
Method Article
2 ב 1: בשלב אחד טיהור זיקה לניתוח מקבילים של חלבון, מתחמי חלבון-מטבוליט
* These authors contributed equally
In This Article
Summary
אינטראקציות חלבון-חלבון, חלבון-מטבוליט מכריעים עבור כל פונקציות הסלולר. במסמך זה, אנו מתארים את פרוטוקול המאפשר ניתוח מקביל של אינטראקציות אלה עם חלבון של בחירה. פרוטוקול שלנו היה אופטימיזציה עבור הצמח תרביות תאים, והוא משלב טיהור זיקה עם חלבון מבוסס ספקטרומטר מסה וזיהוי מטבוליט.
Abstract
תהליכים תאיים מוסדרים על ידי אינטראקציות בין מולקולות ביולוגיות כגון חלבונים, מטבוליטים ו חומצות גרעין. אמנם אין חידוש החקירה של אינטראקציות חלבון (PPI), גישות ניסיוניות מכוון לאפיון אינטראקציות חלבון-מטבוליט אנדוגני (PMI) מהווים התפתחות למדי. במסמך זה, אנו מציגים פרוטוקול המאפשר אפיון סימולטני של PPI, PMI של חלבון של בחירה, המכונה פיתיון. פרוטוקול שלנו היה מותאם עבור תא תודרנית תרבויות, והוא משלב זיקה טיהור (AP) עם ספקטרומטר מסה (MS)-המבוסס על זיהוי חלבונים, מטבוליט. בקיצור, קווים תודרנית מהונדס, המבטאים חלבונים פיתיון דבוקה תג קירבה, הם קודם lysed להשיג תמצית הסלולר מקורית. נוגדנים אנטי-תג משמשים כדי להוריד את השותפים חלבון ו מטבוליט של החלבון פיתיון. מתחמי מטוהרים זיקה מופקים באמצעות מתיל טרטצעד אחד-בוטיל אתר (MTBE) / מתנול/מים בשיטה. בעוד מטבוליטים להפריד את קוטבי או השלב הידרופוביות, ניתן למצוא חלבונים בגדר. מטבוליטים והן חלבונים מכן נותחו על ידי אולטרה-ביצועים נוזלי כרומטוגרפיה-ספקטרומטריית (UPLC-MS או UPLC-MS/MS). ריק-וקטור (EV) בקרת קווי משמשים כדי לא לכלול תוצאות חיוביות שגויות. היתרון העיקרי של פרוטוקול שלנו הוא שהם מאפשרים זיהוי של שותפים חלבון ו מטבוליט של חלבון המטרה במקביל בתנאים ליד-פיזיולוגיים (סלולריים lysate). השיטה הציג היא פשוטה, מהירה, ניתן להתאים בקלות למערכות ביולוגיות חוץ תרביות תאים הצמח.
Introduction
השיטה המתוארים כאן מטרות על הזיהוי של המטבוליט וחלבון השותפים של חלבון של בחירה בתנאים lysate הסלולר ליד -ב- vivo . זה כבר העריכו שיש רבים מטבוליטים יותר מאשר מאופיין היום של פונקציית רגולציה חשוב1. מטבוליטים יכול לשמש מתגים ביולוגי, לשנות את הפעילות, פונקציונליות, ו/או לוקליזציה של שלהם קולטן חלבונים2,3,4. בעשור האחרון מספר שיטות פריצת דרך, המאפשרת זיהוי של PMI ויוו או בתנאים ליד -ב- vivo , כבר מפותחת5. גישות זמין ניתן להפריד לשתי קבוצות. הקבוצה הראשונה כוללת טכניקות להתחיל עם פיתיון ידוע-מטבוליט על מנת ללכוד חלבון הרומן שותפים. שיטות כוללות זיקה כרומטוגרפיה6, סמים זיקה היעד מגיב-יציבות assay7, כימותרפיה-פרוטאומיקס8ו פרוטאום תרמית פרופיל9. הקבוצה השנייה מורכבת שיטה אחת שמתחילה חלבון ידוע על מנת לזהות מולקולה קטנה ליגנדים10,11.
AP בשילוב עם lipidomics מבוססי MS שימש כדי לנתח חלבון-השומנים מתחמי האפייה12. כנקודת התחלה, המחברים השתמשו זני שמרים לבטא 21 אנזימים המעורבים ביוסינטזה ergosterol, 103 kinases דבוקה הוא לטיהור טנדם-זיקה (ברז) תג. 70% של האנזימים ו-20% kinases נמצאו לאגד ליגנדים הידרופובי שונים, ושופך אור לתוך הרשת אינטראקציית חלבון מורכב-השומנים.
בעבר, יכולתי נדגים, באופן דומה כדי שומנים, תרכובת פולאר polar למחצה גם להישאר מאוגד ל קומפלקסי חלבון מבודד lysates הסלולרית13. על סמך ממצאים אלה, החלטנו למטב את AP שיטת שפורסם בעבר10,11 תאי צמחים, תרכובות הידרופילית14. למטרה זו, השתמשנו ברז וקטורים, מתוארת על ידי ואן Leene. ואח 2010, משתמשים בהצלחה במפעל PPI מחקרים15. כדי לקצר את משך הזמן הנדרש להשגת שורות מהונדס, החלטנו על תרביות תאים תודרנית. אנחנו מועסקים מתיל טרטצעד אחד-בוטיל אתר, (MTBE) / מתנול/מים החילוץ שיטה, המאפשר אפיון חלבונים (גלולה), ליפידים (שלב אורגני) ו הידרופילית מטבוליטים (פאזה מימית)16 יחיד זיקה-טיהור הניסוי. EV בקרת קווי הוכנסו כדי לא לכלול תוצאות חיוביות שגויות, כגון חלבונים מחייב את התג לבד. כמו הוכחת לנו מתויגות שלוש (מתוך חמש) nucleoside diphosphate kinases להציג הגנום תודרנית (NDPK1-NDPK3). בין השאר, אנחנו יכול להדגים כי NDPK1 אינטראקציה עם גלוטתיון S-טרנספראז גלוטטיון. כתוצאה מכך נוכל להוכיח כי NDPK1 הוא נתון glutathionylation14.
לסיכום, פרוטוקול הציג היא כלי חשוב עבור אפיון חלבון ורשתות אינטראקציית חלבון-מולקולה קטנה ומהווה התקדמות גדולה על פני שיטות קיימות.
Protocol
הכנת הטרנסגניים תודרנית תרבות שורות תאים, כולל שכפול, שינוי, בחירה, וצמיחה תנאים ניתן למצוא ב-17. הערה EV בקרת קווי המומלצים לתיקון תוצאות חיוביות שגויות. לפני הניסוי, לאשר את ביטוי של החלבון הפיתיון על ידי ניתוח תספיג חלבון, למשל באמצעות נוגדנים IgG נגד החלק חלבון G של התג זיקה טנדם. חשוב להפריד את המדיה צמיחה חומר התרבות של תא צמחי.
1. הכנת חומר צמחי תא לפני הניסוי
- לגדול קו תרבות תא PSB-L לבנה א overexpressing החלבון של עניין18.
- להכין MSMO בינוני, אשר מכיל 4.43 g/L ש-MSMO מעורבב עם סוכרוז 30 g/L. להתאים את ה-pH של המאגר כדי 5.7 עם 1 מ' KOH ואת אוטוקלב הפתרון. לפני הניסוי, תוספת המדיום עם חומצה α-naphthaleneacetic 0.5 מ ג/ליטר, kinetin 0.05 mg/L ו- 50 kanamycin μg/mL.
- לטפח צמח טרנספורמציה תרביות תאים ב- 50 מ של MSMO בינוני בבקבוקון 100 מ ב- פלטפורמה מסלולית בשייקר עם עצבנות עדין (130 סל ד). לגדול בתאים חדר התרבות ב- 20 ° C ועוצמת האור השווה ל- 80 μmol m s-2 -1.
- תת-תרבות תאים לתוך המדיה טריים מדי 7 ימים, דילול אותם 1:10.
