地注射了AAV,携带一个浮游通道二(ChR2)DNA结构植入VTA,然后一个光纤被植入VTA上方。使用提供的RT-PP和NCP协议对这些小鼠进行分析后获得的行为反应表明,在VTA内多巴胺能和谷氨酸神经元的激活如何导致不同的行为反应(图1)。
RT-PP 和 NCP 范例的分步协议提供了从转基因小鼠基因分型、立体病毒注射和光纤放置到激光控制和行为跟踪软件编程等信息评估。此外,还讨论了在刺激参数和实验方面对协议进行修改的建议,这些方面可能影响科学结果。虽然协议在VTA的上下文中描述,它们可以应用于任何大脑区域或神经元群体,只要相关光遗传学工具,如相关的Cre-驱动程序和絮凝蛋白,可用。
研究方案
本研究使用杂音DAT-Cre19和VGLUT2-Cre20小鼠进行,年龄超过8周,体重>20克。所有实验均根据瑞典(动物福利法 SFS 1998:56)和欧盟立法(ETS 123 公约和第2010/63/EU号指令)在当地动物伦理委员会的许可下进行。
1. 小鼠基因分型
- 使用耳打孔器进行耳检,用于转基因小鼠的基因分型。
- 准备耳冲剂,使用专用底漆进行聚合酶链反应 (PCR) 反应。
注:在本协议中,使用了Cre定向引物。- 在每个含有耳孔的1.5 mL管中加入75 μL的发液缓冲液(缓冲液1:250 mM NaOH,2 mM EDTA)。
- 在加热块中孵育在96°C下30分钟。
- 让样品冷却5分钟,然后加入75 μL的中和缓冲液(缓冲液2:400 mM Tris-HCl pHH 8.0)。
- 根据标准程序12、21使用适当的引物(此处:Cre FW 5' - ACGAGTAGGGTCGCAGA-3'、Cre REV 5' - ACCGAAGATATTTTTT-3')。
注意:在 PCR 发动机罩下的冰上工作,注意不要污染试剂和样品。- 准备 PCR 主组合。根据要分析的样本数量(包括适当的控制样本)将以下体积相乘。按以下顺序混合试剂进行单次 25 μL 最终体积反应:蒸馏水 (18.9 μL),10x 缓冲液与 MgCl2 (2.5 μL),10 mM dNTP 混合物 (0.5 μL)、10 μM 正向底漆 (1 μL)、10 μM 反向底漆 (1 μL)、5 U/μL DNA 聚合酶 (0.1 μL) 和模板 DNA (1 μL; 将添加到后续步骤中)。
注: 始终添加负数、阳性和空(无模板 DNA)控件,以确保有效结果。 - 在PCR管中加入24μL的主混合物。
- 在每个 PCR 管中添加 1 μL 模板 DNA(来自每只小鼠的耳孔)。
- 暂时将 PCR 管离心,以确保模板 DNA 位于主混合物内。
- 使用表 1中的循环程序使用热循环器执行 PCR。
- 准备 PCR 主组合。根据要分析的样本数量(包括适当的控制样本)将以下体积相乘。按以下顺序混合试剂进行单次 25 μL 最终体积反应:蒸馏水 (18.9 μL),10x 缓冲液与 MgCl2 (2.5 μL),10 mM dNTP 混合物 (0.5 μL)、10 μM 正向底漆 (1 μL)、10 μM 反向底漆 (1 μL)、5 U/μL DNA 聚合酶 (0.1 μL) 和模板 DNA (1 μL; 将添加到后续步骤中)。
- 准备一个甘蔗凝胶,使用电泳运行样品。
注: 大小取决于需要分析的样本数。- 在玻璃瓶中加入1x三酸乙酯-EDTA(TAE)缓冲液中,无agarose粉。加热在微波炉,直到甘蔗完全溶解,定期检查,它不沸腾。
注意:采取预防措施,避免烧伤。 - 让凝胶冷却到大约50°C,并加入核酸凝胶染色(0.5 μL/50 mL凝胶)。
- 将凝胶倒入含有井梳的铸造托盘中,将其留在室温下,直到完全凝固。轻轻拆下梳子。
- 用 1x TAE 缓冲液填充电泳罐,并将凝胶放入罐中。
- 在每个DNA样本中加入2μL的1xDNA加载染料。
- 在凝胶的第一个井中加载DNA阶梯4μL,然后继续加载剩余井中的样品的全部体积。
- 将电泳的电源设置为 140 V,运行 25-30 分钟。
- 将凝胶置于紫外线源下,并拍摄结果。
- 在玻璃瓶中加入1x三酸乙酯-EDTA(TAE)缓冲液中,无agarose粉。加热在微波炉,直到甘蔗完全溶解,定期检查,它不沸腾。
2. 立体手术
- 基因分型后,分离小鼠保持对Cre呈阳性。等到他们至少8周大,体重+20克,以进行手术。
- 消毒环境,消毒手术工具,在无菌条件下进行手术。
- 手术前30分钟,用镇痛药将小鼠皮下注射。
- 用异常鲁兰麻醉小鼠(在正常空气中诱导2~3%,麻醉维持1.5~2.0%)。通过轻轻捏住鼠标脚趾,通过测试无疼痛反射,确保达到足够的麻醉水平。相应地调整电子分体交付。
