
Method Article
חיתוך גזע המוח השמיעתי של העוף העוברי להערכת שיפועים טונוטופיים ומיקרו-מעגלים
In This Article
Summary
כאן אנו מציגים פרוטוקול להשגת פרוסות גזע מוח שמיעתיות שאינן קורונליות של עובר העוף לחקר תכונות טונוטופיות ומסלולים התפתחותיים בתוך פרוסת גזע מוח אחת. פרוסות אלה כוללות מקטעים סגיטליים, אופקיים ואופקיים/רוחביים המקיפים אזורים טונוטופיים גדולים יותר בתוך מישור פרוסה בודד שהמקטעים הקורונליים המסורתיים.
Abstract
עובר התרנגולת הוא מודל חייתי מקובל לחקר גזע המוח השמיעתי, המורכב ממיקרו-מעגלים מיוחדים מאוד וטופולוגיה עצבית בכיוון דיפרנציאלי לאורך ציר טונוטופי (כלומר, תדר). הציר הטונוטופי מאפשר קידוד מופרד של צלילים בתדר גבוה במישור הרוסטרלי-מדיאלי וקידוד בתדר נמוך באזורים קאודו-לרוחביים. באופן מסורתי, פרוסות גזע מוח קורונליות של רקמה עוברית מאפשרות לחקור למינה בתדר ISO אינדיבידואלי יחסית. אף על פי שמספיק לחקור שאלות אנטומיות ופיזיולוגיות הנוגעות לאזורים בודדים בתדר איזו, חקר השונות הטונוטופית והתפתחותה על פני אזורים גדולים יותר בגזע המוח השמיעתי מוגבל במקצת. פרוטוקול זה מדווח על טכניקות חיתוך גזע המוח מעוברי תרנגולות הכוללות שיפועים גדולים יותר של אזורי תדר בגזע המוח השמיעתי התחתון. השימוש בשיטות חיתוך שונות לרקמת גזע המוח השמיעתית של עוף מאפשר ניסויים אלקטרופיזיולוגיים ואנטומיים בתוך פרוסת גזע מוח אחת, שבה נשמרים טוב יותר שיפועים גדולים יותר של תכונות טונוטופיות ומסלולים התפתחותיים מאשר חתכים קורונליים. טכניקות חיתוך מרובות מאפשרות חקירה משופרת של התכונות האנטומיות, הביופיזיות והטונוטופיות המגוונות של מיקרו-מעגלים שמיעתיים בגזע המוח.
Introduction
עובר התרנגולת הוא מודל מחקרי רב ערך לחקר שאלות ביולוגיות בסיסיות בתחומים מדעיים רבים ומגוונים, כולל ביולוגיה של התא, אימונולוגיה, פתולוגיה ונוירוביולוגיה התפתחותית. המיקרו-מעגלים של גזע המוח השמיעתי של העוף הם דוגמה מצוינת למעגל מיוחד מאוד שניתן להבין במונחים של מורפולוגיה שמיעתית ופיזיולוגיה. לדוגמה, רובל ופארקים (1975) תיארו לראשונה את האוריינטציה הטונוטופית (כלומר, שיפוע התדרים) של גרעין העוף מגנוצלולריס (NM) וגרעין למינריס (NL) כפונקציה ליניארית על פני ציר הגרעינים, בכיוון ~30° ביחס למישור הסגיטאלי. תאי עצב בודדים ב-NM וב-NL מקודדים את תדר הצליל הטוב ביותר שלהם - הידוע בשם התדר האופייני שלהם (CF) - לאורך המישור הרוסטרלי-מדיאלי עד לאזור הקאודו-לרוחב. נוירונים רגישים לתדר גבוה נמצאים באזור הרוסטרלי-מדיאלי ונוירונים רגישים לתדר נמוך ממוקמים באופן קאודו-לרוחבי. לפיכך, שיטות דיסקציה מסורתיות של רקמת גזע המוח השמיעתית לחקר תכונות טונוטופיות השתמשו בפרוסות קורונל עוקבות. ואכן, מיקרו-מעגלים שמיעתיים של עוברי עוף מתפתחים הוקמו כמערכת מודל לחקר עיבוד אותות של תפקודים שמיעתיים טונוטופיים באמצעות פרוסות גזע מוח רציפות של מישור קורונלי קואורלי במשך עשרות שנים 1,2,3,4,5,6.
עם זאת, הארגון הטונוטופי של NM ו- NL הוא מפותל מבחינה טופולוגית ומורפולוגית. תשומות עצב השמיעה מופצות כך שכניסות CF גבוהות מסתיימות במבנים דמויי אנדבולב המכסים לפחות רבע מההיקף הסומטי של תא NM אדנדריטי. לעומת זאת, כניסות CF נמוכות אינן מאורגנות עם הדקים דמויי נורה סופית אלא עם מספר סינפסות בוטון על דנדריטים של נוירוני NM. כניסות CF אמצעיות מסתיימות הן כנורת קצה והן כסינפסות דמויות בוטון 4,7,8,9,10,11,12. ב-NL, השיפוע הדנדריטי הסטריאוטיפי ביותר ניכר לא רק באורך הדנדריטי אלא גם ברוחב הדנדריטי. שיפוע דנדריטי ייחודי זה תואם באופן הדוק את ציר הטונוטופ. הדנדריטים עוברים עלייה של פי 11 באורך ועלייה של פי חמישה ברוחב מתאי עצב בעלי CF גבוה לנמוך,בהתאמה 6. כדי להתגבר על התפלגויות מפותלות כאלה של גרעינים אלה בפרוסות קורונליות, פרוטוקול זה מתאר גישות דיסקציה במישור הסגיטלי, האופקי והאופקי/רוחבי. טכניקות חיתוך אלה מספקות דוגמאות לרקמת גזע מוח שמיעתית המציגה תכונות טונוטופיות מרביות במישור פרוסה בודד.
Protocol
כל הנהלים אושרו על ידי הוועדות המוסדיות לטיפול ושימוש בבעלי חיים של אוניברסיטת נורת'ווסטרן (IACUC) ובוצעו בהתאם להנחיות המכונים הלאומיים לבריאות לטיפול ושימוש בחיות מעבדה. הפרוטוקולים לדיסקציה והכנה של רקמת גזע המוח תואמים את הפרוטוקולים הקודמים 5,13.
1. טיפול בביצים
- רכשו ביציות מופרות (Gallus gallus domesticus) מספק בעלי חיים מקומי שאושר על ידי IACUC.
- יש לאחסן את הביצים מיד עם ההגעה במקרר בטמפרטורה של 14 מעלות צלזיוס ולדגירה תוך 5 ימים.
הערה: כדאיות העובר יורדת במידה ניכרת לאחר שבוע. - לעקר את הביצים עם 70% אתנול לפני הדגירה ב 38 ± 1 מעלות צלזיוס ו ~ 50% לחות.