- איסוף תאים בשלב הצמיחה לוגריתמי באמצעות משפך זכוכית בשילוב עם משאבת ואקום, באמצעות רשת ניילון כמו מסנן. לעטוף לחדור בנייר אלומיניום, להקפיא במצב חנקן נוזלי.
שים לב: זכור כי בחנקן נוזלי הוא קר מאוד. טיפול שגוי עלול לגרום לכוויות. ללבוש ציוד מגן אישי המתאים, לרבות תרמית מבודדים כפפות, משקפי מגן, חלוק מעבדה.
2. הקש על פרוטוקול
הערה: בשלב הבא מותאמת מ 2014 ואח מיאדה11 וואן Leene et al. 201117.
- Homogenize חומר התרבות של תא צמחי שנקטפו וקפואים באמצעות מערבל מיל (2 דקות ב 20 הרץ) או ומכתש כדי להשיג אבקה. Aliquot דור 3 של החומר הקרקע (תואם לבסביבות 90 מ"ג חלבון סה כ) עבור דגימה. הימנע הפשרתו של המדגם במהלך שלב זה באמצעות חנקן נוזלי ציוד מראש צוננת.
הערה: חנות הקרקע חומר צמחי בצינור 50 מ ב- –80 ° C עד תחילתו של הליך AP. - Triturate הדגימה במרגמה נוזלי-חנקן-precooled ' עם 3 מ"ל של המאגר הקר פירוק (0.025 M טריס-HCl pH 7.5; 0.5 M NaCl 1.5 mM MgCl20.5 מ מ DTT; 1 מ"מ NaF; 1 מ מ נה3VO4; 100 x מעכב פרוטאז מסחרי מדולל קוקטייל; 1 מ מ PMSF) עד מפשירה החומר. ברגע המדגם מפשירה, ומיד להמשיך לשלב הבא.
הערה: להכין מאגר פירוק הטרייה. להציג דוגמאות ריק בשלב זה. חומרי ניקוי מומלץ לא כפי שהם יכולים לגרום בעיות בזיהוי MS. - כדי להסיר את הלכלוך הסלולר, לחלק את החומר צינורות microcentrifuge 2 מ"ל, צנטריפוגה ב 20,817 x g 10 דקות ב 4 º C. לאסוף 3 מ"ל של lysate ברור בשפופרת צנטרפוגה חרוט 15-mL.
- במהלך שלב צנטריפוגה, equilibrate איג-Sepharose חרוזים. Aliquot 100 µL של החרוזים לכל דגימה ולשטוף אותם עם 1 מ"ל פירוק המאגר. המערבולת resuspend פלאש-ספין ושרשרות. לבטל את המאגר פירוק, חזור על השלב פעמיים. Resuspend חרוזים ב 400 µL פירוק המאגר.
- להוסיף חרוזים הצמח שנאספו lysate, דגירה את התערובת על גלגל מסתובב עבור h 1-4 מעלות צלזיוס.
- להעביר התערובת לתוך מזרק בשילוב באמצעות סכינים סטריליים-lock כובע עם ספין עמודה עם מסנן נקבוביות בגודל 35 מיקרומטר. החל לחץ כדי לעבור את lysate דרך. חרוזים עם מתחמי המצורפת יישאר על המסנן, ואילו lysate יעבור דרך.
הערה: לחלופין, השתמש במערכת סעפת ואקום. לוודא להפעיל לחץ עדין כדי לא לפגוע החרוזים. - לשטוף חרוזים בהתחלה עם 10 מ"ל לרחוץ את מאגר (0.025 M טריס-HCl pH 7.5; 0.5 M NaCl), ואז עם 1 מ"ל • תנאי מאגר (10 מ מ טריס-HCl pH 7.5; 150 מ מ NaCl; EDTA 0.5 מ מ 1000-E64 ו- 1 מ מ PMSF מדולל). לבצע שטיפה באמצעות מזרק מצורף העמודה או סעפת מערכת ואקום.
הערה: בעת שימוש במערכת סעפת ואקום גורם מסויימים להחלת לחץ עדין כדי לא לפגוע החרוזים. - דגירה חרוזים עם ה 400 µL • תנאי מאגר המכיל 50 U של גרסה משופרת של הטבק לחרוט פרוטאז וירוס (AcTEV). להשתמש בטבלה שאכר 1,000 סל ד למשך 30 דקות ב-16 ° c
הערה: זכור להשתמש פקק כדי לסגור את העמודה בתחתית הוספת המאגר • תנאי. - הוסף של חלק נוסף (50 U) של האנזים לתוך העמודה, דגירה את התערובת הבאה 30 דקות תחת אותו, התנאים לעיל.
- לאסוף את eluate בשפופרת 2 מ"ל microcentrifuge על-ידי צנטריפוגה (1 דקות, 20,817 x g) או יריעה ואקום. כדי להסיר את מתחמי הנותרים, להציג את שלב נוסף • תנאי שימוש µL 200 • תנאי המאגר.
הערה: אחסן את הדגימה מהנהר או –80 ° C, או להמשיך לשלב מיצוי חלבונים ו מטבוליט. הפשרת דגימות קפוא בקרח.
3. תספיג ניתוח
- כדי לאשר את הנוכחות של הפיתיון חלבון eluate שנאספו להשתמש 10 µL של eluate המכילים חלבון – מטבוליט לביצוע מרחביות – דף וניתוח תספיג חלבון. כדי לזהות את החלבון עניין, להשתמש עכבר ראשי נוגדנים נגד streptavidin מחייב חלבון (1:200), חלק התג ברז שנותרו לאחר אס פרוטאז מחשוף, כפי שמתואר ואן Leene. ואח 201117. בשלב הבא, השתמש נוגדנים אנטי עכבר עז משני בשילוב עם HRP.
4. מטבוליט והפקת חלבונים
הערה: פרוטוקול זה מותאמת מ. ואח 2011 Giavalisco16.
הערה: משלב זה ואילך להשתמש בפתרונות UPLC – MS – כיתה.
- להוסיף 1 מ"ל של מתיל טרט-בוטיל אתר (MTBE) / מתנול/למים הממס (3:1:1) eluate שנאספו ומערבבים את הדגימה על-ידי היפוך. ודא כי הממיס הוא מקורר מהנהר לפני השלב החילוץ.
התראה: MTBE מתנול הינם חומרים מזיקים. בצע את שלב החילוץ מתחת למכסה המנוע fume וללבוש ציוד למיגון אישי המתאים, כגון: כפפות. - להוסיף מ 0.4 ל מתנול: מים 1:3 פתרון כל מדגם ומערבבים את התוכן של המדגם על-ידי היפוך.
הערה: תוספת של התערובת עם מתנול: מים פתרון תוצאות שלב ההפרדה. השלב העליון מכיל שומנים, שלב התחתונה מכילה מטבוליטים קוטב ו למחצה קוטבי, חלבונים מצוי בגדר. - כדי להפריד בין שלבים, centrifuge את הדגימה ב 20,817 x g למשך 2 דקות בטמפרטורת החדר, ואז לאסוף את השלב העליון של מידות השומנים (לא עושים את פרוטוקול זה) באמצעות פיפטה טיפול בנוזלים ידנית עם קיבולת נפח 1 מ"ל.
- להוסיף 0.2 מ"ל של מתנול, מיקס על ידי היפוך.
- Centrifuge הדגימה ב g x 20,817 למשך 2 דקות ב RT ולאחר מכן לאסוף את השלב קוטבי מטבוליט מעבדתיים (תרכובת פולאר polar למחצה). כדי להימנע מהפרעה בגדר חלבונים, השאר µL בסביבות 50 של שלב נוזלי בתחתית הצינורית.
- הדוגמאות שנאספו יבש למדידות מטבוליט לינה המאייד צנטריפוגלי. למנוע ייבוש יתר על המידה את כדורי חלבונים על-ידי הסרת דגימות המאייד לאחר 30-60 דקות.
הערה: דגימות מהנהר או –80 ° C, או להמשיך מיד עם הכנת חלבונים לניתוח LC-MS/MS.
5. מכינים את דוגמאות לניתוח פרוטיאומיה מבנית
הערה: השלב זה ממאמרו של אולסן. et al. 200419 ו המדריך הטכני של המיקס טריפסין/ליס-C (ראה טבלה של חומרים).
