
Method Article
Modèle de plaie ex vivo humaine et approche de coloration à montage entier pour évaluer avec précision la réparation de la peau
Dans cet article
Résumé
Nous démontrons ici une technique optimisée pour évaluer la réparation des plaies à l’aide de peau humaine ex vivo combinée à une approche de coloration à montage complet. Cette méthodologie fournit une plate-forme préclinique pour l’évaluation des thérapies potentielles des plaies.
Résumé
Les plaies chroniques non cicatrisantes, qui touchent principalement les personnes âgées et les diabétiques, constituent un domaine important des besoins cliniques non satisfaits. Malheureusement, les traitements actuels des plaies chroniques sont inadéquats, tandis que les modèles précliniques disponibles prédisent mal l’efficacité clinique des nouvelles thérapies. Nous décrivons ici un modèle préclinique à haut débit pour évaluer de multiples aspects de la réponse de réparation de la peau humaine. Des plaies d’épaisseur partielle ont été créées dans la peau ex vivo humaine et cultivées tout au long d’un temps de guérison. Des biopsies de plaies cutanées ont été prélevées en fixateur pour la procédure de coloration complète. Les échantillons fixes ont été bloqués et incubés dans des anticorps primaires, avec une détection réalisée par anticorps secondaire conjugué par fluorescence. Les plaies ont été contre-colorées et imagées par microscopie confocale avant de calculer le pourcentage de fermeture de la plaie (ré-épithélialisation) dans chaque biopsie. En appliquant ce protocole, nous révélons que les plaies excisionnelles de 2 mm créées dans une peau de donneur saine sont entièrement réépithélialisées au jour 4-5 après la blessure. Au contraire, les taux de fermeture des plaies cutanées diabétiques sont considérablement réduits, accompagnés d’une reformation perturbée de la barrière. La combinaison de la plaie de la peau humaine avec une nouvelle approche de coloration à montage entier permet une méthode rapide et reproductible pour quantifier la réparation ex vivo de la plaie. Collectivement, ce protocole fournit une plate-forme humaine précieuse pour évaluer l’efficacité des thérapies potentielles des plaies, transformant les tests précliniques et la validation.
Introduction
Les plaies chroniques et non cicatrisantes, qui sont très répandues chez les personnes âgées et les diabétiques, sont un domaine très peu apprécié des besoins cliniques non satisfaits. Ces blessures représentent un fardeau physique et psychologique majeur pour les patients et coûtent aux prestataires de soins de santé des milliards chaque année à traiter1. Malgré une meilleure compréhension de la biologie des plaies et des progrès technologiques, jusqu’à 40 % des plaies chroniques ne guérissent toujours pas après les meilleurs soins standard2. Ainsi, 14 à 26% des patients atteints d’ulcères du pied diabétique nécessitent par la suite une amputation3, tandis que le taux de mortalité post-amputation à 5 ans s’élève à environ 70%4. En conséquence, il est urgent de développer de nouvelles thérapies efficaces pour améliorer la qualité de vie des patients tout en réduisant le fardeau substantiel des soins de santé imposé par une mauvaise cicatrisation des plaies. Les modèles précliniques mal prédictifs restent un obstacle important au développement de nouvelles thérapies efficaces.
La réparation des plaies est un processus dynamique et multiforme impliquant un large éventail de types de cellules, d’innombrables niveaux de communication et un environnement tissulaire remodelé temporellement. La cicatrisation de la peau est soutenue par quatre grandes étapes réparatrices: l’hémostase, l’inflammation, la prolifération et le remodelage de la matrice. Ces étapes agissent finalement pour prévenir la perte de sang et l’infection, fermer la surface de la plaie (un processus appelé ré-épithélialisation) et ramener la peau à un état non blessé5. Les plaies chroniques sont associées à une étiologie diversifiée et à une perturbation généralisée des processus de guérison6,ce qui complique encore l’identification des cibles thérapeutiques. Néanmoins, un large éventail de modèles ont été développés pour élucider les facteurs moléculaires et cellulaires de la pathologie des plaies et tester de nouvelles approches thérapeutiques7.
Le modèle de réparation de plaie le plus utilisé est la blessure aiguë chez la souris. Les souris sont très faciles à traiter pour les études mécanistes et fournissent des modèles validés du vieillissement et du diabète8. Malgré les similitudes générales montrées entre la guérison de la souris et celle de l’homme, des différences entre les espèces dans la structure de la peau et la dynamique de guérison demeurent. Cela signifie que la plupart des recherches sur les plaies murines ne se traduisent pas facilement à la clinique9. Par conséquent, il y a eu une poussée vers des systèmes humains in vitro et ex vivo avec une applicabilité et une translatabilitéélevées 10,11.
Ici, nous fournissons un protocole approfondi pour effectuer des plaies excisionnelles d’épaisseur partielle dans la peau humaine ex vivo. Nous décrivons également notre approche de coloration à montage complet comme une méthode hautement reproductible d’évaluation ex vivo de la cicatrisation de la peau humaine. Nous montrons la trajectoire de la réparation épidermique (ré-épithélialisation) et la formation ultérieure d’une barrière, en évaluant le taux de fermeture des plaies dans la peau humaine saine par rapport à la peau humaine diabétique. Enfin, nous démontrons comment la coloration à montage entier peut être adaptée pour une utilisation avec une gamme d’anticorps afin d’évaluer divers aspects de la réponse de guérison.
Protocole
La peau humaine a été obtenue auprès de patients subissant une chirurgie reconstructive à l’hôpital Castle Hill et à l’infirmerie royale de Hull (Hull, Royaume-Uni) avec le consentement écrit et pleinement éclairé du patient, les directives institutionnelles et l’approbation éthique (LRECs: 17/SC/0220 et 19/NE/0150). La peau non diabétique a été prélevée chez des patients subissant une chirurgie de routine (âge moyen = 68 ans). La peau diabétique a été sélectionnée parmi des donneurs qui avaient établi un diabète de type II et des antécédents d’ulcération (âge moyen = 81 ans). Les échantillons de la chirurgie ont été transportés dans des milieux de détention et traités immédiatement à leur arrivée au laboratoire. Toutes les étapes expérimentales utilisant des tissus humains non fixés ont été réalisées au niveau de biosécurité 2 (BSL-2) dans une armoire de biosécurité à flux laminaire de classe II.
1. Préparation des milieux de culture cutanés et des réactifs de coloration
REMARQUE: Tous les détails du réactif et du consommable sont fournis dans le tableau des matériaux. S’assurer que tous les réactifs et l’équipement utilisés pour le traitement et la culture des tissus humains sont stériles. Stériliser les instruments avant l’utilisation et décontaminer avec un désinfectant après contact avec le tissu. Décontaminer les déchets dans un désinfectant à 1 % avant leur élimination.
- Milieu de détention : Complétez le milieu Eagle modifié (DMEM) de Dulbecco à haute teneur en glucose avec 2 mM de L-glutamine et une solution antibiotique-antimycotique à 4 % (v/v).
- Solution saline équilibrée (HBSS) de Hank avec antibiotiques : Ajouter une solution antibiotique-antimycotique à 4 % (v/v) à l’HBSS. Conserver à 4 °C jusqu’à utilisation.
- Solution saline tamponnée au phosphate (DPBS) de Dulbecco : Préparer le DPBS en dissolvant 9,6 g de poudre de DPBS par litre d’eau distillée (dH2O). Autoclave pour stériliser et conserver à 4 °C jusqu’à utilisation.
- Milieux de croissance de la peau humaine: Complétez le DMEM à haute teneur en glucose avec 2 mM de L-glutamine, une solution antibiotique-antimycotique à 1% (v / v) et du sérum bovin fœtal à 10% (v / v). Conserver à 4 °C jusqu’à utilisation.
- Fixateur cutané : À 450 mL dedH2O, ajouter 40 mL de solution de formaldéhyde, 10 mL d’acide acétique glacial, 4,5 g de chlorure de sodium et 0,25 g de bromure d’alkyltriméthylammonium. Conserver à température ambiante (RT) et utiliser en quelques jours.
ATTENTION : Le fixateur est dangereux (irritant et inflammable). Manipuler avec soin et éliminer par un itinéraire approprié. - Solution saline tamponnée au phosphate (PBS) : Préparer le PBS pour la coloration complète en ajoutant 6 g de chlorure de sodium à 100 mL de solution tampon de phosphate et 900 mL de dH2O.
- Tampon de lavage de coloration: Dissoudre 0,5% (v / v) Triton X-100 dans PBS.
- Tampon bloquant : Ajouter 0,2 % (p/v) d’azide de sodium et 2 % (v/v) de sérum animal au tampon de lavage de coloration. Conserver à 4 °C jusqu’à deux semaines.
