
Method Article
Contrôle des aflatoxines dans les arachides ARNi médiée: méthode pour analyser la production de mycotoxines et Transgene expression dans la Peanut /
Dans cet article
Résumé
Nous démontrons une méthode pour l'analyse des aflatoxines et l'expression du transgène dans les graines d'arachide qui contiennent des signaux ARN interférence pour réduire au silence des gènes de synthèse de l'aflatoxine dans le champignon Aspergillus flavus. Contrôle de l'ARNi médiée par des mycotoxines dans les plantes n'a pas été signalé précédemment.
Résumé
L'Organisation des Nations Unies pour l'alimentation et l'agriculture estime que 25% des cultures vivrières dans le monde sont contaminées par des aflatoxines. Cela représente 100 millions de tonnes de nourriture détruites ou détournées vers la consommation non-humaine chaque année. Les aflatoxines sont cancérigènes puissants normalement accumulées par les champignons Aspergillus flavus et A. parasiticus dans les céréales, les noix, les légumes racines et les autres produits agricoles. Silence de cinq gènes aflatoxine-synthèse par interférence ARN (ARNi) dans les usines d'arachide a été utilisé pour contrôler l'accumulation de l'aflatoxine après inoculation avec A. flavus. Auparavant, aucune méthode existait pour analyser l'efficacité de l'ARNi dans les événements transgéniques d'arachide individu, que ceux-ci produisent généralement peu de graines, et les méthodes traditionnelles de grandes expériences sur le terrain dans des conditions d'aflatoxine ont été propices pas une option. Sur le terrain, la probabilité de trouver des graines naturellement contaminés est souvent 1/100-1/1,000. En outre, la contamination par l'aflatoxine est pas uniformément répartis. Notre méthode utilise quelques graines par événement transgénique, avec de petits morceaux transformés pour PCR en temps réel (RT-PCR) ou petite séquençage de l'ARN, et pour l'analyse de l'accumulation de l'aflatoxine par chromatographie liquide ultra performance (UPLC). Lignes d'arachide ARNi exprimant 288-72 et 288-74, ont montré jusqu'à 100% de réduction (de p≤0.01) en aflatoxine B 1 et B 2 par rapport à la commande qui se sont accumulés jusqu'à 14.000 ng. G -1 de l'aflatoxine B 1 lorsque inoculé avec aflatoxigènes A. flavus. Comme référence, le total maximum d'aflatoxines autorisées pour la consommation humaine aux Etats-Unis est de 20 ng g -1.. Ce protocole décrit l'application du contrôle de l'ARNi médiée par des aflatoxines dans les graines et les méthodes pour l'évaluation des arachides transgéniques. Nous pensons que son application dans l'élevage d'arachide et autres cultures apportera progrès rapides dans ce domaine important de la science, De la médecine et de la nutrition humaine, et contribueront de manière significative à l'effort international visant à contrôler les aflatoxines, et potentiellement d'autres mycotoxines dans les principales cultures vivrières.
Introduction
Environ 4,5 milliards de personnes sont exposées de façon chronique aux aflatoxines 1, les substances cancérogènes les plus puissantes connues dans la nature 2. Ces mycotoxines contaminent 25% des cultures vivrières dans le monde 3, y compris le maïs, le manioc, le riz, les noix, les céréales et les épices. 4. Les aflatoxines entraînent des retards de croissance chez les enfants de 5, compromettent le système immunitaire 6, sont présents dans 58% des carcinomes hépatocellulaires-en biopsies humaines 7,8, et tuent des centaines de personnes au cours des flambées périodiques de aflatoxicose 9,10. Les aflatoxines sont des mycotoxines polykétide dérivé normalement produites par Aspergillus flavus et A. parasiticus; aflatoxines B 1 et B 2 sont produites par A. flavus, alors que A. parasiticus produit également des G 1 et G 2. La structure chimique de ces composés et un chromatogramme montrant leur séparation par UPLC sont présentées dans la figure 1.

Figure 1. Les aflatoxines et RNAi insérer Haut:. Structure chimique (à gauche) et l'exemple de chromatogramme (à droite) des quatre aflatoxines polyketides dérivés les plus courants: B 1, B 2, G 1 et G 2, produite par Aspergillus parasiticus, A . flavus produit B 1 et B 2 Bottom: Schéma de fragments de gènes dans l'ARNi construire p5XCAPD utilisé pour la transformation d'arachide, numéros sous les flèches sont les numéros d'accession de gènes fragment dans le génome d'Aspergillus flavus;. PIV2: la pomme de terre intron; paires de bases;: pb RT_5X_1 et RT_5X_2:. En temps réel des sites d'amorce PCR S'il vous plaît cliquer ici pour voir une version plus grande de cette figure.
{kind=link}
Les pertes économiques dans les exportations en raison de aflatoxines dans l'arachide dépassent seuls 450 millions de dollars américains si calculé sur la base de 4 ng. G -1 limite de l'aflatoxine autorisé pour la consommation humaine dans l'Union européenne 11. Les aflatoxines sont connus depuis 60 ans 12; Toutefois, si de nombreuses pratiques agricoles ont été développés pour en atténuer les effets, y compris l'application d'autres souches fongiques 13,14, aucune méthode uniforme de contrôle existe, et les variétés de plantes résistantes ne sont pas disponibles. Test matériel phytogénétique pour la résistance aux aflatoxines est particulièrement difficile, parce que même dans des conditions propices à l'invasion de l'agent pathogène, l'accumulation de mycotoxines est imprévisible et ne suit pas une distribution normale. Ainsi, les expériences nécessitent généralement de vastes zones de plantation, des centaines de graines et de multiples échantillons de 100-1,700 g pour réduire la variabilité de la 15,16 de données.
L'interférence ARN étaitdécouvert en 1998 17; et les avantages de "faire taire" sont actuellement explorées dans un certain nombre de nouvelles applications, par exemple., dans les thérapies humaines contre le cancer du sein métastatique 18, le cancer du foie 19, la leucémie myéloïde 20, et dans la protection des plantes contre les insectes 21 et nématodes 22. Dans les usines, les signaux d'interférence d'ARN peuvent voyager cellule en cellule, avec des petits ARN interférents (siRNA) et de l'ARN de haut poids moléculaire étant responsable pour le gène posttranscriptional systémique taire 23,24, même à l'intérieur des agents pathogènes fongiques qui sont en contact étroit avec la plante hôte 25. L'efficacité de l'ARNi sur le silençage des plantes médiée par des gènes fongiques-pathogène a été décrite dans quelques pathosystèmes végétales, pour ces derniers, l'examen visuel des symptômes dans les parties aériennes des plantes (feuilles) a permis la quantification de la maladie, à savoir, oomycète Bremia dans la laitue 26 , Puccinia chez le blé 27 et Fusarium dans la banane 28. Beaucoup plus difficile est d'évaluer l'efficacité ARNi pour contrôler les mycotoxines dans les plantes, en particulier les aflatoxines dans les arachides que les feuilles ne présentent aucun symptôme d'infection, les organes envahis (graines) sont sous plusieurs pouces du sol, l'apparition de l'infection est imprévisible, et seul produit chimique l'analyse permet de déterminer la présence d'aflatoxines. En outre, chaque événement transgénique arachide produit normalement quelques graines (4-6 par plante); Par conséquent, le test traditionnel pour une accumulation sans trait d'aflatoxine dans les grandes parcelles de terrain, d'une durée saisons de culture entières, et en utilisant des centaines de graines est pas possible. Une méthode est décrite ici pour analyser en moins d'une semaine, l'ARNi graines d'arachide pour présence de transgène et pour une accumulation sans trait d'aflatoxine, en utilisant seulement quelques graines.
Protocole
1. construction moléculaire et de transformation d'arachide
- Combiner des fragments d'ADN de cinq A. gènes Flavus AFL2G_07223 (AFLS ou aflJ), AFL2G_07224 (aflR), AFL2G_07228 (CATF / pksA / pksL1), AFL2G_07731 (pes1) et AFL2G_05027 (aflatoxine pompe d'efflux, aflep). Pour cela, utilisez les amorces et ultramers suivantes: DIR-1, à court Dir1-R, DIR-2-inversé, à court Dir2-R, DirAll-NCO-Rv et DirAll-Bameco-Fw, tableau 1.
- Assurez-DIR-1 à double brin par 5 cycles de PCR (réaction de 25 ul, 95 ° C 2 min, suivie de 5 cycles de 94 ° C 45 sec, 55 ° C 30 sec, 68 ° C 15 sec) en utilisant l'ADN polymérase selon fabricant instructions et amorce court Dir1-R pour laisser un CCCGT 3 'en saillie. Répétez ces étapes pour faciliter le double brin DIR-2-inversé en utilisant l'amorce court Dir2-R pour laisser un ACGGG 3 'en saillie complémentaire DIR-1.