- לבצע עיכול אנזימטי של המדגם.
התראה: ממיסים בשימוש במהלך עיכול אנזימטי, desalting של המדגם מזיקים. מתחת למכסה המנוע fume וללבוש ציוד למיגון אישי המתאים, כגון: כפפות.- להמיס חלבון גלולה ב µL 30 מאגר דנטורציה הטרי (40 מ מ אמוניום ביקרבונט המכילה אוריאה thiourea/6 מ' 2 מ', pH 8). כדי להשיג מסיסות חלבון טוב יותר, לבצע צעד sonication 15-מין. חזור על השלב עד בגדר מתמוסס.
- Centrifuge הדגימה ב 20,817 g x 10 דקות ב 4 ° C, ולאחר מכן להעביר את תגובת שיקוע צינור microcentrifuge.
- לקבוע ריכוז חלבון באמצעות את שיטת ברדפורד.
- עבור aliquot בבדיקות נוספות של אמצעי אחסון שקול ל- 100 µg של חלבון ולמלא את הדגימה עד 46 µL עם מאגר דנטורציה.
- להוסיף µL מדגם 2 הפחתת הטרי מאגר (50 מ"מ DTT מומס H2O), תקופת דגירה של 30 דקות בטמפרטורת החדר.
- לטיפול הדגימה עם 2 µL אלקילציה הטרי מאגר (150 מ מ iodoacetamide התפרקה במאגר אמוניום ביקרבונט 40 מ מ), דגירה התערובת בחושך למשך 20 דקות בטמפרטורת החדר.
- לדלל את הדגימה עם µL 30 40 מ מ אמוניום ביקרבונט המאגר ולהוסיף 20 µL של LysC/טריפסין מיקס.
- לאחר אינקובציה 4 h ב 37 מעלות צלזיוס, לדלל את הדגימה עם 300 µL מאגר אמוניום ביקרבונט 40 מ מ.
- להמשיך עם הדגירה לילה ב 37 º C.
- Acidify הדגימה כ-20 µL של 10% חומצה trifluoroacetic (TFA) כדי להשיג את ה-pH < 2. בדוק מדגם pH באמצעות רצועה pH.
הערה: המדגם-מהנהר או להמשיך לשלב הבא.
- Desalt את החלבונים מעוכל.
הערה: רצוי, השתמש במערכת סעפת ואקום. למנוע ייבוש יתר של העמודה.- לשטוף את העמודה סי18 SPE (ראה טבלה של חומרים) עם 1 מ"ל של 100% MeOH ולאחר מכן עם 1 מ"ל של 80% acetonitrile (לחצן מצוקה) המכיל 0.1% TFA מדולל במים. השימוש, כאן ולא ב עוד יותר חלבון desalting צעדים, סעפת מערכת ואקום כדי להאיץ את התהליך. למנוע ייבוש יתר של העמודה.
- Equilibrate לעמודה ע י שטיפת זה פעמיים עם 1 מ"ל של 0.1% TFA מדולל במים.
- לטעון את הדגימה אל העמודה. צינור שטיפה עם µL 200 נוספים של 0.1% TFA והעברת הפתרון על העמודה. הפעל את הפתרונות באמצעות המדור.
- לשטוף את העמודה פעמיים עם 1 מ"ל של 0.1% TFA.
- Elute desalted פפטידים מהעמוד עם 800 µL 60% לחצן מצוקה, פתרון TFA 0.1%. יבש את השבר שנאספו המאייד צנטריפוגלי, הימנעות ייבוש יתר של השבר חלבונים על-ידי הסרת דגימות המאייד לאחר 30-60 דקות.
הערה: דגימות מהנהר או –80 ° C או להמשיך לשלב הבא. בשלבים הבאים, לשמור דגימות על קרח.
6. מדידת הכין דגימות חלבון באמצעות UPLC – MS/MS.
הערה: לפני מדידות פרוטיאומיה מבנית, metabolomic, לסנן (0.2 µm גודל הנקבוביות), דגה כל מאגרי באמצעות משאבת ואקום עבור 1 h.
- Resuspend מיובשים פפטיד צניפה המאוחסן ב- 2 mL צינור microcentrifuge 50 µL של המאגר C (3% v/v לחצן מצוקה, 0.1% חומצה פורמית וי/v) באמצעות פיפטה טיפול בנוזלים ידנית עם 200 קיבולת האחסון µL. Sonicate דגימות למשך 15 דקות באמבט אולטרא בתדירות אולטרה סאונד 35 קילו-הרץ.
התראה: לחצן מצוקה, חומצה פורמית הם חומרים מזיקים. מתחת למכסה המנוע fume וללבוש ציוד למיגון אישי המתאים, כגון: כפפות. - Centrifuge הדגימה ב 20,817 g x 10 דקות ב 4 ° C, ולאחר מכן להעביר 20 µL של תגובת שיקוע בקבוקון זכוכית.
- פפטידים מתעכל נפרד באמצעות עמודה הפוך-פאזי סי18 מחובר כרומטוגרפיה נוזלית ולרכוש ספקטרה המונית באמצעות ספקטרומטר מסה.
- נפרדים בעמודה µL 3 מדגם באמצעות קצב זרימה 300-nl/min. עבור שלב נייד, השתמש מאגר C ו- D (63% v/v לחצן מצוקה, 0.1% חומצה פורמית וי/v), ויוצרים הדרגתי ramping מ-3% לחצן מצוקה עד 15% לחצן מצוקה מעל 20 דקות ולאחר מכן לרשת 30% לחצן מצוקה מעל הבא 10 דקות.
הערה: אחסן את שאר המדגם מהנהר או –80 ° C עד מספר חודשים. לפני המדידה פרוטיאומיה מבנית, להקפיא מחדש את הדגימה על קרח. - לשטוף מזהמים 10 דקות באמצעות 60% לחצן מצוקה, equilibrate את העמודה עם µL 5 מאגר C לפני המדידה של הדוגמה הבאה.
- לקבל ספקטרום המונית באמצעות נתונים תלויי-שיטת MS/MS עם ערכת רזולוציה 70,000, AGC היעד של יונים6 3e, זמן הזרקת מרבי של 100 ms ו- m/z ועד 300 1600. רוכשים מרבי של סריקות 15 MS/MS רזולוציה של 17,500, AGC היעד של 1e5, זמן הזרקת מרבי של 100 אלפיות שניה, יחס underfill של 20%, עם חלון בידוד של 1.6 m/z ו- m/z הנע בין 200 ל- 2000. לאפשר איפקס טריגר (6 – 20 s), הדרה דינמי set כדי s 15, ו אל תכלול חיובים של 1 ו- > 5.
- נפרדים בעמודה µL 3 מדגם באמצעות קצב זרימה 300-nl/min. עבור שלב נייד, השתמש מאגר C ו- D (63% v/v לחצן מצוקה, 0.1% חומצה פורמית וי/v), ויוצרים הדרגתי ramping מ-3% לחצן מצוקה עד 15% לחצן מצוקה מעל 20 דקות ולאחר מכן לרשת 30% לחצן מצוקה מעל הבא 10 דקות.
7. עיבוד נתונים פרוטיאומיה מבנית
- להוריד את מסד הנתונים החדש פרוטאום תודרנית לבנה http://www.uniprot.org/ וכוללים מזהם מסד נתונים. לנתח את הנתונים הגולמיים המתקבל LC-MS פועל באמצעות MaxQuant עם משולב אנדרומדה פפטיד מנוע החיפוש משתמש ברירת המחדל עם התומכים LFQ נורמליזציה20,21,22. למצוא מידע מפורט אודות פרמטרים בשימוש בטבלה S1.
- פתח את הקובץ פלט "חלבון groups.txt". לצורך ניתוח נוסף, לסנן לקבוצות חלבון המזוהה עם פחות שני פפטידים ייחודי. הסר קבוצות חלבון המוגדר על-ידי MaxQuant פוטנציאליים מזהמים ומסנן לבנה א חלבונים (ARATH Fasta כותרות עמודה) קיים במסד הנתונים.
