
Un abonnement à JoVE est nécessaire pour voir ce contenu. Connectez-vous ou commencez votre essai gratuit.
Method Article
Criblage direct en culture liquide pour l’évaluation de la production de protéines hétérologues à l’aide d’un mutant auxotrophe d’Aspergillus oryzae
Dans cet article
Résumé
Une méthode de criblage en culture liquide directe (DLC) a été mise au point qui réduit considérablement le temps nécessaire à la transformation protoplaste du champignon filamenteux, Aspergillus oryzae, par le polyéthylène glycol (PEG), lorsqu’elle est utilisée pour évaluer la production sécrétoire de protéines hétérologues. Cette méthode augmente considérablement le débit du protocole d’évaluation.
Résumé
Aspergillus oryzae, un champignon filamenteux, est l’un des hôtes les plus largement utilisés pour les applications industrielles, y compris la production à grande échelle de protéines. Une méthode de transformation des protoplastes médiée par le polyéthylène glycol (PEG) est généralement utilisée pour l’introduction de gènes hétérologues dans A. oryzae. La méthode conventionnelle nécessite généralement trois semaines pour le criblage des transformants favorables. Ici, une nouvelle technique, la méthode de criblage direct par culture liquide (DLC), est introduite, qui réduit le temps de criblage à six jours dans un format de flacon de 200 ml ou à 10 jours dans un format de microplaque de 24 puits. La méthode de criblage DLC assure l’acquisition de transformants positifs et l’évaluation de la production sécrétoire de protéines hétérologues en une seule étape, contrairement à la méthode de criblage conventionnelle où deux étapes distinctes sont nécessaires pour la même chose. Le protocole de transformation des protoplastes d’A. oryzae par le PEG est décrit, qui comprend cinq étapes : préparation de la suspension de spores fraîches, préculture, préparation des protoplastes, introduction de l’ADN et criblage DLC. Pour obtenir des résultats réussis dans le criblage DLC, il est essentiel d’utiliser un milieu riche en nutriments avec une pression osmotique optimisée. Le protocole devrait populariser davantage l’utilisation d’A. oryzae comme hôte de choix dans la production industrielle de protéines.
Introduction
Aspergillus oryzae est un micro-organisme important dans l’industrie alimentaire japonaise qui est utilisé depuis plus de 1 000 ans dans la production d’aliments fermentés, tels que le saké (vin de riz), le shoyu (sauce soja) et le miso (pâte de soja)1,2. Il a la capacité de sécréter une grande quantité de protéines, telles que les protéases et les amylases3. Des informations sur le séquençage du génome d’A. oryzae sont également disponibles4. De plus, des techniques de génie génétique puissantes et utiles sur le plan pratique ont été mises au point pour ce champignon 5,6,7,8,9,10. Des transformants favorables ont été utilisés comme hôtes pour la production sécrétoire de protéines hétérologues 11,12,13,14.
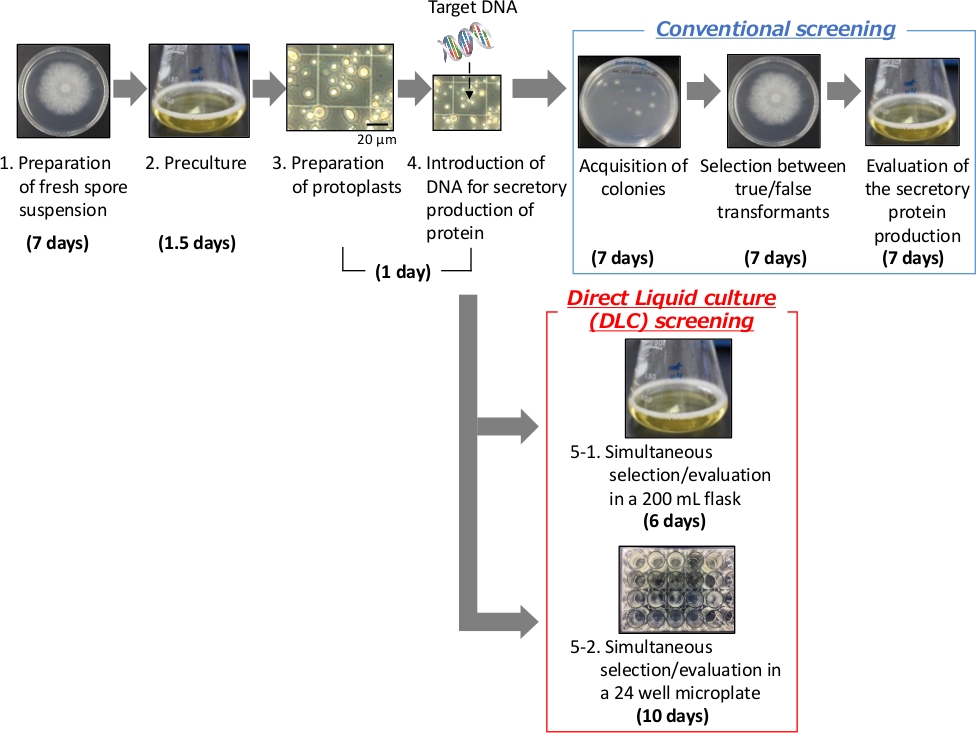