2. הרכב והכנה של נוזל מוחי-שדרה מלאכותי (ACSF)
- יש לערבב את הכימיקלים הבאים ב-1 ליטר של 18.2 MΩcm dH 2 O כדי ליצור תמיסת מלאי ACSF של 10x: NaCl (נתרן כלורי) 130 mM, NaHCO3 (סודיום ביקרבונט) 26 mM, KCl (אשלגן כלורי) 2.5 mM, NaH2PO4 (נתרן דיהידרוגן פוספט) 1.25 mM, דקסטרוז (D-(+)-גלוקוז) 10 mM. שמור את פתרון המלאי במקרר.
- יש להכין את MgCl 2 (מגנזיום כלוריד) 1 M ואת CaCl 2 (סידן כלורי) 1 M תמיסות בנפרד ב-18.2 MΩcm dH2 O ולאחסן במקרר.
- מיד לפני השימוש, יש לדלל 10x ACSF ל-1x ולבעבע ברציפות עם 95% O 2/5% CO 2 למשך 15-20 דקות ולהוסיף MgCl 2 ו-CaCl 2. כדי להכין ACSF ו-dACSF (ניתוח ACSF), התאימו לריכוז הסופי של Mg 2+ 1 mM, Ca 2+ 3 mM ו-Mg 2+ 3 mM, Ca 2+ 1 mM, בהתאמה.
- הגדר את קצב המבעבע עבור ACSF כך שה- pH הוא 7.2-7.4 עם אוסמולליות בין 300 ל- 310 mOsm/L.
הערה: הצבת ה-ACSF באמבט קרח בזמן מבעבע מועילה לשמירה על טמפרטורת תמיסה נמוכה, שתתמוך בשלמות המבנית של הרקמות בזמן הניתוח.
3. אגרוז (5%) הכנת בלוקים
- מערבבים 5 גרם של agarose ב 100 מ"ל של dACSF. יש להשתמש באמבט מים בטמפרטורה של 100 מעלות צלזיוס או במיקרוגל למשך 2-3 דקות, תוך ערבוב כל 30 שניות כדי למנוע גושים עד שהאגרוז מתמוסס לחלוטין ומתחיל לבעבע.
- יוצקים את האגרוז המומס בצלחת פטרי ריקה בעובי של עד 5 מ"מ ושומרים בטמפרטורת החדר להתייצבות. לאחר ההגדרה, אוטמים את צלחת הפטרי באמצעות פרפילם ומאחסנים בטמפרטורה של 4 מעלות צלזיוס.
- חותכים את agarose לתוך בלוקים מעוקב עם להב חד ולהשתמש בהם בזמן דיסקציה.
4. פרוטוקול דיסקציה ובידוד גזע המוח השמיעתי
- נקו את אזור הנתיחה באמצעות תרסיס תמיסת אתיל 70%.
- הדביקו את גוש האגרוז התומך או הזוויתי על מגש הוויברטום.
- בחר ביצים בגיל הרצוי (E20 ו- E21 בפרוטוקול הנוכחי). טפלו ודגרו את הביצים בהתאם לפרוטוקולים המפורטים לעיל כמו בשלב 1.
- אתרו את החלל המלא באוויר על ידי הנחת הביצה תחת אור בהיר (סוכריות) וחפשו את החלל הזה בצד הגדול או העגול יותר של הביצה.
- לאקלם את הביצים לטמפרטורת החדר, לפצח את הקליפה מעל החלל המלא באוויר ולחשוף את שק הממברנה.
- לעשות חתך עדין בשק כדי לחשוף את המקור.
- בעזרת אזמל, משכו בעדינות את הצוואר והראש מתוך הביצה.
- מהיר לערוף את הראש באמצעות מספריים חדים.
- לאחר עריפת הראש, נקו את הראש עם dACSF מקורר קרח כדי להסיר עודפי דם מפד הדיסקציה.
- החזיקו את הראש יציב ב-dACSF מקורר קרח ובצעו חתך רוסטרו-קאודלי. מתחילים את החתך מאחורי ובין העיניים ועוקבים אחר אורך הצוואר שנקטף.
הערה: עוברים צעירים יותר עשויים לדרוש פחות לחץ בעת ביצוע החתך. - להפריד את העור כדי לחשוף את הגולגולת.
- חותכים את הגולגולת מאחורי העין בקו אמצע לכיוון לרוחב. עשו זאת עבור שתי ההמיספרות.
הערה: שלב זה עוזר להפריד את החלק הרוסטרלי של הגולגולת מהמוח המחובר תוך שמירה על רקמת המוח שלמה5. - פורסים את החלק הרוסטרלי של הגולגולת. מניחים את הלהב מאחורי העיניים ומבצעים חיתוך מהיר.
הערה: ייתכן שיידרש מאמץ כדי לחתוך את הגולגולת המצורפת בצורה נקייה. - לטבול את הראש בצלחת של dACSF קר.
- בעזרת זוג מספריים קטן, יש לבצע חתכים בקו האמצע עד לרוחב באזור הקאודאלי של הגולגולת כדי לנסות ולהפריד את המוח מהגולגולת מבלי לגרום נזק לרקמות.
- חשוף בעדינות את גזע המוח ואת המוח הקטן. לסגת את האזור הגבי של הגולגולת כולה, להסיר את גזע המוח בזהירות, ולחשוף אותו בעזרת מכחול עדין עם מזחלת קלה. השתמש במלקחיים מעוקלים כדי לנקות את גזע המוח מחיבור רקמות וכלי דם. שימו לב במיוחד לאזור העצב הגולגולתיה-8 והקפידו להשאיר אורך קצר של סיבי עצב שלמים משני הצדדים.
- להפריד את גזע המוח מהמוח הקטן על ידי חיתוך peduncles והסרת כלי הדם בזהירות. לקצץ את גזע המוח של כלי דם נוספים.
הערה: ודא שההליך כולו מבוצע ב-dACSF מקורר קרח המבעבע ברציפות בקרבוקסיגן (95% O 2/5% CO2).
5. חיתוך ויברטום
הערה: בשלבים הבאים, יש לתמוך בחלק האחורי של הרקמה עם חתיכת אגרוז מעוקבת.
- הניחו את להב הוויברטום לאורך הציר האופקי והדביקו את גזע המוח על מגש חיתוך. הדביקו את הצד הרוסטרלי, תוך שמירה על הציר הרוסטרלי-קאודלי אנכי לפרוסות קורונל.
- שמור על הציר הצדדי-מדיאלי אנכי עבור פרוסות סגיטליות.
- הדביקו את הצד הגחוני, ושמרו על הציר הגבי-גחוני אנכי לפרוסות אופקיות.