- כדי לבדוק את חשיבות העשרה חלבונית בין דגימות, להשתמש LFQ מנורמל עוצמות ולבצע אינטראקצית, דו-זנבית של סטודנט מבחן t ולאחריו השוואה מרובים תיקון (למשל בנימיני & הוכברג שווא גילוי שיעור (פד) תיקון או תיקון Bonferroni).
- חישוב ערך-p על-ידי השוואת עוצמות LFQ השיג עבור EV ובקרה NDPK1. לסנן את כל הערכים לא נקבע. למיין ערכי-p עולה ולהשתמש R script או מחשבון מקוון (למשל, https://www.sdmproject.com/utilities/?show=FDR) לחישוב תיקון רוזוולט. פילטר רוזוולט הערכים מתחת 0.1.
הערה: לשקול את הצורה של ניתוח נתונים מתאימים למחקר. מחקרים כמותיים (ניתוח של העשרה חלבונית בין מדגמים) להשתמש בערך "LFQ העוצמה", ואילו עבור מחקר איכותני (נוכחות או היעדרות של חלבון מסוים) לבחור את הערך "עוצמה". - מסנן עבור קבוצות חלבון כי הם יותר בשפע NDPK1 השוואת לשלוט EV. לקבוע לוקליזציה של הפרטנרים הפוטנציאלים חלבון באמצעות מסד הנתונים סובא23 ותקן החלבונים משותפת לשפות אחרות עם NDPK1.
- חישוב ערך-p על-ידי השוואת עוצמות LFQ השיג עבור EV ובקרה NDPK1. לסנן את כל הערכים לא נקבע. למיין ערכי-p עולה ולהשתמש R script או מחשבון מקוון (למשל, https://www.sdmproject.com/utilities/?show=FDR) לחישוב תיקון רוזוולט. פילטר רוזוולט הערכים מתחת 0.1.
8. מידת דגימות המכיל שלב הקוטב באמצעות UPLC – MS.
- Resuspend מיובשים שלב קוטב של שלב µL 4.5 ב 200 של מים, sonicate את הדגימה במשך 5 דקות.
- Centrifuge הדגימה ב 20,817 g x 10 דקות ב 4 ° C, ולאחר מכן להעביר את תגובת שיקוע בקבוקון זכוכית.
הערה: אחסן את שאר המדגם מהנהר או –80 ° C עבור אפילו מספר חודשים. לפני המדידה metabolomic, להקפיא מחדש את הדגימה על קרח. - לבצע צעד הפרדה באמצעות UPLC בשילוב לעמודה הפוך-פאזי סי18 ולרכוש ספקטרה המוני עם MS.
- להעמיס על µL טור 2 המדגם לכל זריקה עבור כל מצב יינון (חיוביים ושליליים) ולהפריד את השבר באמצעות 400 קצב הזרימה µL/min. כדי ליצור מעבר הצבע הדרוש למדידה מטבוליט, להכין פתרון שלב ניידים כדלקמן: מאגר של (0.1% חומצה פורמית H2O) ואת החיץ B (0.1% חומצה פורמית לחצן מצוקה).
- מטבוליטים נפרד µL 400/min, מעבר הצבע הבאים: 1 דקה 99% של מאגר A, 11-מין צבע ליניארי מ- 99% של מאגר A 60% של מאגר א', 13-מין צבע ליניארי מ- 60% של מאגר של 30% של מאגר A, 15-מין צבע ליניארי מ-30% של מאגר א-1% מאגר A, החזק בריכוז 1% עד למינימום 16 החל מ- 17 דקות, צבע ליניארי שימוש מ 1% של מאגר של 99% של מאגר א equilibrate מחדש את העמודה עבור 3 דקות עם 99% ריכוז במאגר A לפני המדידה של הדוגמה הבאה.
- לרכוש ספקטרה המוני המכסים טווח המונית בין 100 ל 1500 מ'/z עם רזולוציה מוגדר 25,000, טעינת זמן מוגבלת למטרה להגדיר AGC גב' 100 ל 1e6, מתח נימי כדי 3kV עם נדן גז זרימה והערך גז עזר של 60 ו- 20 , בהתאמה. להגדיר טמפרטורה נימי מתח 250 ° C ו מקפה כדי 25V.
9. עיבוד נתונים מטבולומיקס
- תהליך אסף chromatograms שנרכש בשני מצבי יינון. להשתמש בתוכנה כדי לחלץ מסה יחס מטען (מ'/z), זמן השמירה (RT), האינטנסיביות של פסגות משויך, למשל, תוכנה מסחרית (ראו טבלה של חומרים) או חלופיים24.
- הפעל תוכנת עיבוד על-ידי לחיצה כפולה על קובץ ה-.exe
- ליצור זרימת עבודה חדשה, לחפש פעילות "טעינה של קובץ" ולאחר להעביר פעילות זו על-ידי "גרור ושחרר" לחלל ריק זרימת עבודה. הקש את הפעילות עם הכפתור הימני של העכבר, לפתוח את הגדרות של הפעילות.
- בכרטיסייה ' המכיל הגדרות "כללי", להגדיר שם של הניסוי בשדה "שם", הבא לחץ על "בחר קבצים ותיקיות" ולסמן raw chromatograms.
- בכרטיסייה ' המכיל הגדרות "מתקדם", מוגדר "פרופיל נתונים הפסקת" עוצמת 0. לחץ על "החל" ועל "אישור".
- לחיפוש ולהוסיף פעילות "לטאטא נתונים". הקש את הפעילות עם הכפתור הימני של העכבר, לפתוח את הגדרות של הפעילות.
- בכרטיסייה ' המכיל הגדרות "כללי", לסמן "Centroid נתונים" ו- "נתונים MS/MS". להסיר את כל הנתונים MS/MS על-ידי בחירה "הכול" בחלונית ' בחירה '.
- לחיפוש ולהוסיף את פעילות "Chromatogram חיסור רעש כימי". הקש את הפעילות עם הכפתור הימני של העכבר, לפתוח את הגדרות של הפעילות.
- בכרטיסייה ' המכיל הגדרות "כללי", לסמן "Chromatogram החלקה", להגדיר מספר סריקות כדי "3" ו- "משערך" כדי "ממוצע נע". הגדר "RT חלון" סריקות 51, "Quantile" ל- 50%, חיסור "שיטה" ועוצמה 750 "סף".
- בכרטיסייה ' הכולל את "מתקדם" הגדרות, לסמן "RT מבנה להסרת" ולהגדיר "מינימום RT משך" 5 סריקות.
- בכרטיסייה ' המכיל הגדרות "מתקדם", לסמן "m/z מבנה להסרת" ולהגדיר "מינימום ז/ז אורך" 3 נקודות.
- לחיפוש ולהוסיף את פעילות "Chromatogram RT יישור". הקש את הפעילות עם הכפתור הימני של העכבר, לפתוח את הגדרות של הפעילות.
- בכרטיסייה ' המכיל הגדרות "כללי", להגדיר "יישור" מזימתה "של Pairwise יישור בסיס עץ" ו- "מרווח זמן חיפוש RT" מין 0.5.
- בכרטיסייה ' הכולל את "מתקדם" הגדרות, להשתמש בפרמטרים של ברירת המחדל.
- לחיפוש ולהוסיף את פעילות "שיא זיהוי" בקבוצת "Chromatogram" של פעילויות. הקש את הפעילות עם הכפתור הימני של העכבר, לפתוח את הגדרות של הפעילות.
- בכרטיסייה ' המכיל הגדרות "כללי", מוגדר "סיכום חלון" 0.09 דקות, מינימום שיא "גודל" מינימום 0.03 נקודות, "מיזוג המרחק המקסימלי" 5 נקודות ואסטרטגיה "למזג" "מרכזי". ב- "שיא RT פיצול" תיבת להגדיר "יחס פער/שיא" ל- 50%.
- בכרטיסייה ' המכיל הגדרות "מתקדם", מוגדר "החלקת חלון" 5 נקודות, "עידון הסף" 80% ו- "סף עקביות" 1. הגדר "מרכז חישוב" כמו "משוקלל בעוצמה" עם "עוצמת סף" להגדיר ב-70%.