L’électroporation, la transformation médiée par Agrobacterium et la transformation protoplaste médiée par le polyéthylène glycol (PEG) sont les techniques utilisées pour introduire des gènes hétérologues dans A. oryzae 15,16,17. La méthode de transformation du protoplaste médiée par le PEG a été largement utilisée depuis qu’elle a été signalée pour la première fois pour Neurospora crassa en 197918. Dans cette méthode, des protoplastes sont préparés et mélangés avec du PEG et le gène hétérologue qui doit être introduit dans les cellules. Les parois des protoplastes sont compromises enzymatiquement, ce qui rend les cellules vulnérables au stress physique et aux changements de pression osmotique19,20. La méthode de criblage conventionnelle utilisée dans la production sécrétoire de protéines hétérologues comprend trois étapes : l’acquisition de transformants positifs (sur plaque de gélose molle), la sélection de vrais et de faux transformants (sur plaque de gélose) et l’évaluation de la production sécrétoire de protéines hétérologues (en culture liquide) ; chacune de ces étapes prend sept jours (Figure 1). Ainsi, la méthode conventionnelle nécessite généralement environ trois semaines.
Le temps nécessaire au criblage des cellules transformées d’A. oryzae est beaucoup plus long que celui requis pour d’autres micro-organismes couramment utilisés dans la recherche en biotechnologie, ce qui rend le processus plus lourd. Par exemple, lorsqu’on utilise Escherichia coli comme hôte, il faut environ deux jours entre l’introduction de l’ADN et la confirmation de son effet21.
Pour contourner la limitation associée à l’utilisation d’A. oryzae mentionnée ci-dessus, une nouvelle méthode de criblage en culture liquide directe (DLC) est introduite, qui permet un dépistage plus rapide et plus simple pour l’évaluation de la production sécrétoire de protéines hétérologues (Figure 1). Dans la méthode DLC, un mutant auxotrophe uridine et un milieu liquide riche en nutriments sont utilisés. À l’aide de cette méthode, l’étape de criblage peut être terminée dans les six jours suivant l’introduction de l’ADN dans les protoplastes dans un flacon de 200 ml ou dans un délai de 10 jours dans un format de microplaque de 24 puits. De plus, la préparation longue et laborieuse des milieux de plaque de gélose n’est pas nécessaire dans cette méthode. Il y a un énorme avantage à utiliser la méthode nouvellement décrite, surtout si l’on considère le fait que la méthode conventionnelle nécessite deux milieux différents : la plaque de gélose molle pour l’acquisition de transformants positifs, qui nécessite une manipulation minutieuse et un contrôle de la température, et la plaque de gélose solide pour la sélection de vrais transformants.

Figure 1 : Schéma de la transformation protoplaste du champignon filamenteux Aspergillus oryzae par le polyéthylène glycol (PEG).
(Panneau supérieur) Méthode de criblage conventionnelle. (Panneau inférieur) Méthode de criblage par culture liquide directe (DLC). Veuillez cliquer ici pour voir une version agrandie de cette figure.