- Ligaturer les deux fragme 199 pbnts avec l'ADN ligase T4 selon les instructions du fabricant. Amplifier par PCR le fragment 393 pb résultant comme indiqué dans 1.1.1 en utilisant des amorces DirAll-CACC-FW et DirAll-NCO-RV (tableau 1), et de cloner le produit en utilisant des techniques standard en pENTR1A faire plasmide p2 + 4ENTR.
- Recombiner P2 + 4ENTR dans pCAPD 29 (NCBI accession: KC176455.1) en utilisant LR clonase II enzyme mélange selon les instructions du fabricant pour faire plasmide p5XCAPD, et la transformer en Escherichia coli DH5a en utilisant des techniques standard suivie d'un séquençage partiel. Remarque: L'insert complet ARNi est indiquée dans le tableau 1.
- Transformer la souche d'Agrobacterium C58C1 30 avec le plasmide p5XCAPD comme indiqué précédemment 30, et d'utiliser la bactérie résultant pour transformer des plantes d'arachide comme suit:
- Croître à 30 ° C l'Agrobacterium p5XCAPD, utilisez pour cela, 50 ml de LB-Broth supplémenté avec 500pg ml -1 streptomycine, 25 pg ml -1 gentamicine, 10 pg ml -1 kanamycine, et secouer la culture à 250 rpm jusqu'à atteindre 1 260 OD.
- Récolter les cellules d'Agrobacterium par centrifugation (6000 xg) pendant 10 min, remettre en suspension dans 50 ml AB milieu minimal 31 100 acétosyringone uM pendant 1 heure, et le lieu de la suspension bactérienne des explants de 10-14 jour vieux plants Exp27-1516, coureur lignée d'arachide type. Sécher les explants sur 3MM papier buvard après 30 minutes, et de les placer sur un milieu shoot-induction (SIM) [MS sel 32, 3% de saccharose, 20 uM benzylaminopurine (BAP), 10 uM thidiazuron (TDZ), pH 5,8, 0,3 % gomme gellane] sans antibiotiques dans l'obscurité pendant trois jours.
- Avez-sélection de tissus et la régénération comme indiqué avant 33. Déplacez tissus sur la carte SIM (500 um céfotaxime et 100 uM kanamycine) pour la formation de tournage, avec des transferts bi-hebdomadaire pour 2 mois. Puis lieuexpansion pousses sur un milieu shoot-allongement (SEM) [5 uM BAP, l'acide gibbérellique 1 uM (GA 3)], bi-hebdomadaire pendant plusieurs mois.
- Placez pousses individuelles, 2 cm en taille, en milieu root-induction (RIM) [1/2 MS, 1,5% de saccharose, 5 uM α-naphtalène-acétique (NAA), 2,5 uM indole-butyrique (IBA)], puis acclimater les plants et de les transférer à l'effet de serre.
2. Identification des plantes d'arachide hébergeant ARNi Genes Silence aflatoxine Synthèse
- Utiliser plante mini kit dans un poste de travail de robot avec 200 élution ul selon les instructions du fabricant pour extraire l'ADN de jeunes feuilles de plants d'arachide qui ont été soumis à un processus de transformation (comme décrit précédemment) avec ARNi construire p5XCAPD (Figure 1) qui a pour épine dorsale plasmide pCAPD 29 pour le silençage génique.
- Tamiser les échantillons d'ADN par un seul tube PCR nichée (STN-PCR) comme décrit PREVIment 34 pour détecter le marqueur NPTII sélectionnable et l'ARNi insérer de p5XCAPD. Propager par clonage à partir de coupes (3-4 nœuds) PCR plantes positives à produire suffisamment de semences pour les essais dans cette première génération.
- Utilisez quatre dilutions de 2 fois de l'ADN (50-100 ng ul -1 avant dilution) dans toutes les réactions STN-PCR. Pour NPTII, en utilisant des amorces externes PCAPD 5714F: 5'-AGGCTATTCGGCTATGACTG-3 'et PCAPD 6446R: 5'-CGTCAAGAAGGCGATAGAAG-3', et les amorces internes PCAPD 5730F: 5'-ACTGGGCACAACAGACAATC-3 'et PCAPD 6249R: 5'-ATATTCGGCAAGCAGGCATC- 3 '.
- Pour la détection de l'ARNi insérer dans des réactions de PCR en utilisant STN-amorces externes 35S-PDSFw: 5'-CCTAACAGAACTCGCCGTAA-3 'et DirAll-Nco-RV: 5'-ATGCCATGGGGTTATTGGGTGCAGAATGG-3', et les amorces internes Probe_5027_Fw: 5'-3-gtatttgtgaccatgtttctg 'et Probe_7228_Rv: 5'-GGACGGATAGTAAACTGCGG-3'.
- Récolte arachide gousses de plantes positives STN-PCR et de la con plantes TROL cultivées dans les mêmes conditions. Étape critique: Veiller à ce que le contrôle plantes sont cultivées dans les mêmes conditions et la même saison que les plantes ARNi. Jet d'eau les gousses avec un nettoyeur haute pression à faible intensité ou gratter à la main pour enlever exocarpe, déterminer la couleur du mésocarpe en plaçant les gousses sur une planche de maturité et de gousses séparées en groupes (jaune, orange, marron et noir) 35 (Figure 2).

Figure 2. Préparation de gousses d'arachide pour l'analyse. Gauche: tailles d'arachide différents retrouvés à la récolte que l'arachide est une plante à croissance indéterminée; Centre: placer les arachides dans le panier métallique pour l'enlèvement de pression d'eau de exocarpe; droite: groupes de maturité par la couleur de mésocarpe sur une planche de profil d'arachide (jaune, orange, marron et noir).98fig2large.jpg "target =" _ blank "> S'il vous plaît cliquer ici pour voir une version plus grande de cette figure.
3. installation expérimentale
- Retirez la coque, processus jaune et graines brunes séparément. Calculer le nombre de graines à utiliser dans l'expérience selon le tableau 2. Notez qu'un minimum de trois morceaux de terre (pièce 1 de semences = la moitié cotylédons) par ligne d'arachide et la date d'échantillonnage est nécessaire pour effectuer une analyse statistique et de réduire l'erreur-type.
| prénom | Séquence | ||
| DIR-1 | 5'- GCCAGCTCAAAAGTGCGATGCACCAAGGAGAAACCGGCCTGTGCT CGGTGTATCGAACGTGGTCTTGCCTGTCAATACATGGTCGTATTTG TGACCATGTTTCTGGTGGCATTGGACCGTCTTGTCATCTCTACAGCC ATTCCCCAGATCACGGACGAATCCCCTGCATCTACGCGCACGCATC ACTTGGGGTACCCGT-3 ' | ||
| Dir1 court-R | 5'-Phos-TACCCCAAGTGATGCGTGCGCG-3 ' | ||
| DIR-2-inversé | 5'GGTTATTGGGTGCAGAATGGTAAACCACCCAACAGTACGCGAAATG TCAATTCCAGAGTCCCAAACCTCCCTACCGTGGCCTGGACGGATAG TAAACTGCGGAGCTTGGGAACAAAATCCGCTGTCTGATCGCCGAAG AGAAAGAGTTGCCTTGATTGAGCCGCATCGAGGACAGGTTGTGTTG CTGTTGATAGACGGG-3 ' | ||
| Dir2 court-R | 5'-Phos-CTATCAACAGCAACACAACC-3 ' | ||
| DirAll-CACC-Fw | 5'-CACCGCCAGCTCAAAAGTGCGATGC-3 ' | ||
| DirAll-NCO-Rv | ATGCCATGGGGTTATTGGGTGCAGAATGG 5'-3 ' | ||
| DirAll-Bameco Fw- | ATGGGATCCGAATTCGCCAGCTCAAAAGTGCGATGC 5'-3 ' | ||
| Insert ARNi complète | 5'GCCAGCTCAAAAGTGCGATGCACCAAGGAGAAACCGGCCTGTGCT CGGTGTATCGAACGTGGTCTTGCCTGTCAATACATGGTC / GTATTTG TGACCATGTTTCTGGTGGCATTGGACCGTCTTGTCATCTCTACAGCC ATTCCCCAGATCACGGACGAAT / CCCCTGCATCTACGCGCACGCAT CACTTGGGGTACCCGTCTATCAACAGCAACACAACCTGTCCTCGAT GCG / GCTCAATCAAGGCAACTCTTTCTCTTCGGCGATCAGACAGCG GATTTTGTTCCCAAGCTCCGCAGTTTACTATCCGTCCA / GGCCACGG TAGGGAGGTTTGGGACTCTGGAATTGACATTTCGCGTACTGTTGGG TGGTTTACCATTCTGCACCCAATAACC-3 ' | ||
. Tableau 1. Les oligonucléotides et ultramers utilisés pour construire l'ARNi construire p5XCAPD Phos: phosphorylés extrémité 5 '; "/" Sépare les cinq fragments de gènes utilisés; Complete insert d'ARNi: séquence utilisée comme 2 répétitions inversées pour former p5XCAPD.
