
Method Article
Détection simple et rapide de l’activité de la topoisomérase 1 basée sur l’amplification du cercle roulant dans des échantillons biologiques bruts
Dans cet article
Résumé
Un protocole pour la détection sensible et quantitative de l’activité de la topoisomérase 1 à l’aide du test de détection de l’activité enzymatique améliorée du cercle roulant est décrit. La méthode permet de détecter l’activité de la topoisomérase 1 à partir de composants purifiés ou d’extraits cellulaires/tissulaires. Ce protocole a de nombreuses applications dans tous les domaines impliquant la détection de l’activité enzymatique.
Résumé
Des techniques basées sur l’amplification isotherme telles que l’amplification du cercle roulant ont été utilisées avec succès pour la détection d’acides nucléiques, de quantités de protéines ou d’autres molécules pertinentes. Ces méthodes se sont révélées être des alternatives substantielles à la PCR ou à l’ELISA pour les applications cliniques et de recherche. De plus, la détection de la quantité de protéines (par transfert Western ou immunohistochimie) est souvent insuffisante pour fournir des informations pour le diagnostic du cancer, alors que la mesure de l’activité enzymatique représente un biomarqueur précieux. La mesure de l’activité enzymatique permet également le diagnostic et le traitement potentiel des maladies transmises par des agents pathogènes. Chez tous les eucaryotes, les topoisomérases sont les enzymes clés de liaison à l’ADN impliquées dans le contrôle de l’état topologique de l’ADN au cours de processus cellulaires importants et font partie des biomarqueurs importants pour le pronostic et le traitement du cancer.
Au fil des ans, les topoisomérases ont été considérablement étudiées en tant que cible potentielle des médicaments antiparasitaires et anticancéreux avec des bibliothèques de composés naturels et synthétiques à petites molécules qui sont étudiées chaque année. Ici, la méthode d’amplification du cercle roulant, appelée test de détection de l’activité enzymatique améliorée du cercle roulant (REEAD) qui permet la mesure quantitative de l’activité de la topoisomérase 1 (TOP1) d’une manière simple, rapide et sans gel, est présentée. En clivant et en ligaturant un substrat d’ADN spécialement conçu, TOP1 convertit un oligonucléotide d’ADN en un cercle fermé, qui devient le modèle pour l’amplification du cercle roulant, donnant ~103 produits de cercle roulant répétés en tandem. Selon l’incorporation de nucléotides pendant l’amplification, il existe la possibilité de différentes méthodes de lecture, de la fluorescence à la chimiluminescence en passant par la colorimétrique. Comme chaque ligature de clivage médiée par TOP1 génère un cercle d’ADN fermé, le test est très sensible et directement quantitatif.
Introduction
Les topoisomérases appartiennent à la classe des enzymes modifiant l’ADN et beaucoup d’entre elles se sont révélées utiles comme biomarqueurs des maladies humaines 1,2,3,4,5,6. TOP1 est impliqué dans la résolution du stress topologique associé aux processus cellulaires tels que la réplication de l’ADN, la transcription des gènes, la recombinaison et la ségrégation chromosomique7. TOP1 peut résoudre à la fois les superbobines négatives et positives par un mécanisme qui implique la formation d’une cassure monocaténaire transitoire dans l’ADN 8,9. Après s’être lié à l’ADN, TOP1 positionne le site actif tyrosine (Tyr723) pour effectuer une attaque nucléophile sur le squelette du phosphodiester. Un complexe de clivage TOP1-ADN est ensuite généré avec l’enzyme attachée de manière covalente à l’extrémité 3' du brin d’ADN brisé. Cela libère la contrainte de torsion en permettant à l’extrémité 5' du brin clivé de tourner autour du brin intact. Enfin, le groupe hydroxyle de l’extrémité 5' effectue une attaque nucléophile sur la liaison 3'-phosphotyrosyle. En conséquence, TOP1 est libéré et l’épine dorsale de l’ADN est restaurée8.
Plusieurs tests ont été mis au point pour étudier les étapes du cycle catalytique TOP1, notamment le test de relaxation10, le test de déplacement de mobilité électrophorétique (EMSA)11,12, les tests de clivage-ligature du suicide de l’ADN 13,14 et le test in vivo du complexe d’enzymes (ICE) 15 . Cependant, ces tests ont plusieurs limites car ils dépendent de l’électrophorèse sur gel, qui nécessite des agents intercalants de l’ADN, ou ils nécessitent une formation et un équipement hautement spécialisés. De plus, les tests nécessitent de grandes quantités d’enzyme TOP1 purifiée (allant de 1 à 5 ng pour le test de relaxation et de 50 à 200 ng pour l’EMSA et le test de clivage-ligature) ou des extraits d’au moins 106 cellules pour fonctionner de manière optimale. Par conséquent, une méthode très sensible qui permet la détection spécifique de l’activité TOP1 dans des échantillons biologiques bruts et au niveau de l’événement catalytique unique, appelée test REEAD, a été développée16.
Ici, un protocole de détection de l’activité TOP1 à l’aide du test REEAD16 est présenté. Une représentation schématique de l’essai est illustrée à la figure 1. Une grille en silicone conçue sur mesure est fixée à une lame fonctionnalisée pour créer une configuration multipuits à lames de verre, appelée wellmaker dans ce qui suit (Figure 1A). Ceci est suivi par le couplage d’une amorce modifiée 5'-amino aux groupes fonctionnels du NHS dans les puits de la lame (figure 1B). La réaction de clivage et de ligature médiée par TOP1 convertit un substrat d’ADN spécifique en un cercle fermé. Le substrat spécifique au TOP1 (Figure 1C) se replie spontanément en une forme d’haltère contenant une tige double brin et deux boucles simple brin. L’une des boucles est complémentaire à l’amorce ancrée en surface. La région de la tige contient un site de clivage TOP1 privilégié à trois bases en amont de l’extrémité 3' et un surplomb 5'-hydroxyle. La circularisation du substrat peut être réalisée à l’aide d’un extrait cellulaire/tissulaire (Figure 1C) ou d’un TOP1 purifié recombinant (Figure 1D).
Lorsque le substrat est clivé par TOP1, l’enzyme devient transitoirement liée à l’extrémité 3' et le fragment à trois bases diffuse, permettant au surplomb 5' de recuit au substrat et facilitant la ligature médiée par TOP1. La ligature entraîne une circularisation du substrat et par la suite la dissociation de TOP1. Le cercle fermé est hybridé à l’amorce ancrée en surface (Figure 1E) et est utilisé comme matrice pour l’amplification isotherme du cercle roulant (RCA) médiée par la phi29 polymérase, qui peut effectuer une RCA avec déplacement de brins, produisant 103 produits de répétition en tandem. Au cours de l’étape RCA, des nucléotides marqués par fluorescence (Figure 1F) ou couplés à la biotine (Figure 1G) peuvent être incorporés17. L’incorporation de nucléotides marqués par fluorescence permet de détecter les RCP à l’aide d’un microscope à fluorescence ou d’un scanner à fluorescence. Alternativement, un anticorps anti-biotine couplé à la peroxydase de raifort (HRP) (Figure 1H) peut lier les nucléotides biotinylés, permettant ainsi la détection des produits en cercle laminé (RCP) en utilisant soit une chimiluminescence améliorée (ECL), soit par la conversion de la 3,3',5,5'-tétraméthylbenzidine (TMB) en une couleur détectable. L’ACR suit une cinétique de réaction linéaire, ce qui rend le test REEAD directement quantitatif, car un RCP représente une seule réaction de clivage-ligature médiée par TOP1. Ici, il est montré que ce test peut être utilisé pour détecter l’activité de TOP1 en tant qu’enzyme recombinante purifiée ou extraite d’échantillons bruts et comme outil de dépistage de médicaments anti-TOP1.
Protocole
REMARQUE : Trouvez une liste des compositions tampons, de l’équipement et des autres matériaux requis dans le Tableau des matériaux.