- 将鼠标放在立体装置上。添加眼部润滑剂,防止因干燥而眼部病变,并洗去颅骨顶部的头发。使用加热垫保持鼠标温度稳定。
- 在颅骨皮肤下注射100μL局部麻醉剂,5分钟生效。
- 通过酒精或无菌盐水与碘交替的三个循环应用来准备切口部位。使用无菌棉尖,从切口线向外启动应用。
- 用钳子轻轻抬起皮肤,用手术剪刀沿骨刻轴切开±1.5厘米,露出头骨表面。
- 使用棉棒,应用 H2O2溶液去除渗透液。
- 用无菌盐水冲洗头骨,并使用无菌棉尖施用器干燥。
- 找到布格玛和羊肉。
- 通过将注射针的尖端定位在立体框架上,在布雷格玛和 lambda 上调整,确定扁平的颅骨对齐。测量每个位置的通风口坐标并进行比较。当头骨是平的,腹腔坐标为布雷格玛和lambda是相同的。如果没有,请调整头部位置并再次进行测量。
- 找到并标记位置 (AP: -3.45 mm, ML: -0.2 毫米从布雷格马根据富兰克林和 Paxinos22) 注射克里依赖病毒和植入光纤将发生,并使用微钻打一个小孔。
- 使用精密泵将 400 nL 病毒加载到安装在立体仪器上的 10 μL 注射器中。
- 小心降低针头(34 G,下角),在VTA(AP:-3.45 mm, 根据富兰克林和Paxinos 22,ML-0.2毫米,从布雷格玛-0.2毫米,从头骨表面-4.4毫米,使用精密泵以100 nL/min的注射速率。
- 注射后,将针头留在原位10分钟,以便病毒扩散(图2A)。
- 慢慢从注射部位缩回针头。
- 使用微钻打小孔以安装锚定螺钉,以稳定光纤和牙科水泥复合物。
- 根据富兰克林和Paxinos22,再次取取胸膜坐标,并植入光纤(直径200微米,0.37 NA),在:AP:-3.45毫米,ML:-0.2毫米从布雷格玛和-4.0毫米从头骨表面(图2B)根据富兰克林和Paxinos 22。
- 用牙科水泥固定头骨上的纤维。在光纤 ferule 周围涂抹足够的水泥,将其固定在头骨上,但请注意将 3⁄4 mm 的 ferule 顶部留无水泥,以便连接配线(图 2C)。
注意:注意不要用水泥填洞,因为这可能导致脑组织损伤。可将止血材料添加到孔中,以防止这种情况发生。 - 使用组织胶或可吸收缝合线关闭任何开放伤口,让动物恢复至少两周。手术后,再服用一剂镇痛药12⁄24小时。
3. 设置激光源控制
- 使用单板微控制器控制激光源。使用适当的软件编写脚本。使用适当的连接电缆将脚本加载到微控制器板上。
注: 脚本应包括通过 TTL 框从跟踪软件产生的外部调制(输入),以及激光的输出来控制刺激参数。对于 20 Hz 频率的 10 ms 脉冲宽度,请使用补充编码文件中的脚本。 - 将主板连接到跟踪硬件的激光和 TTL 盒。
- 使用网络电缆将 TTL 盒连接到主板(提供脚本的引脚 5)(图3A,C)。
- 确保激光设置为通过外部调制进行控制,并使用 FC/PC 电缆将激光连接到电路板(用于给定脚本的引脚 13)(图 3B,C)。
- 将适当的销连接到电路板的接地部件。
- 将激光源连接到光纤。
- 将激光源连接到旋转接头 (图 3D)。
- 将配线 (图 3E) 连接到旋转接头.
- 将旋转接头稳定在设备上方,但位于记录区域之外。确保光纤配线的长度适合让鼠标在竞技场中移动时无困难(图 3F)。
4. 在跟踪软件中设置RT-PP方法的实验
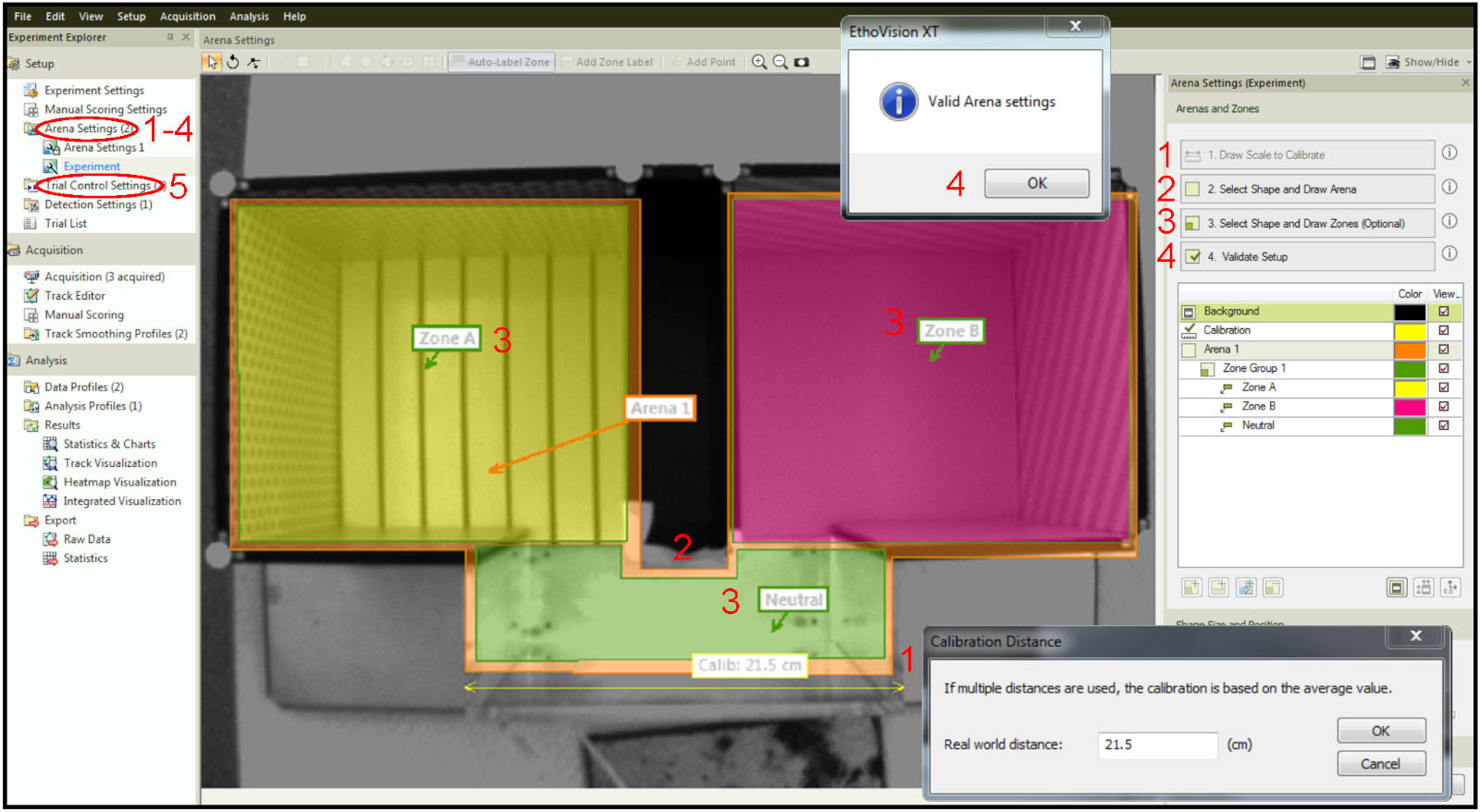
- 校准竞技场设置。使用标尺测量物理设备的特定部分,在"绘制比例到校准"选项卡下绘制一条与软件中图像上测量的零件对应的线,然后输入已知值(图 4中的步骤 1 )。
- 设计竞技场。绘制将记录小鼠移动的区域(图 4中的步骤 2 )。
- 创建区域。绘制最终将指定为激光配对、激光未配对和"中性"的区域(图 4中的步骤 3 )。
- 验证设置以确认没有冲突的参数,例如竞技场外的区域(图 4 中的步骤4)
- 在选项卡试验控制设置下设置实验参数(图 4中的步骤 5 )。
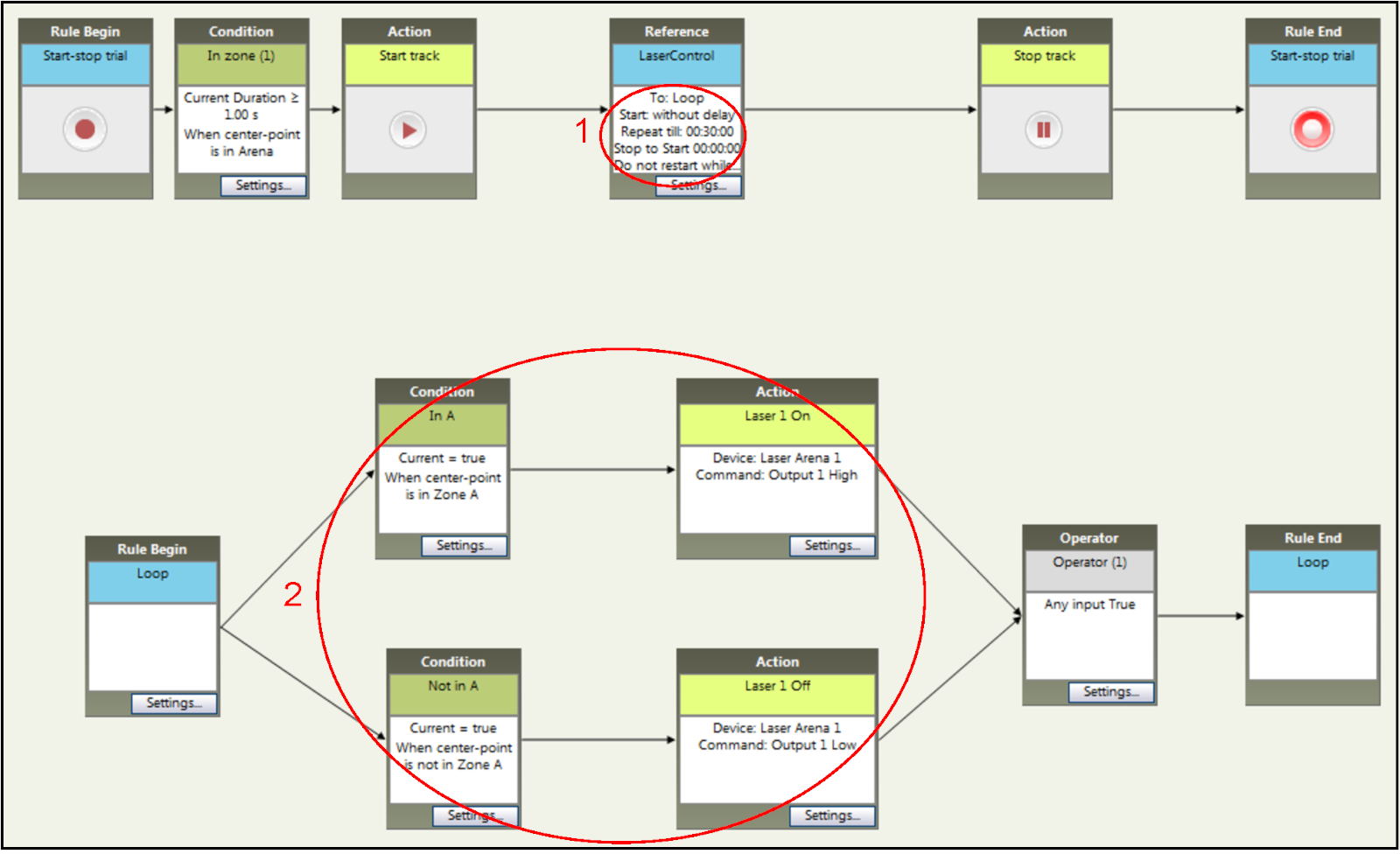
- 设置 30 分钟 RT-PP 会话的试验时间,如图5中的步骤 1 所示。
- 确保启用"硬件控制",将隔间指定为激光配对,其中鼠标进入将通过跟踪软件触发 TTL 信号到微控制器板。在图 5(步骤 2)中,激光配对隔间为隔间 A。对于反转阶段,切换隔间,使隔间 B 进行激光配对,并且隔间 A 取消配对。为此,在软件中用 B 替换 A,将 B 替换为 A。