- Placer graines d'arachide entiers en une seule couche de manière à couvrir le fond du bêcher stérile un. Ajouter 75% d'éthanol / eau (v / v) pour couvrir les graines, puis ajouter un volume égal de la même solution. Incuber à température ambiante pendant 30 secondes, puis rincer avec de l'eau déminéralisée stérile (SDW).
- Ajouter 2% d'hypochlorite dans le bécher contenant de l'éthanol semences traitées comme suit: ajouter suffisamment de solution d'hypochlorite de 2% pour couvrir les graines, puis ajouter un volume égal de la même solution et incuber pendant 5 min. Étape critique: rincer abondamment à trois fois avec un volume de SDW équivalente à 5 fois le volume d'hypochlorite utilisé (Figure 3).

Figure 3. Dispositif expérimental. Haut: la stérilisation de la surface des graines d'arachide avant et après l'hypochlorite, l'enlèvement de couches de terre (testa); Moyen: élimination de l'embryon et de voir de près l'embryon, puis couper cotylédons dans la moitié Bas: cotylédons et demi dans l'eau distillée stérile, buvard sur stérile absorbant papier, bosses sur la gélose de l'eau, et le placement des cotylédons et demi (de côté coupé vers le haut) sur la surface de la gélose. S'il vous plaît cliquer ici pour voir une version plus grande de cette figure.
{kind=link}
- Autoriser les graines stérilisées en surface à Imbibe immergés dans SDW pendant 2 heures. Placez les graines sur une boîte de Pétri stérile, enlever les téguments avec des pinces, séparer les cotylédons, et avec un scalpel supprimer les embryons. Note:Les embryons peuvent être jetés ou utilisés pour régénérer de nouvelles plantes.
- Couper chaque cotylédons de moitié l'aide d'un scalpel. Pour éviter la déshydratation, garder les morceaux de semences de coupe à l'eau stérile jusqu'à ce que toutes les graines sont traitées.
- Ont préparé des boîtes de Pétri contenant de la gélose de l'eau stérile (1,5% de gélose / eau; p / v), une pour chacun des trois morceaux de graines. Faire de petites entailles dans la gélose en utilisant une pince, efface brièvement l'excès d'eau de morceaux de semences sur des serviettes en papier stériles, puis placer les morceaux de semences (face coupée vers le haut) sur les plaques d'agar d'eau /.
- À partir d'une culture fraîche de aflatoxigènes Aspergillus flavus NRRL 3357 dans un milieu agar de Czapek augmenté à 25 ° C pendant 10 jours, préparer une suspension de 50.000 spores par ul SDW, comptés avec un hémocytomètre.
- Placez 2 pi de la suspension de spores sur les surfaces de chaque pièce demi-cotylédons éviter le ruissellement sur les côtés coupés, pour vous assurer que les spores se exposés au tissu de la graine qui abrite ARNi (Figure 4).

Figure 4. L'inoculation et d'incubation pour l'analyse des aflatoxines. Top: Demi cotylédons, inoculation avec une suspension de spores, et Aspergillus flavus croissance du mycélium sur la moitié cotylédons après 24 heures d'incubation bas: à gauche:. Incubation de 48 h sur 1,5% d'agar; centre: une incubation de 72 h sur 1,5% d'agar; droite:. exemple de montage expérimental incorrecte, une incubation de 72 heures sur gélose à 0,5% S'il vous plaît cliquer ici pour voir une version plus grande de cette figure.
{kind=link}
- Incuber les boîtes de Pétri contenant les cotylédons demi inoculés et non inoculés à 30 ° C dans l'obscurité jusqu'à ce que l'échantillonnage.
4. L'échantillonnage pour l'analyse des aflatoxines et Gene Expression
- Prélever des échantillons danstriple à 24, 48 et 72 h (96 h en option) d'incubation, à la fois pour la RT-PCR et pour l'analyse de l'aflatoxine, chaque répétition étant une pièce (la moitié cotylédons). Choisissez au hasard des échantillons de différentes plaques à chaque date d'échantillonnage. Utilisation d'un tissu, retirer doucement spores fongiques agar et l'excès avant de placer des morceaux de semences dans des flacons ou des tubes à essai.
- Pour l'analyse de l'aflatoxine, placer chaque répétition (une pièce) dans un 4 ml vis de verre bouchon du flacon et stocker à -80 ° C. Remarque: Compte tenu de la stabilité chimique des aflatoxines, les échantillons peuvent être stockés dans cet état pendant plusieurs mois (dans les expériences actuelles généralement 1-2 mois).
- Pour lieu RT-PCR chaque répétition (une seule pièce) en prépare déjà 2 ml de broyage des tubes contenant deux billes en acier inoxydable (2,5 mm de diam) et trois perles de zirconium (2 mm de diam).
- Geler immédiatement (de préférence dans de l'azote liquide) tous les échantillons et de les conserver à -80 ° C jusqu'à leur traitement.
5. analyse des aflatoxines de Half individuel Cotyledon Pieces
- Utilisez le A. flavus inoculé des échantillons pour cette analyse. Amener les échantillons à température ambiante pendant environ 30 min, ajouter quatre volumes (habituellement 2-3 ml; p / v) de méthanol (garder une trace pour les calculs ultérieurs), fermer les bouchons, et incuber O / N (~ 16 h) dans le obscurité sans agitation.
- Placer une fritte en une correspondant à 1,5 ml de propylène minicolonne, ajouter 200 mg de base de Al 2 O 3 et le capuchon avec un autre fritte comme décrit précédemment 36; ensuite, placer un Ultra-liquide à haute performance Chomatographer (UPLC) échantillonneur automatique flacon sous la colonne assez près pour éviter une éventuelle évaporation de l'éluat.
- Dans un tube à essai en verre jetable (ne pas utiliser de plastique), placer 0,5 ml de l'extrait de méthanol obtenu à l'étape 4.1, ajouter 0,5 ml d'acétonitrile, mélanger à la pipette et appliquer 0,5 ml du mélange dans la colonne préparée dans l'étape 4.2. Autoriser élution dans le flacon de l'échantillonneur automatique par gravité (ne pas appliquer de pression). Élution prend habituellement 2-4 min, à proximité èmee flacon immédiatement en utilisant un capuchon compatible UPLC avec cloisons. Conserver les flacons à température ambiante et les analyser le même jour sur un UPLC.
- Pour la séparation des aflatoxines, flacons lieu échantillonneur automatique contenant éluats d'échantillons et flacons échantillonneur automatique avec les normes de l'aflatoxine (B 1 et B 2 lors de l'utilisation flavus) dans un instrument UPLC équipé d'un appariement Gestionnaire UPLC solvant quaternaire, UPLC Sample Manager, UPLC Fluorescent détecteur, et un 2,1 mm x 50 mm 18 C, 1,7 um colonne.
- Utilisez une phase mobile isocratique composé d'eau / MeOH / CH 3 CN (64:23:13, v / v / v), au taux de 0,30 ml min -1. Obtenir des chromatogrammes assurant une séparation de base stabilisée pour le calcul précis de la concentration de l'aflatoxine, selon les instructions du fabricant 37.
- Confirmer l'identité des aflatoxines par l'obtention de leurs données spectrales de masse et de les comparer avec les données publiées 38. Utilisez un ile piège spectromètre de masse équipé d'une interface ESI et le logiciel correspondant selon les instructions du fabricant.
- Déterminer les concentrations d'aflatoxines par référence aux courbes d'étalonnage obtenues en injectant des quantités différentes de normes commerciales d'aflatoxines B 1, B 2 correspondant, G 1 et G 2 comme suggéré par le fabricant de l'UPLC et déterminée par le logiciel 37.
- Placez les morceaux de semences déjà extraites avec du méthanol, dans des flacons de verre individuelles pour O / N (~ 16 h) lyophilisation pour déterminer leur poids sec. Ensuite, calculer la concentration d'aflatoxine dans ng. G -1 de poids sec de plantons.
- Pour l'analyse des données, de convertir les résultats de l'aflatoxine pour vous connecter (ng. G -1 +1), suivie d'un test de Tukey pour les comparaisons moyennes.
6. Gene Expression, Traitement RT-PCR des échantillons
- Prenez les 2 ml de broyage tubes contiennenting échantillons du -80 ° C congélateur, et immédiatement (sans décongélation) les broyer dans un homogénéisateur de broyeur à billes à 3100 tours par minute pendant 40 secondes, puis procéder à l'extraction de l'ARN en utilisant Trizol selon les instructions du fabricant.
- Préparer l'ADNc à partir de chaque échantillon en utilisant une ARN pg et des quantités égales d'oligo dT et des hexamères aléatoires, faire une dilution 1: 8 de l'ADNc et en utilisant 2 pi par réaction à RT-PCR (comme décrit précédemment 39).