- לחיפוש ולהוסיף את פעילות "קיבוץ באשכולות איזוטופ" בקבוצת "Chromatogram" של פעילויות. הקש את הפעילות עם הכפתור הימני של העכבר, לפתוח את הגדרות של הפעילות.
- בכרטיסייה ' המכיל הגדרות "כללי", להגדיר "סובלנות RT" כדי מינימום 0.015 z- ו m/סובלנות 5 עמודים לדקה.
- בכרטיסיה לקבוע הגדרות המכילות "המעטפה הולם", "שיטה" כמו "לא" הגבלת צורה"ו-"יינון""פרוטונציה (עבור מצב חיובי), "דה-פרוטונציה" (עבור מצב שלילי). הגדר מינימום ומקסימום החיוב"1 ו-4, בהתאמה.
- בכרטיסייה ' הכולל את "מתקדם" הגדרות, להשתמש בפרמטרים של ברירת המחדל.
- לחיפוש ולהוסיף פעילות "סינגלטון מסנן".
- כדי לייצא את התוצאות של עיבוד נתונים, לחפש ולהוסיף את פעילות "אנליסט" בקבוצת "יצוא" של פעילויות.
- בכרטיסייה ' המכיל הגדרות "כללי", להגדיר "סוג" "אשכולות" ו- "הנצפה" כמו "לסכם בעוצמה". לבחור "ליעד מותאמת אישית", ציין ספריית של קובץ ייצוא.
- בכרטיסייה ' הכולל את "מתקדם" הגדרות, להשתמש בפרמטרים של ברירת המחדל.
- הוספת ביאורים ההמונית תכונות משתמש במסד הפניה-מתחם ללא צורך במיקור חוץ.
- לנתח כיתה MS אחד או מספר הפניה-מתחם באמצעות שימוש UPLC-גב' LC-MS באותה השיטה לניתוח של התייחסות-תרכובות מטבוליטים מטוהרים במשותף עם חלבון עניין.
הערה: במחקר זה, ערכה של כמעט 300 dipeptides היה מנותח, המשמש כספריית הפניה-מתחם. - השתמש ניתוח תוכנה (ראה טבלה של חומרים) כדי לפתוח קובץ raw chromatogram ולחפש עבור m ספציפי/z ו- RT המשויך למדידה הפניה-מתחם (עיין במדריך למשתמש).
שימו לב: מטבוליטים משניים שונים בסוגי יינון. לבדוק הנוכחות של המשותף adducts על-ידי חיפוש עבור יון מסה שווה ל M-1.007276, M + 1.007276, M + 18.033823 מ' + 22.989218 [M-H], [M + H], [M + NH4] ו- [M + Na], בהתאמה. - גיליון אלקטרוני להשתמש כדי לפתוח קובץ "אנליסט" שהושג לאחר העיבוד של chromatograms ושל חיפוש של יונים וברצפטורים ספציפיים מיוצא המוני. השווה RT של התכונה המוני נמדד את הניסוי ואת RT של המתחם הפניה. לאפשר סטייה של 0.005 Da עבור מ/z ו- 0.1 min עבור RT.
- לנתח כיתה MS אחד או מספר הפניה-מתחם באמצעות שימוש UPLC-גב' LC-MS באותה השיטה לניתוח של התייחסות-תרכובות מטבוליטים מטוהרים במשותף עם חלבון עניין.
- כדי לבדוק את המשמעות של המטבוליט העשרה מטוהרים במשותף עם חלבון מסוים בין דגימות (קו עם חלבונים overexpressed השליטה לעומת EV ריבית), להשוות בין ערכי שיא באמצעות דו-זנבית בלתי-זוגי של התלמיד במבחן t ולאחריו מרובות תיקון השוואה (למשל בנימיני & הוכברג גילוי שקר קצב תיקון או תיקון Bonferroni).
תוצאות
במחקר המקורי, בשלושה גנים לבנה א NDPK היו overexpressed בתרבויות PSB-L תא ההשעיה תחת השליטה של יזם 35S מכוננת14 (איור 1). תג קירבה טנדם שהיה מחובר מקצוות carboxy - או אמינו-מסוף חלבון פיתיון לגבו. מתחמי מטוהרים זיקה היו נענשים MTBE/מתנול/מים החילוץ16. משך-זיקה חלבונים ומולקולות קטנות אותרו באמצעות MS (טבלאות S2, S3).
לתיקון תוצאות חיוביות שגויות, דגימות ריק שימשו כדי לא לכלול מזהמים מולקולה קטנה מן כימיקלים מעבדה מתכלים. יתר על כן, מטבוליטים וחלבונים לאגד גם תג קירבה או שרף לבד היה אליבי באמצעות קווי הבקרה EV. כדי לאחזר תוצאות חיוביות נכון, דו-זנבית בלתי-זוגי של התלמיד במבחן t וקצב בנימיני & הוכברג גילוי שקר הוחל תיקון כדי לזהות מטבוליטים (טבלה S4), חלבונים (טבלה S5) באופן משמעותי מועשר בנקודת הגישה NDPKs ניסויים (N - ו C-סופני מתויג NDPKs) לעומת הקווים שליטה EV (פד ר < 0.1). שימו לב כי עבודה קודמות, נהגנו היעדרות/נוכחות קריטריונים ניסחו interactors חלבון ומולקולה קטנה.
נציג תוצאות ניתנות עבור NDPK1, בעוד מטבוליט נתונים להתמקד dipeptides, מחלקה הרומן של הרגולטורים מולקולה קטנה למד בקבוצה שלנו. ניתוח פרוטיאומיה מבנית גילה 26 שותפים חלבון בשם של NDPK1. על-ידי סינון נוסף חלבונים משותפת לשפות אחרות בתא subcellular אותו בתור NDPK1 (ציטוזול), הרשימה צמצמנו 13 חלבון בשם interactors. בין החלבונים שזוהו היו גלוטתיון S-טרנספראז, התארכות שני גורמים חניכה, טובולין, aconitate hydratase. ניתוח Metabolomic גילה ארבעה dipeptides ואל-Leu באיל-Glu, Leu-איל, איל-Phe זה במיוחד משותפת eluted עם NDPK1 (איור 2). שים לב כי כל dipeptides ארבע לשתף משקע הידרופובי ב שלהם אמיני, רומז משותפת מחייב ירידה לפרטים.
לחפש מתחמי חלבון, חלבון-מטבוליט ידוע לנו 13 המאוחזרים לזהות חלבונים ו dipeptides ארבעה נגד תפר מסד25 (איור 3). מספר תצפיות עשוי להיות: (i) אף אחד interactors דווח קודם לכן עבור NDPK1. (ii) APX1 ortholog דווח לקיים אינטראקציה עם אלדהיד דהידרוגנאז משפחה ALDH7B4, תוך תרגום חניכה פקטור FBR12 עם תרגום חניכה גורם נוסף מקודדת על ידי הגן AT2G40290. (iii) dipeptides מזוהה לא דיווחו על חלבון שותפים. Dipeptides eluted משותפת לא דווחו קודם הקשורות אל כל חלבון צמחי שאוחזרו. עם זאת, הם תפקיד חשוב אורגניזמים אחרים: Leu-איל, למשל, יש אפקט neurotrophin-מפעיל קו26תאים אנושיים. שימו לב כי הניסוי אינו מאפשר לזהות את הטופולוגיה המדויק של המערכת. לדוגמה, dipeptide ייתכן אינטראקציה ישירה עם NDPK1 אך עשויות להיות קשורות היטב לכל אחד החלבונים משותפת מטוהרים.
יחדיו, התוצאות שלנו להראות כי ההליך הוקמה, העסקת AP יחד עם ספקטרומטר מסה, מאפשר זיהוי של חלבון, חלבון-מולקולה קטנה interactors, מסייע ליצור מידע נרחב אודות interactome של החלבון היעד.

