
Method Article
ניתוח החלק ההקפאה של האזור התת-כספי למבוגרים לניתוח פרוטאום כמותי מדויק ועמוק
In This Article
Summary
קריו-סעיף-ניתוח מאפשר הכנה טרייה, קפואה של הנישה הנוירוגנית הגדולה ביותר במוח מורין לניתוח פרוטאום כמותי עמוק. השיטה מדויקת, יעילה וגורמת להפרעה מינימלית לרקמות. לכן, הוא מתאים באופן אידיאלי לחקר המיקרו-סביבה המולקולרית של נישה זו, כמו גם איברים, אזורים ומינים אחרים.
Abstract
הנישה הנוירוגנית התת-פננדימלית מורכבת מרצועת מפרקים של קיר החדר לרוחב של החדר הצדדי. האזור התת-קרקעי (SEZ) הוא אזור דק ומובחן החשוף לחדרים ולנוזל השדרתי. הבידוד של נישה זו מאפשר ניתוח של מיקרו-סביבה נוירוגנית של תאי גזע. עם זאת, מיצוי של רקמות קטנות לניתוח פרוטאום הוא מאתגר, במיוחד לשמירה על עומק מדידה ניכר והשגת חוסן אמין. שיטה חדשה המכונה cryo-סעיף-ניתוח (CSD), המשלב דיוק גבוה עם הפרעת רקמות מינימלית, פותחה כדי להתמודד עם אתגרים אלה. השיטה תואמת לשיטות ספקטרומטריית מסה (MS) חדישות המאפשרות זיהוי של רגולטורים נישה בשפע נמוך. מחקר זה השווה את ה- CSD ואת נתוני הפרוטאום שלו לשיטה ולנתונים המתקבלים על ידי מיקרודיסקציה של לכידת לייזר (LCM) וחתך סטנדרטי שלם. שיטת ה- CSD הביאה לעומק כימות כפול בפחות ממחצית זמן ההכנה בהשוואה ל- LCM ובו זמנית עלתה בבירור על דיוק הניתוח של הניתוח כולו. לפיכך, CSD היא שיטה מעולה לאיסוף SEZ לניתוח פרוטאום.
Introduction
כמו neurogenesis מוגבל במוח הבוגר, אסטרטגיות שונות לתיקון מערכת העצבים המרכזית ייהנו מאוד הבנה מוגברת של היסודות של תחליף עצבי למבוגרים. מכרסמים עזרו לנו להבין את המנגנונים הבסיסיים של נוירוגנזה לאחר הלידה, אם כי יש לציין כי נוירוגנזה בוגרת תלויה מאוד במינים. בעכברים, יש שלוש נישות של תאי גזע עצביים בוגרים (NSC). ההיפותלמוס הוא נישה בוגרת של המועצה לביטחון לאומי עם פוטנציאל נוירוגני 1,2, בעוד נוירוגנזה בוגרת מתמשכת מוגבלת בעיקר להיפוקמפוס3 ול- SEZ של הקירות הצדדיים של החדרים לרוחב4,5,6. ה- SEZ הוא אזור הנבטים הגדול ביותר המכיל NSCs (תאי סוג B) המתפתחים לנוירובלסטים (תאים מסוג A) באמצעות תאי אב מגבירי מעבר (תאי C מסוג C). ה-SEZ מכיל 20-35% מתאי סוג B, 1-15% מתאי סוג C, 1-30% מתאי סוג A ו-25-50% מתאי אפנדימאל7. ה- SEZ כולל מיקרו-ארכיטקטורה מורכבת, עם תאי אנדותל, תאים מיקרוגליאליים ותאים אפנדימליים המתגוררים ומשפיעים על נישה של תאי הגזע8,9,10. למרות נוירונים נדירים SEZ, אקסונים הנובעים ממקורות רחוקים כגון סטריאטום, אזור הטגמנטום הגחוני, או ההיפותלמוס להגיע ולהשפיע על סוג B תאים4. מאפיין ייחודי של נישה זו של תאי גזע הוא ההפרדה בין אתר ההתפשטות לבין אתר הבידול. לאחר התפשטות, האבות העצביים נודדים כמה מילימטרים מה- SEZ לנורת הריח, שם הם מתבדלים באופן סופני לנוירונים ומשתלבים במעגלים עצביים קיימים. חקירות של תוכניות תאי-מהותיות הקשורות נוירוגנזה כבר סיפקו ידע חשוב עבור תכנות מחדש של תאים טיפוליים ניסיוניים ואסטרטגיות השתלה15,16,17,17,18,19,20. עם זאת, אותות חיצוניים של תאים גם לווסת neurogenesis, סביבות רקמות יכול לקבוע את הגורל הנוירוגני של תאי גזע11,12,14,21,22,23. כתוצאה מכך, חקירת המיקרו-סביבה של הגומחות הנוירוגניות והאינטראקציה שלה עם תאי הגזע היא בעלת חשיבות מכרעת.
המטריצה החוץ-תאית (ECM) וחלבונים מופרשים אחרים הם חלק גדול מהמיקרו-סביבה. לזיהוי וכימות מדויקים, גישה פרוטאומית מתאימה יותר מגישה תמלולית לקביעת הרכב ECM בשל המתאם הנמוך בין התמלול לרמות החלבון עבור ECM24,25. יתר על כן, יש ראיות משמעותיות כי רגולטורים נישה SEZ אינם מיוצרים באופן בלעדי על ידי תאים המאכלסים את הנישה עצמה. במקומות מרוחקים יותר, כגון מקלעת הכורואידים, מפרישים אותות אפנון המועברים לתאי הגזע באמצעות הנוזל השדרתי22,23. חקירת פרוטאום נישה יכול לעזור לזהות רגולטורים נישה נוכח נישה עצמאית של אתר הייצור שלהם, בהתחשב בכך חלק ניכר של microenvironment חוץ תאי מורכב על ידי חלבונים.
כדי לאסוף את אזור חדרי המורין לניתוח פרוטאומי לא משוחד, נדרשת שיטה עם דיוק גבוה, לכידת סרט paraventricular דק כ -50 מיקרומטר המכיל תאי גזע תוך אי הכללת הרקמה של הסטריאטום הסמוך. יתר על כן, הפרעה רקמות במהלך הניתוח חייב להיות מינימלי לניתוח microenvironment חוץ תאי כי חלבונים מסיסים, כולל גורמי גדילה או ציטוקינים, ניתן לשטוף בקלות. למרות שניתן לנתח את ספקטרום המסה של רקמה קבועה, הסוכן הנדרש, כגון paraformaldehyde, יפחית את עומק זיהוי החלבון ועשוי לגרום לשינויים פוסט-טרנסלנטיים. ניתוח SEZ שלם נפוץ, למשל, עבור אוסף של תאים לניתוח מיון תאים המופעל על ידי פלואורסצנטיות, מסיר את כל ה- SEZ עם מספריים26. ביתור סטנדרטי זה הוא מהיר עם הפרעה מינימלית לרקמות. עם זאת, זיהום striatal של הדגימות לא ניתן להימנע. לעומת זאת, ל- LCM יש את היתרון יוצא הדופן של דיוק ניתוח מעולה. עם זאת, LCM עשוי להציג הפרעות רקמות, למשל, עקב כתמי רקע או denaturation חלבון שנגרם בלייזר. כדי לשלב את נקודות החוזק של הניתוח כולו ו- LCM, פותחה שיטה חדשנית התואמת לטרשת נפוצה, המכונה קריו-קטע-חתך (CSD), (איור 1A-D). ה- CSD מאפשר את החילוץ של SEZ ואת הניתוח של SEZ של הקירות המדיאליים של החדרים לרוחב (MEZ), שהוא אזור בקרה אידיאלי, בעיקר לא נוירוגני עבור SEZ (ראה הפרוטוקול). פרוטאום הנישה שהושג על ידי השילוב של CSD ושיטות MS חדישות הוכיח להיות שימושי עבור אפיון וזיהוי של רגולטורים חדשים בנישה זו NSC למבוגרים 25. לפיכך, שיטה זו תהיה שימושית לקביעת הרכב חלבון רקמת SEZ.