{kind=link}
Protocole
1. Préparation de la suspension de spores fraîches
- Inoculer 20 μL d’une suspension mère de spores (1 x 107 spores/mL) au centre d’une plaque de culture contenant du milieu Czapek-Dox (CD) avec 20 mM d’uridine (autoclavé à 121 °C pendant 20 min, tableau 1).
- Incuber à 30 °C pendant 7 jours pour favoriser la formation de spores.
- Ajouter 1,5 mL de solution de Tween 20 à 0,01 % (autoclavée à 121 °C pendant 20 min) dans la plaque de culture contenant les spores et mettre les spores en suspension en les grattant à l’aide d’un épandeur de cellules.
- Prélever la suspension de spores dans un tube de microcentrifugation stérile de 1,5 mL à l’aide d’une pipette.
- Stockez la suspension à 4 °C jusqu’à l’utilisation.
REMARQUE : Utilisez la suspension de spores dans la procédure suivante dans les 2 semaines. Pour faire un stock destiné à un stockage à long terme, ajoutez du glycérol à la suspension de spores jusqu’à une concentration finale de 15 % et conservez-le à -80 °C.
2. Pré-culture
- Ajouter 100 mL de milieu polypeptone-dextrine (tableau 2) dans un erlenmeyer de 500 mL et l’autoclaver à 121 °C pendant 20 min.
- Après avoir confirmé que la température du milieu est réduite, ajoutez la solution d’uridine, stérilisée en passant à travers un filtre d’une taille de pores de 0,22 μm, jusqu’à une concentration finale de 20 mM.
- Ajouter 200 μL de la suspension de spores (1 x 107 spores/mL) préparée à l’étape 1.4 et incuber à 30 °C en agitant à 120 tr/min pendant 36 h.
3. Préparation des protoplastes
- Récoltez la biomasse fongique préparée à l’étape 2.3 sur un filtre en verre avec une taille de pores de 30 μm (autoclavé à 121 °C pendant 20 min).
- Versez 100 mL d’eau distillée (autoclavée à 121 °C pendant 20 min) sur le filtre en verre et remuez plusieurs fois avec une spatule (autoclavée à 121 °C pendant 20 min).
- Après le lavage avec DW, versez 100 mL de tampon de chlorure de sodium (NaCl) (autoclavé à 121 °C pendant 20 min, tableau 3) sur le filtre en verre et remuez plusieurs fois avec une spatule.
- Transférez environ 1 à 2 mL (volume humide) des cellules dans un tube conique de 50 mL à l’aide d’une spatule (autoclavée à 121 °C pendant 20 min).
- Après avoir ajouté 15 mL de la solution enzymatique (stérilisée en passant à travers un filtre à pores de 0,22 μm, tableau 4) dans un tube conique de 50 mL, ajouter un tampon NaCl jusqu’à ce que le volume total atteigne 30 mL.
- Fermez le couvercle et fermez-le avec un film de paraffine. Incuber à 30 °C en secouant à 60 tr/min pendant 2 h.
- Faire passer la solution obtenue à travers une crépine de 70 μm fixée au tube conique de 50 mL pour éliminer le mycélium n’ayant pas réagi.
- Centrifuger la solution de protoplaste dans un tube conique de 50 mL à 2 150 x g et 4 °C pendant 20 min.
- Jetez délicatement le surnageant pour éviter que la pastille ne se déloge. Ajouter 1 mL de solution B stérilisée glacée (autoclavée à 121 °C pendant 20 min, tableau 5) et suspendre doucement les protoplastes précipités par pipetage.
- Transférez les protoplastes suspendus dans un tube stérile de 1,5 ml.
- Centrifuger à 2 220 x g et 4 °C pendant 5 min et retirer le surnageant. Ajoutez ensuite 1 mL de solution B glacée et suspendez doucement la pastille par pipetage.
- Répétez l’étape 3.9.
- À l’aide d’un hémocytomètre, mesurez le nombre de protoplastes au microscope.
- Préparez une suspension de protoplastes (1−3 x 107 protoplastes/mL) dans la solution B et conservez-la sur de la glace jusqu’à utilisation.
REMARQUE : Utilisez immédiatement la suspension protoplast selon la procédure suivante.