5. 使用 NCP 方法修改设置以测试刺激的厌恶特性
- 按照前面所述的步骤 4.1_4.4 进行操作。
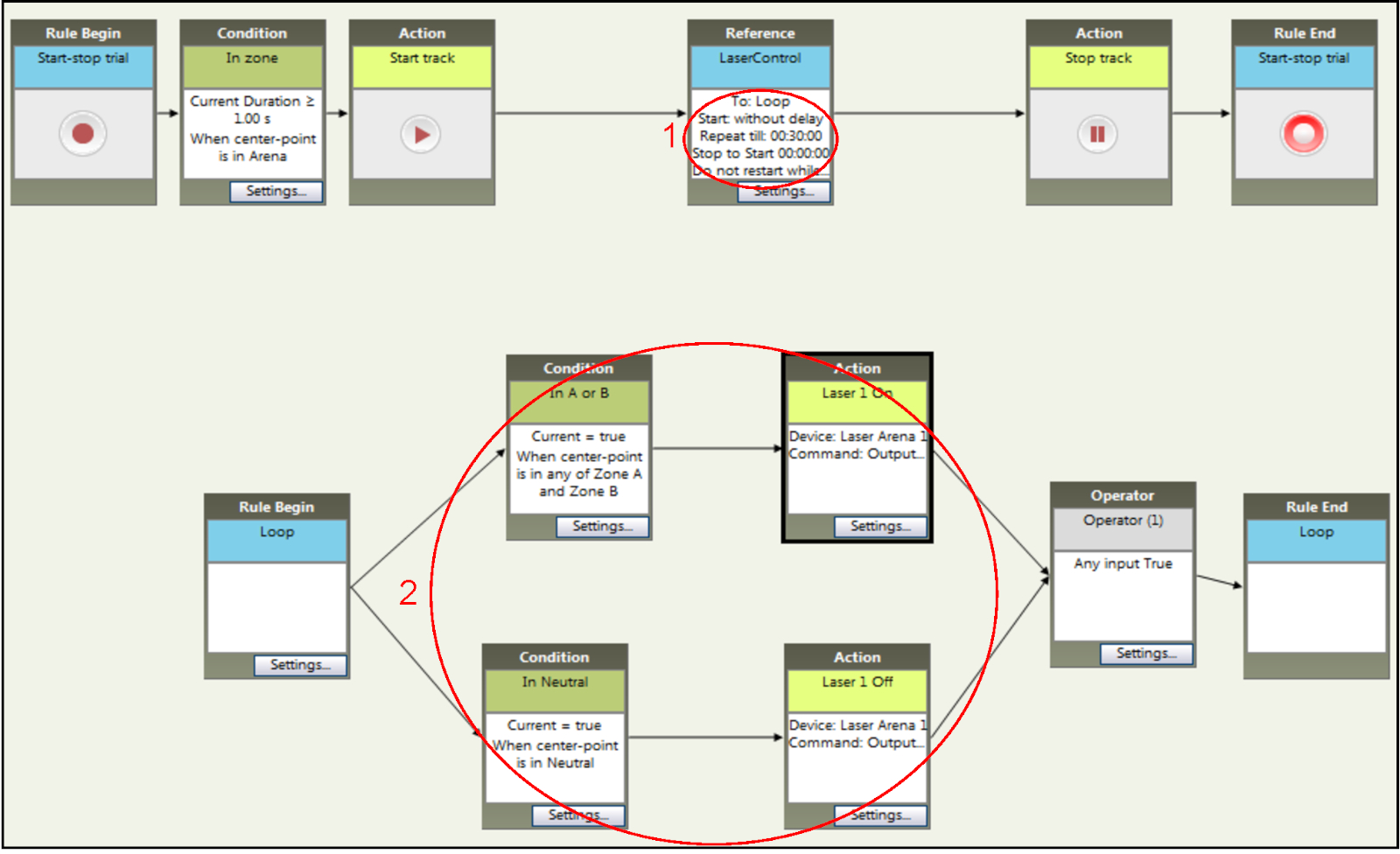
- 通过"参考"框设置中的"重复"选项将实验时间设置为 30 分钟(图 6中的步骤 1)。
- 通过添加与 A 和 B 隔间设置相关的条件框的"当中心点位于区域 A 和区域 B 中的任何位置",将 A 和 B 区域指定为激光配对(图 6中的步骤 2)。请注意,当动物位于中性隔间时,激光将关闭。
6. 使用激光刺激进行实验
- 设置检测设置。
- 使用虚拟人与鼠标类似,以确保适当的检测设置。
- 将虚拟设备放在设备的一个隔间中,并使用带动态减法的自动设置。
- 拆下假人并将其放置在相反的隔间。确保完全检测到虚拟人,如果没有,请通过软件调整设置以实现正确的检测。
- 在此步骤中,还要检查刺激是否按预期工作。使用先前配置的试用控制设置开始采集,并将虚拟人放在激光配对隔间中,并查看是否按照应有的情况触发了刺激。然后,将假人放在未配对和/或中性隔间中,看看刺激是否停止。
- 使用带有传感器的功率计,使用激光上的旋钮将激光功率设置为 10 mW(图 3B)。每次使用激光刺激时,执行此步骤。
注意:使用防护眼设备,因为直接接触激光会导致眼睛永久性损伤。 - 将鼠标放入设备中。
- 轻轻地将鼠标从笼子中拿出来,并使用陶瓷套连接光纤植入物到光纤配线。
- 将鼠标轻轻放在三节装置的中性隔间中。
- 等待,直到软件检测到鼠标。
- 拆下垂直滑动门,限制动物进入主隔间。
- 允许动物自由探索,没有任何干扰。
注:当动物未接受刺激时,遵循相同的程序,但不需要步骤 6.2 且激光一直关闭的情况除外。
结果
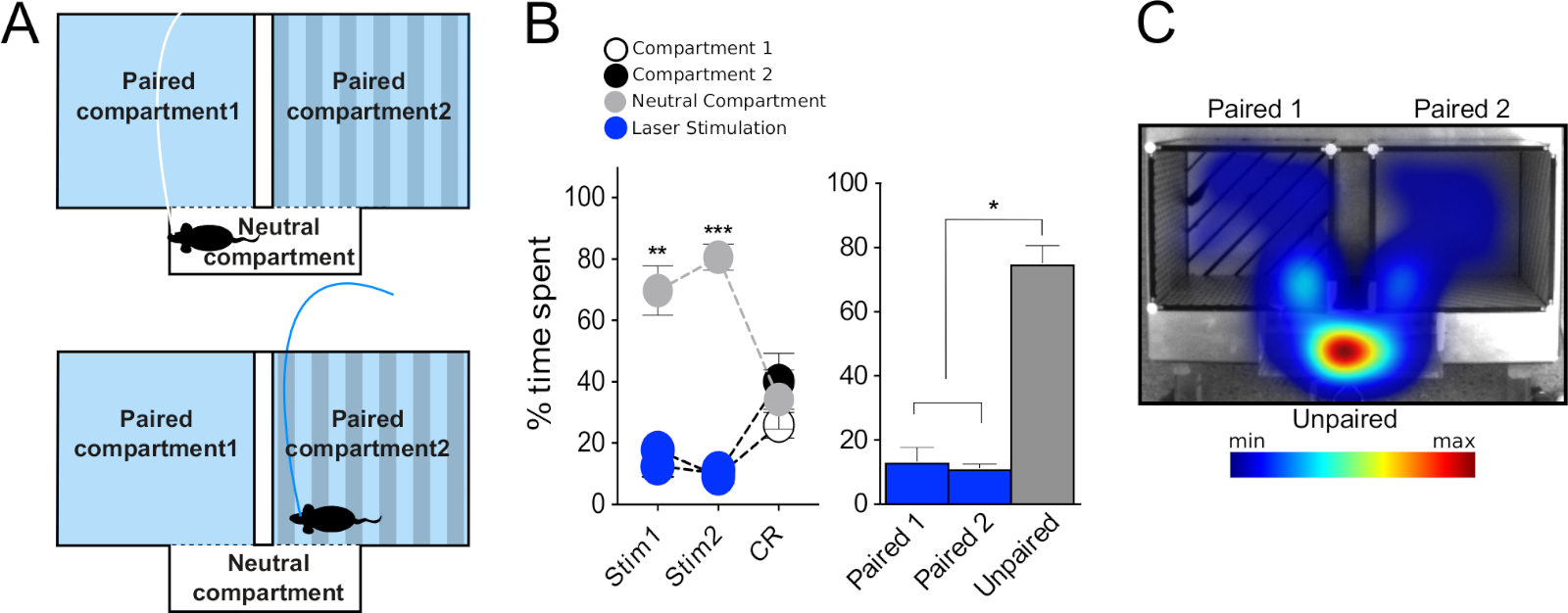
三节装置(图3F)适用于治疗药物的有益效应,并实时评估使用光遗传学直接刺激神经元的有益或厌恶特性。它由两个主隔间(20 厘米 [W] x 18 厘米 [L] x 25 厘米 [高]) 和一个较小的连接隔间 (20 厘米 [W] x 7 厘米 [L] x 25 厘米 [H])。主隔间具有独特的墙壁和地板纹理和图案,以方便关联学习,而连接/中性隔间是狭窄和透明的,因此小鼠自然花更少的时间在它。如上所述,跟踪软件可用于记录小鼠的多个行为参数,包括运动和在每个隔间中花费的时间,并控制激光刺激。整个RT-PP实验在8个会话中进行(图1A),允许评估直接刺激(第3天、第4天、第6天和第7天)的有益或厌恶特性,并形成正或负的关联,以回应以前的经验(第5天和第8天,"CR")。
首先,我们测试了在VTA中注射AAV-ChR2-eYFP病毒的DAT-Cre小鼠,以靶向多巴胺能神经元。根据文献, 我们观察到,小鼠更喜欢在隔间中花时间与刺激配对(图1B,阶段1,第3和第4天,蓝色圆圈,双向重复测量[RM]方差[方差]分析),隔间F(2,18)的影响= 141,p <0.001;会话X隔间F(12,108)的影响=42.1,p<0.01; Tukey 的专位后测试配对与未配对的 p < 0.001)。在反转阶段,将隔间与激光刺激的配对切换,证实了这些观察结果(图1B,第6天和第7天,蓝色圆圈,图基的后点置测试配对与未配对的隔间p <0.001),从而排除了从第1阶段获得的结果与侧偏置或随机参数有关的可能性。在RT-PP的四天中,平均每个隔间所花费的时间证实,与未配对的小鼠相比,它们平均花费70%的时间在激光配对隔间中(+20%)和中立 (+10%)隔间(图 1B条形图、单向 RM 方差分析图、刺激 F(2,6)效果 = 139,p < 0.001,Tukey 的帖子如何配对与未配对和中性隔间p < 0.001)。 此外,在没有刺激的情况下,在第5天和第8天,小鼠在先前与激光刺激配对的隔间中花费的时间明显更多(Tukey的后点p<0.001),表明以前的经验足以诱导反映为"寻求"刺激的关联学习行为。 这些数据与文献一致,证明目前的方法可以可靠地用于研究VTA特定神经元群体的光遗传学刺激的有益作用。