1. Culture cellulaire
- Cultivez les cellules préférées dans un milieu approprié et cultivez-les selon les instructions. À titre d’exemple, cultivez du Caco2, des cellules dérivées de l’adénocarcinome colorectal, dans un milieu essentiel minimal (MEM) complété par 20 % de sérum fœtal bovin (FBS), 1 % d’acides aminés non essentiels (NEAA), 100 unités/ml de pénicilline et 100 mg/mL de streptomycine. Maintenir les cultures cellulaires dans un incubateur humidifié (5% CO2/95% atmosphère d’air à 37 °C). Plaquez les cellules dans des flacons de culture tissulaire et divisez-les tous les 3 jours pour maintenir les cellules à 70% de confluence.
- Récolter les cellules d’une fiole confluente à 70% par traitement à la trypsine (solution de trypsine à 0,25%, solution d’EDTA à 0,02%) et laver avec une solution saline tamponnée au phosphate (PBS).
- Remettez en suspension la pastille de cellule dans le PBS pour ajuster la concentration de la cellule à 0,5 × 106 cellules/tube. Faire tourner à 200 × g pendant 5 min et aspirer soigneusement le surnageant.
REMARQUE : Les cellules utilisées pour le REEAD doivent être utilisées fraîches et conservées sur de la glace. Les résultats obtenus avec les cellules Caco2 ont été récemment publiés17. Le test a été testé avec une variété de lignées cellulaires, y compris les cellules dérivées du cancer du côlon, du sein, du poumon et du col de l’utérus 18,19,20,21,22,23, mais il peut être utilisé avec tous les échantillons biologiques bruts contenant TOP1, tels que les tissus, le sang et la salive.
2. Préparation des diapositives fonctionnalisées
- Fixez la grille d’isolateur en silicone conçue sur mesure à la lame fonctionnalisée, rendant ainsi le faiseur de puits. Appuyez sur le silicone pour éviter la formation de bulles d’air (voir Figure 1A).
- Préparer un mélange d’apprêt 5'-amino 5'-amino 5 μM dans un tampon d’impression 1x. Ajouter 4 μL du mélange à chaque puits et placer le puits dans une chambre d’hybridation avec NaCl saturé à des températures comprises entre 15 °C et 25 °C, à l’abri de la lumière.
REMARQUE: Une chambre d’hybridation peut facilement être fabriquée en utilisant une boîte d’embout de pipette remplie de NaCl saturé. L’hybridation dure un minimum de 16 h et un maximum de 72 h. Voir la séquence de l’amorce 5'-amino à la figure 1B. Les lames sont fonctionnalisées avec des groupes NHS pour permettre la liaison d’oligonucléotides amino-modifiés.
3. Génération de substrats circulaires fermés
- Préparation de substrats circulaires fermés avec TOP1 recombinant ou extrait cellulaire.
- Lyser une pastille cellulaire de 0,5 × 106 cellules dans 500 μL de tampon de lyse pour atteindre une densité cellulaire de 1 000 cellules/μL. Incuber pendant 10 min sur de la glace.
- Préparer un mélange circulaire de 2 μL de substrat spécifique TOP1 de 5 μM (la séquence du substrat est la suivante : 5'-AGA AAA ATT TTT AAA AAA ACT GTG AAG ATC GCT TAT TTT AAA AAT AAA TCT AAG TCT TTT AGA TCC CTC AAT GCA CAT GTT TGG CTC CGA TCT AAA AGA CTT AGA-3') et 2 μL d’enzyme TOP1 recombinante ou 2 μL d’extrait cellulaire (voir étape 3.1.1) dans 16 μL de 1x tampon de réaction TOP1.
REMARQUE : La concentration de TOP1 recombinant utilisée dépend de la méthode de lecture choisie (voir Figure 2, Figure 3, Figure 4 et Figure 5). Voir la séquence du substrat spécifique au TOP1 à la figure 1C. - Incuber pendant 30 min à 37 °C (voir figure 1C,D).
- Arrêtez la réaction en ajoutant 2 μL de SDS à 1%.
- Préparation de substrats circulaires fermés en présence de médicaments
- Préparer un mélange circulaire de 2 μL de substrat spécifique TOP1 de 5 μM et de 1 μL de DMSO à 100 % ou 1 μL de camptothécine (CPT) de 1,6 mM dans 14 μL de 1x tampon de réaction TOP1. Ajouter 2 μL d’enzyme TOP1 recombinante à 5 ng/μL.
REMARQUE: D’autres composés à petites molécules ou médicaments peuvent être utilisés. Dans ce cas, un titrage du composé doit être effectué. Si le composé est dissous dans un solvant autre que le DMSO, celui-ci doit être utilisé comme témoin au lieu du DMSO. - Incuber pendant 1 min à 37 °C.
- Arrêtez la réaction en ajoutant 2 μL de SDS à 1%.
- Préparer un mélange circulaire de 2 μL de substrat spécifique TOP1 de 5 μM et de 1 μL de DMSO à 100 % ou 1 μL de camptothécine (CPT) de 1,6 mM dans 14 μL de 1x tampon de réaction TOP1. Ajouter 2 μL d’enzyme TOP1 recombinante à 5 ng/μL.
REMARQUE: L’étape 3 peut être démarrée en parallèle avec l’étape 2 et les cercles peuvent être stockés à 4 ° C pendant la nuit. Alternativement, les cercles d’ADN peuvent être préparés en même temps que l’étape 4 et utilisés immédiatement. Voir la séquence du substrat spécifique au TOP1 à la figure 1C. Le substrat se plie en forme d’haltère avec une région de tige double contenant un site de clivage TOP1 préféré et deux boucles simple brin. Le substrat d’haltère ouvert est converti en un substrat circulaire fermé par clivage et ligature médiés par TOP1 et est ci-après appelé cercle. Comme il y a un excès de l’amorce 5'-amino, il n’y aura pas de concurrence entre les substrats ouverts et fermés spécifiques au TOP1.
4. Blocage du faiseur de puits
- Plongez le faiseur de puits dans un plateau de 5 cm x 5 cm rempli de tampon 1 qui a été préchauffé à 50 °C. À l’aide d’une pipette, poussez le liquide à l’intérieur des puits pour vous assurer qu’il n’y a pas de bulles d’air. Incuber pendant 30 min à 50 °C.
- Retirer le tampon 1 et laver 2 x 1 min avec dH2O. Agiter vigoureusement à la main.
- Ajouter le tampon 2 qui a été préchauffé à 50 °C. Assurez-vous qu’il n’y a pas de bulles d’air dans les puits. Incuber pendant 30 min à 50 °C.
- Laver 2 x 1 min avec dH2O. Agiter vigoureusement à la main.
- Laver avec 70% d’EtOH pendant 1 min. Secouer vigoureusement à la main.
- Laissez l’installation du puits sécher à l’air.
REMARQUE: Utilisez de l’air comprimé pour sécher les puits. Alternativement, il est possible de souffler de l’air à l’aide d’une pipette Pasteur. Assurez-vous que les puits sont secs avant de continuer.
5. Hybridation des cercles au faiseur de puits
- Ajouter 4 μL des cercles (réalisés à l’étape 3) à chaque puits correspondant.
- Placer le fabricant de puits dans une chambre d’humidité pendant 1 h avecdH2Oà 37 °C.
NOTE: Une chambre d’humidité peut être fabriquée en utilisant une boîte pour les pointes de pipettes remplies de dH2O. Alternativement, l’hybridation peut être effectuée pendant la nuit à 25 ° C dans la chambre d’humidité.
6. Lavage
- Lavez le puits avec Buffer 3.
- Retirez la mémoire tampon 3 et remplacez-la par la mémoire tampon 4.
- Retirer le tampon 4 et laver avec 70% d’EtOH.
- Retirez l’EtOH et laissez sécher le faiseur de puits.
REMARQUE: Toutes les étapes de lavage sont effectuées pendant 1 min dans un plateau de 5 cm x 5 cm en immergeant complètement la glissière. Assurez-vous toujours qu’il n’y a pas de bulles d’air à l’intérieur des puits.