- כדי להשיג את המישור הסאגיטלי-אופקי הזוויתי החריף, הדביקו את הצד הגחוני של גזע המוח, תוך שמירה על הציר הגחוני-גבי אנכי על משטח ההיפוטנוז של גוש האגרוז, שנחתך בזווית של 45°. הדביקו את המשטח הנגדי של גוש האגרוז הפונה למגש החיתוך ושמרו על הציר הרוסטרלי-קאודלי במקביל לקצה הלהב.
6. טיפול בחתיכות שבריריות או גדולות של רקמת גזע המוח
- בגישה חלופית לשלב 5, טבלו את גזע המוח המבודד בנקודת התכה נמוכה (LMP) בטמפרטורה של ~40 מעלות צלזיוס בצלחת פטרי בגודל 35 מ"מ x 10 מ"מ.
- לאחר ששפכו אגרוז על גזע המוח השקוע, הניחו את צלחת הפטרי על קרח כדי להתמצק. חותכים את גוש האגרוז המעוקב עם גזע מוח מוטבע באמצעות סכין גילוח חדה.
- הדביקו את גוש האגרוז LMP על הצד הרוסטרלי שלו, תוך שמירה על הציר הרוסטרלי-קאודלי של גזע המוח אנכי.
- קח פרוסות קורונל עד שניתן יהיה לדמיין את אזור NM.
- הסר את בלוק agarose מן הדבק עם להב חד. כדי לזהות את הגרעינים, הניחו בעדינות 0.5 μL של צבע (טולוידין כחול או כתום G) על NM עם מחט עדינה.
- הרכיבו מחדש בלוק זה על מגש החיתוך לפרוסות סגיטליות או אופקיות וזהו את הגרעינים ביחס לאזור המוכתם.
- לקבלת הביצועים הטובים ביותר, הגדר את מהירות חיתוך הוויברטום ל- 4 - 5 (~ 30 ± 4 מ"מ לדקה), תדר רטט ב- 85-87 הרץ וחותך משרעת ב- 4-6 (~ 1 ± 0.2 מ"מ).
- לאחר חיתוך גזע המוח, הניחו את הפרוסות שנאספו ברצף של 200-300 מיקרומטר בתא פרוסות זמין מסחרית כדי שיווי משקל למשך שעה אחת בטמפרטורת החדר ב-ACSF, עם בועות רציפות בתערובת של 95% O 2/5% CO 2 (pH 7.2-7.4, אוסמולריות 300-310 mOsm/L). בתנאים אלה, פרוסות להישאר קיימא עד 5-6 שעות.
7. אלקטרופיזיולוגיה: הליך מהדק טלאי
- העבר פרוסת גזע מוח לתא ההקלטה עם זלוף מתמשך של ACSF קרבוקסיגנציה ~ 1.5 ± 0.5 מ"ל לדקה.
- משוך פיפטות טלאי עם מושך מיקרופיפט בקוטר קצה 1-2 מיקרומטר והתנגדות בטווח של 3-6 MΩ.
- מלאו פיפטות בתמיסה פנימית מבוססת K-גלוקונט (להקלטת מהדק זרם).
- כדי לבחון את התכונות העצביות באזורים הטונוטופיים השונים בתוך פרוסה, אתר תאי עצב בשני קצות מישור הפרוסה והתקרב באמצעות אלקטרודת ההקלטה.
- שמרו על לחץ אוויר חיובי בקצה הפיפטה תוך כדי התקרבות לנוירון.
- לנוע לכיוון סומה עד כניסה הוא הדמיה על הנוירון. בצע את שני השלבים הבאים במהירות.
- צרו אטימה של ג'יגה-אוהם (1 GΩ) על ידי שחרור לחץ אוויר חיובי.
- שמור על הגדרת המגבר במצב מהדק מתח ותקן את היסט הפיפטה כאפס pA. הפעל בדיקת איטום (דופק בדיקה של 10 mV ב-100 הרץ). החל לחץ אוויר שלילי כדי לקרוע חלק קטן של הממברנה העצבית.
- כדי לבחון את המאפיינים הפנימיים הפעילים של נוירונים שמיעתיים, החל זריקות זרם סומטי היפרפולריזציה ודה-פולריזציה.
הערה: ניתן להציג דוגמאות להליך זה בווידאו משלים S1, S2. פרטים על הליך זה מסופקים באגדות הווידאו.
תוצאות
כל פרוסות גזע המוח המוצגות כאן נרכשו מרקמת גזע המוח (~200-300 מיקרומטר) ודוממו באמצעות אופטיקה של 5x אובייקטיבית וניגודיות הפרעה דיפרנציאלית (DIC). המצלמה הורכבה על מיקרוסקופ החיתוך וחוברה למחשב באמצעות תוכנה לרכישת תמונות (ראו טבלת חומרים). כניסת הלוויין לנתונים אלה (לוחות ימניים) צולמה באמצעות מטרת הגדלה של פי 60 של טבילת מים. הושקעה תשומת לב כדי להבטיח שכל האזורים בפרוסת גזע המוח יוגדלו באופן שווה בעת קבלת תמונות דיגיטליות. התמונות צולמו בבהירות ומיקוד אופטימליים. התמונות הדיגיטליות של פרוסות גזע המוח נתפרו באופן מישורי המבוסס על אזור חופף ויובאו למחשב שולחני לצורך התאמות נוספות של בהירות, ניגודיות וגווני אפור. המיקרו-מעגלים הבסיסיים של גזע המוח השמיעתי של התרנגולת זוהו על פי עבודה קודמת 1,2,5,13. תחת המיקרוסקופ (5x מטרה), גרעינים שמיעתיים זוהו כאזור הסמוך לסיבי העצב המיאלינים בכבדות המסתובבים סביב כל גרעין הן באופן איפסילטרלי והן באופן קונטרלטרלי לאורך האזורים הגביים של הפרוסה.