然后,我们测试了在VTA中注射AAV-ChR2-eYFP的VGLUT2-Cre小鼠,以靶向VTA的谷氨酸神经元。在这个实验中,我们观察到了与DAT-Cre小鼠所展示的行为表型相反的行为表型。因此,小鼠避免与刺激配对的隔间,并在所有 RT-PP 日内花费更多的时间在未配对的天数(图 1C左,双向 RM 方差分析,隔间 F(2,12)的影响 = 40.9,p < 0.001;会话 x 隔间 F(12,72)的影响 = 16.1,p & 0.001; Tukey 的专时测试配对与未配对p < 0.001;图 1C右侧,单向 RM ANOVA 效应刺激 F(2,6) = 162,p < 0.001,图基的后点对与未配对和中性隔间p < 0.001)。 有趣的是,在"CR"第5天和第8天,小鼠没有表现出对先前配对的隔间的明确回避(配对和未配对的隔间之间没有区别)。缺乏有条件的反应可能是由于激光配对隔间所花费的时间不足,从而阻止了激光活化与发生的特定环境之间形成关联。为了进一步探索这种回避表型,我们使用了一个修改后的协议,我们将其命名为"中性隔间首选项",缩写为 NCP。在本实验中,两个主隔间都与刺激配对,中性隔间保持无刺激(图7A)。我们假设,如果刺激具有厌恶性,那么老鼠将被迫花时间在更小的中性隔间,以避免它。事实上,在刺激的两天(Stim1和Stim2)中,老鼠大部分时间都在中性隔间里度过(约80%)与配对隔间相比(图7B,C;左侧:隔间F(2,8)的双向RM ANOVA效应= 70.9,p < 0.001; 会话 x 隔间 F(4,16)的影响 = 6.9,p = 0.002,Tukey 的后档"刺激 1"中性隔间与隔间 1 和 2 p < "刺激 2"中性隔间与隔间 1 和 2 p < 0.001;右: 单向 RM 方差,刺激 F(2, 2) = 54.2, p = 54.2 0.018,Tukey 的专位测试配对 1 和 2 与中性p < 0.05)。与RT-PP测试期间的"CR"天一样,小鼠似乎没有在隔间和刺激之间形成负结;也就是说,在没有刺激(CR)的情况下,他们对所有隔间进行了相同的探索(图7B,在成对隔间和中性隔间中所花费的时间没有差别)。这些实验的结果证实了在RT-PP设置过程中观察到的行为表型,从而支持RT-PP和NCP范式的组合实现。

图1:在RT-PP范式中使用光遗传学的行为测试。(A) 实验时间表的原理表示。(B,C)左上:图表,表示在整个RT-PP实验中为DAT-Cre(N = 10)和VGLUT2-Cre(N = 7)注射AAV-ChR2-eYFP的小鼠在每个隔间中所花费的时间百分比。蓝色圆圈:激光配对隔间;白色、黑色圆圈:主隔间;灰色圆圈:中性隔间。右上角:第 3、4、6 和 7 天(RT-PP)中每个隔间的平均时间百分比。底部:DAT-Cre 和 VGLUT2-Cre 鼠标在每个隔间中花费的时间的代表性热图。所有数据均以正态分布(夏皮罗-威尔克测试)。结果以均值 = SEM=p < 0.001 成对与未配对显示;=p < 0.05, =p < 0.01, =p < 0.001 配对与中性隔间;[p < 0.01, ]p < 0.001 未配对与中性隔间。这个数字已被修改从Bimpisidis等人12。请点击此处查看此图的较大版本。
{kind=link}

图2:光遗传学实验的外科手术。(A) 在VTA中注射与克里相关的病毒载体.(B) 将光纤植入注射部位上方。注意用于稳定的锚固螺钉。(C) 使用牙科水泥将纤维永久固定在头骨上。请点击此处查看此图的较大版本。
{kind=link}

图3:光遗传学实验中使用的设备。(A) 研究中使用的 TTL 框.它接收来自跟踪软件的输入,并将 TTL 信号发送到微控制器板。(B) 用于实验的激光源的前(上)和后视(底部)。(C) 用于控制激光刺激的微控制器板.请注意从 TTL 盒到激光源的连接。(D) 旋转接头.(E) 实验中使用的光纤片带。(F) 用于RT-PP和NCP实验的三节式仪器。请点击此处查看此图的较大版本。
{kind=link}

图 4:在跟踪软件中设计竞技场和区域。第 1 步:设置校准。第2步:绘制整个竞技场。第 3 步:在竞技场内绘制区域。第 4 步:安装验证。步骤 5:用于设置时间和刺激参数的"试用控制设置"选项卡。请点击此处查看此图的较大版本。
{kind=link}

图5:在跟踪软件中设置RT-PP实验的时间和刺激参数。为持续时间(步骤 1)和光刺激条件(步骤 2)添加特定规则。条件可轻松更改,以适应反转阶段的要求。请点击此处查看此图的较大版本。
{kind=link}

图6:在跟踪软件中设置NCP实验的时间和刺激参数。刺激疗程的持续时间(步骤 1)与 RT-PP 的持续时间类似,但轻度刺激激活的条件(步骤 2)不同。进入任一主隔间(此处命名为区域 A 和区域 B)会导致光遗传学刺激,只有当鼠标进入中性隔间时才会终止。请点击此处查看此图的较大版本。
{kind=link}