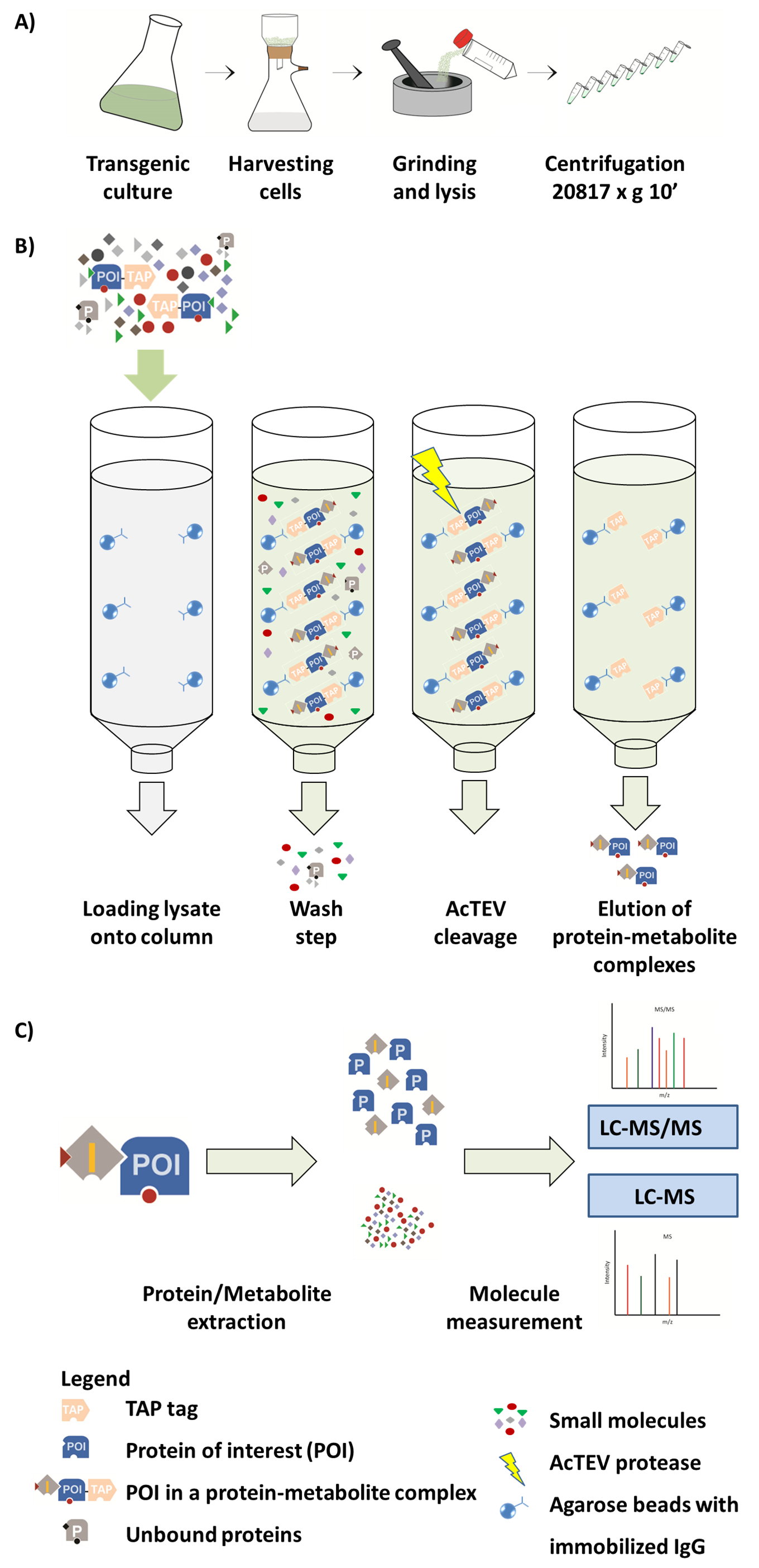
איור 1. ערכה של זרימת עבודה AP-MS. (א) הכנה של השבר מסיסים מקורית של תרבית תאים. השלבים הבאים (ב') במסגרת ההליך AP. לאחר טעינת המדגם אל העמודה, נקשר החלבון עניין (פוי) התמזגו לתגית ברז הנוגדן IgG ותשמרו על החרוזים agarose. כביסה של העמודה מקלה על הסרת חלבונים לא מאוגד מטבוליטים. לאחר ביצוע AcTEV המחשוף, מתחמי חלבון-מטבוליט פוי הם eluted. (ג) ההפרדה של מתחמי לתוך שבר חלבון ו מטבוליט שלאחריה יבוא ניתוח MS הכמותיים למחצה. חלק איור זה מתרבה מ. ואח 2017 Luzarowski14. אנא לחץ כאן כדי להציג גירסה גדולה יותר של הדמות הזאת.
{kind=link}

באיור 2. במיוחד eluting יחד עם NDPK1 dipeptides. עוצמות ממוצע של ארבעה dipeptides ואל-Leu (א), איל-Glu (B), Leu-איל (ג)ואיל-Phe (D) שנמדד בניסוי AP שורטטו. כל dipeptides ארבע להראות העשרה משמעותי בדגימות NDPK1 לעומת שליטה EV (כוכביות מייצגים רוזוולט < 0.1). קווי שגיאה מייצגים שגיאת התקן עבור מדידות 6 (3 ושכפול של N - ו 3 של C-סופני מתויג חלבונים). אנא לחץ כאן כדי להציג גירסה גדולה יותר של הדמות הזאת.
{kind=link}

איור 3. אינטראקציה עם רשת של כל המולקולות eluting בשיתוף עם NDPK1, שאילתה מול מסד הנתונים תפר שוקל רק הקודם ניסיוני, עדויות של מסד הנתונים (ביטחון > 0.2). בטחון גבוהה יותר מצביעה על סיכון גבוה יותר של אינטראקציה, מחושב בהתבסס על הנתונים הופקדו. אנא לחץ כאן כדי להציג גירסה גדולה יותר של הדמות הזאת.
{kind=link}
שולחן S1. MaxQuant פלט טבלת "parameters.txt". העניינים כולל ערכי סף עבור זיהוי, כימות, כמו גם מידע אודות מסדי הנתונים המשמש. אנא לחץ כאן כדי להוריד את הקובץ.
שולחן S2. מידע MaxQuant פלט טבלת "proteinGroups.txt". הטבלה מכילה רשימה של כל קבוצות שזוהה חלבון, עוצמות, פרטים נוספים כגון מספר ייחודי פפטידים והניקוד. אנא לחץ כאן כדי להוריד את הקובץ.
שולחן S3. קובץ פלט ובו ניתוח של מטבוליטים קוטבי. הטבלה מכילה רשימה של כל התכונות המוני מזוהה המאופיין מ ספציפי/z, RT ועוצמה. אנא לחץ כאן כדי להוריד את הקובץ.
שולחן S4. Dipeptides נמצאו בדגימות AP אשר NDPK1, NDPK2 או NDPK3 שימשו בתור פיתיון. Dipeptides נוכח בדגימות ריק לא נכללו ברשימה. שני עצמאית קווים (מתויג ב או או C-אמיני) עבור כל NDPK היו להפעיל בשלושה עותקים. הסטודנט t-מבחן ולמנף תיקון של p-ערך באמצעות בנימיני & שיטה הוכברג שימשו כדי לקבוע באופן משמעותי מועשר שותפים אינטראקטור של NDPKs (פד ר < 0.1). בהתחשב ΔRT מחושבת ביחס הפניה תרכובות של Δppm ביחס monoisotopic המוני נתון Metlin27. אנא לחץ כאן כדי להוריד את הקובץ.
שולחן S5. חלבונים מטוהרים בשיתוף עם NDPK1. שני עצמאית קווים (מתויג ב או או C-אמיני) עבור כל NDPK נוהלו דולר. הסטודנט t-מבחן ולמנף תיקון של p-ערך באמצעות בנימיני & שיטה הוכברג שימשו כדי לקבוע באופן משמעותי מועשר שותפים אינטראקטור של NDPKs (פד ר < 0.1). אנא לחץ כאן כדי להוריד את הקובץ.