REMARQUE: Blocage dans le sérum de l’espèce hôte d’anticorps secondaires. L’azide de sodium empêchera la croissance bactérienne pendant l’incubation. - Solution de travail DAPI : Préparer un stock de 5 mg/mL de 4′,6-diamidino-2-phénylindole (DAPI) dans du diméthylsulfoxyde. Diluer le bouillon 1:1 000 dans un tampon de lavage de coloration pour donner une solution de travail DAPI de 5 μg/mL.
- Bloc de peroxydase : Ajouter 0,3 % (v/v) de peroxyde d’hydrogène au tampon de lavage de coloration. Conserver à 4 °C jusqu’à utilisation. Restez dans l’obscurité pour éviter la décomposition.
- Kit ABC-HRP :
- Anticorps secondaire conjugué HRP : 1 goutte d’IgG anti-chèvre de lapin biotinylé dans 5 mL de tampon de coloration. Conserver à 4 °C jusqu’à deux semaines.
REMARQUE: Le kit / secondaire utilisé dépendra de l’espèce hôte de l’anticorps primaire. - Réactif du complexe avidine-biotine (ABC) : 2 gouttes de réactif A et 2 gouttes de réactif B dans 5 mL de tampon de lavage de coloration. Le réactif ABC doit être préparé au moins 30 minutes avant utilisation. Conserver à 4 °C jusqu’à deux semaines.
- Anticorps secondaire conjugué HRP : 1 goutte d’IgG anti-chèvre de lapin biotinylé dans 5 mL de tampon de coloration. Conserver à 4 °C jusqu’à deux semaines.
- Substrat de peroxydase: 3 gouttes de réactif 1, 2 gouttes de réactif 2, 2 gouttes de réactif 3 et 2 gouttes de peroxyde d’hydrogène dans 5 mL de dH2O. Le substrat de peroxydase doit être fraîchement préparé immédiatement avant utilisation et ne peut pas être stocké.
2. Préparation de la peau pour les plaies
REMARQUE : Ces étapes doivent être effectuées dans une armoire de biosécurité à flux laminaire de classe II.
- Recueillir la peau dans un support de maintien et le transporter jusqu’à l’armoire BSL-2.
- Placez le derme cutané vers le bas dans une boîte de Petri stérile de 90 mm et retirez le tissu adipeux avec des ciseaux stériles.
- Placer la peau dans un tube de 50 mL contenant 25 mL de HBSS avec des antibiotiques pendant 10 min à TA. Agiter par intermittence pour éliminer tout sang résiduel et tissu adipeux.
- Répétez l’étape 2.3 à l’aide d’un nouveau tube de 50 mL.
- Placer la peau dans un tube frais de 50 mL contenant 25 mL de HBSS, cette fois sans antibiotiques pendant 10 min à RT. Agiter comme à l’étape 2.3.
- Effectuez un rinçage final de la peau en plaçant la peau dans un nouveau tube avec 25 mL de DPBS. La peau est maintenant prête à se blesser.
3. Créer des plaies cutanées humaines ex vivo
REMARQUE : Ces étapes doivent être effectuées dans une armoire de biosécurité à flux laminaire de classe II.
- Préparez les plats de culture de la peau avant de blesser. Dans une boîte de Petri de 60 mm, empilez deux tampons absorbants stériles et ajoutez 4 mL de milieu de peau humaine sur le côté du plat. Placez une membrane filtrante stérile en nylon sur la pile de tampons absorbants.
REMARQUE: Les milieux cutanés peuvent être modifiés en fonction des conditions de traitement requises. Jusqu’à trois explants de plaies peuvent être cultivés sur chaque pile. - Sécher le côté dermique de la peau sur de la gaze stérile dans une boîte de Petri de 90 mm pour éliminer le DPBS résiduel.
REMARQUE: Cela empêche la peau de glisser lors de la blessure. - Placez le derme de la peau vers le bas sur un couvercle de boîte de Petri propre de 90 mm et tamponnez l’épiderme avec de la gaze stérile fraîche.
REMARQUE: Il est plus facile d’enrouler la peau dans un couvercle de boîte de Petri que la base. Les travaux ultérieurs doivent être effectués rapidement pour éviter que la peau ne se dessèche. - En maintenant la peau tendue, appuyez sur un coup de biopsie de 2 mm contre la peau et tordez doucement. Ne frappez pas entièrement à travers la peau.
REMARQUE: Les plaies d’épaisseur partielle sont conçues pour percer l’épiderme et partiellement dans le derme. Il peut y avoir une variabilité de donneur à donneur et de site à site dans la force requise pour créer la plaie d’épaisseur partielle. - Utilisez des pinces à tissu denté incurvées pour ramasser chaque côté de la plaie de 2 mm et accrochez des ciseaux à iris incurvés sous la plaie de 2 mm pour la découper uniformément.
- Biopsie autour de la plaie centrale de 2 mm à l’aide d’un poinçon de biopsie de 6 mm pour créer une explantation de 6 mm avec une plaie d’épaisseur partielle de 2 mm au centre.
REMARQUE: Un poinçon de biopsie de 6 mm peut être utilisé pour marquer la peau afin de marquer l’endroit où chaque plaie de 2 mm devrait être. Veillez à ne pas percer complètement le tissu. Créez des explantations de plaies dans un motif en nid d’abeille pour réduire le gaspillage. - Placez l’épiderme des explantations de plaie sur la pile de membranes filtrantes en nylon (préparée à l’étape 3.1).
REMARQUE: Lorsque vous manipulez des explantations de plaies, veillez à ne pas endommager la plaie centrale. Utilisez de petites pinces et ramassez chaque explante sur les côtés opposés. - Incuber les plaies à 32-37 °C et 5% de CO2 dans une atmosphère humidifiée (90-95%) pendant 1-7 jours. Remplacez le support tous les 2-3 jours.
4. Coloration complète des plaies ex vivo
REMARQUE: Cette section décrit les méthodes de coloration par immunofluorescence et immunoperoxydase. Bien mélanger tous les réactifs avant utilisation.
- Méthode de coloration fluorescente
- Prélever les explantations de plaies dans des tubes de microcentrifugation de 1,5 mL contenant 500 μL de fixateur cutané et incuber à 4 °C pendant la nuit.
REMARQUE: Le fixateur utilisé dans ce protocole fonctionne bien pour les anticorps décrits. L’optimisation sera nécessaire pour d’autres anticorps. La fixation tissulaire plus de 24 heures peut entraîner une surfixe. - Le lendemain, retirez le fixateur et remplacez-le par 1 mL de tampon de lavage de coloration. Les biopsies peuvent être stockées dans un tampon de lavage de coloration à 4 °C jusqu’à 2 semaines avant la coloration.
REMARQUE: Pour toutes les étapes du tampon de lavage, utilisez une pipette sérologique ou une pointe de pipette, en prenant soin de ne pas endommager la plaie. - Aspirer le tampon de lavage de coloration et effectuer un rinçage de plus avec 1 mL de tampon de lavage de coloration.
- Calculer la quantité de tampon de blocage requise pour les étapes 4.1.5 à 4.1.6 (nombre d’échantillons x 300 μL = quantité de tampon de blocage en μL). Créez un tampon supplémentaire si nécessaire.
- Ajouter 150 μL de tampon bloquant à chaque échantillon et incuber pendant 1 h à TA. Pour toutes les étapes de coloration, assurez-vous que chaque échantillon est suffisamment couvert et qu’il n’y a pas de bulles recouvrant la surface de la plaie de biopsie.
REMARQUE: Cette étape peut être effectuée dans des tubes de microcentrifugation de 1,5 mL ou dans une plaque de 48 puits. Si vous utilisez une plaque de 48 puits, incuber les plaies face vers le bas dans chaque puits. - Diluer l’anticorps primaire dans le tampon bloquant restant.
REMARQUE: La kératine anti-souris 14 (K14) diluée 1: 1 000 dans le tampon de blocage fonctionne bien. Optimisez cette étape pour une utilisation avec d’autres anticorps ou plusieurs sondes. - Aspirer le tampon bloquant et ajouter 150 μL d’anticorps primaire par puits/tube de microcentrifugation. Incuber les explantations de plaies dans un anticorps primaire à 4 °C pendant la nuit.
- Le lendemain, aspirer l’anticorps primaire et rincer dans un tampon de lavage contenant 0,2 % d’azide de sodium pendant 1 h à RT (500 μL par échantillon).
- Effectuez trois autres étapes de rinçage à l’aide d’un tampon de lavage de coloration (30 min par lavage, 500 μL par échantillon).
- Diluer l’anticorps secondaire conjugué par fluorescence dans un tampon de lavage de coloration (p. ex., chèvre anti-souris 488 à une dilution de 1:400).