图7:在NCP范式中使用光遗传学的行为测试。(A) 实验设置的原理表示。(B) 左图:图表,表示在两天刺激(Stim1 和 Stim 2)期间和在 VTA (N = 5) 中注射AAV-ChR2的 VGLUT2-Cre 小鼠在条件响应 (CR) 会话期间在每个隔间中花费的时间百分比。右图:在 NCP 刺激的两天内,每个隔间的平均时间百分比。(C) 在刺激日的一天中,VGLUT2-Cre小鼠在每个隔间中花费的时间的代表性热图。数据是正态分布的(夏皮罗-威尔克测试)。结果以平均值 = SEM=p < 0.05, =p < 0.01, =p < 0.001 未配对与配对 1 和配对 2 个隔间。这个数字已被修改从Bimpisidis等人12。请点击此处查看此图的较大版本。
{kind=link}
| 步 | 温度 | 时间 | 周期 |
| 1. 初始变性 | 95 °C | 4分钟 | 1 |
| 2. 变性 | 95 °C | 30 s | 30 |
| 3. 退火 | 55 °C | 30 s | |
| 4. 扩展 | 72 °C | 40 s | |
| 5. 最终延期 | 72 °C | 6分钟 | 1 |
| 6. 保持 | 4 °C | 直到被实验者拦住 | 1 |
表1:PCR循环程序。
补充编码文件。请点击此处查看此文件(右键单击下载)。
讨论
在目前的研究中,我们提出了两个分步协议,即如何使用小鼠的光遗传学执行不同类型的位置偏好分析。所概述的协议用于评估VTA神经元的有益或厌恶行为表型(图1和图6)12,但也可用于探索神经元在其他大脑区域的行为作用。
最近的几项研究描述了RT-PP模式在两节23,24和三节仪器13,14,15,16,17,18。目前的协议描述了RT-PP和NCP协议在三节式仪器中的详细设置,类似于在CPP实验中传统上用于评估滥用药物管理行为影响的设备。虽然结果仅以鼠标在每个隔间中花费的时间百分比的形式显示,但跟踪软件确实允许分析其他几个行为参数,例如转换为区域、速度、移动时间等。分析不同的参数对解释数据非常重要。
目前的RT-PP协议是灵活的,可以修改,以测试不同类型的刺激模式是否有有益的效果。激光控制的参数可以通过微控制器板的脚本或跟踪软件轻松更改,从而演示了设置的多功能性。我们建议在以前研究中应用频率范围(有时较低)的20 Hz刺激频率,使用同一蛋白酶变体(ChR2/H134R)来研究多巴胺能和谷氨酸神经元及其终端13、14、16、17、18、23、24、25、26、27。最近的研究表明,较高的刺激频率对行为的影响可能大于低刺激频率,并且这些效应通过由较高频率28引起的去极化块进行调节。同样,在侧视区15中刺激谷氨酸神经元和GABAergic神经元时,行为输出的差异也表现出差异。这些研究检查了与VTA不同区域的神经元,对非谷胱甘肽神经元的高频观察到了最大的影响15,28。我们对20 Hz的选择是基于以前对谷胱甘肽和多巴胺能VTA神经元的研究,表明通过不同的刺激频率,与奖励相关的行为输出没有显著改变24,26。
可以调整并可能影响实验结果的另一个参数是光源的功率。较高的激光功率可以增加光刺激区域的大小,这在某些类型的实验中可能是有益的,但温度增加5的缺点。事实上,最近的一项研究表明,激光引起的温度升高可以改变大脑生理,影响行为测量29。这些观察强调了在实验设计中纳入蛋白酶负控制的重要性。在目前的协议中,我们使用的激光功率是10 mW的,这是类似的,以前已被证明是有效的刺激多巴胺能和谷氨酸神经元在VTA16,24,26。在设置实验时,请务必注意感兴趣的细胞所在的区域的大小以及光纤和配线属性(数值孔径、芯直径)。在执行与激光功率相关的计算时,这些参数是必须考虑的。详细资料,卡尔·戴塞罗斯实验室开发的计算器(http://web.stanford.edu/group/dlab/cgi-bin/graph/chart.php)可以使用。
在应用光遗传学实验时,对Cre-Lox重组进行组织学验证是另一个关键方面。重组效率的验证应始终在试验组进行,然后开始对一大群动物进行任何行为实验。这因伦理原因很重要,但对于优化的实验输出也很重要。每种病毒结构可能显示不同神经元类型和不同区域5的可变特异性,这个参数可以以不可预测甚至误导性的方式影响实验。例如,我们之前已经验证了 DAT-Cre 小鼠 VTA 中AAV5病毒的 Cre-Lox 重组模式,并发现单侧注射足以针对大多数感兴趣领域。然后,当我们研究VTA内的空间受限亚群,如NeuroD6表达特征的亚群时,我们观察到双边病毒注射更有效,能够靶向更多的神经元,对光遗传学光刺激产生更明显的行为影响12。此外,从手术到开始行为实验的时间必须谨慎处理。两周的时间足以让ChR2DNA结构在细胞体中表达,正如我们在这里展示的那样,但如果研究者在投影区13、14、15、17中测试刺激的效果,则可能需要更长的等待时间(±8周)。