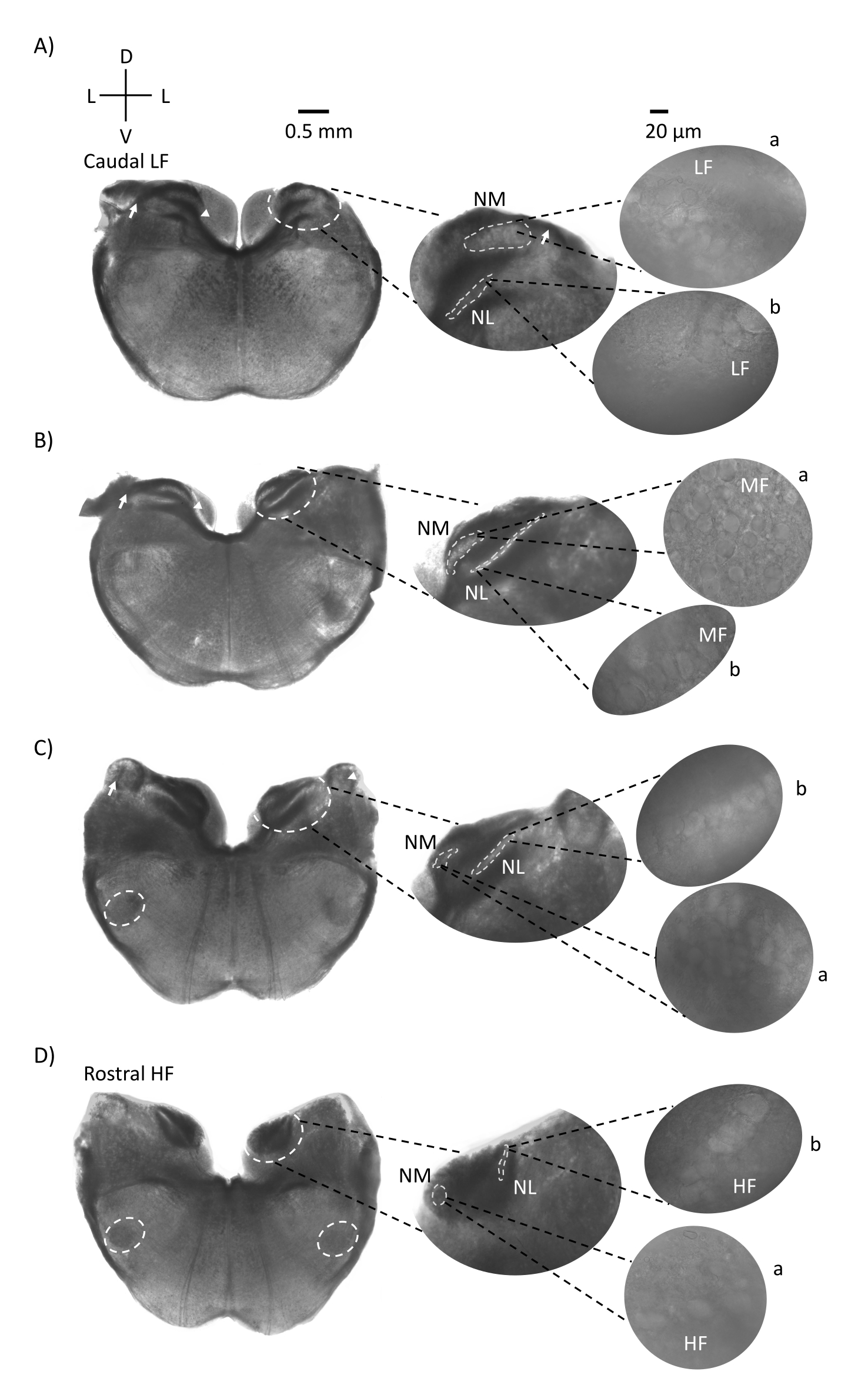
איור 1 מראה את המקטעים הקורונליים המסורתיים של רקמת גזע המוח (200-300 מיקרומטר) מעובר עוף E21. ארבע פרוסות הכתר המוצגות כאן מייצגות את אזורי התדר היחסי של גרעיני גזע המוח השמיעתיים, מהאזור השמיעתי בעל ה-CF הנמוך ביותר (איור 1A, caudo-lateral) המתקדם לאזור השמיעתי בעל ה-CF הגבוה ביותר (איור 1D, רוסטרלי-מדיאלי). עבור כל ארבע פרוסות הכתר באיור 1A-D, אזורים מוגדלים של NM ו-NL מסומנים מוצגים בעמודה האמצעית ומוגדלים (מטרה פי 60) על המראה הימני של לוח האיור (a ו-b, בהתאמה, בכניסות לוויין). החץ באיור 1A,B מראה את הקלט של סיבי עצב השמיעה, וראש החץ מראה ביפורקציה של אקסונים NM בצד שמאל של הפרוסה. איור 1C מראה מבנה נוסף של גרעין שבלול העופות המכונה גרעין זוויתי (NA, חץ משמאל וראש חץ מימין). שתי פרוסות הכתר הרוסטרליות ביותר מראות את גרעין האוליברי העליון (SON) הממוקם לאורך האזור הגחוני-צדדי של הפרוסה הקורונלית (איור 1C,D, עיגולים מקווקווים לבנים).
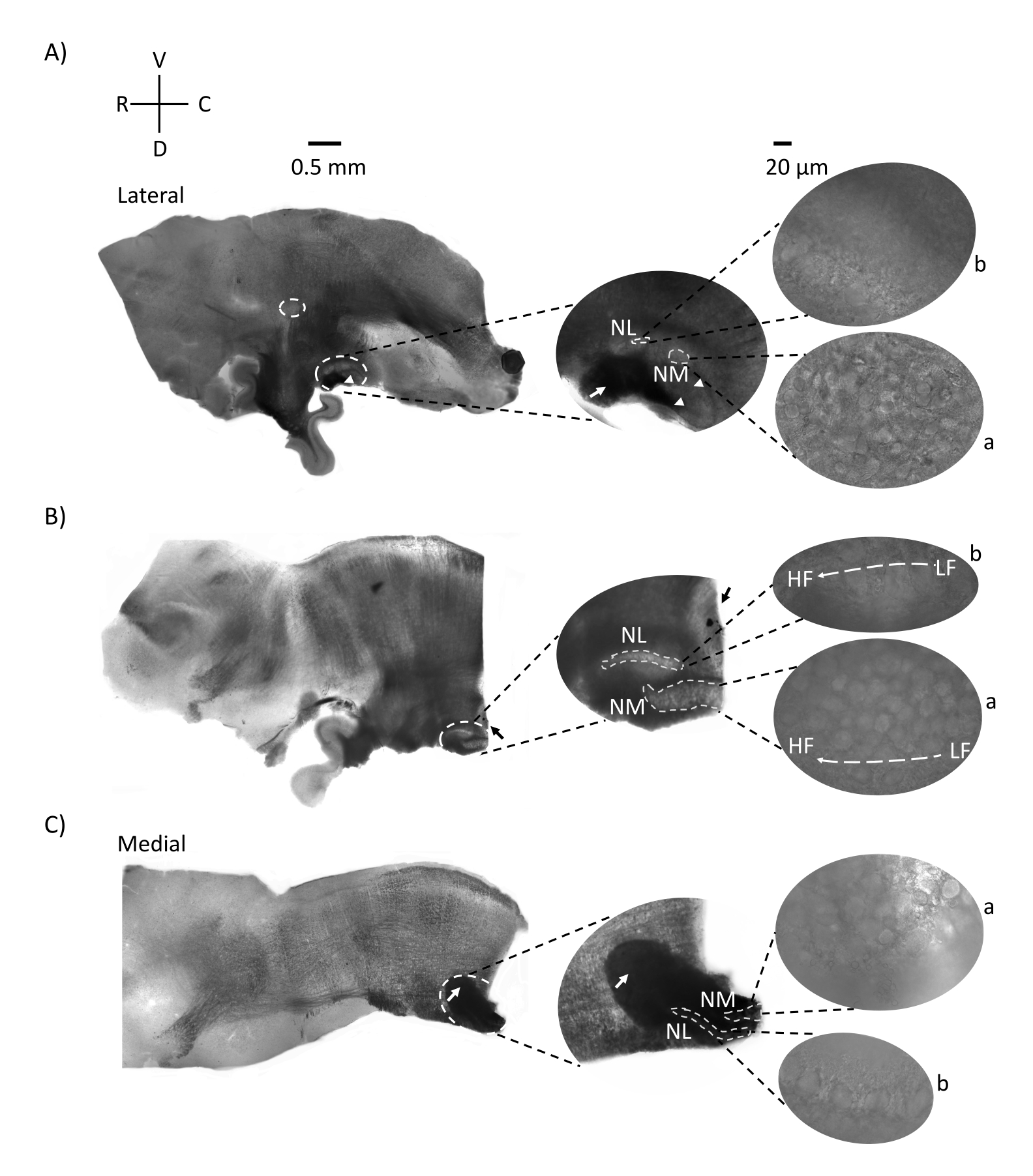
איור 2 מראה מקטעים סגיטליים של רקמת גזע המוח (200-300 מיקרומטר) מעובר עוף E21. עבור כל שלוש הפרוסות הסגיטליות (איור 2A-C), אזורים מוגדלים של NM ו-NL מסומנים מוצגים בעמודה האמצעית ומוגדלים (60x אובייקטיביים) במראה הימני של לוח האיור (a ו-b, בהתאמה בתצלומי לוויין). NM ו-NL זוהו במקום שבו סיבי עצב השמיעה (איור 2A, חץ אמצעי) נכנסו לאשכול הנוירונים שנצפו בהגדלה גבוהה יותר (איור 2A, עיגולים מקווקווים וראשי חץ באמצע, קטנים, לבנים) והדגישו את נקודת ההתחלה של אזור השמיעה (איור 2A, שמאלי, גדול, עיגול מקווקו לבן וראש חץ). SON זוהה באזור הרוסטרו-לטרלי של הפרוסה הצידית ביותר (איור 