Protocol
כל ההליכים הניסיוניים במחקר זה בוצעו בהתאם להנחיות גרמניה והאיחוד האירופי ואושרו על ידי הוועדה המוסדית לטיפול בבעלי חיים וממשלת בוואריה העליונה (Regierung von Oberbayern). רק עכברי C57Bl6 זכרים בין הגילאים 8-10 שבועות שימשו לניסויים.
1. הכנת מוח העכבר (~ 15 דקות לעכבר)
- הכן את מדיום החיתוך על ידי הוספת 5 מ"ל של 1 M HEPES (ריכוז סופי 10 מ"מ) ל 500 מ"ל של 1x פתרון מלח מאוזן של האנק (HBSS).
הערה: זמן האחסון של אמצעי הניתוח (+4 °C)) לא יעלה על שבועיים. - להקריב את העכברים על ידי נקע צוואר הרחם ולנתח בזהירות את המוח.
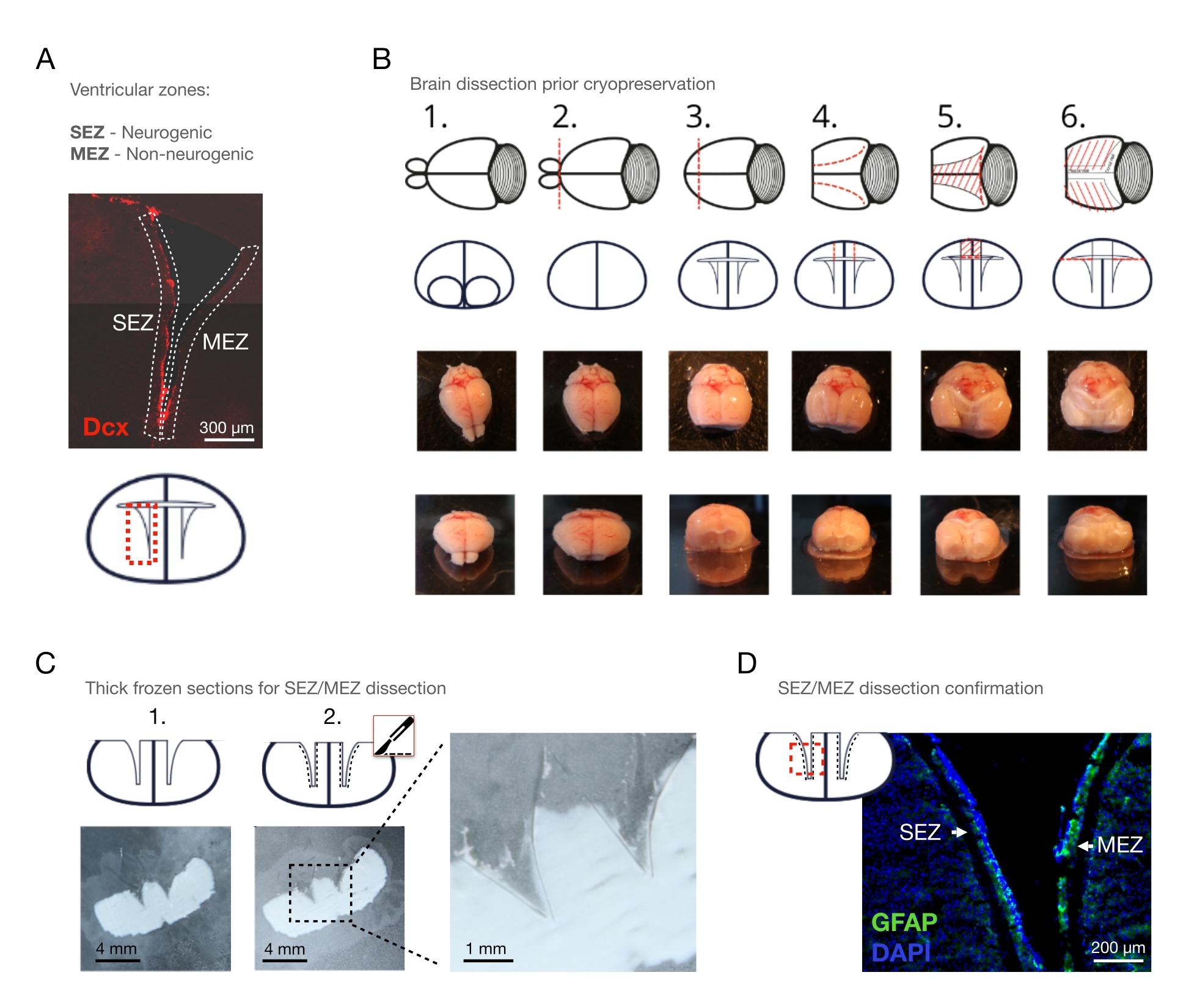
הערה: בעת חקירת ECM, הרקמה צריכה להיות לא שונה. נקע צוואר הרחם שומר על זמן הניתוח קצר ככל האפשר, ובכך מונע עיכול עצמי אנזימטי פוסט-תמותתי ככל האפשר. אם הסרת הדם היא קריטית לשאלת המחקר, פשוט להחדיר את העכבר transcardially עם תמיסת מלח חוצץ פוספט (PBS) לפני הסרת המוח. - יש לחלץ את המוח בניתוח ידני ולהניח אותו בצלחת תרבית המכילה מדיום ניתוח קר כקרח (איור 1B – 1).
הערה: שמור את המוח במדיום ביתור על קרח לאורך כל הניתוח. - הסר את נורת הריח (OB) עם אזמל (איור 1B - 2) על ידי חתך קורונל ישר בין OB לבין הקוטב הקדמי של קליפת המוח.
- הסר את הקוטב הקדמי של קליפת המוח עם האזמל באמצעות חתך קורונלי כדי להפוך את החדרים לרוחב גלויים במישור הקורונלי (איור 1B - 3).
הערה: ודא את החתך coronal נעשה ~ 5 מ"מ rostrally מן chiasm אופטי; אחרת, החלק rostral של SEZ / MEZ יאבד. - בעזרת מספריים, פתחו את שני החדרים הצדדיים מלמעלה, החל בקטע קשתי מהמשטח הקליפתי וכלה בלומן החדרי, והאריכו את החתך הזה בצורה בצורת C בעקבות כיפוף החדר (איור 1B – 4).
- חבר את הקצוות הקודליים של החתך הקשת השמאלי והימני באמצעות חתך קורונלי נוסף עם המספריים.