- Calculer la quantité requise d’anticorps secondaires (nombre d’échantillons x 150 μL = quantité en μL).
- Ajouter 150 μL d’anticorps secondaire à chaque puits/tube de microcentrifugation. Incuber pendant 1 h à TA. Effectuer les étapes d’incubation 4.1.10 - 4.1.16 dans l’obscurité car l’anticorps secondaire est sensible à la lumière.
REMARQUE: Cette étape peut être effectuée à 4 ° C pendant la nuit si nécessaire. Optimiser la concentration d’anticorps secondaires nécessaire pour un signal adéquat et une coloration de fond limitée. - Retirez l’anticorps secondaire et effectuez 3 rinçages de 3 x 30 minutes avec un tampon de lavage de coloration (500 μL par échantillon).
- Jetez le tampon de lavage restant et calculez la quantité de solution de travail DAPI requise (conformément à l’étape 4.1.11).
- Contre-maintenir chaque explante avec 150 μL de solution de travail DAPI pendant 10 min à RT.
REMARQUE: DAPI va tacher les noyaux cellulaires en bleu. Le colorant Hoechst peut être utilisé comme alternative au DAPI. - Effectuez deux lavages finaux de 30 minutes avec un tampon de lavage de coloration (500 μL par échantillon). Les biopsies peuvent être stockées dans un tampon de lavage de coloration à 4 °C dans l’obscurité jusqu’à deux semaines avant l’imagerie.
- Prélever les explantations de plaies dans des tubes de microcentrifugation de 1,5 mL contenant 500 μL de fixateur cutané et incuber à 4 °C pendant la nuit.
- Méthode de coloration en champ clair.
- Effectuez les étapes 4.1.1 à 4.1.3.
- Étancher l’activité endogène de la peroxydase avec un bloc de peroxydase à 4 °C pendant la nuit.
REMARQUE: Cette étape est importante lors de l’utilisation d’un anticorps conjugué HRP pour réduire la coloration de fond non spécifique du tissu. Les tissus hautement vascularisés contiendront plus d’activité peroxydase endogène. - Jeter le bloc de peroxydase et rincer deux fois pendant 30 min dans un tampon de lavage de coloration.
- Effectuez les étapes 4.1.4 à 4.1.8.
REMARQUE: Les lavages après l’étape 4.1.7 sont particulièrement importants pour éliminer l’azide de sodium des échantillons. Si l’azide de sodium n’est pas éliminé de manière adéquate, il inactivera le HRP et interférera avec la détection des taches. - Ajouter 150 μL d’anticorps secondaire conjugué HRP à chaque tube de puits/microcentrifugeuse et incuber pendant la nuit à 4 °C ou 1 h à TA.
- Retirez l’anticorps secondaire et effectuez 3 lavages de 30 minutes dans un tampon de lavage de coloration.
- Ajouter 150 μL de réactif ABC à chaque puits/tube de microcentrifugation et incuber pendant la nuit à 4 °C ou 1 heure à TA.
- Aspirer le réactif ABC et effectuer 3 lavages de 3 min dans un tampon de lavage de coloration.
- Ajouter 150 μL de substrat de peroxydase à une explante et déterminer le temps nécessaire pour détecter un changement de couleur notable.
REMARQUE: Choisissez un échantillon où une forte coloration est attendue. Dans ce cas, un anneau rouge pour montrer l’épiderme migrateur (K14). La 3,3'-diaminobenzidine-4, ou tout autre substrat chromogène approprié, peut être utilisé en remplacement de ce substrat de peroxydase. - Une fois qu’un changement de couleur est observé, retirez le substrat de peroxydase et remplacez-le par 1 mL de dH2O.
- Répétez la détection du substrat de peroxydase pour les autres explants, en incubation pendant la durée déterminée à l’étape 4.2.11.
- Rincer toutes les explantations avec 1 mL de dH2O pour éliminer le substrat résiduel de peroxydase. Bien que les explantes puissent être stockées jusqu’à une semaine à 4 °C avant l’imagerie, il est préférable de les imager dès que possible pour éviter la lixiviation du substrat de peroxydase dans le dH2O au fil du temps.
5. Imagerie et quantification
- Imagerie fluorescente
REMARQUE: L’imagerie fluorescente est réalisée à l’aide d’un microscope confocal à balayage laser. Cependant, un microscope fluorescent inversé peut être suffisant pour acquérir des images 2D afin de quantifier les taux de fermeture des plaies. Lors de la sélection des anticorps secondaires, assurez-vous que les fluorochromes choisis sont compatibles avec les spectres d’excitation et d’émission de l’équipement de microscopie disponible.- Utilisez un microscope confocal à balayage laser équipé d’un objectif 2,5x, 10x et 20x, d’un étage motorisé x-y-z, d’un appareil photo numérique et d’un logiciel d’acquisition. Allumez le détecteur de lumière transmise (TPMT) pour permettre une visualisation facile de chaque biopsie et pour permettre la mesure de la fermeture totale de la plaie. Alternativement, mesurez chaque plaie par microscopie à fond clair après l’imagerie par fluorescence.
- Placez une base de boîte de Petri de 60 mm sur la plate-forme d’imagerie et ajoutez une fine couche (environ 1 mL) de DPBS.
REMARQUE: Si trop de DPBS est utilisé, la biopsie se déplacera pendant l’imagerie. Vous pouvez également utiliser une plaque de 48 puits si un support de plaque est disponible. - Utilisez de petites pinces tissulaires pour transférer les explantes de plaies des puits / tubes de microcentrifugation vers la boîte de Petri contenant du DPBS. Placez la plaie de biopsie du côté vers le bas dans la boîte de Pétri.
- Utilisez l’oculaire et la lampe fluorescente pour localiser et concentrer la plaie. Si des bulles sont piégées sous l’échantillon dans le champ de vision, ramassez la plaie avec une pince à tissu et repositionnez-la.
- Configurez le logiciel d’imagerie, en assurant une taille de sténopé égale entre les canaux pour une confocalité optimale. Pour cela, vérifiez la valeur d’une unité aérée pour chaque canal et sélectionnez la valeur la plus grande. Sélectionnez la vitesse de numérisation, la qualité de l’image et la moyenne.
REMARQUE: Les fluorochromes des anticorps secondaires conjugués et la contre-tache choisie (par exemple, DAPI) dicteront les canaux requis. - Allumez le logiciel d’acquisition en direct et ajustez la puissance laser et le gain de chaque canal aux niveaux requis pour visualiser la coloration. Réduisez le bruit de fond en augmentant le décalage numérique.
- Placez la plaie au centre du plan d’imagerie.
REMARQUE: Si la plaie ne remplit pas toute l’image en raison de l’utilisation d’un objectif plus petit ou de la création d’une plaie plus grande, prenez un panneau d’images et assemblez-les ensemble (manuellement ou avec une fonction de mosaïque dans le logiciel d’imagerie approprié). - Acquérir des images des biopsies de la plaie. Utilisez les mêmes paramètres d’imagerie entre les explants.
REMARQUE: Des images de puissance supérieure permettront d’évaluer les structures tissulaires et l’expression et l’emplacement des marqueurs cellulaires. - Collectez les piles Z en série à travers la plaie, en particulier lorsque le tissu n’est pas complètement plat contre la boîte de Pétri. Utilisez un logiciel d’analyse pour réduire la pile Z en une seule image de projection d’intensité maximale.
- Imagerie en champ clair
REMARQUE: L’imagerie en champ clair des biopsies colorées par immunoperoxydase peut être effectuée de plusieurs façons.- Imagerie au microscope inversé : Préparez les explants de plaies pour l’imagerie en les plaçant dans une boîte de Petri/puits comme décrit aux étapes 5.1.2-5.1.3. Acquérez des images numériques sous éclairage en champ clair sur un microscope inversé équipé d’un appareil photo numérique. Assemblez plusieurs images si nécessaire.
- Imagerie au microscope numérique sans fil : Utilisez un microscope numérique sans fil connecté à un téléphone ou à un ordinateur portable pour obtenir des images de haute qualité de manière rentable. Placez les explantations enroulées sur certains tissus et retirez tout résidu de dH2O (ou tampon de lavage de coloration) de l’entreposage de l’échantillon. Placez l’explantation de la plaie au centre du champ de vision du microscope. Obtenez des images à l’aide de la caméra connectée.
- Quantification
REMARQUE: Le pourcentage de fermeture de la plaie peut être quantifié dans n’importe quel logiciel qui permet de dessiner et de mesurer à main levée des formes. ImageJ peut être utilisé pour effectuer la quantification comme suit :- Ouvrez l’image à quantifier dans le logiciel ImageJ.