2A, עיגול קטן, לבן ומקווקו). איור 2B מראה אזורים טונוטופיים מורחבים המכילים אזורים שמיעתיים נמוכים יחסית וגבוהים של CF מ-NM ו-NL לאורך הציר הרוסטרלי-קאודלי (אזורים בעלי קווי מתאר לבנים, ראו גם כניסת לוויין). איור 2C מראה את הניבים האקסונליים האיפסילטרליים והקונטרלטרליים בפרוסה המדיאלית ביותר ובנקודת הקצה של אזור השמיעה (חצים שמאלה ואמצעיים). כיוון הפרוסות המוצג כאן מנוגד לכיוון המסורתי של פרוסות כפי שניתן לראות באיור 1 (כלומר, קורונל). זה בוצע כדי להציג את הכיוון המתאים ביותר לגישה של פיפטה זכוכית הנדרשת להקלטות אלקטרופיזיולוגיות.
כדי לאשר שאזור גדול של ציר הטונוטופיק היה מיוצג באיור 2B, בוצעו הקלטות אלקטרופיזיולוגיות של מהדק זרם מנוירוני NM. איור 3 מראה דמיון תפקודי והבדלים של תאי עצב בוגרים (E21) NM שנרשמו מפרוסה קורונלית (איור 3A,B) ופרוסה סגיטלית (איור 3C,D, וידאו משלים S1, S2). שני נוירוני NM נבחרו מהקצוות המדיאליים והלטרליים של פרוסה קורונלית (בדומה לפרוסה שמוצגת באיור 1B), ושני נוירוני NM נבחרו מהקצוות הרוסטרליים והקאודליים של NM בפרוסה סגיטלית (כמו בפרוסה שמוצגת באיור 2B). איור 3A,B מראה תכונות תגובה אלקטרופיזיולוגיות דומות לזריקות זרם סומטי (−100 pA עד +200 pA, +10 דרגות pA, משך 100 אלפיות השנייה). תבנית הירי של שני נוירוני NM אלה מציגה הבדלים עדינים במישור פרוסה זה, מה שמעיד על למינה בתדר ISO יחסית עבור נוירוני NM בתדר בינוני. איור 3C,D מראה שלדפוסי הירי יש הבדלים מהותיים על פני הציר הרוסטרלי-קאודלי, מה שמצביע על שיפוע טונוטופי גבוה יחסית מנוירון NM בתדר נמוך (איור 3C) לנוירון NM בתדר גבוה (איור 3D). שני תאי העצב הציגו את דפוסי הירי הסטריאוטיפיים שלהם כפי שדווחו קודםלכן 14,15.