הערה: שלושת החתכים יוצרים כעת טרפז ויקלו על הסרת קליפת המוח והקורפוס קאלוזום בשלב הבא. - הסירו את קליפת המוח ואת הקורפוס קאלוסום המכסים את החדרים הצדדיים באמצעות מלקחיים (איור 1B – 5). לאחר מכן, להסיר את קליפת המוח ואת corpus callosum המכסים את קירות החדר המדיאלי. כאן, לבצע חתכים נוספים אם הרקמה מחוברת לקירות החדר המדיאלי, או פשוט להרים את קליפת המוח ואת corpus callosum עם מספריים כדי לחלץ את הרקמה.
- יש לפזר בזהירות את קירות החדר עם מלקחיים (איור 1B – 6). הסר את מקלעת choroid עם מלקחיים.
הערה: הסרה מלאה של מקלעת choroid חשובה כדי למנוע הפרעה עם שלבי הניתוח הבאים ולמנוע זיהום פוטנציאלי של דגימות SEZ / MEZ. - שים את המוח על שקופית זכוכית ומניחים את שקופית הזכוכית על גבי קרח יבש כדי להקפיא את המוח. שמור על קירות החדר בתצורה הפתוחה.
הערה: ודאו מרחק מספיק בין הקירות הצדדיים והמדיאליים של החדר כדי להקל על ניתוח מדויק ובלעדי של SEZ ו-MEZ. אם הרקמה מתכווצת בחזרה לתצורה סגורה, השתמש במלקחיים כדי לתקן את הקירות במצב הרצוי במהלך ההקפאה. הימנע מכל נזק ל- SEZ / MEZ. נסה להפעיל כוח מינימלי, בעיקר בקצה העליון של החדרים הפתוחים.
2. חתך של המוח המוכן (~ 15 דקות לעכבר)
- חותכים 50-100 מיקרומטר קטעים קורונליים בעובי של המוח עד סוף החדר לרוחב באמצעות cryostat ולהרכיב את החלקים על מגלשות זכוכית. ודא כי המוח מחובר ללוח ההתקשרות cryostat ב hindbrain עם מדיום OCT וכי אין OCT בא במגע עם המוח הקדמי, במיוחד בחדרים.
הערה: מדיום OCT יפריע למדידות טרשת נפוצה. עם זאת, אם הרקמה תשמש לבדיקת נוגדנים, אין צורך לא לכלול מדיום OCT. השימוש במגלשות זכוכית מצופה אינו מומלץ. שקופיות מצופות מפעילות יותר מדי כוח דבק על הרקמה, ובכך מעכבות את המעבר של דגימת הרקמה מהמגלשות לצינור המיקרוצנטריפוגה בשלבים הבאים.
3. ניתוח ביד חופשית של פרוסות המוח (~ 30 דקות לעכבר)
- הניחו את מגלשות הזכוכית עם חלקי המוח על קרח יבש מתחת למיקרוסקופ ניתוח (איור 1C - 1).
- הכן את צינורות microcentrifuge על קרח יבש, ולוודא כי הצינורות להישאר על קרח יבש לפחות 1 דקות כדי להיות קר מספיק לפני העברת דגימה.
הערה: השתמש בצינורות microcentrifuge באיכות גבוהה, כמו צינורות באיכות נמוכה מסוימים עשויים לשפוך פלסטיק בשלבי העיכול הבאים של הרקמות הקשורים מדידות טרשת נפוצה. - הרימו את הפרוסות מהקרח היבש למשך 15-30 שניות כדי להשיג הפשרה קצרה ולא שלמה כדי להפוך את המיאלין הקומפקטי של הסטריאטום לנצפות כנקודות לבנות צפופות.
הערה: איתור הגבול בין ה-SEZ לסטריאטום הופך לאפשרי (איור 1C – 2, ראו איור 2A להרחקת המיאלין ולהשוואה עם שיטת הטעינה כולה). אם הפשרה אורכת זמן רב מדי, ניתן להאיץ את התהליך על-ידי לחיצה על אצבע מכוסה כפפה בצד הנגדי של מגלשת הזכוכית. עם זאת, תמרון זה צריך להיות מתורגל כמו הפשרה מוגזמת מתרחשת בקלות. - הפרד את ה-SEZ עם אזמל מקורר מראש מהסטריאטום הסמוך (איור 1C, D).
- מעבירים את ה- SEZ כחתיכה שלמה או מחולקים ל -2-4 חלקים לצינור מיקרוצנטריפוגה באמצעות הקצה הקהה של האזמל המקורר. אם הרקמה תשמש לסוג אחר של ניתוח שאינו MS, העבר את דגימת הרקמה למיכל המתאים במקום זאת (למשל, צלחת של 96 בארות).
הערה: חיתוך הרקמה הקפואה לחלוטין עלול להוביל לפירוק מהיר של הרקמה ולנפילה מהמגלשה. חיתוך רקמה מופשרת לחלוטין מוביל להתפוררות הרקמה. ודא כי הרקמה אינה קפואה לחלוטין או מופשרת לחלוטין.
תוצאות
בעת ביצוע השלבים לעיל, דגימות הרקמה בצינורות microcentrifuge מוכנים ותואמים להכנת מדגם MS. לאחר הכנת מדגם, השגנו ~ 5-7 מיקרוגרם של פפטידים לכל מדגם של SEZ או MEZ לכל עכבר. עם זאת, הכמויות הסופיות של הפפטידים עשויות להיות תלויות בשיטת ההכנה לטרשת נפוצה. בהשוואות הפרוטאום שלהלן, זיהוי חלבונים ועומק כימות (500-1,000 חלבונים לדגימה) הוגדלו על ידי התאמה חישובית של ספקטרום הפפטיד לספריות ספקטרום פפטיד שנוצרו עבור כל אזור רקמה25,27. ראוי לציין, שיטת הפירוק ננו ללא הפסד המשמשת כאן ליצירת ספריות ספקטרום פפטיד אינו זמין כרגע מסחרית. נתוני הטרשת הנפוצה הגולמיים נותחו באמצעות תוכנת MaxQuant28, והגיעו לדיוק המוני בחלקים לכל טווח מיליארד29. סביבת Max Quant מאפשרת התאמה בין הפעלות טרשת נפוצה. שפע חלבונים היה כימות באמצעות אלגוריתם כימות ללא תוויות30. כתמים אימונוהיסטוכימיים נעשו על רקמות קפואות טריות ובוצעו כפי שדווח בעבר25 (ראו טבלת החומרים).
Cryo-section-dissection
ה-SEZ וה-MEZ המלאים של עכברים בוגרים (n = 4) הושגו באמצעות CSD (ראו איור 1 ופרוטוקול). קליפת המוח הסומטו-סנסורית (Cx) נותחה באמצעות מספריים כירורגיות. 4 עכברים נוספים נותחו באותו אופן; עם זאת, הרקמה המנותחת אוחדה לדגימה אחת לכל אזור כדי ליצור את ספריית הפרוטאום (10,923 חלבונים מזוהים) לזיהוי וכימות חלבונים מוגברים בדגימות בודדות25. בארבע הדגימות הבודדות, (ממוצע ± SD) 6,673 ± 317.4 חלבונים היו כימות ב- SEZ ו 6,747 ± 37.7 ב- MEZ. כל נתוני הפרוטאומיקה של MS הופקדו בקונסורציום ProteomeXchange באמצעות מאגר השותפים PRIDE31 , ומספר הכניסה לפרוטומים שדווחו כאן הוא ProteomeXchange: PXD016632 (http://proteomecentral.proteomexchange.org).