- Utilisez l’outil de forme à main levée pour dessiner autour de l’extérieur de la plaie réétitélialisée où elle rencontre la peau normale. Appuyez sur M (ou Analyser | Mesure) pour acquérir une mesure de zone « extérieure ».
REMARQUE: La texture du tissu de la plaie ré-épithélialisée diffère de la peau normale. Les images n’ont pas besoin d’être mises à l’échelle avant ce type d’analyse. - Utilisez l’outil de forme à main levée pour dessiner autour de la zone ouverte de la plaie. C’est là que la plaie ouverte rencontre le bord intérieur du tissu ré-épithélialisant. Appuyez sur M (ou Analyser | Mesure) pour acquérir une mesure de la zone « intérieure ».
- Utilisez l’équation suivante pour déduire le pourcentage de rééthélialisation/fermeture de la plaie :
% de fermeture = (zone externe de la plaie - zone interne de la plaie) / (zone extérieure de la plaie) x 100
REMARQUE: La couverture en pourcentage de la surface des anticorps peut être déduite de la même manière (par exemple, K14) ou en pourcentage de la surface totale de la plaie. L’intensité en pourcentage peut également fournir des informations semi-quantitatives sur l’expression au niveau tissulaire des marqueurs d’intérêt, tandis que l’imagerie à haute puissance présente des données d’expression au niveau cellulaire.
Résultats
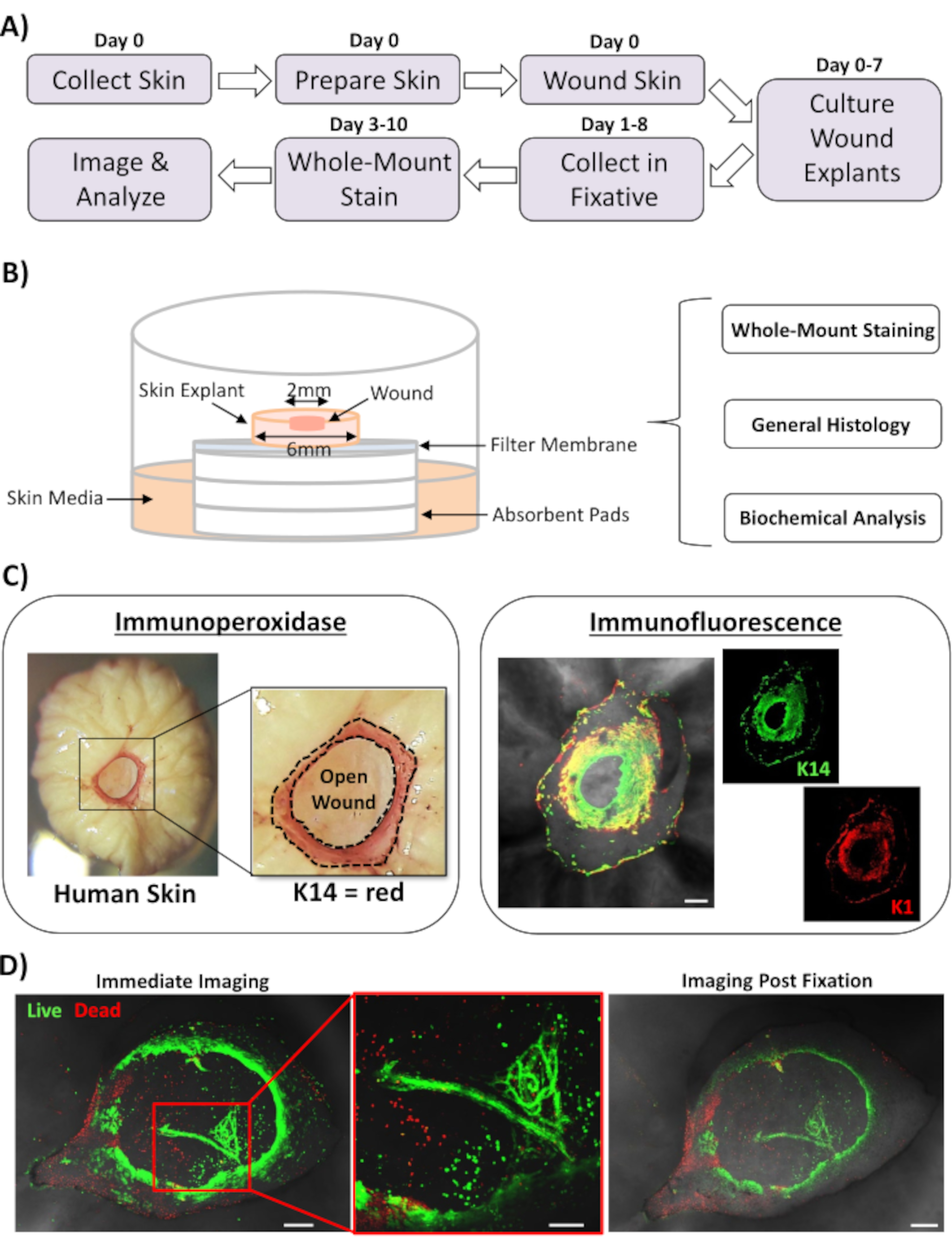
Dans ce rapport, nous présentons une nouvelle approche ex vivo de la plaie cutanée et de coloration complète pour évaluer les facteurs qui influencent la réponse de réparation de la peau humaine. La figure 1A montre un schéma du pipeline procédural, qui peut être effectué en 3 à 10 jours, en fonction des temps d’incubation de la plaie. Les plaies d’épaisseur partielle sont cultivées sur des piles de membranes à l’air : interface membranaire et peuvent être collectées pour une coloration complète, incorporées dans de la paraffine ou un milieu OCT pour l’histologie générale, ou congelées dans de l’azote liquide pour l’analyse biochimique (Figure 1B). Nous créons généralement des plaies d’épaisseur partielle de 2 mm au centre des explants de 6 mm. Cependant, la taille de la plaie et de l’explantation environnante peut être modifiée en fonction des besoins. La procédure de montage complet a été adaptée avec succès aux méthodes de coloration par immunoperoxydase et immunofluorescence (Figure 1C).
L’immunofluorescence permet de sonder les tissus avec de multiples anticorps. Pour cela, nous vous conseillons d’utiliser des anticorps primaires élevés chez différentes espèces et des anticorps secondaires conjugués par fluorescence appariés aux espèces pour limiter la réactivité inter-espèces. Les concentrations d’anticorps et les temps d’incubation devront être optimisés. Si une coloration de fond est observée, réduisez les concentrations d’anticorps, augmentez les étapes de lavage et ajoutez un tampon bloquant à l’anticorps secondaire. La viabilité des tissus frais peut être directement évaluée à l’avec des colorants de viabilité commerciale (voir tableau des matériaux). Nous montrons également que les tissus peuvent être fixés après la coloration de viabilité et imagés avec succès lorsqu’ils sont pratiquementappropriés (Figure 1D).

Figure 1: L’approche de la blessure ex vivo humaine et de la coloration à montage entier. (A) Pipeline illustrant le flux de travail procédural, de la collecte de la peau et de la blessure ex vivo à la coloration des tissus et à l’analyse des données. (B) Diagramme démontrant le système de culture de plaies cutanées ex vivo chez l’homme avec des analyses effectuées régulièrement sur le tissu. (C) La coloration à montage entier peut être utilisée à l’aide de techniques d’immunoperoxydase et d’immunofluorescence. K14 = kératine 14. (D) Les tissus vivants peuvent être colorés avec des colorants de viabilité commerciale et imagés avec succès après la fixation. Barre = 100 μm. Cette coloration a été réalisée sur une peau non diabétique. Veuillez cliquer ici pour voir une version agrandie de cette figure.