7. Amplification du cercle roulant
- Préparer un mélange RCA de 1x tampon réactionnel Phi29 complété par 0,2 μg/μL de BSA, 1 unité de polymérase Phi29 et 0,25 mM de dNTP et 0,0125 mM d’ATTO-488-dUTP pour la lecture de fluorescence, ou 0,1 mM dATP, 0,1 mM de DTTP, 0,1 mM dGTP, 0,09 mM dCTP et 0,01 mM de biotine-dCTP pour la lecture colorimétrique/chimiluminescente. Ajouter 4 μL à chaque puits. Voir la figure 1E.
NOTE: La préparation du mélange RCA doit être faite sur glace. Lors de l’incorporation de nucléotides fluorescents dans le RCA, évitez la lumière directe de cette étape pour protéger les fluorophores. - Incuber pendant 2 h à 37 °C dans la chambre d’humidité. En cas d’utilisation de nucléotides fluorescents, placez la chambre d’humidité dans l’obscurité. Voir la figure 1F,G.
8. Lavage
- Pour les protocoles de lecture chimiluminescence ou colorimétrique, lavez le fabricant de puits comme à l’étape 6, puis passez à l’étape 11.
- Pour le protocole de lecture de fluorescence, retirez la grille en silicone à l’aide d’une pince à épiler avant de la laver comme dans les étapes suivantes.
- Laver la lame pendant 10 min dans la mémoire tampon 3.
- Retirez la mémoire tampon 3 et remplacez-la par la mémoire tampon 4. Laver pendant 5 min.
- Retirer le tampon 4 et laver pendant 1 min dans de l’EtOH à 70 %.
- Retirez l’EtOH et laissez sécher le faiseur de puits.
9. Visualisation de produits de cercles de laminage fluorescents à l’aide d’un scanner à fluorescence
- Scannez la lame dans un scanner à fluorescence à l’aide des filtres correspondant au fluorophore utilisé. Utilisez le photomultiplicateur maximum possible qui ne donne pas de saturation.
REMARQUE: Le scanner fluorescent utilisé pour l’acquisition d’images dans ce protocole est équipé d’un laser 473 nm et d’un filtre FAM à excitation 490 nm, émission 520 nm. - Importez l’image dans ImageJ et changez le type d’image en 8 bits (Image | Type | 8 bits). Pour mesurer les bandes séparément, limitez la surface mesurée à l’aide de l’outil de dessin rectangulaire de la barre d’outils. Dessinez la zone souhaitée et mesurez l’intensité (Analyser | mesure). Ensuite, déplacez la zone dessinée à l’origine vers la bande suivante et mesurez.
- Tracez les données dans le logiciel souhaité. Des images représentatives, y compris la quantification extraite d’ImageJ, sont illustrées à la figure 2.
10. Visualisation de produits de cercles roulants fluorescents à l’aide d’un microscope à fluorescence
- Avec un stylo résistant à l’EtOH, dessinez la position des puits avant de retirer la grille en silicone et lavez la lame comme à l’étape 8.2. Après les étapes de lavage, montez la lame avec 1 μL de support de montage sans 4',6-diamidino-2-phénylindole (DAPI) et ajoutez un verre de couverture. Pour permettre la visualisation dans la caméra, collez la lame sur une lame de microscope de 76 mm x 26 mm.
- Analysez à l’aide d’un microscope à fluorescence équipé d’un objectif d’immersion dans l’huile 60x et d’une caméra. Prenez 12 à 15 images de chaque échantillon/puits.
- Importez les images dans ImageJ et empilez-les (Image | Piles | Image à empiler). Remplacez le type d’image par 8 bits (Image | Type | 8 bits).
- Définir le seuil (Image | Ajuster | seuil).
- Définissez le seuil comme suit : réglez la barre inférieure et ajustez la barre supérieure de sorte que seuls les bons signaux soient rouges et que l’arrière-plan soit noir. Assurez-vous que le seuil est aussi bas que possible avant que les signaux réels ne commencent à disparaître.
- Parcourez les images individuelles et assurez-vous qu’elles correspondent aux images avant de définir le seuil. Notez que le seuil peut changer d’une expérience à l’autre.
- Compter les signaux (Analyser | Analyser les particules). Assurez-vous que le champ résumé dans les paramètres ImageJ est coché.
- Exportez les résultats vers une feuille de calcul ou un autre logiciel pour un traitement ultérieur des données. Des images représentatives, y compris la quantification extraite d’ImageJ, sont illustrées à la figure 3.
11. Couplage d’anticorps anti-biotine conjugués HRP aux produits de cercle roulant marqués à la biotine
- Ajouter 4 μL d’anticorps anti-biotine conjugués HRP dilués 1:300 dans le tampon 5 complétés par 5 % de lait écrémé écrémé et 5 % de BSA à chaque puits.
- Incuber pendant 50 min à 15-25 °C dans la chambre d’humidité (voir Figure 1H).
- Lavez 3 x 3 min avec Buffer 5.
- Sécher le faiseur de puits.
12. Visualisation des produits de cercle roulant marqués à la biotine
- Visualisation avec ECL
- Mélanger 40 μL de Luminol ECL et 40 μL deH2O2immédiatement avant utilisation. Ajouter 2 μL à chaque puits.
- Visualisez la diapositive à l’aide d’une caméra CCD ou sur des films radiographiques.
- Visualisation avec TMB
- Après l’étape 11.4, retirez la grille en silicone. Ensuite, ajoutez 400 μL de TMB sur toute la lame et placez la lame dans une chambre d’humidité. Attendez ~5-10 min pour le développement des couleurs. Après le développement de la couleur, lavez la lame avec 70% d’EtOH.
- Visualisez le développement des couleurs à l’œil nu. Prenez une photo de la diapositive avec un appareil photo ou un téléphone portable.
- Importez l’image dans ImageJ et changez le type d’image en 8 bits (Image | Type | 8 bits). Pour mesurer les bandes séparément, limitez la surface mesurée à l’aide de l’outil de dessin rectangulaire de la barre d’outils. Dessinez la zone souhaitée et mesurez l’intensité (Analyser | mesure). Ensuite, déplacez la zone dessinée à l’origine vers la bande suivante et mesurez.
- Tracez les données dans le logiciel souhaité. Des images représentatives, y compris la quantification extraite d’ImageJ, sont illustrées à la figure 4, à la figure 5 et à la figure 6.
Résultats
Ici, un protocole de détection de l’activité TOP1 à l’aide du test REEAD est présenté16. Le protocole a été utilisé pour détecter l’activité recombinante de TOP1 à l’aide de quatre méthodes de lecture différentes: scanner à fluorescence, microscope à fluorescence, chimiluminescence et colorimétrique. Le protocole a également été utilisé pour détecter l’activité de TOP1 extrait des cellules Caco2, comme exemple d’extrait brut, avec lecture par chimiluminescence ou TMB. De plus, le protocole a été utilisé comme outil de dépistage de médicaments, pour détecter l’inhibition de l’activité recombinante de TOP1 par CPT, comme exemple d’inhibiteur spécifique de TOP1, en utilisant la méthode de lecture par chimiluminescence.