השוואה לחיתוך בעל טעינה מלאה
ניתוח שלם בוצע על פי פרוטוקול סטנדרטי26. ניתוח מלא חשף מספר דומה של חלבונים (כ-6,000 עבור SEZ ו-6,000 עבור Cx, n = 4 לקבוצה) בהשוואה ל- CSD25. אחד השיפורים המיועדים של שימוש ב- CSD עבור ה- SEZ, במקום פרוטוקול ביתור שלם, הוא הפחתת זיהום פוטנציאלי. בדגימות SEZ מזוהמות ברקמה מאזור אחר, חלבונים מועמדים שזוהו לא ניתן להקצות לאזור כמו העשרה משמעותית יכולה לנבוע מאזור העניין והמזהם. מבחינה חיסונית, הגליקופרוטאין (MAG) הקשור למיאלין הקשורים למיאלין, כמוסות פנימיות חיוביות עשירות במיאלין של הסטריאטום זוהו בדגימות המלאות, אך לעתים רחוקות בדגימות CSD (איור 2A). הזיהום החריצי בדגימות המלאות יכול להיות מאושר על ידי זיהוי ההעשרה של חלבוני מיאלין ב- SEZ בהשוואה לדגימות קליפת המוח הסומטו-חושית (Cx) חומר אפור (GM) (איור 2B). שים לב שחלקים גדולים של ה- Cx GM, במיוחד שכבות ה- Cx העליונות, אינם ממושמעים32.
כאשר חבילות סיבים גדולות עוברות דרך הסטריאטום, זיהום באזור זה הביא להעשרת חלבוני המיאלין בהשוואה ל- Cx. חלבוני המיאלין ששימשו כסמנים לזיהום סטריאטלי בדגימות SEZ היו החלבון הבסיסי של המיאלין (MBP), הגליקופרוטאין הקשור למיאלין (MAG), הפרוטאוליפיד-חלבון 1 (Plp1) ו-2',3'-ציקלאי-נוקלאוטיד 3'-פוספודיסטראז (Cnp). כל חלבוני סמן המיאלין הועשרו באופן משמעותי ב-SEZ בהשוואה ל-Cx. לעומת זאת, ההשוואות לארבעת חלבוני סמן המיאלין בערכת הנתונים של CSD לא הניבו הבדלים משמעותיים בהשוואת SEZ ל- Cx (איור 2B). נתונים פרוטאומיים של הסטריאטום33 תומכים בהשערה כי העשרת חלבוני המיאלין בדגימות SEZ של הניתוח כולו נגרמה על ידי הזיהום עם רקמת סטריאטל. לפיכך, CSD מנע במידה רבה זיהום על ידי רקמת סטריאטל (עשיר במיאלין קומפקטי) לעומת ביתור שלם.
ניתוח פרוטאום לא משוחד של רקמות לא מנותקות יכול לחשוף חלבונים חוץ תאיים מעניינים. עם ניתוח משופר באמצעות ה-CSD, חלבונים חוץ-תאיים הועשרו באופן משמעותי בדגימות בהשוואה לדגימות המלאות (איור 2C, בדיקת העשרת ביאורים). ה- CSD והניתוח המלא מציגים העשרה דומה של המונחים אונטולוגיה של הגנים (GO) "אקסוזום שלפוחית חוץ-תאית" ו"חלק אזור חוץ-תאי". עם זאת, המונח GO "Matrisome הקשורים" מועשר מעט יותר ב- CSD מאשר בחיתוך כולו. בהתאם לכך, האנזים חוצה מחייב ECM והתגלה לאחרונה נוירוגנזה רגולטור transglutaminase-2 (Tgm2) נמצאו מועשרים SEZ לעומת Cx באמצעות CSD25. לעומת זאת, לא נמצא הבדל בין דגימות SEZ ו- Cx שהושגו על-ידי הניתוח המלא (איור 2D). נתונים פרוטאומיים של striatum33 תומכים בהשערה כי הגילוי של רגולטור neurogenesis Tgm2 על ידי ניתוח שלם היה מונע על ידי הזיהום עם רקמה סטריאטלית. לפיכך, בסך הכל, ההקפאה-סעיף-ניתוח הוא שיפור מוצלח אך גם הכרחי לחיתוך הסטנדרטי לניתוח פרוטאום ספציפי לנישה.
השוואה למיקרוסקופיה של לכידת לייזר
החצי הקדמי של ה-SEZ וה-MEZ של 3 עכברים בוגרים הושגו עבור LCM (איור 3A). בסך הכל, שיטת LCM מציגה כמה חסרונות, במיוחד לגבי הפרעה רקמות ויעילות. כדי לדמיין את אזור העניין תחת מיקרוסקופ החיתוך, יש צורך בהכתמת רקע, שעלולה לשטוף חלבונים קטנים או מסיסים של עניין, למשל, גורמי גדילה, ציטוקינים או רגולטורים ECM כגון אנזימים. יתר על כן, שקופיות מבלות פעמים שונות בטמפרטורת החדר במהלך הסרת לייזר. יתר על כן, הלייזר עצמו עשוי לפגוע בחלבונים בעלי עניין.
ל- CSD יש יתרון ניכר על פני LCM לגבי הזמן והמאמץ הדרושים לביצוע הניתוח: שלב 1 של הפרוטוקול חייב להתבצע באופן דומה הן עבור CSD והן עבור LCM; ללא שלב זה, קירות החדר נשארים דבקים, מה שהופך את ההפרדה של דגימות MEZ ו- SEZ קשה. בהתחשב בכך שסעיפי ה- CSD (100 מיקרומטר) עבים פי 6-7 מהעובי המרבי34 של מקטעי LCM (15 מיקרומטר), שלב 2 (חתך של המוח) ושלב 3 (הסרת MEZ ו- SEZ מכל מקטע קורונלי) ייקח לפחות 6-7 פעמים יותר זמן עבור LCM. הכתמת הרקע הדרושה והגדרת מיקרוסקופ הלייזר ידרשו זמן נוסף. כאן, זה לקח שלוש פעמים יותר זמן לקצור 50% של SEZ ו MEZ של 3 בעלי חיים על ידי LCM לעומת 100% של SEZ ו MEZ של 4 בעלי חיים על ידי CSD, המהווה יתרון מהירות פי שמונה של CSD. לסיכום, LCM לא רק דורש כמות ניכרת של מאמץ נוסף, אבל הרקמה כפופה גם לתקופה ארוכה יותר באופן משמעותי של מניפולציה ושינויי טמפרטורה שיכולים לסכן את הדינמיקה והאמינות של נתונים שנוצרו על ידי ניתוח עוקב.