4. Introduction de l’ADN pour la production sécrétoire de protéines
- En utilisant un plasmide contenant une cassette d’ADN pour la production sécrétoire de protéines comme matrice, amplifiez le fragment d’ADN requis pour la production sécrétoire par PCR.
- Préparez l’échantillon d’ADN à utiliser pour la transformation en traitant le produit PCR à l’aide d’un kit de purification PCR.
- Ajouter 1 mL de la solution de protoplaste, 200 μL de la solution C (autoclavée à 121 °C pendant 20 min, tableau 6) et 50 μL de l’échantillon d’ADN (0,5 à 1 μg/μL) dans un tube conique stérile de 15 mL prérefroidi, et mélanger doucement la solution par pipetage.
REMARQUE : La solution C est très visqueuse et peut être difficile à manipuler ; Assurer un mélange complet par pipetage. - Incuber le tube sur de la glace pendant 30 min.
- Ajouter 1,5 mL de solution C, mélanger doucement par pipetage, puis laisser la solution à température ambiante pendant 20 à 30 min.
5. Criblage direct de la culture liquide
REMARQUE : Sélectionner le système de culture à l’aide d’erlenmeyers (section 5.1) ou de microplaques (section 5.2).
- Système de culture à l’aide d’un erlenmeyer de 200 mL
- Ajouter 50 mL de milieu de DP contenant 0,8 M de sorbitol dans un erlenmeyer de 200 mL et stériliser à l’autoclave à 121 °C pendant 20 min.
- Après avoir confirmé que la température du milieu est réduite, ajouter la suspension de protoplastes préparée à l’étape 4.5 à une concentration finale de 1 x 105 protoplastes/mL.
- Incuber la culture pendant 6 jours à 30 °C en secouant à 120 tr/min.
- Prélever 1 mL du milieu et le conserver dans un tube de 1,5 mL à 4 °C. Conservez l’échantillon de culture à -20 ° C, s’il n’est pas analysé immédiatement.
- Mélanger 20 μL d’échantillon de culture et 20 μL de tampon d’échantillon de dodécylsulfate de sodium (SDS) (tableau 7) dans un tube de 1,5 mL et faire bouillir à 95 °C pendant 5 min.
- Après refroidissement sur glace, chargez 20 μL de l’échantillon bouilli et 5 μL d’étalon de protéine précoloré sur un gel prémoulé et analysez la production sécrétoire de la protéine hétérologue cible par électrophorèse sur gel de dodécylsulfate de sodium-polyacrylamide (SDS-PAGE)10.
- Système de culture à l’aide d’une microplaque de 24 puits
- Ajouter la suspension de protoplaste préparée à l’étape 4.5 dans un milieu de DP stérilisé contenant 0,8 M de sorbitol jusqu’à une concentration finale de 1 x 105 protoplastes/mL, et mélanger délicatement par pipetage.
- Ajouter 1 mL de la solution préparée à l’étape 5.2.1 dans trois puits d’une microplaque de 24 puits.
- Fixez le couvercle et incubez à 30 °C en secouant à 175 tr/min pendant 10 jours.
- Prélever 1 mL de milieu et stocker dans un tube de 1,5 mL à 4 ° C. Stocker l’échantillon de culture à -20 ° C, s’il n’est pas analysé immédiatement.
- Mélanger 20 μL de l’échantillon de culture et 20 μL de tampon d’échantillon SDS dans un tube de 1,5 mL et faire bouillir à 95 °C pendant 5 min.
- Après refroidissement sur glace, chargez 20 μL de l’échantillon bouilli et 5 μL d’étalon protéique précoloré sur un gel prémoulé et analysez la production sécrétoire de la protéine hétérologue cible par SDS-PAGE10.
Résultats
Les résultats de l’introduction de la cassette d’expression de l’ADN codant pour Talaromyces cellulolyticus cellobiohydrolase (CBH : numéro d’enregistrement GenBank E39854) dans un uridine auxotrophe de la souche HO422 d’A. oryzae et du criblage de la production sécrétoire de la protéine hétérologue sont décrits ci-dessous.