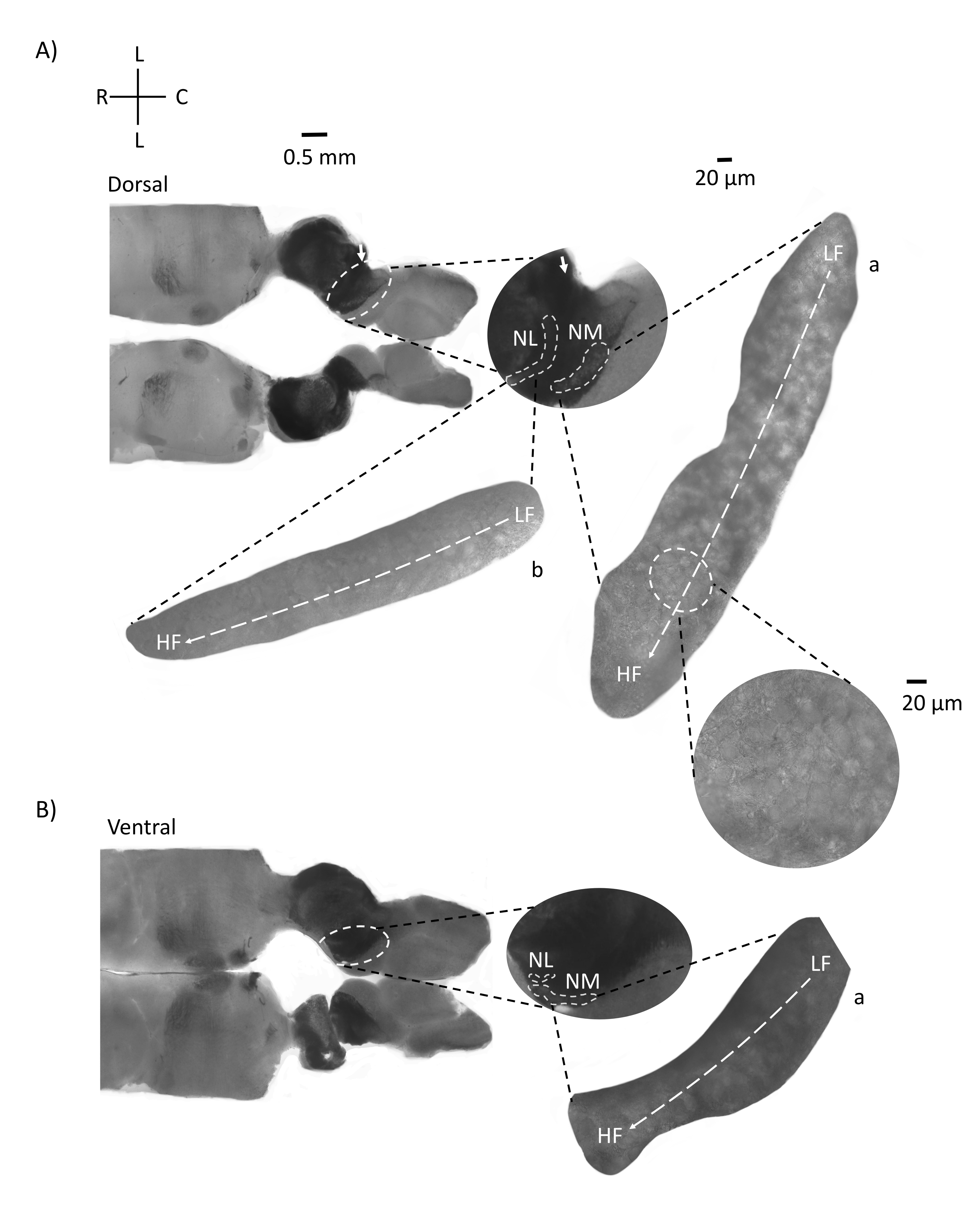
איור 4 מראה מקטעים אופקיים של רקמת גזע המוח (200-300 מיקרומטר) של עובר עוף E21. עבור שתי הפרוסות האופקיות (איור 4A,B), אזורים מוגדלים של NM ו-NL מסומנים מוצגים בעמודה האמצעית ומוגדלים (60x אובייקטיביים) במראה הימני של לוח האיור (a ו-b, בהתאמה, בכניסות לוויין). בפרוסות האופקיות, NM ו-NL זוהו לכיוון קו האמצע, ותאי עצב התפשטו לאורך הציר הלטרלי-מדיאלי (איור 4A,B, אזורי מתאר אמצעיים, לבנים ומקווקווים). התמונות המוגדלות מראות את ההיקף הגדול של שיפוע הטונוטופי. נוירונים בתדר נמוך נמצאים באזורים הקאודו-צדדיים, ותאי עצב בתדר גבוה נמצאים באזורים הרוסטרליים-מדיאליים (איור 4A,B, מימין, לוויינים). הסיבים העוברים דרך קו האמצע לאורך הציר הרוסטרלי-קאודלי מראים את הקשרים הקונטרלטרליים של גרעיני השמיעה, אך הארגון של סיבים אלה אינו במישור פשוט. עם זאת, פרוסות זוויתיות חריפות מחתך אופקי/רוחבי יכולות לעקוב אחר הסיבים האקסונליים הללו לכיוון המישור הסגיטלי. פרוסות של רקמת גזע מוח בעובי 200-300 מיקרומטר בזווית חדה (45°) ממישור אופקי מוצגות באיור 5. ניתן לראות גרעינים שמיעתיים בגזע המוח על פני התפשטות אלכסונית גדולה שמתחילה מהפרוסה הצדדית ביותר ומסתיימת בפרוסה המדיאלית ביותר (איור 5A-C, המסומן בלוחות אמצעיים, אזור קו מתאר לבן). יתר על כן, ניתן להמחיש את הכיוון הזוויתי של אזורי NM ו-NL גם בפרוסות אסימטריות עוקבות (איור 5A-C, לוחות אמצעיים מסומנים, אזור לבן עם קו מתאר מקווקו). תמונות מוגדלות (אובייקטיביות פי 60) מראות את הציר הטונוטופי של גרעיני השמיעה כשהוא נע לאורך הציר הרוסטרלי-מדיאלי עד הקאודו-לרוחבי (איור 5A-C, מימין, כניסת לוויין). הכיוון של פרוסות באיור 5 דומה לזה שבאיור 2. הם מנוגדים להצגה המסורתית של תמונות אך מתאימים יותר לניסויים אלקטרופיזיולוגיים.

איור 1: מקטעים טוריים קורונליים מייצגים של גזע המוח. (א-ד) משמאל: פרוסות מציר קאודלי לציר רוסטרלי, גרעיני השמיעה וסיבים מחברים המסומנים בעיגול מקווקו לבן. התוספת האמצעית היא תצוגה גדולה יותר של אזור השמיעה, שבו גרעינים מוצגים בתוך עיגולים מקווקווים לבנים a: NM ו- b: NL. החצים מראים סיבים אפרנטיים של עצב השמיעה, וראש החץ מראה ביפורקציה של אקסון NM ב-A,B. החץ מציג את NA ב-C. עיגול מקווקו לבן לרוחב מראה SON ב- C,D. מימין: תוספת לוויין מראה את הגרעינים האלה ב-60x מטרה: a: NM ו-b: NL. קיצורים: NM = גרעין מגנוצלולריס; NL = גרעין למינריס; NA = גרעין זוויתי; SON = גרעין אוליברי מעולה; LF = נוירונים בתדר נמוך יחסית; MF = נוירונים בתדר בינוני; HF = נוירונים בתדר גבוה; D = גב; L = לרוחב; V = גחון. אנא לחץ כאן כדי להציג גרסה גדולה יותר של נתון זה.