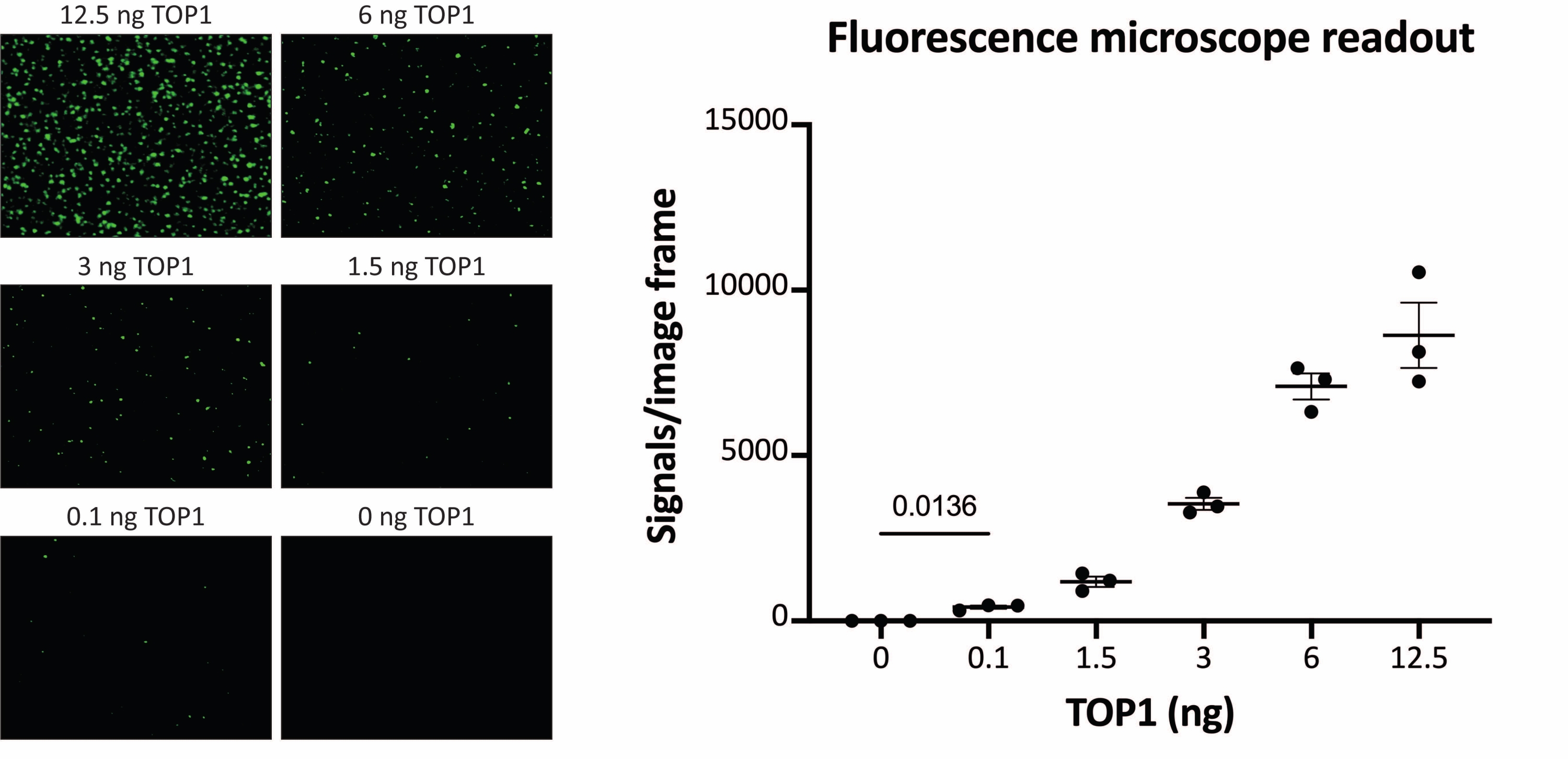
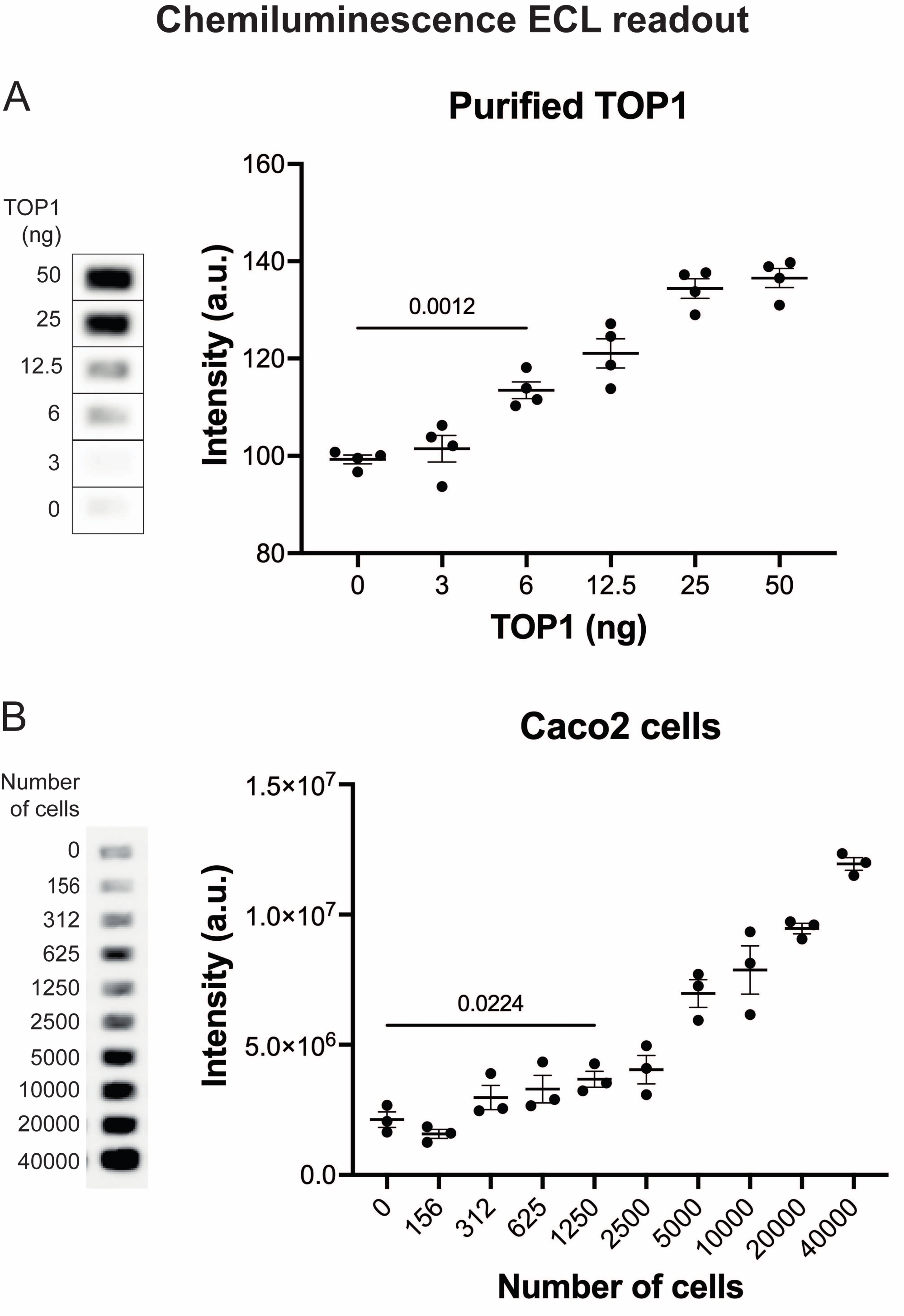
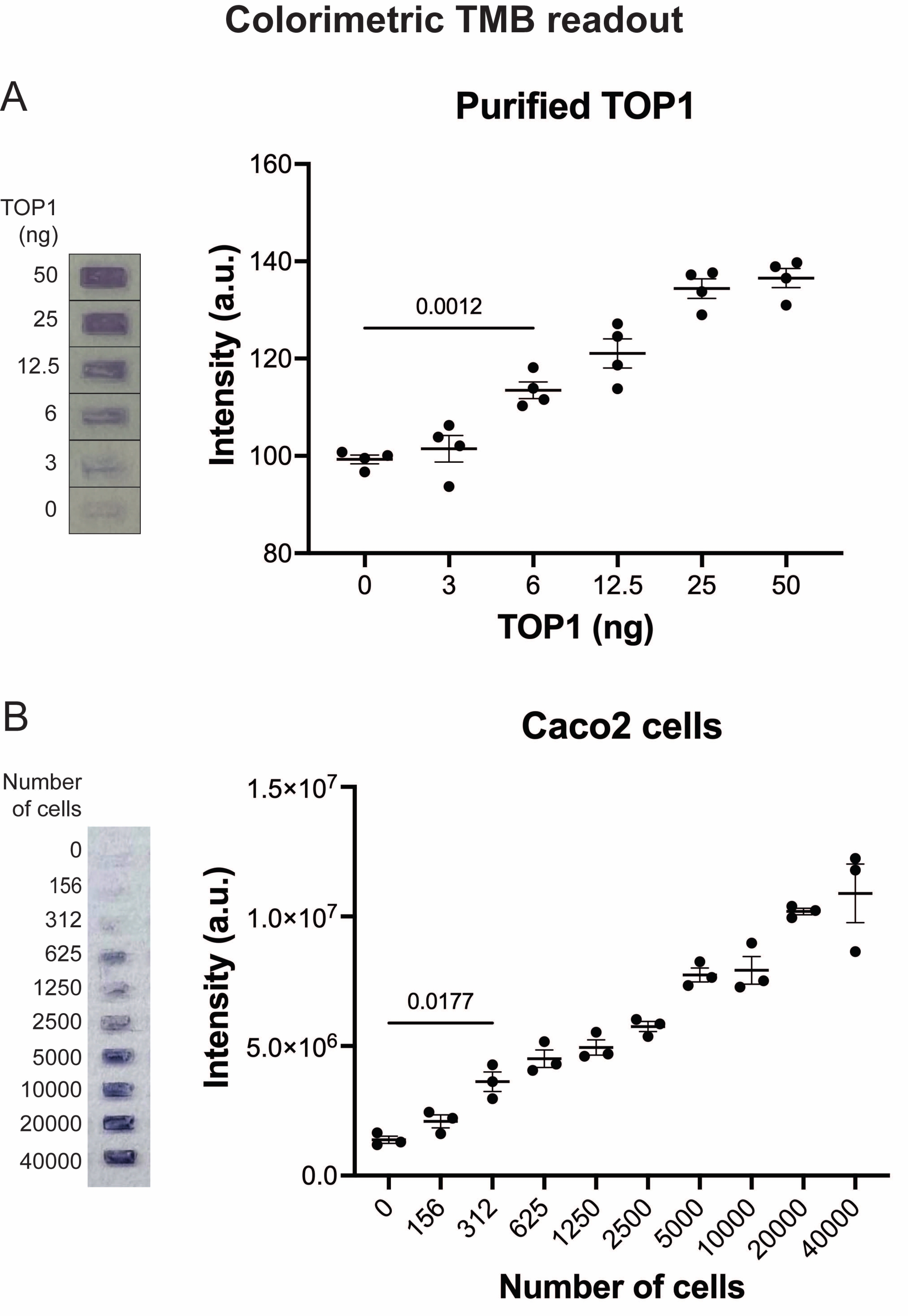
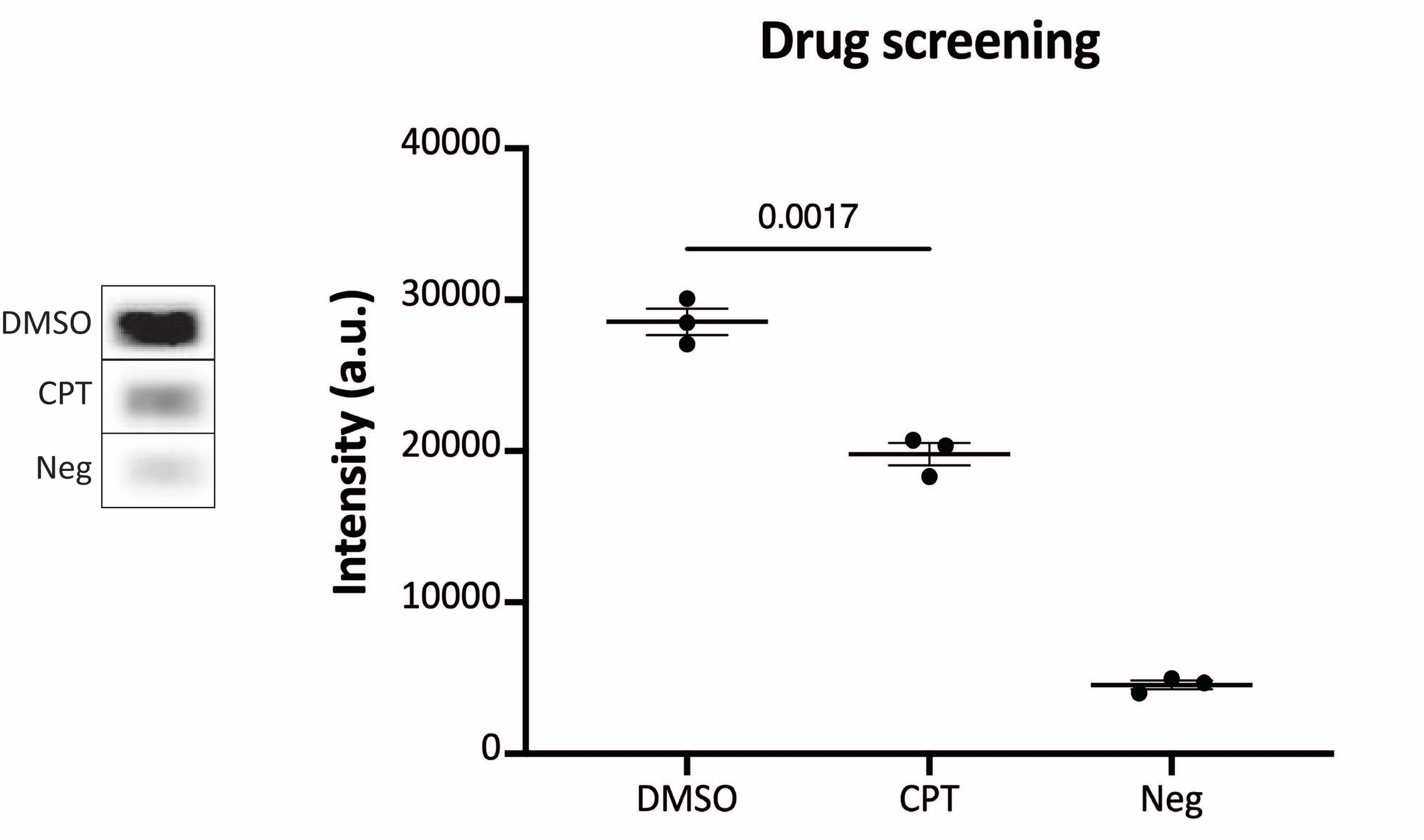
Les résultats obtenus lors de l’analyse de l’activité TOP1 à l’aide de la lecture de fluorescence sont illustrés à la figure 2 et à la figure 3. Un balayage représentatif de la lame fluorescente et la quantification résultante sont présentés à la figure 2. Comme le montre la quantification, cette lecture a une limite de détection de 12,5 ng de TOP1. Les images représentatives et la quantification obtenue lors de l’utilisation de la lecture du microscope fluorescent sont présentées à la figure 3. En utilisant cette méthode de lecture, la limite de détection est aussi faible que 0,1 ng de TOP1. Les résultats obtenus lors de l’analyse de l’activité TOP1 à l’aide de la lecture de la chimiluminescence sont représentés à la figure 4, et les résultats de la lecture colorimétrique sont représentés à la figure 5. La limite de détection de ces méthodes de lecture est de 6 ng de TOP1 ou sous forme de TOP1 extrait de 312 cellules Caco2. La figure 6 montre les mesures de l’activité TOP1 en présence de 80 μM CPT ou DMSO à titre d’exemple d’application de dépistage de médicaments. L’image représentative et la quantification résultante de trois expériences indépendantes montrent que CPT inhibe la circularisation médiée par TOP1 du substrat comme prévu24,25.