{kind=link}
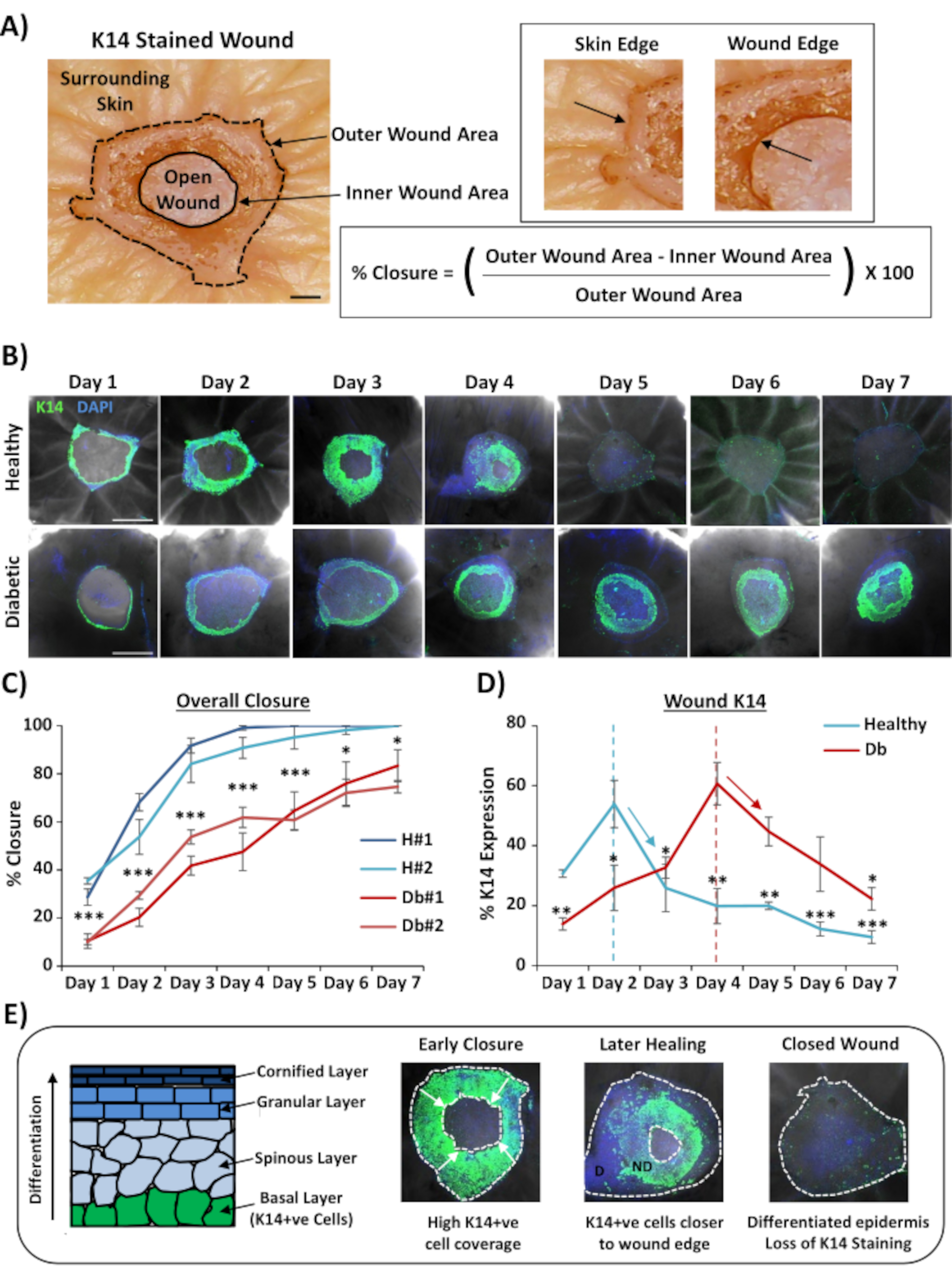
L’utilisation la plus largement applicable pour la coloration complète des plaies est de déterminer le taux de fermeture de la plaie d’une manière plus reproductible que celle qui peut être fournie par sectionnement histologique. Le pourcentage de fermeture a été quantifié en pourcentage de réétalisation de la surface de la plaie, comme le montre la figure 2A. La couverture en pourcentage de la surface de marqueurs spécifiques peut être mesurée à partir de la surface totale de la plaie ou en pourcentage de la plaie réétitélialisée. Nous avons caractérisé la cicatrisation dans une peau saine (non diabétique) par rapport à une peau diabétique sur une période de sept jours, en recueillant les plaies chaque jour après la blessure (images représentatives, Figure 2B). Les plaies cutanées saines se sont refermables au fil du temps comme prévu, avec une fermeture complète observée dans la plupart des échantillons aux jours 4-5. Au contraire, les plaies cutanées diabétiques ne se sont pas complètement refermables au cours de la période d’analyse de sept jours(figure 2C). Un retard significatif dans la fermeture de la plaie a été observé entre les plaies cutanées saines et diabétiques lors de la comparaison des taux de cicatrisation à chaque point temporel après la blessure(P < 0,001 au jour 6, P < 0,05 au jour 6 et P < 0,05 à P < 0,001 au jour 7).
Après évaluation des taux globaux de fermeture des plaies; nous avons mesuré le pourcentage de toute la zone de la plaie (zone externe sur la figure 2A)où les cellules K14 positives pouvaient être visualisées (coloration verte sur la figure 2B). Fait intéressant, nous avons observé que dans les plaies cutanées ex vivo saines, la coloration K14 a culminé au jour 2, puis a rapidement diminué (signification à chaque point temporel par rapport au pic du jour 2, Figure 2D). Cela reflète probablement la reformation de la barrière épidermique précoce, à l’exclusion de la pénétration des anticorps K14 à travers des couches épidermiques différenciées (voir la figure 2E schématique). Au cours du processus de réépithélialisation, les kératinocytes de la couche basale (K14 + ve) migrent vers l’intérieur sur la plaie ouverte, de sorte que l’épiderme plus proche du bord externe de la plaie se forme plus tôt que l’épiderme plus près du bord interne de la plaie (front migrateur). Alors que le bord avant de l’épiderme nouvellement formé continue de migrer pour fermer la plaie ouverte restante, l’épiderme du bord externe commence à se différencier pour reformer les autres couches épidermiques. Au début de la guérison, nous nous attendrions donc à voir la majeure partie de la zone réépithélialisée constituée de cellules basales (K14 + ve), tandis que dans la réparation ultérieure, la coloration K14 est perdue à mesure que l’épiderme se différencie de l’extérieur vers l’intérieur (voir les images de monture entière à la figure 2E). Par conséquent, la diminution de la coloration K14 montrée à la figure 2D (flèches vers le bas) est en corrélation avec une différenciation épidermique accrue. Fait intéressant, la coloration visible de K14 a culminé plus tôt dans les plaies saines (jour 2) par rapport aux plaies diabétiques (jour 4), démontrant ainsi que la rééchélialisation et la différenciation épidermique subséquente sont retardées dans les plaies cutanées diabétiques.

Figure 2: La coloration en montage entier révèle des taux decicatrisation perturbés chez les diabétiques par rapport aux peaux saines. (A) Méthode utilisée pour quantifier la fermeture des plaies à partir de mesures externes et internes des plaies. Des images en champ clair montrent une coloration à la kératine 14 (K14) en rouge. Bar = 300 μm. (B) Images représentatives de la guérison au fil du temps (jour après la blessure) dans une peau saine et diabétique. Bar = 500 μm. K14 = vert. DAPI = noyaux bleus. (C) Quantification des taux de fermeture des plaies (pourcentage de réétalialisation) montrant que les plaies ex vivo d’une peau saine se ferment significativement plus rapidement que les plaies ex vivo de peau diabétique. H = sain. Db = diabétique. (D) Le pourcentage de coloration K14 culmine plus tôt dans la peau saine par rapport à la peau diabétique, puis diminue en fonction de l’augmentation de la différenciation épidermique (flèches vers le bas). (E) K14 (cellule épidermique basale) coloration est perdue à mesure que l’épiderme se différencie. D = différencié. ND = non différencié. Des lignes pointillées blanches représentent les bords intérieurs et extérieurs des plaies. Flèches blanches = direction de migration. n = 6 plaies par donneur, par point temporel. Moyenne +/- SEM. * = P < 0,05, ** = P < 0,01 et *** = P < 0,001. En bonne santé et diabétiques comparés à chaque point de temps de guérison en C (valeurP pour la comparaison la moins significative). Changement temporel de la coloration K14 par rapport au pic pour chaque donneur en D. Veuillez cliquer ici pour voir une version agrandie de cette figure.