Préparation de la suspension de spores fraîches
Discussion
Nous avons développé un système qui permet le criblage des transformants d’A. oryzae plus rapidement que la méthode conventionnelle, en réalisant une culture liquide des protoplastes. L’aspect le plus critique de cette méthode est la pression osmotique du milieu liquide. La pression osmotique adaptée à la culture liquide a été optimisée à l’aide de sorbitol. La croissance de la souche HO4 d’A. oryzae a été la plus active en présence de 0,8 M de so...
Déclarations de divulgation
Les auteurs n’ont rien à divulguer.
Remerciements
Les auteurs remercient Rinkei Okano pour son aide dans les expériences. Les auteurs remercient le professeur Katsuya Gomi de l’Université de Tohoku et le professeur Masayuki Machida de l’Institut de technologie de Kanazawa pour leurs précieuses discussions. Ce travail a été financé par Honda Research Institute Japan Co., Ltd.
matériels
| Name | Company | Catalog Number | Comments |
| 1 M NaOH | NACALAI TESQUE, INC. | 37421-05 | |
| 1 M Tris-HCl (pH 7.5) | FUJIFILM Wako Pure Chemical Corporation | 318-90225 | |
| 1 M Tris-HCl (pH 6.8) | FUJIFILM Wako Pure Chemical Corporation | 2106-100 | |
| 1.5-mL Microcentrifuge tube | AS ONE Corporation | 1-1600-03 | |
| 15-mL Conical centrifuge tube | Becton, Dickinson and Company | 352196 | |
| 2-mercaptoethanol | Bio-Rad Laboratories, Inc. | 1610710 | |
| 24-well Microplate | AGC TECHNO GLASS CO., LTD. | 3820-024 | |
| 50-mL Conical centrifuge tube | Becton, Dickinson and Company | 352070 | |
| 70-µm Cell strainer | Becton, Dickinson and Company | 352350 | |
| Agar | FUJIFILM Wako Pure Chemical Corporation | 010-15815 | |
| Autoclave | TOMY SEIKO CO.,LTD. | LSX-700 | |
| Bromophenol blue | FUJIFILM Wako Pure Chemical Corporation | 021-02911 | |
| CaCl2 | FUJIFILM Wako Pure Chemical Corporation | 038-24985 | |
| Casamino acid | FUJIFILM Wako Pure Chemical Corporation | 393-02145 | |
| Cellulase R-10 | Cosmo Bio Co., Ltd. | 16419 | |
| Dextrin hydrate | FUJIFILM Wako Pure Chemical Corporation | 044-00585 | |
| D-Sorbitol | FUJIFILM Wako Pure Chemical Corporation | 191-14735 | |
| e-PAGEL | ATTO CORPORATION | E-T/R1020L | Used for precast gel in SDS-PAGE |
| Electrophoresis device | ATTO CORPORATION | WSE-1150 | Used for SDS-PAGE |
| FeSO4·7H2O | FUJIFILM Wako Pure Chemical Corporation | 098-01085 | |
| Glass filter 17G3 | Tokyo Garasu Kikai Co., Ltd. | 0000094147 | |
| Glycerol | FUJIFILM Wako Pure Chemical Corporation | 070-04941 | |
| Hemocytometer | Funakoshi Co., Ltd. | 521-10 | |
| High speed refrigerated centrifuge | KUBOTA CORPORATION | 7780 | Used for centrifugation of samples in 50-mL conical centrifuge tubes |
| Incubator | TAITEC CORPORATION. | G·BR-200 | Used for flask liquid culture and preparation of protoplasts |
| Incubator | TAITEC CORPORATION. | BR-43FL | Used for microplate liquid culture and plate culture |
| KCl | FUJIFILM Wako Pure Chemical Corporation | 163-03545 | |
| KH2PO4 | NACALAI TESQUE, INC. | 28721-55 | |
| Lysing enzyme | Sigma-Aldrich | L1412-10G | |
| MgSO4·7H2O | FUJIFILM Wako Pure Chemical Corporation | 131-00405 | |
| Micro refrigerated centrifuge | KUBOTA CORPORATION | 3740 | Used for centrifugation of samples in 1.5-mL microcentrifuge tubes |