תוצאות הטרשת הנפוצה של CSD הושוו לתוצאות המיקרודיסקציה של לכידת הלייזר (LCM). שתי ערכות הנתונים הותאמו לספריה הפרוטאומית שנוצרה על-ידי איגום דגימות CSD. בממוצע, LCM הניב 3,441 ± 270.0 ו-3,613 ± 238.7 חלבונים בודדים באזור SEZ וחדרי הביניים, בהתאמה (איור 3B). בהתחשב בהבדל המדהים בזיהוי חלבונים, ניתוח רכיבים עיקרי (PCA) הציג הפרדה ברורה על פי שיטת הניתוח (רכיב 1: 62.7%, לא מוצג). רכיב 2 הציג את ההפרדה הגדולה ביותר עבור SEZ ו- MEZ בין דגימות LCM (8.5%, איור 3C). רכיב 3 נראה גם להפריד LCM ו- CSD; עם זאת, הבדל זה עשוי לנבוע מהבדלים מבוססי שיטה ולא ממספר החלבונים המזוהים (6.4%). עם זאת, ההפרדה האזורית הכוללת נותרה בולטת להפליא עבור נתוני ההקפאה-ניתוח וטובה בהרבה מאשר עבור LCM. אי התאמה זו בדינמיקת הנתונים עשויה לנבוע מזמנים שונים שהושקעו על ידי הדגימות בטמפרטורת החדר במהלך ניתוח הלייזר או רגישות גבוהה יותר של כמויות רקמות קטנות לשונות בפרוטוקולי הפרוטאומיקה הבאים ובמדידות ספקטרומטריית המסה.
כדי לחפש הבדלים בפרופיל הפרוטאום של ה-ECM, בוצעה בדיקת העשרת ביאורים דו-ממדית בין CSD ל-LCM עבור ה-SEZ וה-MEZ (איור 3D). חישוב ההעשרה היחסית של מונחי GO בין דגימות LCM ו- CSD מאפשר השוואה של דינמיקת פרוטאום יחסית של אשכולות חלבון ECM בין שתי השיטות למרות כמות הרקמה הלא שוויונית וההבדלים בפרוטוקול הניתוח. העלילות חושפות מתאם טוב בין LCM ל- CSD. הביאורים "חלק אזור חוץ תאי" ו "אברון קרום חוץ תאי" מועשרים באופן דומה בשתי השיטות והאזורים. לפיכך, הביקוש המוגבר לזמן של LCM לא נראה פיצוי על ידי רגישות גבוהה יחסית עבור חלבונים הקשורים ECM. במקום זאת, CSD מספק זיהוי/כימות חזקים יותר בעת השוואת נתוני המדגם עבור חלבוני ECM הקשורים לנוירוגנזה ול- SEZ Tgm2, תרומבוסופונדין-4 (Thbs4), S100a6 וטנסין-C (Tnc) (איור 3E). במקרה של TnC, למרות כימות בכל הדגימות, רק CSD הציג העשרה עבור SEZ לעומת MEZ. עם זאת, חלבוני קרום הבסיס הקשורים SEZ Nidogen-1 (Nid1), Laminin subunit beta-2 (Lamb2), ו מרתף קרום ספציפי heparan גופרתי פרוטאוגליקן חלבון הליבה (Hspg2)35 הציג העשרה חזקה עוד יותר SEZ (לעומת MEZ) בדגימות LCM מאשר בדגימות CSD (לא מוצג). לפיכך, CSD יכול לספק דגימות רקמה המספקות פרוטאום כמותי מדויק ועמוק לאפיון SEZ במסגרת זמן סבירה, מבלי לדאוג לגבי שלמות רקמות נפגעת או אובדן חלבון.
סטטיסטיקה
בדיקות סטטיסטיות, בדיקות העשרת ביאורים דו-ממדיות ו-PCA נעשו בסביבת פרסאוס. חלבונים נכללו בניתוח אם זוהה ערך חוקי עבור כל שיטה במדגם אחד לפחות. שפע חלבונים והשוואות מספרים הוצגו באופן חזותי באמצעות תוכנת ניתוח נתונים (ראו טבלת החומרים). בקרה המבוססת על תמורות של שיעור הגילוי הכוזב (FDR) (רוזוולט הוגדר ל- 0.05, 250 אקראיים) שימשה להשוואות חלבונים. עבור בדיקות העשרת ביאור דו-ממדי36, מונחי GO המוצגים מועשרים באופן משמעותי (רוזוולט הוגדר ל- 0.02 בשיטת בקרת ה-FDR של Benjamini-Hochberg).

איור 1: שיטת ההקפאה-סעיף-ניתוח. (א) סקירה כללית של אזור העניין: החדר הצדדי עם ה-SEZ הנוירוגני וה-MEZ הלא-נוירוגני. נוירובלסטים מחוסנים עם DCX. (B) הסרת Stepwise של OB, הקוטב הקדמי, קליפת המוח, וקורפוס callosum מעל החדרים ואת מקלעת choroid: 1. מיקום במדיום ניתוח, 2. הסרת OB, 3. הסרת הקוטב הקדמי של קליפת המוח, 4. חתכים קשתיים של החלק העליון של החדר, 5. הסרת החלק העליון של החדר, 5. הסרת החלק העליון של החדר, 6. התפשטות קירות החדר. (C) 100 מיקרומטר פרוסות קורונל של מוח העכבר הקפוא הטרי, (1.) לפני ו -(2.) לאחר הסרת קירות החדר עם אזמל קר כקרח. סרגלי קנה מידה = 4 מ"מ (D) כתמים של קטע קורונלי של חדר לרוחב (GFAP: ירוק; דאפי: כחול), מראה את SEZ ו MEZ ניתח עם CSD. סרגלי קנה מידה = 300 מיקרומטר (A), 200 מיקרומטר (D). קיצורים: CSD = ניתוח קריו-סעיף; SEZ = אזור תת-ייצוגי; MEZ = אזור אפנדימלי מדיאלי; Dcx = Doublecortin; OB = נורת חוש הריח; GFAP = חלבון חומצי פרברי גליה; DAPI = 4′,6-diamidino-2-פנילינדולה. לחץ כאן כדי להציג גירסה גדולה יותר של איור זה.
{kind=link}

איור 2: דיוק חתך מעולה עם ההקפאה-קטע-ניתוח בהשוואה לחתך בעל משקל מלא. (A) תמונה אימונוהיסטוכימית של דגימת SEZ שהתקבלה על ידי ניתוח מלא (משמאל). ההכללה של רקמת סטריאטל עשיר במיאלין הוא דמיינו על ידי כתמים נגד MAG (ירוק). הכתמת SEZ המנותח באמצעות ה- CSD (מימין). ב- CSD, כמעט כל המיאלין striatal (כתמים נגד MAG, ירוק) אינו נכלל בסרט המדגם. גרעינים הוצגו באמצעות DAPI (כחול). (B) השוואה של העשרת סמן מיאלין ב- SEZ לעומת Cx מ- wholemount (MBP: p = 0.0074; MAG: p = 0.0016; Plp1: p = 0.0011; CNP: p = 0.0029) ו- CSD (MBP: p = 0.0667; MAG: p = 0.0236; Plp1: p = 0.3420; CNP: p = 0.1842). (C) מבחן העשרה ביאור דו-ממדי המשווה את כל ה-SEZ עם דגימות CSD-SEZ. המונחים GO מרחב חוץ-תאי ו- Matrisome הקשורים למטריזום מועשרים יותר בנתוני ה- CSD מאשר בנתונים המלאים. (ד) שפע החלבון של הרגולטור המל"ל Tgm225 שתוכנן עבור הניתוח המלא וה- CSD. Tgm2 מועשר באופן משמעותי ב- SEZ בהשוואה ל- Cx ב- CSD (CSD: p = 0.0029; שלם: p = 0.1775). עבור B ו- D: כהפניה, נתוני פרוטאום מ שארמה ואח ' 33 עם מדידות של סטריאטום וקליפת המוח שרטטו עבור החלבונים המתאימים המוצגים בדגימות כל הבמה ו- CSD. סרגלי קנה מידה = 200 מיקרומטר (A). קיצורים: CSD = ניתוח קריו-סעיף; SEZ = אזור תת-ייצוגי; MAG = גליקופרוטאין הקשור למיאלין; Cx = קליפת המוח הסומטו-חושית; MBP = חלבון בסיסי מיאלין; Plp1 = פרוטאוליפיד-חלבון 1; CNP = 2',3'-מחזורי-נוקלאוטיד 3'-phosphodiesterase; GO = אונטולוגיה גנטית; NSC = תא גזע עצבי; Tgm2 = tranglutaminase 2; DAPI = 4′,6-diamidino-2-פנילינדולה; LFQ = כמות ללא תוויות. לחץ כאן כדי להציג גירסה גדולה יותר של איור זה.