Discussion
פרוטוקול הציג מאפשר זיהוי מקבילי של מתחמי PP ו- PM של חלבון המטרה. מ שיבוט על התוצאות הסופיות, ניתן להשלים את הניסוי קצת כמו 8-12 שבועות. AP מלאה אורכת כ 4-6 h עבור קבוצה של 12 עד 24 דגימות, עיבוד פרוטוקול שלנו מתאימים לניתוח תפוקה באמצע.
הפרוטוקול, למרות להיות בסך הכל פשוטה, יש מספר שלבים קריטיים. (i) כמות מספקת של חלבון קלט וחרוזים זיקה חיוני להגיע טווח דינמי של המטבוליט זיהוי. פירוק התא יעיל לכן צעד מכריע בהליך. התשואות המסכן חלבון יכול להיות תוצאה של pulverization לא מספקת של החומר או של שיוצרת יחס פירוק-מאגר/חומר. (ii) להקפיד כי ריאגנטים בשימוש הם ידידותיים MS. חומרי ניקוי חזקים, גליצרול או כמויות מוגזמות של מלח יש להימנע ככל מתערבות MS זיהוי. (iii) חרוזים Agarose לא צריך להיות יבשים יתר במהלך שטיפת שלבים, כאשר משתמש של יריעה ואקום זה חשוב להחיל קצב זרימה איטית כדי לא כדי להרוס את החרוזים או להשפיע על יציבות מורכבים.
ישנם כמה שינויים אפשריים חשובים בפרוטוקול הציג: (i) נשתמש האמרגן CaMV35S המכונן כדי למקסם את כמות החלבון פיתיון. ביטוי, בעוד מאוד שימושי, ניתן להיות השפעה רצינית על הומאוסטזיס תא28 , להוביל להיווצרות אינטראקציות לא רלוונטי מבחינה פיזיולוגית. ביטוי של חלבונים מתויגים באמצעות היזמים מקורי שבו האפשר רקע אובדן-של-פונקציה נחשב נעלה לאחזור interactors הביולוגית האמיתית. חלבונים בדרך כלל לא באה לידי ביטוי בתרבויות תא צמח, צמח רקע יוכיח הדרושות כדי לזהות interactors רלוונטיים. (ii) בעת עבודה עם קרום חלבונים, המאגר פירוק צריך להיות בתוספת של דטרגנט התואמים ל- MS. (iii) הקדמה של שלב טיהור זיקה שני יכול לשפר שווא-תוצאות חיוביות ליחס חיובי true, לבטל את הצורך פקדים EV29. תג טנדם הרומן עם שני אתרים פרוטאז עצמאית-המחשוף מציג חלופה אטרקטיבית השלב גודל-הדרה כרומטוגרפיה שנוספו על-ידי. et al. 2014 מיאדה11, זה מפרך וגם זמן רב.
החיסרון החמורה ביותר של נקודת הגישה הוא שיעור גבוה של תוצאות חיוביות שגויות. הסיבות הן רבות. ביטוי מכוננת כבר הוזכר. מקור נוסף של אינטראקציות לא רלוונטי מבחינה פיזיולוגית, אלא אם כן עובד עם organelles מבודד, הוא הכנת כל-תא lysates המכיל תערובת של חלבונים ושל מטבוליטים של תאים subcellular שונים. לוקליזציה subcellular אמור לשמש כדי לבצע סינון עבור interactors אמיתי. ובכל זאת, הרוב המכריע של תוצאות חיוביות שגויות כתוצאה איגוד לא ספציפי בין חלבונים שרפים agarose. הקדמה של שלב טיהור שני, כפי שתוארה לעיל, מציעה את הפתרון הטוב ביותר לבעיה, עם זאת מגיע על חשבון זמן ותפוקת. יתר על כן, אינטראקציה חלשה עלול ללכת לאיבוד כמו הפרוטוקול מתארך. אזהרה נוספת של AP הוא למרות מידע מקיף שמספק לגבי interactome של חלבון המטרה, המבדילים בין היעדים הישירים והעקיפים של החלבון פתיונים הוא בלתי אפשרי. גישות ריאקציה דו-מולקולרית יישוב נדרשים לאשר אינטראקציות.
AP בשילוב עם גליקומיקס מבוססי MS שימש ללמוד חלבון-מתחמי cerevisiae ס12. עבודה זו, יחד עם שלנו תצפית קודמות13 , באופן דומה כדי שומנים, תרכובת פולאר polar למחצה להישאר מאוגד ל קומפלקסי חלבון מבודד תאית lysates, סיפק והיערכות מושגית עבור פרוטוקול שהוצגו. פרוטוקול שלנו מאופיין על ידי שלוש נקודות ייחודיות: (i) לעומת זאת כדי השמרים לעבוד12, הוא מדגים כי AP מתאים מאחזר לא רק חלבון הידרופובי אבל גם הידרופילית ליגנדים. (ii) בהצגת נוהל חילוץ 3-in-one, AP יחיד ניתן ללמוד interactors חלבון ו מטבוליט של החלבון פיתיון. (iii). שינינו את פרוטוקול לשתול תאים.
המאמצים העתידיים יתמקד ביצירת תג טנדם הרומן עם שני אתרים פרוטאז עצמאית-המחשוף. אנחנו רוצים גם לחקור את התאמתו של הפרוטוקול כדי נמוך-שפע מולקולות קטנות כמו הורמונים צמחיים.
Disclosures
המחברים אין לחשוף.
Acknowledgements
ברצוננו להודות בחביבות פרופ ' ד ר לותאר Willmitzer בשל מעורבותו בפרוייקט, בדיונים פרודוקטיביים ולאחר השגחה מעולה. אנחנו אסירי תודה ד ר דניאל Veyel שעזרת עם מדידות MS פרוטיאומיה מבנית. אנחנו מעריכים מיכאליס Änne גברת מי סיפק לנו עזרה טכנית שלא תסולא בפז מדידות LC-MS. יתר על כן, ברצוננו להודות ד ר מוניקה Kosmacz, ד ר Ewelina Sokołowska שלהם לעזרה ומעורבות בעבודה על כתב היד המקורי, ולא כדי Weronika Jasińska לקבלת תמיכה טכנית.