{kind=link}
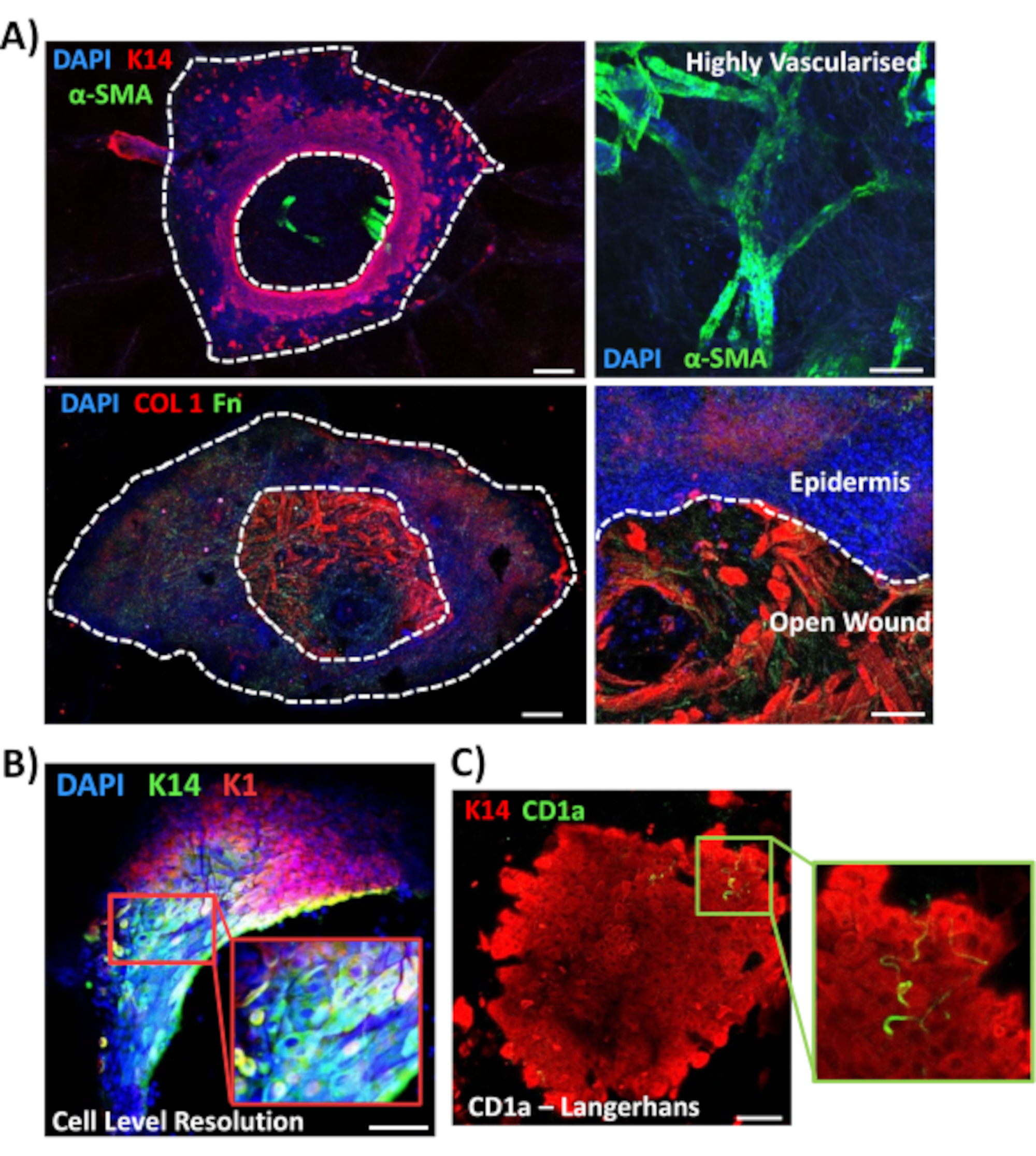
Nous avons ensuite utilisé la coloration à montage entier pour explorer l’expression tissulaire et la localisation d’autres marqueurs pertinents pour la plaie dans la peau non diabétique (Figure 3). Tous les anticorps utilisés et leurs concentrations de travail sont indiqués dans la Table des matériaux. Les vaisseaux sanguins de la plaie ouverte se sont colorés positivement avec l’anticorps alpha de l’actine du muscle lisse (a-SMA), utilisé en association avec K14 pour délimiter les bords épidermiques dans des images de puissance inférieure(Figure 3A). La matrice dermique a été colorée avec des anticorps contre le collagène de type I (COL 1) et la fibronectine (Fn). Ici, le collagène a été observé comme des fibres épaisses abondantes tandis que les fibres de fibronectine étaient clairsemées, ondulées et minces(Figure 3A). Notre approche de coloration globale est également capable de fournir une résolution de coloration au niveau cellulaire, comme démontré pour les kératinocytes K14 positifs(Figure 3B).
Enfin, nous montrons que les plaies ex vivo humaines possèdent des cellules immunitaires résidentes, avec des cellules de Langerhans détectées autour de l’épiderme nouvellement formé au jour 3 après la blessure (Figure 3C). En effet, ces résultats suggèrent que la coloration à montage entier peut être utilisée pour étudier les caractéristiques clés de la réponse de guérison, y compris l’inflammation, la prolifération et la matrice extracellulaire(Figure 4A). Prises ensemble, nos données révèlent que la procédure combinée de blessure cutanée ex vivo et de coloration complète est une méthode valide pour évaluer divers aspects de la réparation de la peau humaine saine et diabétique (pathologique).

Figure 3: Optimisation de l’approche de coloration globale pour une utilisation avec d’autres anticorps. (A) Les vaisseaux sanguins ont été colorés avec de l’actine musculaire alpha lisse (α-SMA, vert) et de la kératine 14 (K14, rouge), tandis que les fibres matricielles ont été colorées avec du collagène I (COL 1, rouge) et de la fibronectine (Fn, vert). (B) La procédure de montage complet fournit jusqu’à la résolution de localisation au niveau de la cellule (K14, vert; K1, rouge). (C) CD1a+ve Cellules de Langerhans (vert) observées dans l’épiderme nouvellement formé. DAPI = noyaux bleus. Barre = 100 μm. Des lignes pointillées blanches montrent les bords intérieurs et extérieurs de la plaie et séparent la plaie de l’épiderme. Cette coloration a été réalisée sur une peau non diabétique. Veuillez cliquer ici pour voir une version agrandie de cette figure.
{kind=link}

Figure 4: Validité de la procédure de coloration à montage complet pour évaluer la cicatrisation des plaies. (A) Illustration illustrant comment la technique de coloration à montage entier peut évaluer les processus pertinents pour la plaie. Anticorps utilisés = texte rouge. K14 = kératine 14. COL 1 = collagène 1. Fn = fibronectine. (B) La procédure de coloration à montage entier (flèches bleues) introduit moins de variabilité dans les mesures de fermeture de la plaie que l’analyse histologique standard (flèches rouges). S1 = section 1. WE = bord de la plaie. Barre = 300 μm. Cette coloration a été réalisée sur une peau non diabétique. Veuillez cliquer ici pour voir une version agrandie de cette figure.
{kind=link}
Discussion
Dans ce protocole expérimental, nous décrivons une méthode optimisée pour évaluer la fermeture des plaies dans la peau ex vivo humaine en utilisant la coloration tissulaire à montage entier. Il s’agit d’une ressource importante pour permettre une évaluation critique des traitements potentiels des plaies et pour mieux comprendre la réponse de réparation des plaies humaines. Nous avons publié une évaluation de la cicatrisation dans les plaies cutanées ex vivo précédemment12,13, mais dans ces rapports, l’approche de coloration globale n’a pas été utilisée pour mesurer la fermeture de la plaie. La coloration à montage entier est beaucoup plus facile et nécessite moins d’expérience technique que l’histologie standard, qui implique l’incorporation de paraffine ou d’OCT et la section d’échantillons. La procédure de montage complet réduit également la variabilité expérimentale, permettant de quantifier l’ensemble de la plaie et pas seulement une seule section transversale à une position définie dans le tissu (voir la figure 4B pour une illustration comparative). Nous soutenons pleinement l’importance de quantifier la guérison de l’ensemble de la structure non symétrique de la plaie, comme l’ont clairement souligné Rhea et Dunnwald pour les plaies aiguës murines14. Ces auteurs ont montré l’importance de la coupe en série in vivo des plaies excisionnelles pour des mesures reproductibles et précises de la morphologie des plaies. La section en série pourrait également être appliquée aux plaies ex vivo humaines; cependant, pour une quantification précise de la fermeture de la plaie et de la réétitélialisation, la coloration à montage entier à haut débit devrait être la méthode préférée. Nous notons que ce protocole de coloration à montage complet devrait également être compatible avec le traitement ultérieur (cire ou OCT) pour l’analyse histologique traditionnelle.
La coloration à montage entier n’est pas sans inconvénients. Bien qu’il offre une reproductibilité plus élevée dans les expériences de cicatrisation des plaies, il nécessite l’utilisation de plus de tissus pour l’analyse que les techniques histologiques standard. Il peut s’agir d’un problème où l’accès aux tissus est limité, en particulier lorsque plusieurs anticorps doivent être évalués. Une autre approche consisterait à utiliser une méthode de plaie incisionnelle où la largeur de la plaie est relativement uniforme et la variabilité réduite (comme le montrent les plaies de souris et d’hommes15,16). Cependant, les plaies excisionnelles restent plus applicables à la plupart des types de plaies pathologiques17.