{kind=link}

איור 3: כימות חלבון חוץ-תאי משופר עם ניתוח החלקה בהקפאה בהשוואה ל- LCM. (A) כתמים סגולים של קרסיל של חדר לרוחב לפני ואחרי לכידת הלייזר של ה- SEZ וה- MEZ (משמאל). לשם השוואה, חתך CSD של SEZ ו- MEZ (מימין). סרגלי קנה מידה = 150 מיקרומטר. (B) השוואה של מספר החלבונים שזוהו בדגימות SEZ ו- MEZ מ- CSD ו- LCM. הנתונים מוצגים כממוצע ± SD. (C) ניתוח רכיבים עיקרי של דגימות SEZ ו- MEZ המשווה CSD ו- LCM (רכיב 2: 8.5% מהשונות; רכיב 3: 6.4%). (D) העשרת ביאור דו-ממדי של ה-MEZ (למעלה) ו-SEZ (חלק עליון) ו-SEZ (למטה). המונחים GO אברון חוץ-תאי וחלק אזור חוץ-תאי מועשרים באופן משמעותי (נקודות אדומות). (E) שפע של חלבוני סמן חוץ-תאיים הקשורים SEZ ב- SEZ ו- MEZ עבור LCM (Tnc: p = 0.3789) ודגימות CSD (Tgm2: p = 0.2940; S100a6: p = 0.0218; THBS4: p = 0.3941; Tnc: p = 0.0004). קיצורים: CSD = ניתוח קריו-סעיף; LCM = לייזר-לכידת-microdissection; SEZ = אזור תת-ייצוגי; MEZ = אזור אפנדימלי מדיאלי; GO = אונטולוגיה גנטית; Tnc = Tenacin-C; Tgm2 = transglutaminase 2; S100a6 = S100 חלבון מחייב סידן A6; THBS4 = תרומבוסופונדין-4; LFQ = כמות ללא תוויות. לחץ כאן כדי להציג גירסה גדולה יותר של איור זה.
{kind=link}
Discussion
שיטת CSD אפשרה לחלץ במדויק רקמת SEZ וליצור פרוטאום אמין עם עומק משמעותי באמצעות MS. CSD מציג יתרון ברור בהשוואה לחיתוך מלא במונחים של זיהום סטריאטלי מופחת מאוד של דגימות SEZ והעשרת חלבונים חוץ תאיים. כפי שניתן גם לזהות מספר דומה של חלבונים בדגימות בודדות (~ 6,500 חלבונים לדגימה) עם CSD וחיתוך מלא, הזמן הנוסף עבור CSD שווה את המאמץ. LCM מספק ניתוח SEZ מדויק יותר אך הגיע לעומק פרוטאום נמוך יותר, עם רק 3,500 חלבונים לדגימה למרות שהשתמש באותו פרוטוקול MS כמו CSD (התאמת ספריה וכימות ללא תוויות). חשוב לציין שהשונות הייתה גדולה בהרבה, כנראה בגלל זמן ההכנה הארוך פי שמונה לדגימה. PCA של הדגימות שהושגו על ידי LCM ו- CSD חושף הפרדה ברורה של שתי השיטות עם אשכולות הדוקים ספציפיים לאזור המופרדים בחוזקה זה מזה. לעומת זאת, דגימות LCM הציגו התפלגות מפוזרת יותר, אשר כנראה בחלקה בשל אורך ההכנה. לא ברור אם איסוף דגימות רבות יותר על פני תקופה ארוכה יותר היה מניב פרוטאום של חוסן ועומק שווים עם LCM. חישוב הערכה, איסוף נפח מדגם דומה כפי שנעשה עבור CSD ייקח 5-8 פעמים יותר עם LCM, אפילו עד פי 15 יותר אם דגימות שסופקו עבור ספריות ספקטרום פפטיד נכללו, וחלק גדול ממנו בתנאים מופשרים. יתר על כן, בהתחשב בהפרעות הנוספות של הרקמה הדרושה ל- LCM (כתמי רקע, ניתוח לייזר), LCM סיפק מעט, אם בכלל, רווח על פני CSD. לפיכך, CSD יכול להיחשב מתאים יותר למחקר פרוטאום חוץ תאי, במיוחד עבור SEZ.
ראוי לציין, אם אזור העניין קטן יותר מאשר SEZ (למשל, חוקר רק את שכבת התא ependymal), גישה חופשית נופלת מאחורי הדיוק של LCM. לדוגמה, השימוש ב- CSD להפרדת האפנדימלי לשכבה התת-קרקעית קשה מכיוון שהשכבה האפנדימלית היא רק קוטר תא רחב, והתיחום כלפי השכבה התת-כספית אינו גלוי לעין בלתי ברקמה קפואה טרייה. לפיכך, LCM יהיה בחירה טובה יותר מאשר CSD אם ניתוח מדויק בקנה מידה מתחת 50 מיקרומטר חשוב יותר מאשר רקמה ללא הפרעה או שמירה על זמן הניתוח קצר. עבור אזורים עם רוחב של 50 מיקרומטר ויותר, עם זאת, הדיוק של CSD דומה לזה של LCM לניתוח חלבון ECM.
CSD כבר הוכיח להיות שימושי על ידי תרומה לחקירה של התפקיד התפקודי של ECM בנישה neurogenic25. לפיכך, המשך היישום של CSD ב- SEZ עבור חקירות חלבון ופרוטום שונים (או אפילו רצף RNA גרעין יחיד) עשוי להוביל לאיתור של ויסות נוירוגנזה נוספים, סמני הפעלת תאי גזע, והבנה עמוקה יותר של פיזיולוגיית נישה של תאי גזע SEZ. בהתחשב בירידה של neurogenesis בהזדקנות SEZ37, ניתוח תמציתי של שינויים ECM של SEZ של עכברים בגיל לעומת צעירים עשוי לקדם את ההבנה של מנגנוני נישה מדויקים טיפוח פיתוח ותחזוקה NSC38,39. יתר על כן, ההשפעה של דלקת ופציעה על נוירוגנזה SEZ מבוססת היטב40,41,42,43. ההעשרה של פיברינוגן שמקורו בדם ב- SEZ לאחר פגיעה מוחית בקליפת המוח והשפעתה על אסטרוגלוגנזה SEZ והיווצרות צלקת44 מדגישה את ההשפעה הפוטנציאלית של שינויי מיקרו-סביבה הנגרמים מטראומה על הפיזיולוגיה של תאי הגזע SEZ. לפיכך, חקירת פרוטאום SEZ-ECM בשיתוף עם פגיעה מוחית באמצעות CSD יכול לעזור להבהיר את המנגנונים שבאמצעותם פגיעה ודלקת להשפיע על neurogenesis. חשוב לציין, השיטה יכולה להיות ישימה גם נישות neurogenic המוח האנושי בבריאות ומחלות כמו רקמה קפואה טרי לעתים קרובות ניתן להשיג מניתוחים. יתר על כן, בהתחשב בהבדלים המינים נוירוגנזה בוגרת, זה יהיה גם מרתק ליישם את שיטת CSD על מינים אחרים, למשל, בשיתוף נוירוגנזה striatal. יתר על כן, עם שיטות אחרות לזיהוי חלבונים, הבדלים בגורמי גדילה המיוצרים באופן מקומי ניתן לחקור במדויק וביעילות באמצעות CSD עבור SEZ ו- MEZ (למשל, ELISA).