- Pour la détection de l'expression d'ARNi de l'utilisation des amorces insert: RT_5X_1_105F: 5'GGTGGCATTGGACCGTCTTG-3 ', RT_5X_1_232R: 5'-CGCATCGAGGACAGGTTGTG-3'; et RT_5X_2_95F: 5'-CCATGTTTCTGGTGGCATTG-3 ', RT_5X_2_229R: 5'-ATCGAGGACAGGTTGTGTTG-3'.
- Pour la détection de l'expression du marqueur sélectionnable NPTII, l'utilisation des amorces: RT_NPTII_1_6871F: 5'-CTCGCTCGATGCGATGTTTC-3 ', RT_NPTII_1_7004R: 5'-GCAGGATCTCCTGTCATCTC-3'. Utilisez le gène de ménage actine pour la normalisation, et les amorces: Actine-Fw: CACATGCCATCCTTCGATTG; Actine-Rv: CCAAGGCAACATATGCAAGCT 40.
- Analyser les résultats par Delta-delta méthode C T 41 normalisé pour l'expression d'actine. Représenter les résultats sous forme de multiplication sur le contrôle.
Résultats
Plasmide p5XCAPD a été fait comme un dérivé de pCAPD 29, et l'a utilisé pour transformer des plantes d'arachide; Ce vecteur porte répétitions inversées de cinq petits fragments, 70-80 pb chacun, de gènes de synthèse de l'aflatoxine-A. flavus séparés par un intron (Figure 1). Fragments de AFL2G_07224 (aflR), AFL2G_07223 (AFLS ou aflJ), AFL2G_05027 (aflatoxine pompe d'efflux, aflep), AFL2G_07228 (CATF / pksA / pksL1), et AFL2G_07731 (pes1) ont été utilisés pour la construction, les numéros sur la figure 1 correspondent à une . flavus annotation du génome au Broad Institute, Cambridge, MA, et la littérature 42. Un total de 99 lignes d'arachide ont été régénérés après être passé par le processus de transformation, 50 étaient positifs par PCR pour NPTII détecté par STN-PCR, et 33 lignes étaient PCR positives et les graines produites. Seulement sept lignes positifs PCR ont été propagées par clonage et testés par le présenméthode de t pour l'accumulation de l'aflatoxine, tous les sept montré entre 60% et 100% moins d'accumulation de l'aflatoxine que le contrôle. Ici, nous montrons les résultats de deux de ces sept lignes. Comme les événements transgéniques individuels produisent généralement peu de graines, une méthode a été développée pour utiliser un nombre minimum de graines tout en étant capable de faire l'analyse statistique paramétrique. Un organigramme de la préparation de l'échantillon et montage expérimental est représenté sur la Figure 5 et le Tableau 1. Bien que la première génération de graines transgéniques est généralement hémizygotes, il est prévu que la cellule à cellule et la circulation systémique de petits ARN interférents (siRNA) générés par interférence ARN devrait conférer aflatoxine synthèse silence dans toute la plante.

Figure 5. organigramme schématique de la méthode pour analyser l'efficacité de l'ARNi à faire taire Aspergillus gènes aflatoxine-synthèse dans les graines d'arachide. Représentation graphique du flux de travail lors du traitement des échantillons d'arachide pour l'expression du gène ou de l'analyse de l'aflatoxine. S'il vous plaît cliquer ici pour voir une version plus grande de cette figure.
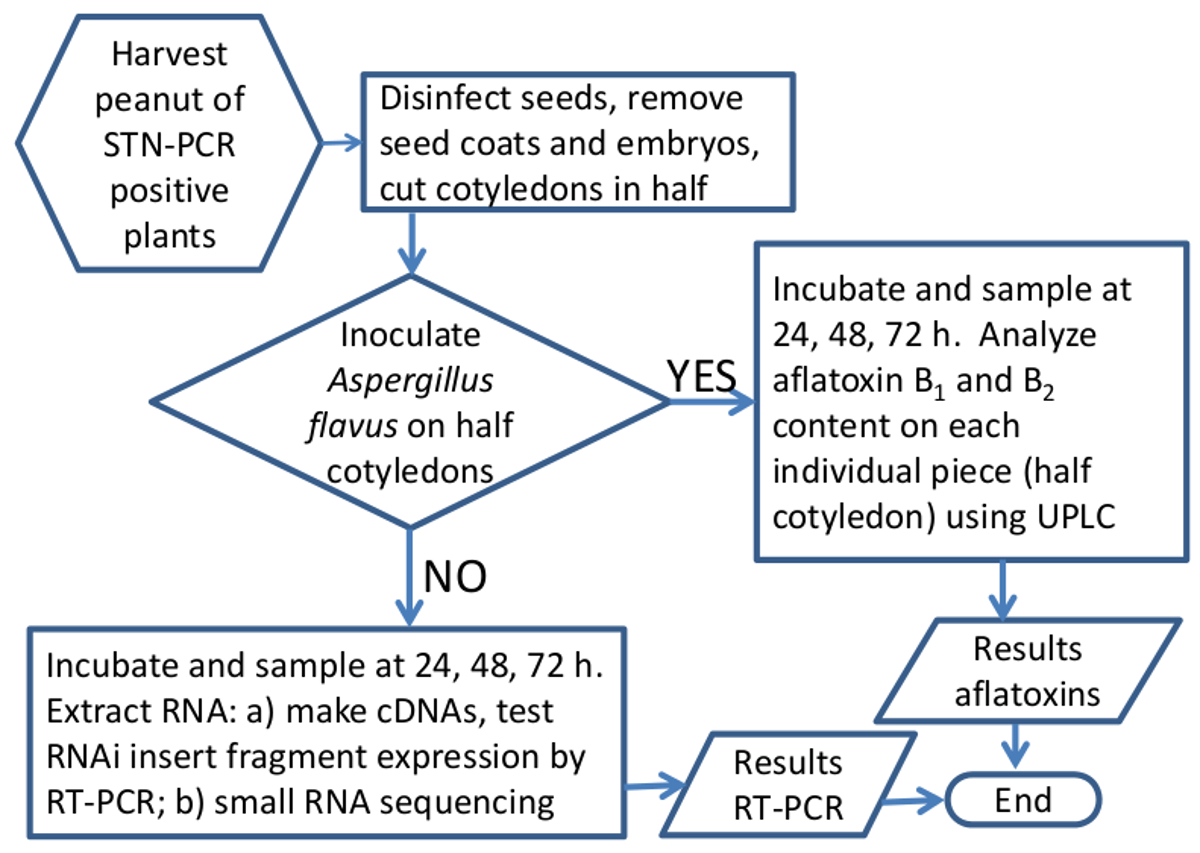{kind=link}
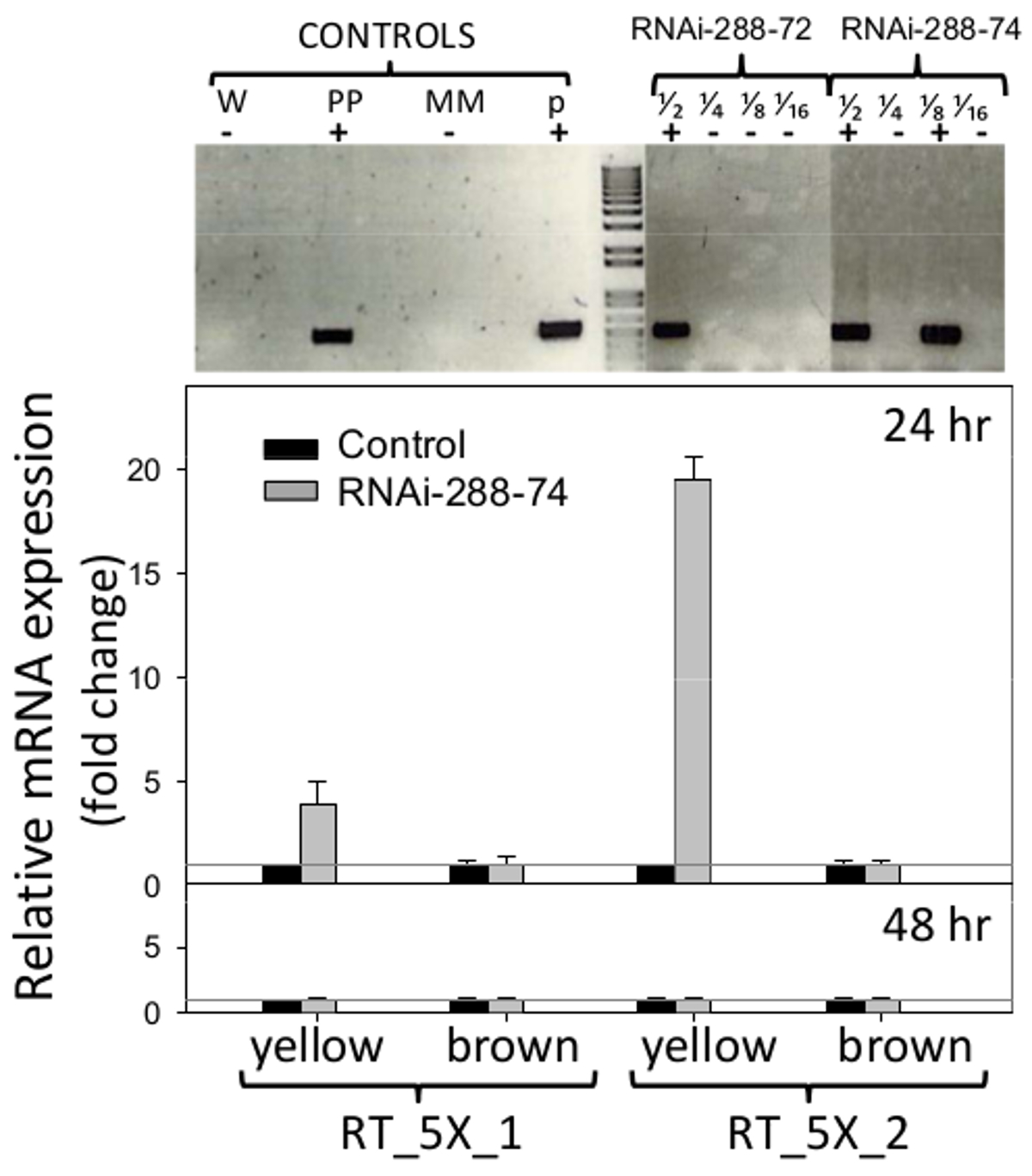
ARNi lignes d'arachide 288-72 et 288-74 ont montré la présence de l'sélectionnable fabricant NPTII lorsqu'il est testé par STN-PCR, sections de gel sont présentés dans la figure 6 (en haut), les images originales sont disponibles auprès des auteurs sur demande. Plasmide pCAPD n'a pas été utilisé comme témoin de la transformation car il encode répétitions inversées de deux gènes Cmr (résistance au chloramphénicol) et la CCDB (toxine) de l'effet inconnu sur les plantes. PCR en ligne d'arachide négative 288-9 et d'autres qui sont passés par le processus de régénération et a été cultivé dans les mêmes conditions que les lignes ARNi, ont été utilisés comme con négativecontrôle.