值得注意的是,注射的病毒量(在我们的例子中为300 nL)可能适合在VTA中研究神经元时,但体积和度定必须根据转导效率和所研究的结构大小进行调整。此外,对于位于中微线轴远处的双边结构,可能需要进行双边注射,并双边植入光纤,以确保两个半球的激活/抑制。
最后,始终需要执行死后组织分析,以验证和确认 Cre-Lox 重组的效率,并验证光纤在预定位置的正确植入位点。意外、过度受限或过度的 Cre-Lox 重组可能发生,因为在预定区域边界外表达 Cre 的神经元分布未知,或者由于病毒血清型的差异、病毒处理不当、阻塞用于病毒输送或其他手术相关问题的注射器。为了得出安全的结论,必须验证令人满意的Cre-Lox重组和正确的光纤植入,以确认行为评估的任何统计结果。
根据这里提供的数据作为如何使用两种行为范式的例子,根据先前的发现23、24、25、26、27,在VTA中通过分析DAT-Cre小鼠,对VTA中多巴胺能神经元的光遗传学刺激获得的光对侧,基于先前的发现23、24、25、26、27,而VGLUT2-Cre小鼠所显示的这一侧是预料不到的。VTA的VGLUT2神经元及其预测已被证明参与奖励和厌恶16,17,24,30,31,因此,我们进行了NCP分析,以评估在当前RT-PP设置中观察到的明显回避行为更详细。通过使用狭窄的透明走廊作为唯一的非光成对隔间来确认VTA谷氨酸神经元刺激的厌恶特性,很明显,在这个特定的三腔设置中,这些神经元的光遗传学激活会引起厌恶反应。这些实验在这里展示,以举例说明可能受益于使用RT-PP和NCP协议的情况,是最近发表的研究的一部分,完整的数据集以及有关这些发现的讨论可以在本出版物12中找到。
除了NCP之外,确认厌恶的替代方法包括将竞技场的其余部分配对到激光激活时对开放场区域内的强照明,或者执行主动回避任务,即鼠标必须执行特定的行为模式才能终止激光刺激15。
总之,所述协议提供了如何以最有效的方式成功执行RT-PP和NCP分析的关键信息,以解开神经元激活在奖励和厌恶中的作用。根据科学假设,可以使用这些协议分析一系列参数,并且每个协议还可以与其他经过验证的范例相结合,以优化行为分析,实现光遗传学,以解决特定的大脑感兴趣的区域和神经元。
披露声明
作者没有什么可透露的。
致谢
我们的资金来源是感激的承认:厄普萨拉大学,维滕斯卡普斯卡斯雷德(瑞典研究理事会),Hjörnfonden,帕金森丰登,贝尔蒂尔·赫尔斯滕研究基金会,OE&EdlaJohansson,ZoologiskForskning和@hlén。动物被保存在Uppsala大学,并在Uppsala大学行为设施进行实验。
材料
| Name | Company | Catalog Number | Comments |
| AAV-Cre dependent virus | UNC Vector Core | - | a great variety of viruses to suit any project's needs |
| Agarose | VWR Life Science | 443666A | |
| Buffer for PCR | KAPA BIOSYSTEMS | KB1003 | 10x, contains 1.5mM MgCl2 at 1x |
| Bupivacaine (Marcain) | Aspen | N01BB01 | local anesthetic, 5 mg/ml solution, requires prescription |
| Carprofen (Norocarp) | N-Vet | 27636 | anti-inflammatory, analgesic; 50 mg/ml solution, requires prescription |
| dNTP set | Thermo Fisher Scientific | R0181 | 100mM, have to be dilluted to 10mM and mixed |
| DNA ladder | Thermo Fisher Scientific | SM0243 | 100 bp, 50 μg Gene Ruler |
| DNA loading dye | Thermo Fisher Scientific | R0611 | 6x, dilute to 1x before using |