Figure 1 : Représentation schématique du protocole REEAD. (A,B) Préparation des diapositives fonctionnalisées. La grille en silicone est fixée à la lame fonctionnalisée. Ensuite, l’amorce 5'-amino est couplée à la lame, permettant ainsi l’amplification du substrat spécifique au TOP1. (C, D) Génération de substrats circulaires fermés. Le substrat spécifique au TOP1 se plie en forme d’haltère avec un site de clivage TOP1 préféré dans la tige double brin et une séquence de liaison d’amorce dans l’une des deux boucles simple brin. (C) TOP1 est libéré lors de la lyse des cellules. Lors du clivage et de la ligature médiés par TOP1, le substrat est converti en un cercle fermé; (D) du TOP1 recombinant purifié est utilisé. E) Amplification du cercle roulant. Les substrats circulaires fermés sont hybridés à l’amorce ancrée en surface et amplifiés par RCA à l’aide de la polymérase Phi29. Le RCA peut être obtenu soit par incorporation de nucléotides fluorescents comme dans F, soit de nucléotides biotinylés comme dans G. Les produits de cercle roulant fluorescent sont visualisés à l’aide d’un microscope à fluorescence ou d’un scanner à fluorescence. (H) Couplage de l’anticorps anti-biotine conjugué HRP. Les produits à cercle roulant biotinylés sont incubés avec l’anticorps anti-biotine conjugué HRP. Le développement du signal est médié par l’enzyme HRP, et les signaux sont visualisés soit par ECL et détectés à l’aide d’une caméra CCD, soit à l’aide de TMB générant une visualisation colorimétrique. Abréviations : REEAD = détection de l’activité enzymatique améliorée en cercle roulant; RCA = amplification du cercle roulant; TOP1 = topoisomérase 1; HRP = peroxydase de raifort; LMTEC = chimiluminescence accrue; TMB = 3,3',5,5'-tétraméthylbenzidine. Veuillez cliquer ici pour voir une version agrandie de cette figure.
{kind=link}

Figure 2 : Mesures de l’activité TOP1 à l’aide du scanner à fluorescence. Le panneau de gauche montre une image représentative de la diapositive lors de l’analyse de l’activité de 3 à 50 ng de TOP1 à l’aide du protocole de lecture du scanner de fluorescence. Le panneau de droite montre la quantification résultante. test t avec correction de Welch, p = 0,0064, n = 4. Abréviation : TOP1 = topoisomérase 1. Veuillez cliquer ici pour voir une version agrandie de cette figure.
{kind=link}

Figure 3 : Mesures de l’activité du TOP1 à l’aide du microscope à fluorescence. Le panneau de gauche montre des images représentatives de la lame lors de l’analyse de l’activité de 0,1 à 12,5 ng de TOP1 à l’aide du protocole de lecture du microscope à fluorescence. Notez que les images sont des versions recadrées pour afficher correctement les points. Pour cette raison, ils ne ressemblent pas au nombre de signaux dans la quantification indiquée dans le panneau de droite. test t avec correction de Welch, p = 0,0136, n = 3. Abréviation : TOP1 = topoisomérase 1. Veuillez cliquer ici pour voir une version agrandie de cette figure.
{kind=link}

Figure 4 : Mesures de l’activité de la TOP1 à l’aide de la lecture de la chimiluminescence. (A) Le panneau de gauche montre une image représentative de la diapositive lors de l’analyse de l’activité de 3 à 50 ng de TOP1 à l’aide du protocole de lecture de la chimiluminescence. Le panneau de droite montre la quantification résultante. test t avec correction de Welch, p < 0,0001, n = 4. (B) Le panneau de gauche montre une image représentative de la diapositive lors de l’analyse de l’activité de TOP1 extraite de 156 à 40 000 cellules Caco2 à l’aide du protocole de lecture par chimiluminescence. Le panneau de droite montre la quantification résultante. test t avec correction de Welch, p = 0,0224, n = 3. Abréviations : TOP1 = topoisomérase 1. Veuillez cliquer ici pour voir une version agrandie de cette figure.
{kind=link}

Figure 5 : Mesures de l’activité TOP1 à l’aide de la lecture colorimétrique. (A) Le panneau de gauche montre une image représentative de la diapositive lors de l’analyse de 3 à 50 ng de TOP1 à l’aide du protocole de lecture colorimétrique. Le panneau de droite montre la quantification résultante. test t avec correction de Welch, p = 0,0012, n = 4. (B) Le panneau de gauche montre une image représentative de la diapositive lors de l’analyse de l’activité de TOP1 extraite de 156 à 40 000 cellules Caco2 à l’aide du protocole de lecture colorimétrique. Le panneau de droite montre la quantification résultante. test t avec correction de Welch, p = 0,0117, n = 3. Abréviations : TOP1 = topoisomérase 1; TMB = 3,3',5,5'-tétraméthylbenzidine. Veuillez cliquer ici pour voir une version agrandie de cette figure.
{kind=link}

Figure 6 : Mesures de l’activité de TOP1 après traitement médicamenteux à l’aide des protocoles de lecture par chimiluminescence. Le panneau de gauche montre une image représentative de la diapositive lors de l’analyse de 10 ng de TOP1 en présence de 5% de DMSO ou 80 μM CPT pendant 1 min. Le panneau de droite montre la quantification résultante. test t avec correction de Welch, p = 0,0017, n = 3. Abréviations : TOP1 = topoisomérase 1; CPT = camptothécine; DMSO = diméthylsulfoxyde; Neg = contrôle négatif. Veuillez cliquer ici pour voir une version agrandie de cette figure.
{kind=link}
Discussion
Les topoisomérases représentent une classe d’enzymes modifiant l’ADN qui suscitent un intérêt scientifique et de recherche élevé, étant la cible de composés à petites molécules ayant un effet potentiel dans le traitement anticancéreux ou dans la lutte contre les maladies infectieuses. De plus, l’activité du TOP1 humain s’est avérée être un biomarqueur efficace du pronostic et du traitement du cancer18,19,23. Bien que ce soit l’activité TOP1 qui influence l’efficacité d’un inhibiteur chimiothérapeutique26, c’est souvent la quantité d’ADN-ARN ou la quantité de TOP1 qui est évaluée en milieu clinique. Cela est dû au manque d’outils gratuits rapides, faciles et à base de gel qui peuvent fournir une quantification précise et précise de l’activité topoisomérase dans tous les environnements de laboratoire.
Ici, un protocole pour le test REEAD permettant la mesure de l’activité TOP1 de manière sans gel est décrit. Dans ce protocole, le TOP1 purifié ou extrait brut de cellules est incubé avec un substrat d’ADN spécialement conçu en forme d’haltère qui, lors du clivage / ligature médié par TOP1, est converti en une molécule circulaire fermée. Les cercles sont ensuite hybridés sur une surface vitrée et amplifiés par RCA. Pour faciliter l’exécution des réactions se produisant sur la lame, une grille en silicone est fixée au verre, créant des puits individuels où les réactions ont lieu. De cette façon, le protocole tire parti d’un système multipuits - une grille en silicone - appelé wellmaker.
Le protocole a été utilisé pour détecter l’activité des TOP1 et TOP1 recombinants extraits des cellules d’adénocarcinome colorectal Caco2, utilisés à titre d’exemple. De plus, à titre d’exemple de REEAD à utiliser comme outil de dépistage de médicaments, le protocole a été utilisé pour détecter l’inhibition de l’activité de TOP1 par l’inhibiteur bien connu de TOP1 CPT24,25. Le test a été couplé à différentes méthodes de lecture - la microscopie à fluorescence, qui donne une sensibilité élevée, et un scanner à fluorescence, chimiluminescence ou colorimétrique, qui nécessitent un équipement et une formation moins spécialisés. La lecture du microscope à fluorescence très sensible a les limites de la nécessité d’un réglage de microscope à fluorescence de bonne qualité, d’un personnel qualifié et d’une acquisition et d’une analyse d’images chronophages.