| Microscope | Leica Microsystems | DMI6000 B | |
| NaCl | FUJIFILM Wako Pure Chemical Corporation | 190-13921 | |
| NaH2PO4·2H2O | NACALAI TESQUE, INC. | 31718-15 | |
| NaNO3 | FUJIFILM Wako Pure Chemical Corporation | 195-02545 | |
| Parafilm M | Bemis Company, Inc | PM-996 | |
| PCR Purification Kit | QIAGEN K.K | 28104 | |
| Petri dish | Sumitomo Bakelite Co., Ltd. | MS-11900 | Used as culture plate |
| Polyethylene glycol | Sigma-Aldrich | P3640-500G | |
| Polypeptone peptone | Becton, Dickinson and Company | 211910 | |
| Protein ladders | Bio-Rad Laboratories, Inc. | 161-0377 | Used as molecular weight marker in SDS-PAGE |
| Sodium dodecyl sulfate | Bio-Rad Laboratories, Inc. | 1610301 | |
| Sterile filter | Merck KGaA | SLGP033RB | |
| Sucrose | NACALAI TESQUE, INC. | 30404-45 | |
| Tween 20 | Tokyo Chemical Industry Co., Ltd. | T0543 | |
| Uridine | Sigma-Aldrich | U3750-25G | |
| Yatalase | Takara Bio Inc. | T017 |
Références
- Tsuboi, H., et al. Improvement of the Aspergillus oryzae enolase promoter (P-enoA) by the introduction of cis-element repeats. Bioscience, Biotechnology, and Biochemistry. 69, 206-208 (2005).
- Nemoto, T., Maruyama, J. -. I., Kitamoto, K. Contribution ratios of amyA, amyB, amyC genes to high-level α-amylase expression in Aspergillus oryzae. Bioscience, Biotechnology, and Biochemistry. 76, 1477-1483 (2012).
- Shinkawa, S., Mitsuzawa, S. Feasibility study of on-site solid-state enzyme production by Aspergillus oryzae. Biotechnology for Biofuels. 13, 31 (2020).
- Machida, M., et al. Genome sequencing and analysis of Aspergillus oryzae. Nature. 438 (7071), 1157-1161 (2005).
- Veenstra, A. E., van Solingen, P., Bovenberg, R. A. L., vander Voort, L. H. M. Strain improvement of Penicillium chrysogenum by recombinant DNA techniques. Journal of Biotechnology. 17 (1), 81-90 (1991).
- Takahashi, T., Masuda, T., Koyama, Y. Enhanced gene targeting frequency in ku70 and ku80 disruption mutants of Aspergillus sojae and Aspergillus oryzae. Molecular Genetics and Genomics. 275 (5), 460-470 (2006).
- Mizutani, O., et al. A defect of LigD (human Lig4 homolog) for nonhomologous end joining significantly improves efficiency of gene-targeting in Aspergillus oryzae. Fungal Genetics and Biology. 45 (6), 878-889 (2008).
- Bergès, T., Barreau, C. Isolation of uridine auxotrophs from Trichoderma reesei and efficient transformation with the cloned ura3 and ura5 genes. Current Genetics. 19 (5), 359-365 (1991).
- Takeno, S., et al. Cloning and sequencing of the ura3 and ura5 genes, and isolation and characterization of uracil auxotrophs of the fungus Mortierella alpina 1S-4. Bioscience, Biotechnology, and Biochemistry. 68 (2), 277-285 (2004).
- Ji, Y. W., et al. Application of membrane filtration method to isolate uninuclei conidium in Aspergillus oryzae transformation system based on the pyrG marker. Food Science and Biotechnology. 22 (1), 93-97 (2013).
- Mitsuzawa, S., Fukuura, M., Shinkawa, S., Kimura, K., Furuta, T. Alanine substitution in cellobiohydrolase provides new insights into substrate threading. Scientific Reports. 7 (1), 16320 (2017).
- Fleissner, A., Dersch, P. Expression and export: recombinant protein production systems for Aspergillus. Applied Microbiology and Biotechnology. 87, 1255-1270 (2010).