| Ear puncher | AgnThos | AT7000 | ear puncher to take tissue samples for PCR or to mark animals |
| Fiberoptic patchcords | Doric Lenses | MFP_200/240/900-0.22_1m_FC-ZF1.25 | |
| Implantable fiberoptics | Doric Lenses | MFC_200/245-0.37_5mm_ZF1.25_FLT | the properties of the fibers might change depending on the experiment |
| Infusion pump for virus injections | AgnThos | Legato 130 | contains remote pump to secure the syringe directly on the stereotexi frame |
| Isoflurane (Forane) | Baxter | N01AB06 | Volatile anesthetic, requires prescription |
| Jewelry screws | AgnThos | MCS1x2 | |
| Laser source | Marwell Laser Systems | CNI MBL-III-473-100mW | |
| Microcontroller board | Arduino | "Uno" board | can be ordered from several companies |
| Microdrill | AgnThos | 1474 | could be ordered with or without stereotaxic holder |
| Needle (34G) | World Precision Instruments | NF36BV | |
| Nucleic Acid gel stain - GelRed | Biotium | 41003-T | |
| PCR tubes | Axygen | PCR-0208-CP-C | |
| Power meter | Thorlabs | PM100D | |
| Place Preference Apparatus | Panlab | 76-0278 | |
| Rotary joint | Doric Lenses | FRJ_1x1_FC-FC | |
| Sleeves | Doric Lenses | SLEEVE_ZR_1-25 | mating sleeve to connect the patchcord with the implanted optic fiber |
| Stabilization cement | Ivoclar Vivadent | Tetric EvoFlow | |
| Syringe (10ul) | World Precision Instruments | NanoFil | |
| Taq polymerase | KAPA BIOSYSTEMS | KE1000 | 500U |
| TAE buffer | Omega BIO-TEK | SKU:AC10089 | 50x concentration, has to be dilluted before use |
| Thermal cycler | BIO-RAD S1000 | 1852148 | necessary to perfrom PCR reactions |
| Tissue glue | AgnThos | Vetbond | |
| Tracking software | Noldus | Ethovision XT | |
| TTL box | Noldus | Noldus USB-IO box | |
| UV transilluminator | Azure Biosystems | c200 model |
参考文献
- Fenno, L., Yizhar, O., Deisseroth, K. The Development and Application of Optogenetics. Annual Review of Neuroscience. 34 (1), 389-412 (2011).