. Figure 6. Détection de transgéniques et d'expression en temps réel de l'ARNi insert Haut: monotube, la détection PCR nichée de lignes d'arachides transgéniques ARNi-288-72 et 288-74 ARNi, plantes positives. Contrôles: W (eau), PP (usine positif), MM (Master Mix), p (plasmide p5XCAPD); Les fractions 1/2 1/4 1/8 1/16 représentent des dilutions de 2 fois de l'ADN de fond. PCR en temps réel détection de l'expression du RNAi insert (jeux d'amorces: RT_5X_1, RT_5X_2, comme sur la figure 1, sur immature ( jaune) et matures (cotylédons) brunes de lignées transgéniques à 24 et 48h d'incubation; ligne grise: C T = 1. Les histogrammes représentent les moyens et les barres d'erreur standard (T) de trois échantillons biologiquesavec trois répétitions techniques. La quantification relative de l'insert ARNi a été normalisée par rapport à la gène de ménage actine comme contrôle interne et comparative fois l'expression du transgène, calculée comme indiqué en 5.2.2 et 5.3. S'il vous plaît cliquez ici pour voir une version plus grande de cette figure.
{kind=link}
Pour tester l'efficacité du contrôle éventuelle des plantes-hôte ARNi médiée de l'accumulation de l'aflatoxine, conidies fraîchement récoltées d'Aspergillus flavus NRRL 3357 ont été appliquées sur la surface de coupe des cotylédons et demi à partir de laquelle avaient été enlevés les embryons et testa (figures 3, 4). A. flavus NRRL 3357, dont le génome a été séquencé et a servi de base pour la conception p5XCAPD, a été aimablement fourni par le Dr Horn USDA-ARS-NPRL. L'invasion fongique résultant de la moitié cotylédons après 24, 48 et 72 heures à 30 ° C est spectacle n sur la figure 4. Des échantillons de cotylédons demi inoculés ont été prélevés à 24, 48, 72, 96 h d'incubation et analysés pour l'essentiel quatre aflatoxines B 1, B 2, G 1 et G 2 en utilisant UPLC et confirmée par LC-MS ; résultats sont présentés sur la figure 6. Les concentrations de l'aflatoxine ont été déterminées en utilisant un procédé publié, avec les adaptations 36. A 96 heures d'incubation, les cotylédons commencent à se désintégrer en raison de l'infection fongique. Ligne ARNi 288-72 exposé baisse significative des niveaux d'aflatoxines que le contrôle à toutes les dates d'échantillonnage dans les cotylédons immatures et à la plupart des dates d'échantillonnage dans les arrivées à maturité. Ligne ARNi 288-74 montré des niveaux significativement plus bas d'aflatoxines dans la plupart des dates d'échantillonnage. Les niveaux de signification du test de Tukey sont indiqués par des astérisques dans le graphique de la figure 7.
g7.jpg "/>
. Figure 7. aflatoxines B 1 et B 2 dans la moitié cotylédons d'arachide après incubation (24, 48, 72 et 96 h) avec Aspergillus flavus contrôle: graines de ligne 288-9 non transgénique; ARNi: graines de RNAi-288-72 et 288-74 ARNi, transgénique pour ARNi p5XCAPD au silence cinq gènes aflatoxine-synthèse. (A) l'aflatoxine B 1 dans les graines matures (brun); (B) les aflatoxines B 2 graines matures; (C) l'aflatoxine B 1 dans les graines immatures (de jaune) et (D) les aflatoxines B 2 dans les graines immatures. Les valeurs moyennes avec les barres d'erreur standards correspondants (T) d'échantillons biologiques en double sont représentés. Des différences statistiquement significatives test * de Tukey: p ≤ 0,05, **: p ≤ 0,01, ***: p ≤ 0,001.s: //www.jove.com/files/ftp_upload/53398/53398fig7large.jpg "target =" _ blank "> S'il vous plaît cliquer ici pour voir une version plus grande de cette figure.
Dans l'ensemble, l'ARNi a montré 288-72 long de l'expérience (24 à 96 h d'incubation), une réduction de 94% -100% de l'aflatoxine B 2, et une réduction de 90% -100% de l'aflatoxine B 1 par rapport au témoin. RNAi-288-74 a montré une réduction de 63% -100% de l'aflatoxine B 2 et 60% -100% de réduction dans l'aflatoxine B 1, la figure 7.
Amorces utilisées pour la détection par PCR en temps réel de l'expression de l'insert ARNi sont présentés dans la figure 1. Cotylédons arachides semences ont été analysés sans embryons, de retirer leurs défenses naturelles et être capable de détecter l'effet potentiel de l'ARNi sur la zone la plus exposée aux l'invasion fongique, les cotylédons. Expression de l'ARNi insert détecté dans les cotylédons immatures (jaune) de la ligne 288 à 74 par jeu d'amorces RT_5X_1 était quatre fois au cours de la C T = 1 seuil du contrôle négatif, et en ensemble d'amorces RT_5X_2 était 19 fois au-dessus du seuil, le tout à 24 heures d'incubation. Au moins trois des cinq fragments de gène consécutifs utilisés dans la transformation d'arachide, 5027, 7223 et 7228 (aflep, AFLS / aflJ, et AFLC / pksA, respectivement) ont été détectés par RT-PCR (Figure 1, 6). Expression de l'insert ARNi n'a pas été détecté dans les cotylédons matures à 24 h, ou sur cotylédons matures ou immatures, à 48 heures d'incubation, la figure 6 (en bas).
1| Peanut Ligne | Durée de l'échantillonnage | Les échantillons pour RT-PCR | Les échantillons destinés à l'analyse des aflatoxines (inoculé) | Nombre de semences | ||||
| (non inoculé) | ||||||||
| Rep 1 | Rep 2 | Rep 3 | Rep 1 | Rep 2 | Rep 3 | |||
| ARNi (jaune) | 24 h | 1 | 1 | 1 | 1 | 1 | 1 | 4.5 |
| 48 h | 1 | 1 | 1 | 1 | 1 | |||
| 72 h | 1 | 1 | 1 | 1 | 1 | 1 | ||
| Contrôle | 24 h | 1 | 1 | 1 | 1 | 1 | 1 | 4.5 |
| (jaune) | 48 h | 1 | 1 | 1 | 1 | 1 | 1 | |
| 72 h | 1 | 1 | 1 | 1 | 1 | 1 | ||
Tableau 2. Exemple de montage de petits échantillons pour analyser l'expression des gènes et de l'accumulation de l'aflatoxine dans les ARNi graines d'arachide. L'analyse complète d'expression et d'aflatoxines pour un groupe de maturité (ie., Jaune), avec trois périodes d'échantillonnage (24, 48 et 72 h) , en trois exemplaires, exigerait 4,5 semences, chaque numéro un dans le tableau représente la moitié cotylédons.