Dans cette étude, des plaies d’épaisseur partielle de 2 mm ont été créées au centre des explantes cutanées de 6 mm. Cette méthode peut être optimisée pour d’autres tailles de plaies excisionnelles et d’explants à différentes profondeurs de peau18. De plus, la force nécessaire pour générer des plaies variera d’un donneur à l’autre, où la peau âgée nécessitera moins de force pour la biopsie. Nous éviterions également d’utiliser une peau présentant des vergetures proéminentes ou d’autres altérations structurelles. Nous avons validé une gamme d’anticorps pour prendre en compte différents aspects de la réponse de guérison ex vivo. Ce protocole peut également être utilisé avec d’autres anticorps pertinents pour la peau, où les concentrations d’anticorps et les temps d’incubation devront être optimisés. Néanmoins, nous pensons que notre protocole est le plus adapté à la quantification absolue de la fermeture totale de la plaie, suivie d’une évaluation spatiale de protéines spécifiques d’intérêt. Alors que la monture entière offre une résolution réduite de l’immunolocalisation par rapport à l’analyse histologique standard des coupes de tissus, elle fournit des informations 3D supplémentaires qui manquent à l’histologie 2D standard.
Une mise en garde de l’évaluation de la guérison dans la peau ex vivo par rapport aux modèles in vivo est qu’elle manque de réponse systémique. Un aspect important de la réparation des plaies est l’inflammation et la granulation tissulaire ultérieure, qui est causée par un afflux de cellules inflammatoires et de cellules endothéliales du système vasculaire19. Malgré cette limitation, la peau ex vivo offre toujours une meilleure récapitulation de la cicatrisation clinique que les tests de plaies à base de cellules. Les expériences in vitro impliquent généralement des monocouches de type cellulaire unique ou des co-cultures cultivées sur du plastique de culture tissulaire, tandis que la peau ex vivo fournit un environnement natif pour explorer le comportement cellulaire. Plus récemment, un certain nombre de systèmes équivalents cutanés ont émergé, où la peau est cultivée en laboratoire à partir d’une matrice artificielle et de cellules cutanées isolées20,21. Bien que ces modèles imitent mieux la peau humaine que la plupart des approches in vitro, ils ne simulent toujours pas complètement l’environnement tissulaire natif et sont généralement trop fragiles pour se blesser de manière reproductible. De plus, nous (et d’autres) avons démontré que le tissu cutané humain ex vivo conserve les cellules immunitaires résidentes, ce qui contribuera sans aucun doute à la réparation22,23. Les travaux futurs devraient maintenant se concentrer sur l’extension de la viabilité et de l’immunocompétence du modèle ex vivo pour l’évaluation de la guérison à un stade avancé24. Une option consiste à faire progresser les technologies prometteuses d’organes sur puce capables de prolonger la viabilité des tissus et de maintenir l’architecture de la peau native jusqu’à deux semaines en culture25. Les modèles ex vivo ont également commencé à considérer l’importance de la réponse inflammatoire de la peau en incorporant avec succès des cellules immunitaires, telles que les neutrophiles, dans le tissu hôte26 ou en injectant des anticorps dans le tissu hôte pour provoquer une réaction immunitaire27. Nous nous attendons à ce que ces résultats ouvrent la voie au développement de méthodes plus raffinées et traduisibles à l’avenir.
L’un des principaux avantages de l’utilisation de la peau ex vivo pour mesurer la fermeture des plaies est la capacité de comparer les taux de guérison dans les tissus sains (p. ex., non diabétiques) et pathologiques (p. ex., diabétiques ou âgés). Ici, nous avons montré que la rééphatalisation et la formation de barrière sont en effet altérées dans les plaies diabétiques par rapport aux plaies ex vivo saines. En effet, cela fournit une voie pour l’évaluation préclinique de la réparation pathologique, où le vieillissement et le diabète sont des facteurs de risque majeurs pour développer des plaies chroniques1. Bien qu’il existe des modèles pathologiques in vitro, tels que des cellules isolées de tissus âgés et diabétiques, ou des cellules cultivées dans un taux élevé de glucose pour imiter l’hyperglycémie28,29,ces cellules peuvent rapidement perdre leur phénotype une fois retirées du microenvironnement in vivo. Une composante importante de l’environnement de guérison pathologique extrinsèque est la matrice dermique, qui est altérée à la fois dans le vieillissement et le diabète30. En effet, cette matrice perturbée affecte le comportement des fibroblastes résidents et naïfs31,32. Ainsi, l’importance d’étudier les cellules dans leur environnement tissulaire hôte ne peut être sous-estimée.
En résumé, notre protocole fournit une plate-forme importante pour quantifier la réétalisation des plaies humaines, explorer les facteurs régulateurs et tester la validité et l’efficacité des thérapies potentielles12,13. Bien que les essais précliniques nécessitent toujours des approches in vivo, une stratégie combinée utilisant des tissus humains ex vivo et des blessures murines in vivo devrait affiner la voie préclinique, réduisant l’utilisation animale tout en augmentant la translatabilité entre espèces.
Déclarations de divulgation
Les auteurs ne déclarent aucun conflit d’intérêts.
Remerciements
Nous tenons à remercier M. Paolo Matteuci et M. George Smith pour avoir fourni des tissus aux patients. Nous sommes également reconnaissants à Mlle Amber Rose Stafford pour son aide à la collecte de tissus et à l’Appel Daisy pour avoir fourni des installations de laboratoire.
matériels
| Name | Company | Catalog Number | Comments |
| 50 mL Falcon Tubes | Falcon | 352070 | For skin washing |
| 1.5 ml TubeOne Microcentrifuge Tubes, Natural (Sterile) | Starlab | S1615-5510 | For whole-mount staining |
| 48-Well CytoOne Plate, TC-Treated | Starlab | CC7682-7548 | For whole-mount staining |
| Acetic Acid Glacial | Fisher Chemical | A/0400/PB15 | Part of fixative |
| Alkyltrimethylammonium Bromide | Sigma-Aldrich | M7635 | Part of fixative |
| Anti-Alpha Smooth Muscle Actin Antibody [1A4] | Abcam | ab7817 | Stains blood vessels |
| Anti-Collagen I Antibody | Abcam | ab34710 | Stains collagen |
| Anti-Cytokeratin 14 Antibody [LL002] | Abcam | ab7800 | Stains epidermis |
| CD1A Antibody (CTB6) | Santa Cruz Biotechnology | sc-5265 | Stains Langerhans cells |
| DAPI (4',6-diamidino-2-phenylindole, dihydrochloride) | Thermo Fisher Scientific | 62247 | Counterstain for cell nuclei |
| Falcon 60mm Petri dishes | Falcon | 353004 | Human ex vivo culture |
| Fibronectin Antibody (EP5) | Santa Cruz Biotechnology | sc-8422 | Stains fibronectin |
| Formaldehyde, Extra Pure, Solution 37-41%, SLR | Fisher Chemical | F/1501/PB17 | Part of fixative |
| Gauze Swabs | Medisave | CS1650 | To clean skin |
| Gibco™ Antibiotic-Antimycotic Solution | Thermo Fisher Scientific | 15240062 | Human ex vivo culture |
| Gibco DMEM, high glucose, no glutamine | Thermo Fisher Scientific | 11960044 | Human ex vivo culture |
| Gibco Fetal Bovine Serum | Thermo Fisher Scientific | 10500064 | Human ex vivo culture |
| Gibco HBSS, no calcium, no magnesium | Thermo Fisher Scientific | 14170088 | Human ex vivo culture |
| Gibco L-Glutamine (200 mM) | Thermo Fisher Scientific | 25030081 | Human ex vivo culture |
| Hydrogen Peroxide | Sigma-Aldrich | H1009-100ML | For immunoperoxidase staining |
| ImageJ Software | National Institutes of Health | N/A | For image analysis |
| Invitrogen IgG (H+L) Cross-Adsorbed Goat anti-Mouse, Alexa Fluor 488 | Thermo Fisher Scientific | A11001 | Secondary antibody used depends on required fluorochromes and primary antibody |
| Invitrogen IgG (H+L) Cross-Adsorbed Goat anti-Rabbit, Alexa Fluor 594 | Thermo Fisher Scientific | A11012 | Secondary antibody used depends on required fluorochromes and primary antibody |
| Invitrogen LIVE/DEAD Viability/Cytotoxicity Kit, for mammalian cells | Thermo Fisher Scientific | L3224 | For viability assessment of tissue |
| Iris Forceps, 10 cm, Curved, 1x2 teeth | World Precision Instruments | 15917 | To create wounds |
| Iris Scissors, 11 cm, Curved, SuperCut, Tungsten Carbide | World Precision Instruments | 501264 | To create wounds |
| Iris Scissors, 11 cm, Straight, SuperCut, Tungsten Carbide | World Precision Instruments | 501263 | To remove adipose tissue |
| Keratin 1 Polyclonal Antibody, Purified | Biolegend | 905201 | Stains epidermis |
| Keratin 14 Polyclonal Antibody, Purified | Biolegend | 905301 | Stains epidermis |
| LSM 710 Confocal Laser Scanning Microscope | Carl Zeiss | Discontinued | For fluorescent imaging |
| Merck Millipore Absorbent pads | Merck Millipore | AP10045S0 | Human ex vivo culture |
| Merck Millipore Nylon Hydrophilic Membrane Filters | Merck Millipore | HNWP04700 | Human ex vivo culture |
| Normal Goat Serum Solution | Vector Laboratories | S-1000-20 | Animal serum used depends on secondary antibody |
| Phosphate Buffer Solution | Sigma-Aldrich | P3619 | For wash buffer |
| Sodium Azide | Sigma-Aldrich | S2002 | For blocking buffer |
| Sodium Chloride | Fisher Bioreagents | BP358-212 | Part of fixative |
| Sterilisation Pouches | Medisave | SH3710 | To sterilise instruments |
| Stiefel 2mm biopsy punches | Medisave | BI0500 | For partial thickness wound |
| Stiefel 6mm biopsy punches | Medisave | BI2000 | For outer explant |
| Thermo Scientific Sterilin Standard 90mm Petri Dishes | Thermo Fisher Scientific | 101VR20 | To prepare skin |
| Triton X-100 | Fisher Chemical | T/3751/08 | For wash buffer |
| VECTASTAIN Elite ABC-HRP Kit, Peroxidase (Rabbit IgG) | Vector Laboratories | PK-6101 | For immunoperoxidase staining; HRP kit used depends on primary antibody |
| Vector NovaRED Substrate Kit, Peroxidase (HRP) | Vector Laboratories | SK-4800 | For immunoperoxidase staining |
| Wireless Digital Microscope | Jiusion | N/A | For brightfield imaging |
Références
- Lindholm, C., Searle, R. Wound management for the 21st century: combining effectiveness and efficiency. International Wound Journal. 13, 5-15 (2016).