- Yoon, J., Maruyama, J. -. I., Kitamoto, K. Disruption of ten protease genes in the filamentous fungus Aspergillus oryzae highly improves production of heterologous proteins. Applied Microbiology and Biotechnology. 89, 747-759 (2011).
- Lin, H., et al. Engineering Aspergillus oryzae A-4 through the chromosomal insertion of foreign cellulase expression cassette to improve conversion of cellulosic biomass into lipids. PLoS One. 9, 108442 (2014).
- Chakraborty, B. N., Patterson, N. A., Kapoor, M. An electroporation-based system for high-efficiency transformation of germinated conidia of filamentous fungi. Canadian Journal of Microbiology. 37 (11), 858-863 (1991).
- Sun, Y., et al. A dual selection marker transformation system using Agrobacterium tumefaciens for the industrial Aspergillus oryzae 3.042. Journal of Microbiology and Biotechnology. 29 (2), 230-234 (2019).
- Gomi, K., Iimura, Y., Hara, S. Integrative transformation of Aspergillus oryzae with a plasmid containing the Aspergillus nidulans argB Gene. Agricultural and Biological Chemistry. 51 (9), 2549-2555 (1987).
- Case, M. E., Schweizer, M., Kushner, S. R., Giles, N. H. Efficient transformation of Neurospora crassa by utilizing hybrid plasmid DNA. Proceedings of the National Academy of Sciences of the United States of America. 76 (10), 5259-5263 (1979).
- Lim, F. Y., Sanchez, J. F., Wang, C. C., Keller, N. P. Toward awakening cryptic secondary metabolite gene clusters in filamentous fungi. Methods in Enzymology. 517, 303-324 (2012).
- Li, D., Tang, Y., Lin, J., Cai, W. Methods for genetic transformation of filamentous fungi. Microbial Cell Factories. 16 (1), 168 (2017).
- Chung, C. T., Niemela, S. L., Miller, R. H. One-step preparation of competent Escherichia coli: transformation and storage of bacterial cells in the same solution. Proceedings of the National Academy of Sciences of the United States of America. 86 (7), 2172-2175 (1989).
- Shinkawa, S., et al. Base sequence for protein expression and method for producing protein using same. U.S. Patent. , (2018).
- Tamano, K., et al. The β-1, 3-exoglucanase gene exgA (exg1) of Aspergillus oryzae is required to catabolize extracellular glucan, and is induced in growth on a solid surface. Bioscience, Biotechnology, and Biochemistry. 71 (4), 926-934 (2007).
- Endo, Y., et al. Novel promoter sequence required for inductive expression of the Aspergillus nidulans endoglucanase gene eglA. Bioscience, Biotechnology, and Biochemistry. 72 (2), 312-320 (2008).
- Kubodera, T., Yamashita, N., Nishimura, A. Pyrithiamine resistance gene (ptrA) of Aspergillus oryzae: cloning, characterization and application as a dominant selectable marker for transformation. Bioscience, Biotechnology, and Biochemistry. 64 (7), 1416-1421 (2000).
- Yamada, O., Lee, B. R., Gomi, K. Transformation system for Aspergillus oryzae with double auxotrophic mutations, niaD and sC. Bioscience, Biotechnology, and Biochemistry. 61 (8), 1367-1369 (1997).
- Nødvig, C. S., Nielsen, J. B., Kogle, M. E., Mortensen, U. H. A CRISPR-Cas9 system for genetic engineering of filamentous fungi. PLoS One. 10 (7), 0133085 (2015).
- Shi, T. Q., et al. CRISPR/Cas9-based genome editing of the filamentous fungi: the state of the art. Applied Microbiology and Biotechnology. 101 (20), 7435-7443 (2017).
- Katayama, T., et al. Forced recycling of an AMA1-based genome-editing plasmid allows for efficient multiple gene deletion/integration in the industrial filamentous fungus Aspergillus oryzae. Applied Environmental Microbiology. 85 (3), 01896-01918 (2019).
Réimpressions et Autorisations
Demande d’autorisation pour utiliser le texte ou les figures de cet article JoVE
Demande d’autorisationThis article has been published
Video Coming Soon
À PROPOS DE JoVE
Copyright © 2025 MyJoVE Corporation. Tous droits réservés.