Discussion
Plante-hôte ARNi médiée par le silençage de gènes dans des pathogènes fongiques a été démontré 27,43, cependant, il n'y a pas de publications montrant la faisabilité du contrôle ARNi médiée par l'accumulation de mycotoxines dans les plantes. Un facteur limitant pour ces études à l'arachide était l'absence d'une méthode pour évaluer une accumulation phénotype sans l'aflatoxine dans les plantes individuelles, que les feuilles ne montrent pas de symptômes sur une infection fongique de gousses souterraines. En outre, l'accumulation sans normalement distribué des aflatoxines, et la nécessité pour les grands échantillons pour l'analyse chimique 15,16 ont entravé la quantification de l'effet potentiel sur l'ARNi une seule plante. La méthode présentée ici se compose de 72 expériences de RH en utilisant cinq graines pour effectuer trois prélèvements 24 h-intervalle en trois exemplaires (tableau 1, figure 7). Par rapport à l'analyse des aflatoxines typique qui ne nécessite pas moins de 100 g de graines, notre procédé est particulièrement approprié pour individual d'événements transgéniques de plants d'arachide qui produisent initialement pas plus de deux ou trois gousses.
RNA silencing médiée par la synthèse de l'aflatoxine a été démontrée par la transformation génétique Aspergillus flavus et A. parasiticus. Depuis aflR est un régulateur principal de la production d'aflatoxine dans A. flavus et A. 44,45 parasiticus, il devient une cible intéressante pour le silençage de l'ARN à médiation par les plantes. Cependant, les variations génétiques dans aflR a été démontré chez les espèces d'Aspergillus 46, et ces variantes génétiques pourraient échapper taire si il n'y a pas de séquence parfaite correspondant au signal ARNi produite dans la plante hôte. Ainsi, aflR était l'une des cibles pour réduire au silence dans le vecteur p5XCAPD, mais n'a pas été le seul. Répétitions inversées du gène aflR introduits dans A. flavus et A. parasiticus par transformation conduit à taire et peu ou pas de produition des aflatoxines 47 (McDonald et al., 2005b). Aussi, faire taire le gène AFLD a empêché la production d'aflatoxine jusqu'à 98% en A. flavus et A. parasiticus dans la transformation directe 48. Pour augmenter la probabilité de succès dans notre système, l'arachide a été transformé avec des fragments inversées répétées de cinq gènes impliqués dans la production d'aflatoxine dans A. flavus. Ici, il est indiqué que l'utilisation de p5XCAPD qui cible pour faire taire plusieurs gènes dans la voie de synthèse de l'aflatoxine, 90% des niveaux inférieurs à 100% de l'aflatoxine B 1 et B 2 ont été atteints dans la ligne de 288 à 72, et les niveaux 60-100% inférieurs accumulé dans ligne 288 à 74 par rapport à la commande, lorsque les cotylédons et demi ont été inoculés avec A. flavus, les figures 4, 7. Plus important encore, cette méthode a détecté des différences statistiquement significatives dans l'accumulation de l'aflatoxine par des lignes 288-72, 288-74 contre le contrôle tout au long de l'expérience en appliquant statis paramétriquetics, figure 7. Compte tenu de la petite taille de l'échantillon, il est important de souligner la nécessité d'utiliser une méthode puissante pour détecter les aflatoxines, ces expériences ont été analysés par UPLC qui a une haute résolution, cinq fois de meilleures performances et une sensibilité trois fois plus élevé que HPLC 49.
Expression de l'ARNi insert dans 288-74 n'a été détectée dans les cotylédons immatures (jaune) à 24 heures d'incubation. L'insert ARNi n'a pas été détecté par RT-PCR sur des cotylédons matures de 288 à 74 à 24 h, ou sur un groupe d'échéance à 48 h, la figure 6. Ce même phénomène a été observé dans d'autres lignes d'arachides transgéniques ARNi (Arias, RS 2015 non publié), où généralement les transcriptions ARNi ont été détectés que sur des cotylédons immatures à 24 h. Des échantillons d'ARN ont été traités par la DNase avant la synthèse d'ADNc, les données ont été normalisées pour le niveau d'expression d'actine et aucun signe de contamination de l'ADN a été observée. Si l'ADN a pu être présente dans les échantillons, il devraitont été détectés dans les échantillons de 48 h aussi, mais de façon constante que fut pas le cas. Expression sous le contrôle de la 35S-promoteur est pas toujours uniforme; il peut être affecté par des conditions environnementales 50, type de tissu et le stade de développement 51,52. Dans le même temps, dans la voie de l'interférence ARN, le taux de dégradation de l'ARNm et le taux de décroissance siRNA peuvent varier considérablement 53. Il est possible que la dégradation rapide de l'ARNm par le mécanisme de l'interférence ARN aurait pu empêcher la détection de l'ARNm et 48 heures d'incubation. Que l'absence d'expression à 48 h était due à de faibles 35S-promoteur entraîné la transcription ou à la dégradation rapide de l'ARN double brin par Dicer reste sans réponse. Ainsi, la détection de petits ARN par séquençage à haut débit donnerait une meilleure idée sur les processus qui se déroulent à travers ARNi 54 dans ces expériences. Cependant, depuis RNA silencing propage systémique, principalement à travers le phloème de photosynthaïr sources au saccharose puits (dans ce cas arachide graines) 55, le musellement de l'aflatoxine-synthèse peut se produire dans les graines sans expression locale de l'ARNi insert. Beaucoup de recherche reste à faire pour déterminer le niveau de petits ARN interférents (siRNA) nécessaires pour empêcher l'accumulation de l'aflatoxine dans les graines de seuil. Il est important de souligner le fait que les deux, expression de l'ARNm de l'ARNi construire (Figure 6), et l'accumulation des aflatoxines B 1 et B 2 (figure 7) a montré des résultats différents pour les immatures (jaune) vs. mature (marron) cotylédons. Plants d'arachide ont une croissance indéterminée, qui est, ils présentent à la récolte d'une gamme de dosettes échéance, la figure 2. En outre, les graines de différents groupes de maturité diffèrent par leur composition chimique, par exemple., 2,4% de saccharose dans les graines immatures, et de 1,9% en graines matures dans les mêmes conditions sur le terrain 56,57. Ainsi, pour comprendre la réelle efficiency de contrôle d'ARN médiée par l'accumulation de l'aflatoxine, il est important d'analyser séparément les groupes de maturité.
Une défense naturelle de graines d'arachide est la production de phytoalexines, qui varie dans la diversité des composés produits et leurs quantités relatives en fonction de la maturité des graines et des conditions environnementales 58-61, et il est particulièrement élevé dans les embryons comparativement à 62 cotylédons. Les embryons ont également des concentrations significativement plus élevées d'acides nucléiques, aussi bien ADN et l'ARN que les cotylédons (RS Arias, non publié). Comme graines d'arachide mûrissent, les changements dans leur physiologie et leur composition chimique se produisent 63. Antioxydants phénoliques dans les arachides forme de testa tanins condensés avec une activité fongistatique 64; cela est également évidente dans la couleur de la mésocarpe qui reflète les stades de maturité, jaune au noir 35, comme sa teneur en tanins et composés phénoliques augmente avec la maturité 65. Ainsi, la présence de testa ou d'embryons dans l'expérience, compte tenu de leurs propriétés antimicrobiennes, auraient pu limiter la croissance des champignons et donc surestimé l'effet de l'ARNi au silence, par conséquent, ils ont été enlevés. En outre, la suppression de Testa et embryons permet de limiter les sources de variation dans l'analyse, que la moitié des cotylédons qui porte l'embryon aura plus de phytoalexines et plus de contenu de l'ARN.
En plus de l'analyse par groupes de maturité et l'élimination des testa et de l'embryon dans ces expériences, il est important de souligner quelques observations: a) si les résultats sont indiqués pour un maximum de 96 heures d'incubation, il est recommandé de ne pas utiliser plus de 72 h pour obtenir des résultats cohérents, que les graines se dégradent de 96 h; et b), tandis que les cotylédons demi de la même graine, bien échantillonné au hasard, ne constituent pas des échantillons parfaitement indépendants, RT-PCR et l'accumulation de l'aflatoxine dans les événements transgéniques ont montré variation minimale entre les graines. En outre, un spores fongiques précise compte, le volume de l'inoculums de 2 pl, et l'application de spores sur la surface des cotylédons évitant dégoulinant sur les côtés de coupe sont importants pour assurer que les spores germées sont exposés à du tissu végétal. L'eau / agar sur les plaques devrait être à 1,5% (p / v), l'agar mou provoque le ruissellement des spores comme indiqué sur la dernière image de la figure 4 (en bas). Si la disponibilité des semences à partir d'un événement transgénique notamment être limité, l'échantillonnage peut être fait en double exemplaire au lieu de triple obtenir des résultats similaires (par exemple, la figure 7); Cependant, les échantillons en triple aideront à réduire l'erreur-type. La seule limitation de cette méthode est qu'elle nécessite un système très sensible (UPLC) pour la détection de l'aflatoxine / quantification, mais en même temps, ce qui réduit la probabilité d'une surestimation de l'effet de l'ARNi doit aflatoxines pas être détecté par des méthodes moins sensibles.