Materials
| Name | Company | Catalog Number | Comments |
| Murashige and Skoog Basal Salts with minimal organics | Sigma-Aldrich | M6899 | |
| 1-Naphthylacetic acid | Sigma-Aldrich | N1641 | |
| Kinetin solution | Sigma-Aldrich | K3253 | |
| Tris base | Sigma-Aldrich | 10708976001 | |
| NaCl | Sigma-Aldrich | S7653 | |
| MgCl2 | Carl Roth | 2189.1 | |
| EDTA | Sigma-Aldrich | 3609 | |
| NaF | Sigma-Aldrich | S6776 | |
| DTT | Sigma-Aldrich | D0632 | |
| PMSF | Sigma-Aldrich | P7626 | |
| E-64 protease inhibitor | Sigma-Aldrich | E3132 | |
| Protease Inhibitor Cocktail | Sigma-Aldrich | P9599 | |
| Na3VO4 | Sigma-Aldrich | S6508 | |
| AcTEV Protease | Thermo Fischer Scientific | 12575015 | |
| Rotiphorese Gel 30 (37,5:1) | Carl Roth | 3029.2 | |
| TEMED | Carl Roth | 2367.3 | |
| PageRuler Prestained Protein Ladder | Thermo Fischer Scientific | 26616 | |
| SBP Tag Antibody (SB19-C4) | Santa Cruz Biotechnology | sc-101595 | |
| Goat anti-mouse IgG-HRP | Santa Cruz Biotechnology | sc-2005 | |
| Bradford Reagent | Sigma-Aldrich | B6916 | |
| Trypsin/Lys-C Mix, Mass Spec Grade | Promega | V5071 | |
| Urea | Sigma-Aldrich | U5128 | |
| Thiourea | Sigma-Aldrich | T8656 | |
| Ammonium bicarbonate | Sigma-Aldrich | 9830 | |
| Iodoacetamide | Sigma-Aldrich | I1149 | |
| MTBE | Biosolve | 138906 | |
| Methanol | Biosolve | 136806 | |
| Water | Biosolve | 232106 | |
| Acetonitrile | Biosolve | 12006 | |
| Trifluoroacetic acid | Biosolve | 202341 | |
| Formic acid | Biosolve | 69141 | |
| Unimax 2010 Platform Shaker | Heidolph | 5421002000 | |
| Nylon Mesh (Wire diameter 34 µM, thickness 55 µM, open area 14%) | Prosepa | Custom order | |
| Glass Funnel, 47 mm, 300 ml | Restek | KT953751-0000 | |
| Filter Bottle Top 500 mL 0,2 µM Pes St | VWR International GmbH | 514-0340 | |
| Mixer Mill MM 400 | Retsch GmbH | 207450001 | |
| IgG Sepharose 6 Fast Flow | GE Healthcare Life Sciences | 17-0969-02 | |
| Mobicol ""Classic"" with 2 different screw caps without filters | MoBiTec GmbH | M1002 | |
| Filter (small) 35 µM pore size, for Mobicol M 1002, M1003, M1050 & M1053 | MoBiTec GmbH | M513515 | |
| Variable Speed Tube Rotator SB 3 | Carl Roth | Y550.1 | |
| Rotary dishes for rotators SB 3 | Carl Roth | Y555.1 | |
| Resprep 24-Port SPE Manifolds | Restek | 26080 | |
| Finisterre C18/17% SPE Columns 100mg / 1ml | Teknokroma | TR-F034000 | |
| Autosampler Vials | Klaus Trott Chromatographie-Zubehör | 40 11 01 740 | |
| Acclaim PepMap 100 C18 LC Column | Thermo Fischer Scientific | 164534 | |
| EASY-nLC 1000 Liquid Chromatograph | Thermo Fischer Scientific | LC120 | |
| Q Exactive Plus Hybrid Quadrupole-Orbitrap Mass Spectrometer | Thermo Fischer Scientific | IQLAAEGAAPFALGMBDK | |
| Acquity UPLC system | Waters | Custom order | |
| ACQUITY UPLC HSS C18 Column, 100A, 1.8 µM, 2.1 mM X 100 mM, 1/pkg | Waters | 186003533 | |
| High-power ultrasonic cleaning baths for aqueous cleaning solutions | Bandelin | RK 31 | |
| Genedata Expressionist | Genedata | NaN | |
| Xcalibur Software | Thermo Fischer Scientific | NaN | |
| MaxQuant | NaN | NaN |
References
- Li, X., Snyder, M. Metabolites as global regulators: A new view of protein regulation. Bioessays. 33 (7), 485-489 (2011).
- Jacob, F., Monod, J. Genetic regulatory mechanisms in the synthesis of proteins. Journal of Molecular Biology. 3 (3), 318-356 (1961).
- Schlattner, U., et al. Dual Function of Mitochondrial Nm23-H4 Protein in Phosphotransfer and Intermembrane Transfer a cardiolipin-dependent switch. Journal of Biological Chemistry. 288 (1), 111-121 (2013).
- Ramírez, M. B., et al. GTP binding regulates cellular localization of Parkinson's disease-associated LRRK2. Human Molecular Genetics. , ddx161 (2017).
- Jung, H. J., Kwon, H. J. Target deconvolution of bioactive small molecules: the heart of chemical biology and drug discovery. Archives of Pharmacal Research. 38 (9), 1627-1641 (2015).
- Harding, M. W., Galat, A., Uehling, D. E., Schreiber, S. L. A receptor for the immunosuppressant FK506 is a cis-trans peptidyl-prolyl isomerase. Nature. 341 (6244), 758-760 (1989).
- Lomenick, B., et al. Target identification using drug affinity responsive target stability (DARTS). Proceedings of the National Academy of Sciences of the United States of America. 106 (51), 21984-21989 (2009).
- Manabe, Y., Mukai, M., Ito, S., Kato, N., Ueda, M. FLAG tagging by CuAAC and nanogram-scale purification of the target protein for a bioactive metabolite involved in circadian rhythmic leaf movement in Leguminosae. Chemical Communications. 46 (3), 469-471 (2010).
- Pantoliano, M. W., et al. High-density miniaturized thermal shift assays as a general strategy for drug discovery. Journal of Biomolecular Screening. 6 (6), 429-440 (2001).
- Li, X., Snyder, M. Analyzing In vivo Metabolite-Protein Interactions by Large-Scale Systematic Analyses. Current Protocols in Chemical Biology. , 181-196 (2010).
- Maeda, K., Poletto, M., Chiapparino, A., Gavin, A. -. C. A generic protocol for the purification and characterization of water-soluble complexes of affinity-tagged proteins and lipids. Nature Protocols. 9 (9), 2256-2266 (2014).
- Li, X., Gianoulis, T. A., Yip, K. Y., Gerstein, M., Snyder, M. Extensive in vivo metabolite-protein interactions revealed by large-scale systematic analyses. Cell. 143 (4), 639-650 (2010).
- Veyel, D., et al. System-wide detection of protein-small molecule complexes suggests extensive metabolite regulation in plants. Scientific Reports. 7, (2017).
- Luzarowski, M., et al. Affinity purification with metabolomic and proteomic analysis unravels diverse roles of nucleoside diphosphate kinases. Journal of Experimental Botany. , (2017).
- Van Leene, J., et al. Targeted interactomics reveals a complex core cell cycle machinery in Arabidopsis thaliana. Molecular systems biology. 6 (1), 397 (2010).
- Giavalisco, P., et al. Elemental formula annotation of polar and lipophilic metabolites using 13C, 15N and 34S isotope labelling, in combination with high-resolution mass spectrometry. The Plant Journal. 68 (2), 364-376 (2011).
- Van Leene, J., et al. Isolation of transcription factor complexes from Arabidopsis cell suspension cultures by tandem affinity purification. Plant Transcription Factors: Methods and Protocols. , 195-218 (2011).
- Van Leene, J., et al. A tandem affinity purification-based technology platform to study the cell cycle interactome in Arabidopsis thaliana. Molecular & Cellular Proteomics. 6 (7), 1226-1238 (2007).
- Olsen, J. V., Ong, S. -. E., Mann, M. Trypsin cleaves exclusively C-terminal to arginine and lysine residues. Molecular & Cellular Proteomics. 3 (6), 608-614 (2004).
- Cox, J., Mann, M. MaxQuant enables high peptide identification rates, individualized p.p.b.-range mass accuracies and proteome-wide protein quantification. Nature Biotechnology. 26 (12), 1367-1372 (2008).
- Cox, J., et al. Andromeda: A peptide search engine integrated into the MaxQuant environment. Journal of Proteome Research. 10 (4), 1794-1805 (2011).
- Tyanova, S., Temu, T., Cox, J. The MaxQuant computational platform for mass spectrometry-based shotgun proteomics. Nature Protocols. 11 (12), 2301 (2016).
- Hooper, C. M., et al. SUBAcon: a consensus algorithm for unifying the subcellular localization data of the Arabidopsis proteome. Bioinformatics. 30 (23), 3356-3364 (2014).
- Katajamaa, M., Orešič, M. Data processing for mass spectrometry-based metabolomics. Journal of Chromatography A. 1158 (1-2), 318-328 (2007).
- Szklarczyk, D., et al. STITCH 5: augmenting protein-chemical interaction networks with tissue and affinity data. Nucleic Acids Research. 1277, (2015).
- Tanaka, K. -. i., et al. Dipeptidyl compounds ameliorate the serum-deprivation-induced reduction in cell viability via the neurotrophin-activating effect in SH-SY5Y cells. Neurological Research. 34 (6), 619-622 (2012).
- Smith, C. A., et al. METLIN: A metabolite mass spectral database. Therapeutic Drug Monitoring. 27, 747-751 (2005).
- Bhattacharyya, S., et al. Transient protein-protein interactions perturb E. coli metabolome and cause gene dosage toxicity. Elife. 5, (2016).
- Rigaut, G., et al. A generic protein purification method for protein complex characterization and proteome exploration. Nature Biotechnology. 17 (10), 1030-1032 (1999).
Reprints and Permissions
Request permission to reuse the text or figures of this JoVE article
Request PermissionExplore More Articles
This article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. All rights reserved