לבסוף, הליך הניתוח יכול להיות שונה באופן פוטנציאלי עבור מיצוי מדויק של אזורי מוח אחרים, גם עבור שאלות מחקר שאינן קשורות neurogenesis. לדוגמה, CSD כולל צעד הפשרה למחצה קצר, שבמהלכו מיאלין קומפקטי נראה כאזורים לבנים הנבדלים מרקמת המוח השיורית השקוף יותר. עם שינוי פשוט של השיטה, תכונה זו תאפשר את הניתוח המדויק של רקמת מיאלין קומפקטית קורפוס callosum בלבד, אשר יכול להיות נתון לניתוח פרוטאומי של שינויים הקשורים לפציעה. הצעה לשינוי פרוטוקול שיאפשר את הניתוח הקשוח של הקורפוס היא להשמיט את שלבים 1.5-1.9 של הפרוטוקול ולהמשיך ישירות להכנת קטעי הקורונה במקום לפתוח את החדרים כדי להפוך את SEZ ו MEZ נגישים. לאחר מכן, מניחים את החלקים על קרח יבש, להרים לזמן קצר ולהפשיר למחצה את הפרוסות, ופשוט להסיר את קורפוס callosum עם אזמל. הכנה זו צריכה להיות מוכנה כעת לכל ניתוח הדורש ניתוח יעיל של רקמת קורפוס קאלוזום מקומית.
לסיכום, מחקר זה מציג שיטת מיקרו-ניתוח שיכולה לשמש לניתוח פרוטאום נישה נוירוגנית חדרית אמינה. הנתונים מדגישים את התאימות והתועלת של שיטת CSD יחד עם ניתוח פרוטאומי מבוסס MS של מיקרו-סביבה SEZ. השילוב של דיוק, יעילות וסטירת רקמות מינימלית הופכים את ה- CSD להרחבה רבת ערך של שיטות קיימות.
Disclosures
המחברים מצהירים שאין אינטרסים מתחרים
Acknowledgements
אנו מבקשים להודות בכנות למתיאס מאן על שאיפשר לנו לבצע חלקים גדולים מהניסויים במעבדה שלו, פביאן קוסיה על העזרה עם LCM וניתוח פרוטאום, טטיאנה סימון-אברט על ניתוחים מלאים, וקורביניאן מאייר ואיגור פארון על עזרתם הטכנית. אנו מודים על מימון ממועצת המחקר הגרמנית ל- MG (SFB870, TFR274), האיחוד האירופי (Eranet S-700982-5008-001), ה- ERC (aERC "NeuroCentro" ל- MG), האגודה השבדית למחקר רפואי (SSMF, ל- JK) מענק פוסט-דוקטורט, וקרנות KI וקרנות (2020-01351, ל- JK).
Materials
| Name | Company | Catalog Number | Comments |
| Cryostat CM3050S | Leica | ||
| Dissecting microscope | Leica | ||
| Dumont no. 5SF forceps, Inox super fine tip | Fine Science Tools | cat. no. 11252-00 | |
| Hank’s Balanced Salt Solution with CaCl2 and MgCl2 | Invitrogen | cat. no. 24020 | |
| HEPES buffer solution (1 M) | Invitrogen | cat. no. 15630 | |
| Microscope slides | RS France | cat. no. BPB018 | |
| Safe-lock tubes, PCR clean 2.0 mL | Eppendorf | cat. no. 0030123344 | |
| Spring scissors, Vannas-Tubingen 5 mm | Fine Science Tools | cat. no. 15003-08 | |
| Surgical disposable scalpels | B. Braun | cat. no. 5518083 | |
| Tissue culture dishes 60 mm | Greiner Bio-One | cat. no. 633180 | |
| Antibodies | |||
| Alexa Fluor secondary antibodies (488, 555) (1/1,000) | ThermoFisher Scientific | cat. no. A-11001 | |
| DAPI | Sigma | cat. no. D9542 | |
| guinea pig polyclonal anti-DCX 1:500 | Millipore | cat. no. AB2253, | |
| mouse monoclonal anti-GFAP 1:500 | Sigma | cat. no. G3893 | |
| mouse monoclonal anti-MAG 1:400 | Millipore | cat. no. MAB1567 | |
| Software | |||
| GraphPad Prism version 9 | GraphPad Software, San Diego California USA | www.graphpad.com | |
| Perseus Version 1.6.10.50 | Max-Planck Institute for Biochemistry, Munich Bavaria Germany | https://maxquant.net/perseus/ | |
| ZEN imaging software | Carl Zeiss |
References
- Goodman, T., Hajihosseini, M. K. Hypothalamic tanycytes-masters and servants of metabolic, neuroendocrine, and neurogenic functions. Frontiers in Neuroscience. 9, 387(2015).
- Chaker, Z., et al. Hypothalamic neurogenesis persists in the aging brain and is controlled by energy-sensing IGF-I pathway. Neurobiology of Aging. 41, 64-72 (2016).
- Kuhn, H. G., Toda, T., Gage, F. H. Adult hippocampal neurogenesis: a coming-of-age story. Journal of Neuroscience. 38 (49), 10401-10410 (2018).
- Obernier, K., Alvarez-Buylla, A. Neural stem cells: origin, heterogeneity and regulation in the adult mammalian brain. Development. 146 (4), (2019).
- Obernier, K., et al. Adult neurogenesis is sustained by symmetric self-renewal and differentiation. Cell Stem Cell. 22 (2), 221-234 (2018).
- Bordiuk, O. L., Smith, K., Morin, P. J., Semënov, M. V. Cell proliferation and neurogenesis in adult mouse brain. PloS One. 9 (11), 111453(2014).
- Doetsch, F., García-Verdugo, J. M., Alvarez-Buylla, A. Cellular composition and three-dimensional organization of the subventricular germinal zone in the adult mammalian brain. Journal of Neuroscience. 17 (13), 5046-5061 (1997).
- Lim, D. A., Alvarez-Buylla, A. The adult ventricular-subventricular zone (V-SVZ) and olfactory bulb (OB) neurogenesis. Cold Spring Harbor Perspectives in Biology. 8 (5), 018820(2016).
- Sato, K. Effects of microglia on neurogenesis. Glia. 63 (8), 1394-1405 (2015).