- Kim, C. K., Adhikari, A., Deisseroth, K. Integration of optogenetics with complementary methodologies in systems neuroscience. Nature Reviews Neuroscience. 18 (4), 222-235 (2017).
- Deisseroth, K. Optogenetics: 10 years of microbial opsins in neuroscience. Nature Neuroscience. 18 (9), 1213-1225 (2015).
- Nagy, A. Cre recombinase: The universal reagent for genome tailoring. Genesis. 26 (2), 99-109 (2000).
- Yizhar, O., Fenno, L. E., Davidson, T. J., Mogri, M., Deisseroth, K. Optogenetics in Neural Systems. Neuron. 71 (1), 9-34 (2011).
- Pupe, S., Wallén-Mackenzie, &. #. 1. 9. 7. ;. Cre-driven optogenetics in the heterogeneous genetic panorama of the VTA. Trends in Neurosciences. 38 (6), 375-386 (2015).
- Spanagel, R. Animal models of addiction. Dialogues in Clinical Neuroscience. 19 (3), 247-258 (2017).
- Tzschentke, T. M. Measuring reward with the conditioned place preference (CPP) paradigm: Update of the last decade. Addiction Biology. 12 (3-4), 227 (2007).
- Hoffman, D. C. The use of place conditioning in studying the neuropharmacology of drug reinforcement. Brain Research Bulletin. 23 (4-5), 373-387 (1989).
- Huston, J. P., Silva, M. A. D. S., Topic, B., Müller, C. P. What's conditioned in conditioned place preference?. Trends in Pharmacological Sciences. 34 (3), 162-166 (2013).
- Bardo, M. T., Bevins, R. A. Conditioned place preference: What does it add to our preclinical understanding of drug reward?. Psychopharmacology. 153 (1), 31-43 (2000).
- Bimpisidis, Z., et al. The NeuroD6 subtype of VTA neurons contributes to psychostimulant sensitization and behavioral reinforcement. eNeuro. 6 (3), e0066 (2019).
- Root, D. H., Mejias-Aponte, C. A., Qi, J., Morales, M. Role of Glutamatergic Projections from Ventral Tegmental Area to Lateral Habenula in Aversive Conditioning. Journal of Neuroscience. 34 (42), 13906-13910 (2014).
- Steidl, S., Wang, H., Ordonez, M., Zhang, S., Morales, M. Optogenetic excitation in the ventral tegmental area of glutamatergic or cholinergic inputs from the laterodorsal tegmental area drives reward. European Journal of Neuroscience. 45 (4), 559-571 (2017).
- Barker, D. J., et al. Lateral Preoptic Control of the Lateral Habenula through Convergent Glutamate and GABA Transmission. Cell Reports. 21 (7), 1757-1769 (2017).
- Wang, H. L., Qi, J., Zhang, S., Wang, H., Morales, M. Rewarding Effects of Optical Stimulation of Ventral Tegmental Area Glutamatergic Neurons. The Journal of Neuroscience. 35 (48), 15948-15954 (2015).
- Qi, J., et al. VTA glutamatergic inputs to nucleus accumbens drive aversion by acting on GABAergic interneurons. Nature Neuroscience. 19, 725-733 (2016).
- Qi, J., et al. A glutamatergic reward input from the dorsal raphe to ventral tegmental area dopamine neurons. Nature Communications. 5, 5390 (2014).
- Ekstrand, M. I., et al. Progressive parkinsonism in mice with respiratory-chain-deficient dopamine neurons. Proceedings of the National Academy of Sciences of the United States of America. 104 (4), 1325-1330 (2007).
- Borgius, L., Restrepo, C. E., Leao, R. N., Saleh, N., Kiehn, O. A transgenic mouse line for molecular genetic analysis of excitatory glutamatergic neurons. Molecular and Cellular Neuroscience. 45 (3), 245-257 (2010).
- Papathanou, M., et al. Targeting VGLUT2 in Mature Dopamine Neurons Decreases Mesoaccumbal Glutamatergic Transmission and Identifies a Role for Glutamate Co-release in Synaptic Plasticity by Increasing Baseline AMPA/NMDA Ratio. Frontiers in Neural Circuits. 12, 64 (2018).
- Franklin, K. B. J., Paxinos, G. . The mouse brain in stereotaxic coordinates. , (2008).
- Tsai, H. C., et al. Phasic firing in dopaminergic neurons is sufficient for behavioral conditioning. Science. 324 (5930), 1080-1084 (2009).
- Yoo, J. H., et al. Ventral tegmental area glutamate neurons co-release GABA and promote positive reinforcement. Nature Communications. 7, 1-13 (2016).
- Pascoli, V., Terrier, J., Hiver, A., Lüscher, C. Sufficiency of Mesolimbic Dopamine Neuron Stimulation for the Progression to Addiction. Neuron. 88 (5), 1054-1066 (2015).
- Ilango, A., Kesner, A. J., Broker, C. J., Wang, D. V., Ikemoto, S. Phasic excitation of ventral tegmental dopamine neurons potentiates the initiation of conditioned approach behavior: parametric and reinforcement-schedule analyses. Frontiers in Behavioral Neuroscience. 8, 155 (2014).
- Kim, K. M., et al. Optogenetic mimicry of the transient activation of dopamine neurons by natural reward is sufficient for operant reinforcement. PLoS ONE. 7 (4), 1-8 (2012).
- Kroeger, D., et al. Galanin neurons in the ventrolateral preoptic area promote sleep and heat loss in mice. Nature Communications. 9, 4129 (2018).
- Owen, S. F., Liu, M. H., Kreitzer, A. C. Thermal constraints on in vivo optogenetic manipulations. Nature Neuroscience. 22 (7), 1061-1065 (2019).
- Root, D. H., Estrin, D. J., Morales, M. Aversion or Salience Signaling by Ventral Tegmental Area Glutamate Neurons. iScience. 2, 51-62 (2018).
- Lammel, S., et al. Diversity of transgenic mouse models for selective targeting of midbrain dopamine neurons. Neuron. 85 (2), 429-438 (2015).
转载和许可
请求许可使用此 JoVE 文章的文本或图形
请求许可探索更多文章
This article has been published
Video Coming Soon
版权所属 © 2025 MyJoVE 公司版权所有,本公司不涉及任何医疗业务和医疗服务。