{kind=link}

איור 2: מקטעים סדרתיים סגיטליים מייצגים של גזע המוח. (A-C) משמאל: פרוסות מהציר הלטרלי לציר המדיאלי כאשר גרעיני השמיעה מסומנים בעיגול מקווקו לבן. התוספת האמצעית מציגה את אותו אזור גרעיני שמיעה במבט גדול יותר, המסומן בתוך עיגולים מקווקווים לבנים. (A) עיגול מקווקו לבן במרכז הפרוסה מדגיש את ה-SON; חץ המציג סיבי עצב שמיעתי וראש חץ המציג NA. כתם שחור כהה בקצה הימני של פרוסה הוא חפץ הדמיה. ניתן לראות אזורים במוח הקטן הגב לאזור השמיעה בשתי הפרוסות A ו- B בלוח השמאלי. (B) פרוסה סגיטלית שהכיוון שלה השתנה למישור הקורונלי (במהלך החיתוך). אזור השמיעה זוהה בצבע כחול (חץ שחור) ושוב נחתך במישור הסגיטלי. (א-ג) הוסף באמצע אזור NM ו- NL המסומנים תחת קווים לבנים מקווקווים. מימין: תצוגת לוויין מראה a: NM ו- b: NL שנצפתה בהגדלה אובייקטיבית של פי 60. שיפוע טונוטופי LF ו-HF בגרעינים שמיעתיים מוצג לאורך ציר רוסטרו-קאודלי. חצים המצביעים על האזור הכהה ב-(C) מראים סיבי NM מיאלינים כבדים העוברים לאורך קו האמצע דרך הציר המדיאלי. הסיבים מחברים משני צדי גרעיני השמיעה. קיצורים: NM = גרעין מגנוצלולריס; NL = גרעין למינריס; NA = גרעין זוויתי; SON = גרעין אוליברי מעולה; LF = נוירונים בתדר נמוך יחסית; HF = נוירונים בתדר גבוה; D = גב; V = גחון; R = רוסטרל; C = קאודלי. אנא לחץ כאן כדי להציג גרסה גדולה יותר של נתון זה.
{kind=link}

איור 3: הקלטות אלקטרופיזיולוגיות של תגובה עצבית לזריקות זרם סומטי (−100 pA עד +200 pA, +10 דרגות pA, משך 100 אלפיות השנייה) במצב מהדק נוכחי. תאי עצב נבחרו להקלטות באותה פרוסה אך באזורים מנוגדים קיצוניים של NM. (א,ב) תגובות עצביות מייצגות בפרוסה קורונלית אחת המציינות תכונות יחסיות בתדר ISO עם הבדלים עדינים. תכונות התגובה מייצגות שני נוירוני MF שונים שנרשמו מהאזורים המדיאליים (A) והלטרליים (B) ביותר של NM בפרוסה קורונלית. (ג,ד) הקלטות עצביות מייצגות מפרוסה סגיטלית אחת. ההקלטות מראות תגובת LF NM (C) ותגובת HF NM (D), המדגישות את ההבדלים המהותיים בשיפוע הטונוטופי לאורך מקטע סגיטלי יחיד. קיצורים: NM = גרעין מגנוצלולריס; LF = נוירונים בתדר נמוך יחסית; MF = נוירונים בתדר בינוני; HF = נוירונים בתדר גבוה. אנא לחץ כאן כדי להציג גרסה גדולה יותר של נתון זה.
{kind=link}

איור 4: מקטעים טוריים אופקיים מייצגים של גזע המוח. (א,ב) משמאל: פרוסות לאורך הציר הגבי עד הגחוני, גרעינים שמיעתיים מסומנים בעיגולים מקווקווים לבנים. סיבי עצב הגולגולתה-8 מחברים גרעינים שמיעתיים המסומנים בחץ. התוספת האמצעית היא תצוגה גדולה יותר של אזור גרעיני השמיעה עם גרעינים שמיעתיים המסומנים תחת קווים מקווקווים לבנים NM ואזורי NL מוצגים. תנועה טופולוגית ברורה של גרעיני שמיעה ניתן לראות ב-A,B. (א,ב) מימין: תצוגת לוויין גדולה המציגה a: NM ו - b: NL. התוספת הימנית מראה גרעינים שמיעתיים שנצפו בהגדלה אובייקטיבית של פי 60 ואת הציר הטופולוגי המעוקל מ-LF ל-HF לאורך ציר קאודו-לרוחב לציר רוסטרלי-מדיאלי. קיצורים: NM = גרעין מגנוצלולריס; NL = גרעין למינריס; LF = נוירונים בתדר נמוך יחסית; HF = נוירונים בתדר גבוה; L = לרוחב; R = רוסטרל; C = קאודלי. אנא לחץ כאן כדי להציג גרסה גדולה יותר של נתון זה.
{kind=link}

איור 5: מקטעים טוריים אופקיים/רוחביים אקוטיים מייצגים (45°). (A-C) משמאל: מקטעים טוריים של גזע המוח, גרעינים שמיעתיים המסומנים בעיגול מקווקו לבן. התוספת האמצעית היא תצוגה גדולה יותר של אזור השמיעה. (A) התוספת האמצעית מראה את ההתפשטות הגדולה ביותר של נוירוני NM ו-NL בפרוסות אלה. (ב,ג) תוספת אמצעית: גרעינים שמיעתיים המסומנים בקווים מקווקווים לבנים מראים שינוי טופולוגי הדרגתי בהשוואה ל- (A-C). מימין: תוספת לוויין המציגה גרעינים שמיעתיים a: NM ו- b: NL בהגדלה אובייקטיבית של פי 60. הציר הטונוטופי מאזורי LF לאזורי HF ב-NM וב-NL מסתובב באופן זוויתי מפרוסות רוחביות למדיאליות. קיצורים: NM = גרעין מגנוצלולריס; NL = גרעין למינריס; LF = נוירונים בתדר נמוך יחסית; HF = נוירונים בתדר גבוה; V = גחון; R = רוסטרל; D = גב; C = קאודלי. אנא לחץ כאן כדי להציג גרסה גדולה יותר של נתון זה.
{kind=link}
וידאו משלים S1: זריקות זרם סומטי היפרפולריזציה ודה-פולריזציה. תכונות תגובה מנוירון בתדר נמוך ובתדר גבוה להזרקות זרם סומטי של 100 אלפיות השנייה במצב מהדק זרם. תאי עצב נבחרו מתוך אותה פרוסת גזע מוח סגיטלית. ההזרקות נעות בין -100 ל +200 pA בשלבים של +10 pA במרווחים, משך זמן של 100 אלפיות השנייה. פוטנציאל הפעולה נראה בתגובה לצעדים הנוכחיים המספקים של דה-פולריזציה. הסרטון תואם את העקבות הסופיים המוצגים באיור 3C. אנא לחץ כאן כדי להוריד קובץ זה.
וידאו משלים S2: זריקות זרם סומטי היפרפולריזציה ודה-פולריזציה. בדומה לווידאו משלים S1, סרטון זה מציג תכונות תגובה מנוירון בתדר נמוך ותדר גבוה להזרקות זרם סומטי של 100 אלפיות השנייה במצב מהדק נוכחי. תאי עצב נבחרו מתוך אותה פרוסת גזע מוח סגיטלית. ההזרקות נעות בין -100 ל +200 pA בשלבים של +10 pA במרווחים, משך זמן של 100 אלפיות השנייה. פוטנציאל הפעולה נראה בתגובה לצעדים הנוכחיים המספקים של דה-פולריזציה. הסרטון תואם את העקבות הסופיים המוצגים באיור 3D. אנא לחץ כאן כדי להוריד קובץ זה.