- Tavazoie, M., et al. A specialized vascular niche for adult neural stem cells. Cell Stem Cell. 3 (3), 279-288 (2008).
- Sirko, S., et al. Chondroitin sulfates are required for fibroblast growth factor-2-dependent proliferation and maintenance in neural stem cells and for epidermal growth factor-dependent migration of their progeny. Stem Cells. 28 (4), 775-787 (2010).
- Mercier, F. Fractones: extracellular matrix niche controlling stem cell fate and growth factor activity in the brain in health and disease. Cellular and Molecular Life Sciences: CMLS. 73 (24), 4661-4674 (2016).
- Mercier, F., Kitasako, J. T., Hatton, G. I. Anatomy of the brain neurogenic zones revisited: fractones and the fibroblast/macrophage network. Journal of Comparative Neurology. 451 (2), 170-188 (2002).
- Nascimento, M. A., Sorokin, L., Coelho-Sampaio, T. Fractone bulbs derive from ependymal cells and their laminin composition influence the stem cell niche in the subventricular zone. Journal of Neuroscience. 38 (16), 3880-3889 (2018).
- Chen, G., et al. In vivo reprogramming for brain and spinal cord repair. eNeuro. 2 (5), 0106-0115 (2015).
- Zhang, Q., Chen, W., Tan, S., Lin, T. Stem cells for modeling and therapy of Parkinson's disease. Human Gene Therapy. 28 (1), 85-98 (2017).
- Wei, C., Xiong, S., Cheng, L. Reprogramming of fibroblasts to neural stem cells by a chemical cocktail. Methods in Molecular Biology. 2117, 265-270 (2020).
- Tian, Z., Zhao, Q., Biswas, S., Deng, W. Methods of reactivation and reprogramming of neural stem cells for neural repair. Methods. 133, 3-20 (2018).
- Heinrich, C., et al. Sox2-mediated conversion of NG2 glia into induced neurons in the injured adult cerebral cortex. Stem Cell Reports. 3 (6), 1000-1014 (2014).
- Masserdotti, G., et al. Transcriptional mechanisms of proneural factors and REST in regulating neuronal reprogramming of astrocytes. Cell Stem Cell. 17 (1), 74-88 (2015).
- Seidenfaden, R., Desoeuvre, A., Bosio, A., Virard, I., Cremer, H. Glial conversion of SVZ-derived committed neuronal precursors after ectopic grafting into the adult brain. Molecular and Cellular Neurosciences. 32 (1-2), 187-198 (2006).
- Lepko, T., et al. Choroid plexus-derived miR-204 regulates the number of quiescent neural stem cells in the adult brain. EMBO Journal. 38 (17), 100481(2019).
- Silva-Vargas, V., Maldonado-Soto, A. R., Mizrak, D., Codega, P., Doetsch, F. Age-dependent niche signals from the choroid plexus regulate adult neural stem cells. Cell Stem Cell. 19 (5), 643-652 (2016).
- Angelidis, I., et al. An atlas of the aging lung mapped by single cell transcriptomics and deep tissue proteomics. Nature Communications. 10 (1), 963(2019).
- Kjell, J., et al. Defining the adult neural stem cell niche proteome identifies key regulators of adult neurogenesis. Cell Stem Cell. 26 (2), 277-293 (2020).
- Mirzadeh, Z., Doetsch, F., Sawamoto, K., Wichterle, H., Alvarez-Buylla, A. The subventricular zone en-face: wholemount staining and ependymal flow. Journal of Visualized Experiments: JoVE. (39), e1938(2010).
- Kulak, N. A., Geyer, P. E., Mann, M. Loss-less nano-fractionator for high sensitivity, high coverage proteomics. Molecular & Cellular Proteomics: MCP. 16 (4), 694-705 (2017).
- Tyanova, S., Temu, T., Cox, J. The MaxQuant computational platform for mass spectrometry-based shotgun proteomics. Nature Protocols. 11 (12), 2301-2319 (2016).
- Cox, J., Mann, M. MaxQuant enables high peptide identification rates, individualized p.p.b.-range mass accuracies and proteome-wide protein quantification. Nature Biotechnology. 26 (12), 1367-1372 (2008).
- Cox, J., et al. Accurate proteome-wide label-free quantification by delayed normalization and maximal peptide ratio extraction, termed MaxLFQ. Molecular & Cellular Proteomics: MCP. 13 (9), 2513-2526 (2014).
- Perez-Riverol, Y., et al. The PRIDE database and related tools and resources in 2019: improving support for quantification data. Nucleic Acids Research. 47 (1), 442-450 (2019).
- Tomassy, G. S., et al. Distinct profiles of myelin distribution along single axons of pyramidal neurons in the neocortex. Science. 344 (6181), 319-324 (2014).
- Sharma, K., et al. Cell type- and brain region-resolved mouse brain proteome. Nature Neuroscience. 18 (12), 1819-1831 (2015).
- Datta, S., et al. Laser capture microdissection: Big data from small samples. Histology and Histopathology. 30 (11), 1255-1269 (2015).
- Kerever, A., et al. Novel extracellular matrix structures in the neural stem cell niche capture the neurogenic factor fibroblast growth factor 2 from the extracellular milieu. Stem Cells. 25 (9), 2146-2157 (2007).
- Cox, J., Mann, M. 1D and 2D annotation enrichment: a statistical method integrating quantitative proteomics with complementary high-throughput data. BMC Bioinformatics. 13, Suppl 16 12(2012).
- Daynac, M., Morizur, L., Chicheportiche, A., Mouthon, M. -A., Boussin, F. D. Age-related neurogenesis decline in the subventricular zone is associated with specific cell cycle regulation changes in activated neural stem cells. Scientific Reports. 6, 21505(2016).
- Navarro Negredo, P., Yeo, R. W., Brunet, A. Aging and rejuvenation of neural stem cells and their niches. Cell Stem Cell. 27 (2), 202-223 (2020).
- Smith, L. K., White, C. W., Villeda, S. A. The systemic environment: at the interface of aging and adult neurogenesis. Cell and Tissue Research. 371 (1), 105-113 (2018).
- Neuberger, E. J., Swietek, B., Corrubia, L., Prasanna, A., Santhakumar, V. Enhanced dentate neurogenesis after brain injury undermines long-term neurogenic potential and promotes seizure susceptibility. Stem Cell Reports. 9 (3), 972-984 (2017).
- Fisch, U., Brégère, C., Geier, F., Chicha, L., Guzman, R. Neonatal hypoxia-ischemia in rat elicits a region-specific neurotrophic response in SVZ microglia. Journal of Neuroinflammation. 17 (1), 26(2020).
- Götz, M., Sirko, S., Beckers, J., Irmler, M. Reactive astrocytes as neural stem or progenitor cells: In vivo lineage, In vitro potential, and Genome-wide expression analysis. Glia. 63 (8), 1452-1468 (2015).
- Kernie, S. G., Parent, J. M. Forebrain neurogenesis after focal Ischemic and traumatic brain injury. Neurobiology of Disease. 37 (2), 267-274 (2010).
- Pous, L., et al. Fibrinogen induces neural stem cell differentiation into astrocytes in the subventricular zone via BMP signaling. Nature Communications. 11 (1), 630(2020).
Reprints and Permissions
Request permission to reuse the text or figures of this JoVE article
Request PermissionExplore More Articles
This article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. All rights reserved