Pour ces raisons, la lecture du scanner de fluorescence qui permet une acquisition et une analyse plus rapides, même au détriment de la sensibilité, est présentée. En l’absence d’un scanner à fluorescence, deux excellentes alternatives, la chimiluminescence et les méthodes de lecture colorimétrique, peuvent être envisagées. Les deux méthodes sont rapides et simples, ne nécessitant aucun équipement coûteux ou formation spécialisée. Dans tous les formats de lecture, REEAD présente de nombreux avantages par rapport aux tests de pointe, qui sont plus longs, à base de gel (avec les exigences des agents intercalants) et moins directement quantitatifs. Cependant, il y a quelques étapes critiques dans le protocole présenté. Lors de la manipulation du dépistage du médicament, les points temporels, la concentration du médicament et le rapport quantité de TOP1/substrat d’ADN doivent être optimisés par rapport aux paramètres décrits optimisés pour la CPT. Le médicament peut être étudié en effectuant une préincubation avec le substrat d’ADN ou une préincubation avec l’enzyme TOP1. Cela peut donner des informations précieuses sur la capacité de la molécule à inhiber TOP1 et offrir un aperçu du mécanisme d’action du médicament.
De plus, si les signaux sont faibles ou absents, cela est probablement dû à une lyse inefficace de l’échantillon brut ou à une dégradation de TOP1 dans l’extrait en raison d’une mauvaise utilisation des inhibiteurs de la protéase. En outre, cela peut également être dû à un stockage inadéquat des composants importants du test, tels que la TOP1 recombinante, la phi29 polymérase, les nucléotides, le substrat et l’apprêt. Enfin, les cycles répétés de gel-dégel des oligonucléotides doivent être évités, car cela affecte considérablement les performances du test. Lorsque l’extrait brut est utilisé, l’efficacité d’extraction de l’échantillon biologique spécifique doit être optimisée et la quantité et l’activité de TOP1 peuvent différer de l’exemple rapporté ici. Pour cette raison, chaque extrait brut à tester nécessitera un titrage pour l’identification de la limite de détection et de la plage de sensibilité de l’essai lors de l’utilisation de cet échantillon particulier. Le protocole a été optimisé pour la détection de TOP1 humain. L’activité d’autres TOP1 eucaryotes peut être mesurée, mais la concentration de NaCl et le temps d’incubation peuvent devoir être optimisés en fonction de la méthode de purification enzymatique et de l’activité enzymatique optimale. Un plus grand nombre de substrats circularisés résulteront d’une incubation prolongée, tandis que moins de substrats circularisés résulteront d’une incubation raccourcie.
En plus des applications présentées, REEAD permet la mesure de l’activité TOP1 dans des extraits bruts de petites biopsies de patients cancéreux18, la prédiction de l’effet anticancéreux cytotoxique de la CPT dans les lignées cellulaires cancéreuses 20,21,22, et même la détection de l’activité enzymatique dans des cellules individuelles20,21 . De plus, le réglage REEAD présenté à l’aide du wellmaker permet un criblage multipuits de médicaments de bibliothèques de composés synthétiques ou naturels. En outre, une version modifiée du test REEAD, appelée REEAD C/L, a été développée. Cette configuration permet d’étudier séparément les étapes de clivage et de ligature de la réaction TOP127. Avec le REEAD C/L, il est possible de déterminer le mécanisme d’action des inhibiteurs de petites molécules et de les caractériser comme inhibiteurs catalytiques TOP1 ayant des effets antiparasitaires potentiels28 ou comme poisons TOP1 à utiliser pour le traitement anticancéreux24,25. Enfin, avec la refonte spécifique des substrats d’ADN pour répondre aux différentes exigences (pour la liaison à l’ADN ou le clivage-ligature) d’autres enzymes TOP1, REEAD a également été utilisé pour la détection de maladies infectieuses (comme dans le cas de Plasmodium falciparum, causant le paludisme29), ou Leishmania donovani et du virus de l’orthopoxvirose simienne (données non présentées). Ou, il peut être utilisé pour identifier de petites molécules avec des effets antipathogènes. Le protocole présenté fournit aux scientifiques du domaine TOP1 une méthode simple pour détecter l’activité enzymatique avec peu ou pas d’optimisation et avec la possibilité d’être adapté à des applications encore plus larges à l’avenir.
Déclarations de divulgation
Les auteurs C.T. et K.M. sont des employés de VPCIR biosciences ApS. C.T., B.R.K. et M.S. sont actionnaires et/ou détenteurs d’options d’achat d’actions. C.T., B.R.K., et M.S. déclarent être les inventeurs nommés du brevet EP2022/057172 déposé au nom de VPCIR biosciences ApS. Les autres auteurs déclarent qu’ils n’ont pas d’intérêts concurrents.
Remerciements
Les auteurs tiennent à remercier Noriko Y. Hansen, technicienne de laboratoire du Département de biologie moléculaire et de génétique de l’Université d’Aarhus, pour son assistance technique en ce qui concerne les purifications enzymatiques Phi29 et TOP1.
matériels
| Name | Company | Catalog Number | Comments |
| Equipment | |||
| Camera for fluorescence image analysis | |||
| CCD camera | For chemiluninescence image analysis | ||
| Fluorescence microscope | Olympus | Equipped with 60x oil immersion objective and a GFP filter ( https://www.edmundoptics.com/p/gfp-filter-cube-set-olympus/21527/) | |
| Fluorescence scanner | Typhoon | FLA 9500 | Equipped with 473 laser and FAM filter |
| Humidity chamber with dH2O | |||
| Hybridization chamber with saturated NaCl | |||
| ImageJ software | Fiji | Download at: https://imagej.net/software/fiji/downloads | |
| Materials | |||
| Cell culture | |||
| Cell growth medium appropiate for the chosen cell line | |||
| Trypsin | Sigma | #T4174 | |
| Pen strep | Sigma | #P4300 | |
| Tissue culture flasks | ThermoFisher | #178983 | |
| FBS | Gibco | #10270106 | |
| NEEA | Sigma | #M7145 | |
| PBS | ThermoFisher | #10010023 | |
| Functionalized slides | |||
| 5'-amine anti-TOP1 primer | Sigma | 5’-/5AmMC6/CCA ACC AAC CAA CCA AAT AAG-3’ | |
| Activated Codelink HD slides | SurModics | #DHD1-0023 | |
| Silicon grid | Grace-biolabs | Custom made | |
| Pertex glue | Histolabs | #00801 | |
| Microscope slide 76 x 26 mm | Hounisen | #2510 1201BL | Other microscope slide of same dimension can be used |
| DNA circles and rolling circle amplification | |||
| TOP1 specific substrate | Sigma | 5’-AGA AAA ATT TTT AAA AAA ACT GTG AAG ATC GCT TAT TTT TTT AAA AAT AAA TCT AAG TCT TTT AGA TCC CTC AAT GCA CAT GTT TGG CTC CGA TCT AAA AGA CTT AGA-3’ | |
| ATTO-488-dUTP | Jena Biosciences | #95387 | |
| Biotin-dCTP | Jena Biosciences | #NU-809-BIO16L | |
| dNTP | ThermoFisher | #R0181 | |
| Phi29 polymerase | VPCIR Biosciences | #10010041 | |
| Recombinant TOP1 | VPCIR Biosciences | #10010001 | |
| Detection of rolling circle products | |||
| BioFX TMB | SurModics | #ESPM-0100-01 | |
| Cover glass | |||
| ECL mixture | Cytiva | #RPN2236 | |
| HRP conjugated anti-biotin antibody | Merck | #A4541 | |
| Mounting medium (vectachield) | Vector laboratories | #H-1000 | |
| Buffer list | |||
| 1x PE Buffer | 1 mM EDTA, 8.12 mM Na2HPO4·2H2O, 1.88 mM NaH2PO4·H2O | ||
| 6x Print Buffer | 300 mM Na3PO4, pH 8.5 | ||
| Blocking solution | Buffer 5 supplemented with 5% non-fat milk and 5% BSA pH 9 | ||
| Lysis Buffer | 10 mM Tris-HCl pH 7.5, 1 mM EDTA + protease inhibitors | ||
| 10x TOP1 Reaction Buffer | 50 mM CaCl2, 50 mM MgCl2, 100 mM Tris-HCl pH 7.5 | ||
| 10x Phi29 Reaction Buffer | 330 mM Tris-acetate pH 7.5, 100 mM Mg-acetate, 660 mM K-acetate, 1% Tween20, 200 mM DTT | ||
| Buffer 1 | 50 mM Tris, 50 mM Tris-HCl, 32 mM ethanolamine. NB: stored at 50 °C. | ||
| Buffer 2 | 4x SSC, 0.1% SDS. NB: stored at 50 °C. | ||
| Buffer 3 | 100 mM Tris-HCl pH 7.5, 150 mM NaCl, 0.3% SDS | ||
| Buffer 4 | 100 mM Tris-HCl pH 7.5, 150 mM NaCl, 0.05% Tween20 | ||
| Buffer 5 | 20 mM Tris-HCl pH 7.5, 150 mM NaCl, 0.05% Tween20 | ||
| Chemicals for buffers | |||
| Tris | Sigma | #T1506 | |
| HCl | Sigma | #1.00317.2011 | |
| EDTA | Sigma | #1.08418.1000 | |
| PMSF | Sigma | #05056489001 | |
| SDS | Applichem | #436143 | |
| Na2HPO4 | Sigma | #1.06580.1000 | |
| NaH2PO4 | Sigma | #1.06346.1000 | |
| Ethanolamine | Sigma | #411000 | |
| SSC | Invitrogen | #15557-036 | |
| Tris-acetate | Sigma | #T1258 | |
| Mg-acetate | Sigma | #M0631 | |
| K-acetate | Sigma | #1.04820.1000 | |
| Tween20 | Sigma | #8.22184.0500 | |
| DTT | Sigma | #D0632 | |
| CaCl2 | Sigma | #1.02382.1000 | |
| MgCl2 | Sigma | #M2670 | |
| NaCl | Sigma | #1.06404.5000 | |
| Skimmed milk powder | Sigma | #70166 | |
| BSA | Sigma | #A4503 |
Références
- Baldwin, E. L., Osheroff, N. Etoposide, topoisomerase II and cancer. Current Medicinal Chemistry-Anti-Cancer Agents. 5 (4), 363-372 (2005).