- Guest, J. F., et al. Health economic burden that different wound types impose on the UK's National Health Service. International Wound Journal. 14 (2), 322-330 (2017).
- Guest, J. F., Fuller, G. W., Vowden, P. Diabetic foot ulcer management in clinical practice in the UK: costs and outcomes. International Wound Journal. 15 (1), 43-52 (2018).
- López-Valverde, M. E., et al. Perioperative and long-term all-cause mortality in patients with diabetes who underwent a lower extremity amputation. Diabetes Research and Clinical Practice. 141, 175-180 (2018).
- Wilkinson, H. N., Hardman, M. J. The role of estrogen in cutaneous ageing and repair. Maturitas. 103, 60-64 (2017).
- Frykberg, R. G., Banks, J. Challenges in the treatment of chronic wounds. Advances in Wound Care. 4 (9), 560-582 (2015).
- Wilkinson, H. N., Hardman, M. J. Wound healing: cellular mechanisms and pathological outcomes. Open Biology. 10 (9), 200223 (2020).
- Ansell, D. M., Holden, K. A., Hardman, M. J. Animal models of wound repair: Are they cutting it. Experimental Dermatology. 21 (8), 581-585 (2012).
- Elliot, S., Wikramanayake, T. C., Jozic, I., Tomic-Canic, M. A modeling conundrum: murine models for cutaneous wound healing. Journal of Investigative Dermatology. 138 (4), 736-740 (2018).
- Mazio, C., et al. Pre-vascularized dermis model for fast and functional anastomosis with host vasculature. Biomaterials. 192, 159-170 (2019).
- Wilkinson, H. N., Iveson, S., Catherall, P., Hardman, M. J. A novel silver bioactive glass elicits antimicrobial efficacy against Pseudomonas aeruginosa and Staphylococcus aureus in an ex vivo skin wound biofilm model. Frontiers in Microbiology. 9, 1450 (2018).
- Wilkinson, H. N., et al. Elevated local senescence in diabetic wound healing is linked to pathological repair via CXCR2. Journal of Investigative Dermatology. 139 (5), 1171-1181 (2019).
- Wilkinson, H. N., et al. Tissue iron promotes wound repair via M2 macrophage polarization and the chemokine (CC motif) ligands 17 and 22. The American Journal of Pathology. 189 (11), 2196-2208 (2019).
- Rhea, L., Dunnwald, M. Murine excisional wound healing model and histological morphometric wound analysis. Journal of Visualized Experiments. 162, e61616 (2020).
- Ansell, D. M., Campbell, L., Thomason, H. A., Brass, A., Hardman, M. J. A statistical analysis of murine incisional and excisional acute wound models. Wound Repair and Regeneration. 22 (2), 281-287 (2014).
- Rizzo, A. E., Beckett, L. A., Baier, B. S., Isseroff, R. R. The linear excisional wound: an improved model for human ex vivo wound epithelialization studies. Skin Research and Technology. 18 (1), 125-132 (2012).
- Olsson, M., et al. The humanistic and economic burden of chronic wounds: a systematic review. Wound Repair and Regeneration. 27 (1), 114-125 (2019).
- Mendoza-Garcia, J., Sebastian, A., Alonso-Rasgado, T., Bayat, A. Optimization of an ex vivo wound healing model in the adult human skin: Functional evaluation using photodynamic therapy. Wound Repair and Regeneration. 23 (5), 685-702 (2015).
- Brownhill, V. R., et al. Pre-clinical assessment of single-use negative pressure wound therapy during in vivo porcine wound healing. Advances in Wound Care. , (2020).
- Diekmann, J., et al. A three-dimensional skin equivalent reflecting some aspects of in vivo aged skin. Experimental Dermatology. 25 (1), 56-61 (2016).
- Vidal Yucha, S. E., Tamamoto, K. A., Nguyen, H., Cairns, D. M., Kaplan, D. L. Human skin equivalents demonstrate need for neuro-immuno-cutaneous system. Advanced Biosystems. 3 (1), 1800283 (2019).
- Dijkgraaf, F. E., et al. Tissue patrol by resident memory CD8+ T cells in human skin. Nature Immunology. 20 (6), 756-764 (2019).
- He, X., de Oliveira, V. L., Keijsers, R., Joosten, I., Koenen, H. J. Lymphocyte isolation from human skin for phenotypic analysis and ex vivo cell culture. Journal of Visualized Experiments. (110), e52564 (2016).
- Pupovac, A., et al. Toward immunocompetent 3D skin models. Advanced Healthcare Materials. 7 (12), 1701405 (2018).
- Ataç, B., et al. Skin and hair on-a-chip: in vitro skin models versus ex vivo tissue maintenance with dynamic perfusion. Lab on a Chip. 13 (18), 3555-3561 (2013).
- Kim, J. J., et al. A microscale, full-thickness, human skin on a chip assay simulating neutrophil responses to skin infection and antibiotic treatments. Lab on a Chip. 19 (18), 3094-3103 (2019).
- Jardet, C., et al. Development and characterization of a human Th17-driven ex vivo skin inflammation model. Experimental Dermatology. 29 (10), 993-1003 (2020).
- Chen, J. L., et al. Metformin attenuates diabetes-induced tau hyperphosphorylation in vitro and in vivo by enhancing autophagic clearance. Experimental Neurology. 311, 44-56 (2019).
- Demirovic, D., Rattan, S. I. Curcumin induces stress response and hormetically modulates wound healing ability of human skin fibroblasts undergoing ageing in vitro. Biogerontology. 12 (5), 437-444 (2011).
- Wilkinson, H. N., Hardman, M. J. Wound senescence: A functional link between diabetes and ageing. Experimental Dermatology. 30 (1), 68-73 (2020).
- Fisher, G. J., et al. Collagen fragmentation promotes oxidative stress and elevates matrix metalloproteinase-1 in fibroblasts in aged human skin. The American Journal of Pathology. 174 (1), 101-114 (2009).
- Quan, T., Little, E., Quan, H., Voorhees, J. J., Fisher, G. J. Elevated matrix metalloproteinases and collagen fragmentation in photodamaged human skin: impact of altered extracellular matrix microenvironment on dermal fibroblast function. Journal of Investigative Dermatology. 133 (5), 1362 (2013).
Réimpressions et Autorisations
Demande d’autorisation pour utiliser le texte ou les figures de cet article JoVE
Demande d’autorisationThis article has been published
Video Coming Soon
À PROPOS DE JoVE
Copyright © 2025 MyJoVE Corporation. Tous droits réservés.