En conclusion, cette méthode offre pour la première fois une approche fiable pour étudier laeffet de l'ARNi dans le contrôle des aflatoxines. Réduire le temps pour une expérience de toute une saison de culture à moins d'une semaine, cette méthode sera énormément accélérer la recherche sur l'ARNi arachide / Aspergillus pathosystème vers l'atténuation et / ou de l'élimination des aflatoxines.
Déclarations de divulgation
The authors have nothing to disclose or any conflicts of interest.
Remerciements
This work received the financial support of USDA-ARS CRIS project 6604-21000-004-00D, CRIS project 6604-42000-008-00D, and USAID Feed-the-Future program Agreement number 58-0210-3-012. We thank Valerie Orner, LaTanya Johnson, Joseph Powell and Kathy Gray for their technical assistance. Mention of trade names or commercial products in this article is solely for the purpose of providing specific information and does not imply recommendation or endorsement by the US Department of Agriculture.
matériels
| Name | Company | Catalog Number | Comments |
| Primers, oligonucleotides | DNA Technologies, Coralville, IA, USA | n/a | |
| Dneasy Plant Mini Kit | Qiagen, Valencia, CA | 69106 | |
| Czapek Dox agar medium | Oxoid, by Thermo Fisher Scientific, Waltham, MA | CM0095 | |
| Agar | Thermo Fisher Scientific, Waltham, MA | BP 1423 | |
| Freezer -80 °C | n/a | n/a | |
| Aluminum Oxide, Al2O3 | Fisher Scientific | A941 | |
| SPE Reservoirs 1.5 ml | Grace Davison Discovery Scientific | 210011 | |
| Frits for 1.5 ml SPE reservoir | Grace Davison Discovery Scientific | 211401 | |
| Autosampler vials | Waters Corporation, Milford, MA | 186005221 | |
| Waters Acquity Ultra-Performance Liquid-Chromatography (UPLC) instrument; UPLC-H-Class Quaternary Solvent Manager; UPLC Sample Manager; UPLC Fluorescent detector (FLR); UPLC BEH C18 2.1 mm x 50 mm, 1.7 mm column | Waters Corporation, Milford, MA | ||
| Finnigan LCQ Advantage MAX ion trap mass spectrometer, with Xcalibur version 1.4 software | Thermo Electron Corp., San Jose, CA | ||
| Aflatoxin standards, B1, B2, G1 and G2 | Sigma-Aldrich, St. Louis, MO | A6636; A9887; A0138; A0263 | |
| Systat Software 12.2 | SYSTAT Software Inc., Point Richmond, CA | ||
| Trizol reagent | Invitrogen, CA | 15596-018 | |
| SuperScript III First Strand Synthesis Super Mix | Invitrogen, CA | 11752-050 | |
| ABI 7500 Real-Time PCR | Lifetechnologies, Grand Island, NY | 4406984 | |
| Luria Broth-Miller | Fisher Scientific | R453642 | |
| pENTR1A | Invitrogen, CA | A10462 | |
| LR Clonase II enzyme mix | Invitrogen, CA | 11791-020 | |
| T4 DNA Ligase | NEB Biolabs | M0202L | |
| Gelrite | Sigma-Aldrich, St. Louis, MO | G1919 | |
| Acetosyringone | Sigma-Aldrich, St. Louis, MO | D134406 | |
| QIAcube robot workstation | Qiagen, Valencia, CA | 9001292 | |
| Antibiotics: kanamycin, cefotaxime, gentamicin; streptomycin | Goldbio, St. Louis, MO | cef.: C-104-25; kan: K-120-5; gent.: G-400-1; strep.: S-150-50 | |
| Platinum Taq DNA Polymerase High Fidelity | Invitrogen, CA | 11304-029 |
Références
- Williams, J. H., et al. Human aflatoxicosis in developing countries: a review of toxicology, exposure, potential health consequences, and interventions. The American Journal of Clinical Nutrition. 80, 1106-1122 (2004).
- American Association for Cancer Research: AACR. An evaluation of chemicals and industrial processes associated with cancer in humans based on human and animal data: IARC Monographs Volumes 1 to 20. Cancer Research. 40, 1-12 (1980).
- Turner, P. C. The molecular epidemiology of chronic aflatoxin driven impaired child growth. Scientifica. , (2013).
- Rasooly, R., Hernlem, B., He, X., Friedman, M. Non-linear relationships between aflatoxin B1 levels and the biological response of monkey kidney vero cells. Toxins (Basel). 5, 1447-1461 (2013).
- Gong, Y. Y., et al. Determinants of aflatoxin exposure in young children from Benin and Togo, West Africa: the critical role of weaning. International Journal of Epidemiology. 32, 556-562 (2003).
- Eaton, D. L., Groopman, J. D. The toxicology of aflatoxins: human health, veterinary, and agricultural significance. , Academic Press. (1994).
- Murugavel, K. G., et al. Prevalence of aflatoxin B1 in liver biopsies of proven hepatocellular carcinoma in India determined by an in-house immunoperoxidase test. Journal of Medical Microbiology. 56, 1455-1459 (2007).
- Wang, J. S., et al. Hepatocellular carcinoma and aflatoxin exposure in Zhuqing Village, Fusui County, People's Republic of China. Cancer Epidemiology, Biomarkers & Prevention. 10, American Association for Cancer Research. 143-146 (2001).
- Azziz-Baumgartner, E., et al. Case-control study of an acute aflatoxicosis outbreak, Kenya, 2004. Environmental Health Perspectives. 113, 1779-1783 (2005).
- Lye, M. S., Ghazali, A. A., Mohan, J., Alwin, N., Nair, R. C. An outbreak of acute hepatic encephalopathy due to severe aflatoxicosis in Malaysia. American Journal of Tropical Medicine and Hygiene. 53, 68-72 (1995).
- Villers, P. Aflatoxins and safe storage. Frontiers in Microbiology. 5, 158(2014).
- Kensler, T. W., Roebuck, B. D., Wogan, G. N., Groopman, J. D. Aflatoxin: A 50-year odyssey of mechanistic and translational toxicology. Toxicological Sciences. 120, S28-S48 (2011).
- Dorner, J. W., Cole, R. J., Wicklow, D. T. Aflatoxin reduction in corn through field application of competitive fungi. Journal of Food Protection. 62, 650-656 (1999).
- Cotty, P. J., Bhatnagar, D. Variability among atoxigenic Aspergillus flavus strains in ability to prevent aflatoxin contamination and production of aflatoxin biosynthetic-pathway enzymes. Applied and Environmental Microbiology. 60, 2248-2251 (1994).
- Whitaker, T. B. Standardisation of mycotoxin sampling procedures: an urgent necessity. Food Control. 14, 233-237 (2003).
- Whitaker, T. B., Dorner, J. W., Giesbrecht, F. G., Slate, A. B. Variability among aflatoxin test results on runner peanuts harvested from small field plots. Peanut Science. 31, 59-63 (2004).
- Fire, A., et al. Potent and specific genetic interference by double-stranded RNA in Caenorhabditis elegans. Nature. 391, 806-811 (1998).
- Rafael, D., et al. EMT blockage strategies: Targeting Akt dependent mechanisms for breast cancer metastatic behaviour modulation. Current Gene Therapy. , (2015).
- Li, G., Chang, H., Zhai, Y. P., Xu, W. Targeted silencing of inhibitors of apoptosis proteins with siRNAs: a potential anti-cancer strategy for hepatocellular carcinoma. Asian Pacific. Journal of Cancer Prevention: APJCP. 14, 4943-4952 (2013).
- Koldehoff, M. Targeting bcr-abl transcripts with siRNAs in an imatinib-resistant chronic myeloid leukemia patient: challenges and future directions. Methods in Molecular Biology. 1218, 277-292 (2015).
- Zhang, J., et al. Pest control. Full crop protection from an insect pest by expression of long double-stranded RNAs in plastids. Science. 347, 991-994 (2015).
- Ajjappala, H., Chung, H. Y., Sim, J. S., Choi, I., Hahn, B. S. Disruption of prefoldin-2 protein synthesis in root-knot nematodes via host-mediated gene silencing efficiently reduces nematode numbers and thus protects plants. Planta. 241, 773-787 (2015).
- Jose, A. M., Hunter, C. P. Transport of sequence-specific RNA interference information between cells. Annual Review of Genetics. 41, 305-330 (2007).
- Vazquez, F., Hohn, T. Biogenesis and biological activity of secondary siRNAs in plants. Scientifica. , Hindawi Publishing Corporation. (2013).
- Tinoco, M. L. P., Dias, B. B. A., Dall'Astta, R. C., Pamphile, J. A., Aragao, F. J. L. In vivo trans-specific gene silencing in fungal cells by in planta expression of a double-stranded RNA. BMC Biology. 8, (2010).