- Gilbert, D. C., Chalmers, A. J., El-Khamisy, S. F. Topoisomerase I inhibition in colorectal cancer: Biomarkers and therapeutic targets. British Journal of Cancer. 106 (1), 18-24 (2012).
- Ikeguchi, M., et al. TopoisomeraseI expression in tumors as a biological marker for CPT-11 chemosensitivity in patients with colorectal cancer. Surgery Today. 41 (9), 1196-1199 (2011).
- Meisenberg, C., et al. Clinical and cellular roles for TDP1 and TOP1 in modulating colorectal cancer response to irinotecan. Molecular Cancer Therapeutics. 14 (2), 575-585 (2015).
- Palshof, J. A., et al. Topoisomerase I copy number alterations as biomarker for irinotecan efficacy in metastatic colorectal cancer. BMC Cancer. 17 (1), 1-10 (2017).
- Proszek, J., et al. Topoisomerase I as a biomarker: Detection of activity at the single molecule level. Sensors. 14 (1), 1195-1207 (2014).
- Leppard, J. B., Champoux, J. J. Human DNA topoisomerase I: Relaxation, roles, and damage control. Chromosoma. 114 (2), 75-85 (2005).
- Champoux, J. J. DNA topoisomerases: structure, function, and mechanism. Annual Review of Biochemistry. 70 (1), 369-413 (2001).
- Redinbo, M. R., et al. Crystal structures of human topoisomerase I in covalent and noncovalent complexes with DNA. Science. 279 (5356), 1504-1513 (1998).
- Nitiss, J. L., Soans, E., Rogojina, A., Seth, A., Mishina, M. Topoisomerase assays. Current Protocols in Pharmacology. 57 (1), 3(2012).
- Tesauro, C., et al. Erybraedin C, a natural compound from the plant Bituminaria bituminosa, inhibits both the cleavage and religation activities of human topoisomerase I. Biochemical Journal. 425 (3), 531-539 (2010).
- Keller, J. G., et al. Topoisomerase 1 inhibits MYC promoter activity by inducing G-quadruplex formation. Nucleic Acids Research. 50 (11), 6332-6342 (2022).
- Christiansen, K., Westergaard, O. Characterization of intra- and intermolecular DNA ligation mediated by eukaryotic topoisomerase I. Role of bipartite DNA interaction in the ligation process. Journal of Biological Chemistry. 269 (1), 721-729 (1994).
- Svejstrup, J. Q., Christiansen, K., Andersen, A. H., Lund, K., Westergaard, O. Minimal DNA duplex requirements for topoisomerase I-mediated cleavage in vitro. Journal of Biological Chemistry. 265 (1), 12529-12535 (1990).
- Anand, J., Sun, Y., Zhao, Y., Nitiss, K. C., Nitiss, J. L. Detection of topoisomerase covalent complexes in eukaryotic cells. Methods in Molecular Biology. , Clifton, NJ. 283-299 (2018).
- Stougaard, M., et al. Single-molecule detection of human topoisomerase I cleavage-ligation activity. ACS Nano. 3 (1), 223-233 (2009).
- Keller, J. G., Petersen, K. V., Knudsen, B. R., Tesauro, C. Simple and fast DNA-based tool to investigate topoisomerase 1 activity, a biomarker for drug susceptibility in colorectal cancer. Recent Underst. Colorectal Cancer Treatment. , (2022).
- Jakobsen, A. K., et al. Correlation between topoisomerase I and tyrosyl-DNA phosphodiesterase 1 activities in non-small cell lung cancer tissue. Experimental and Molecular Pathology. 99 (1), 56-64 (2015).
- Jakobsen, A. K., et al. TDP1 and TOP1 as targets in anticancer treatment of NSCLC: Activity and protein level in normal and tumor tissue from 150 NSCLC patients correlated to clinical data. Lung Cancer. 164, 23-32 (2022).
- Keller, J. G., et al. On-slide detection of enzymatic activities in selected single cells. Nanoscale. 9 (36), 13546-13553 (2017).
- Keller, J. G., Stougaard, M., Knudsen, B. R. Enzymatic activity in single cells. Trends in Biotechnology. 29 (5), 222-230 (2019).
- Tesauro, C., et al. Different camptothecin sensitivities in subpopulations of colon cancer cells correlate with expression of different phospho-isoforms of topoisomerase i with different activities. Cancers. 12 (5), 1240(2020).
- Tesauro, C., et al. Topoisomerase i activity and sensitivity to camptothecin in breast cancer-derived cells: A comparative study. BMC Cancer. 19 (1), 1-15 (2019).
- Pommier, Y. DNA topoisomerase I Inhibitors: Chemistry, biology, and interfacial inhibition. Chemical Reviews. 109 (7), 2894-2902 (2009).
- Pommier, Y. Drugging topoisomerases: lessons and challenges. ACS Chemical Biology. 8 (1), 82-95 (2013).
- Bailly, C. Irinotecan: 25 years of cancer treatment. Pharmacological Research. 148, 104398(2019).
- Petersen, K. V., et al. Simple and fast dna based sensor system for screening of small-molecule compounds targeting eukaryotic topoisomerase 1. Pharmaceutics. 13 (8), 1255(2021).
- García-Estrada, C., Prada, C. F., Fernández-Rubio, C., Rojo-Vázquez, F., Balaña-Fouce, R. DNA topoisomerases in apicomplexan parasites: promising targets for drug discovery. Proceedings of the Royal Society B: Biological Sciences. 277 (1689), 1777-1787 (2010).
- Hede, M. S., et al. Detection of the malaria causing Plasmodium parasite in saliva from infected patients using topoisomerase I activity as a biomarker. Scientific Reports. 8 (1), 1-12 (2018).
Réimpressions et Autorisations
Demande d’autorisation pour utiliser le texte ou les figures de cet article JoVE
Demande d’autorisationThis article has been published
Video Coming Soon
À PROPOS DE JoVE
Copyright © 2025 MyJoVE Corporation. Tous droits réservés.