- Govindarajulu, M., Epstein, L., Wroblewski, T., Michelmore, R. W. Host-induced gene silencing inhibits the biotrophic pathogen causing downy mildew of lettuce. Plant Biotechnology Journal. , (2014).
- Yin, C., Jurgenson, J. E., Hulbert, S. H. Development of a host-induced RNAi system in the wheat stripe rust fungus Puccinia striiformis f. sp. tritici. Molecular Plant-Microbe Interactions. 24, 554-561 (2011).
- Ghag, S. B., Shekhawat, U. K., Ganapathi, T. R. Host-induced post-transcriptional hairpin RNA-mediated gene silencing of vital fungal genes confers efficient resistance against Fusarium. wilt in banana. Plant Biotechnology Journal. 12, 541-553 (2014).
- Filichkin, S. A., et al. Efficiency of gene silencing repeats vs. transitive RNAi in Arabidopsis: direct inverted vectors. Plant Biotechnology Journal. 5, 615-626 (2007).
- Sciaky, D., Montoya, A. L., Chilton, M. D. Fingerprints of Agrobacterium Ti Plasmids. Plasmid. 1, 238-253 (1978).
- Clark, D. J., Maaloe, O. DNA Replication and Division Cycle in Escherichia coli. Journal of Molecular Biology. 23, 99-112 (1967).
- Murashige, T., Skoog, F. A revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiol Plantarum. 15, 473-497 (1962).
- Srinivasan, T., Kumar, K. R. R., Kirti, P. B. Establishment of efficient and rapid regeneration system for some diploid wild species of Arachis. Plant Cell Tissue and Organ Culture. 101, 303-309 (2010).
- Gomes, A. L. V., et al. Single-tube nested PCR using immobilized internal primers for the identification of dengue virus serotypes. Journal of Virology Methods. 145, 76-79 (2007).
- Williams, E. J., Drexler, J. S. A non-destructive method for determining peanut pod maturity. Peanut Science. 8, 134-141 (1981).
- Sobolev, V. S., Dorner, J. W. Cleanup procedure for determination of aflatoxins in major agricultural commodities by liquid chromatography. Journal of AOAC International. 85, 642-645 (2002).
- Empower Software, Getting Started Guide. , Waters Corporation. Milford, MA. Available from: http://sites.chem.colostate.edu/diverdi/C431/experiments/high%20pressure%20liquid%20chromatography/references/Empower%20getting%20started%2071500031203rA.pdf (2002).
- Biselli, S., Hartig, L., Wegner, H., Hummert, C. Analysis of Fusarium. toxins using LC-MS-MS: Application to various food and feed matrices. LC GC North America. 23, 404-413 (2005).
- Arias, R. S., Sobolev, V. S., Orner, V. A., Dang, P. M., Lamb, M. C. Potential involvement of Aspergillus flavus laccases in peanut invasion at low water potential. Plant Pathology. 63, 353-363 (2014).
- Dang, P. M., Chen, C. Y., Holbrook, C. C. Evaluation of five peanut (Arachis hypogaea) genotypes to identify drought responsive mechanisms utilising candidate-gene approach. Functional Plant Biology. 40, 1323-1333 (2013).
- Schmittgen, T. D., Livak, K. J. Analyzing real-time PCR data by the comparative C-T method. Nature Protocols. 3, 1101-1108 (2008).
- Amaike, S., Keller, N. P. Aspergillus flavus. Annual Review of Phytopathology. 49, 107-133 (2011).
- Nowara, D., et al. HIGS: Host-induced gene silencing in the obligate biotrophic fungal pathogen Blumeria graminis. Plant Cell. 22, 3130-3141 (2010).
- Woloshuk, C. P., et al. Molecular characterization of aflR, a regulatory locus for aflatoxin biosynthesis. Applied and Environmental Microbiology. 60, 2408-2414 (1994).
- Price, M. S., et al. The aflatoxin pathway regulator AflR induces gene transcription inside and outside of the aflatoxin biosynthetic cluster. FEMS Microbiology Letters. 255, 275-279 (2006).
- Ehrlich, K. C., Montalbano, B. G., Cotty, P. J. Sequence comparison of aflR from different Aspergillus. species provides evidence for variability in regulation of aflatoxin production. Fungal Genetics and Biology. 38, 63-74 (2003).
- McDonald, T., Brown, D., Keller, N. P., Hammond, T. M. RNA silencing of mycotoxin production in Aspergillus and Fusarium species. Molecular Plant Microbe Interactions. 18, 539-545 (2005).
- Abdel-Hadi, A. M., Caley, D. P., Carter, D. R., Magan, N. Control of aflatoxin production of Aspergillus flavus. and Aspergillus parasiticus. using RNA silencing technology by targeting aflD. (nor-1) gene. Toxins (Basel). 3, 647-659 (2011).
- Swartz, M. E. Ultra performance liquid chromatography (UPLC): An introduction: Separation Science Redefined. LCGC North America. , Suppl ement 8. 8-14 (2005).
- Maghuly, F., Khan, M. A., Fernandez, E. B., Druart, P., Watillon, B., Laimer, M. Stress regulated expression of the GUS-marker gene (uidA) under the control of plant calmodulin and viral 35S promoters in a model fruit tree rootstock: Prunus incisa x serrula. Journal of Biotechnology. 135, 105-116 (2008).
- de Mesa, M. C., Santiago-Doménech, N., Pliego-Alfaro, F., Quesada, M. A., Mercado, J. A. The CaMV 35S promoter is highly active on floral organs and pollen of transgenic strawberry plants. Plant Cell Reports. 23, 32-38 (2004).
- Sunilkumar, G., Mohr, L., Lopata-Finch, E., Emani, C., Rathore, K. S. Developmental and tissue-specific expression of CaMV 35S promoter in cotton as revealed by GFP. Plant Molecular Biology. 50, 463-474 (2002).
- Groenenboom, M. A. C., Maree, A. F. M., Hogeweg, P. The RNA silencing pathway: The bits and pieces that matter. PLoS Computational Biology. 1, 155-165 (2005).
- Zhao, D., Song, G. Q. High-throughput sequencing as an effective approach in profiling small RNAs derived from a hairpin RNA expression vector in woody plants. Plant Science: an International Journal of Experimental Plant Biology. 228, 39-47 (2014).
- Kamthan, A., Chauduri, A., Kamthan, M., Datta, A. Small RNAs in plants: recent development and application for crop improvement. Frontiers in Plant Science. 6, 208(2015).
- Manda, A., Bodapati, P. N., Rachaputi, N. C., Wright, G., Fukai, S. Aflatoxins and their relationship with sugars in peanut (Arachis hypogaea L). 4th International Crop Science Congress, 2004, , Available from: http://www.cropscience.org.au/icsc2004/poster/5/1/3/625_manda.htm (2004).
- Uppala, S. S. Factors affecting pre-harvest aflatoxin contamination of peanut (Arachis hypogaea L). , Auburn University. (2011).
- Sobolev, V. S. Localized production of phytoalexins by peanut (Arachis hypogaea) kernels in response to invasion by Aspergillus species. Journal of Agricultural and Food Chemistry. 56, 1949-1954 (2008).
- Sobolev, V. S., Guo, B. Z., Holbrook, C. C., Lynch, R. E. Interrelationship of phytoalexin production and disease resistance in selected peanut genotypes. Journal of Agricultural and Food Chemistry. 55, 2195-2200 (2007).
- Sobolev, V. S., Neff, S. A., Gloer, J. B. New stilbenoids from peanut (Arachis hypogaea) seeds challenged by an Aspergillus caelatus strain. Journal of Agricultural and Food Chemistry. 57, 62-68 (2009).
- Dorner, J. W., Cole, R. J., Sanders, T. H., Blankenship, P. D. Interrelationship of kernel water activity, soil temperature, maturity, and phytoalexin production in preharvest aflatoxin contamination of drought-stressed peanuts. Mycopathologia. 105, 117-128 (1989).
- Sobolev, V. S. Production of phytoalexins in peanut (Arachis hypogaea) seed elicited by selected microorganisms. Journal of Agricultural and Food Chemistry. 61, 1850-1858 (2013).
- Basha, S. M. M., Cherry, J. P., Young, C. T. Changes in free amino acids, carbohydrates, and proteins of maturing seeds from various peanut (Arachis hypogaea L.) cultivars. Cereal Chemistry. 53, 586-596 (1976).
- Lansden, J. A. Aflatoxin inhibition and fungistasis by peanut tannins. Peanut Science. 9, 17-20 (1982).
- Yen, G. C., Duh, P. D., Tsai, C. L. Relationships between antioxidant activity and maturity of peanut hulls. Journal of Agricultural and Food Chemistry. 41, 67-70 (1993).
Réimpressions et Autorisations
Demande d’autorisation pour utiliser le texte ou les figures de cet article JoVE
Demande d’autorisationThis article has been published
Video Coming Soon
À PROPOS DE JoVE
Copyright © 2025 MyJoVE Corporation. Tous droits réservés.