Discussion
מקטעים קורונליים של רקמת גזע המוח העוברי של עוף אפשרו לחקור למינה בתדירות iso אינדיבידואלית יחסית במשך עשרות שנים 1,2,5. עם זאת, הארגון הטונוטופי (כלומר, התדירות) של גזע המוח השמיעתי של התרנגולת מפותל מבחינה טופולוגית ועשוי להיות נגיש יותר בצירים אנטומיים אחרים בהתאם לשאלת המחקר הספציפית. אף על פי שמספיק לחקור שאלות אנטומיות ופיזיולוגיות הנוגעות לאזורים בודדים בתדר איזו, חקר הווריאציות הטונוטופיות והתפתחותן על פני אזורים גדולים יותר בגזע המוח השמיעתי מוגבלים במידה מסוימת על ידי חתכים קורונליים. כדי להתגבר על מגבלה זו, פרוטוקול זה מתאר גישות במישור הסגיטאלי, האופקי והאופקי/רוחבי כדי לספק דוגמאות נוספות לרקמת גזע המוח השמיעתית המציגה תכונות טונוטופיות ושיפועים מרביים במקטע גזע מוח בודד.
מקטעים סגיטליים של אזורי גזע המוח השמיעתיים מראים כי אזורים טונוטופיים שונים מפוזרים על פני אזור גדול יותר בתוך הפרוסה בהשוואה למקטעים קורונליים (אזור שמיעתי סגיטלי = ~300-600 מיקרומטר, אזור שמיעתי קורונלי = ~200-350 מיקרומטר). לדוגמה, אזורי NM ו-NL הודגמו על פני שטח גדול יותר לאורך הציר הרוסטרו-קאודלי במקטעים סגיטליים (לדוגמה, איור 2B), והשיפוע הטונוטופי הפונקציונלי שעובר לאורך הציר האנטומי הזה נכלל במידה רבה בתוך פרוסה סגיטלית אחת. זה אושר עוד יותר עם הקלטות של מהדק זרם של הבדלים עצביים פנימיים המשתנים לאורך השיפוע הרוסטרלי-קאודלי כפי שדווח קודם לכן14,15 (למשל , איור 3C,D). ניסויים עתידיים המדגישים תכונות אנטומיות ואימונוהיסטוכימיות לאורך ציר הטונוטופיק יוכלו להמשיך ולחקור שיפועים ידועים של תכונות שמיעתיות במישור פרוסה סגיטלית אחת. אלה כוללים, אך אינם מוגבלים, לדפוסי צביעת MAP2 וביטוי תעלות אשלגן, שהם שיפועים ידועים של ארכיטקטורה דנדריטית ותכונות פנימיות של NM ו- NL שהוצגו בעבר בסעיפים קורונליםעוקבים 16.
מקטעים אופקיים של אזורים שמיעתיים בגזע המוח מראים שה-NM וה-NL ממוקמים לכיוון קו האמצע. חלק מהסיבים האקסונאליים השמיעתיים עוברים באלכסון או בניצב למישור האופקי (איור 4). סיבים אלה יכולים להיות מלווים על ידי יצירת פרוסה זוויתית חריפה 45° למישור sagittal. הפרוסות האופקיות/רוחביות שהתקבלו היו גדולות יותר מהפרוסות הסגיטליות או האופקיות, וסיבים אקסונאליים ארוכים עברו דרך הציר הרוסטרו-קאודלי הן עבור הצדדים האיפסילטרליים והן עבור הצדדים הקונטרלטרליים. גם NM וגם NL ניתנים להדמיה באזור אלכסוני גדול יותר (~ 400-700 מיקרומטר) כך שניתן לדמיין קשרים קונטרלטרליים לאורך ציר רוחבי-מדיאלי. בנוסף, מישור הפרוסה האופקית/רוחבית מראה גם כיצד אזורי השמיעה והשיפוע הטונוטופי שנוצר כתוצאה מכך מבצעים סיבוב זוויתי (איור 5). חשיפה זוויתית של קשרים קונטרלטרליים באזור גדול יותר הופכת את הפרוסות הללו למתאימות יותר לגירוי אלקטרופיזיולוגי ולמחקרי מיקרו-מעגלים מאשר פרוסות קורונליות מסורתיות.
יתרונות נוספים
היווצרותם של מיקרו-מעגלים שמיעתיים דורשת תיאום מרחבי-טמפורלי של רמזים המקדמים הישרדות עצבית, סינפטוגנזה, התמיינות אקסונאלית, ארכיטקטורה דנדריטית והתבגרות. לפיכך, ניתן להשתמש בחלקי גזע מוח חלופיים של המיקרו-מעגל השמיעתי של עובר העוף לנושאי המחקר הבאים: ארגון מורפולוגי של נוירונים בממדים שונים מבחינה טופוגרפית; ארגון ומיפוי החיבורים של כל גרעיני השמיעה ושיווי המשקל; זיהוי ואפיון דפוסי הפעילות של מרכיבי מעגלים במישורים איזו-תדרים וטונוטופיים; הארגון הטופוגרפי של מיקרו-מעגלים מעוררים לעומת מעכבים ויחסים לאוכלוסיות נוירונים מיוחדות (גרעינים); מיקום מרחבי של גרעינים שמיעתיים נוירונים וחיזוי CF17; מיקוד שיטתי של סוגים עצביים טונוטופיים ספציפיים; מעקב אחר תאי אב והתפתחותם לגרעינים משומרים; שושלת גנטית של תאים לאבולוציה של מעגלים עצביים18; אנטומיה השוואתית של גזע המוח בין מינים; חקירה של מעגלים שיווי משקל כמו קומפלקס שיווי המשקל של דייטר (DC)19; וסינכרוניזציה ושיחות צולבות בין גרעיני שיווי המשקל.
גישה רבת פנים המשתמשת במישורי פרוסה שונים עשויה לעזור לענות על שאלות בסיסיות לגבי תכונות אנטומיות וביופיזיות לא ידועות של מיקרו-מעגלים בגזע המוח. דוגמה טובה לכך היא הקשר בין גרעיני השמיעה העיקריים (NM, NA, NL ו-SON) לבין גרעיני שיווי המשקל, כולל הגרעין הגבי של הלמניסקוס הלטרלי (LLDp), הגרעין הסמי-לונארי (SLu)20 והגרעין המשיק (TN)3. עם זאת, לפרוטוקול זה ולמחקרים מבוססי פרוסות אלה יש כמה מגבלות.
אמצעי זהירות ומגבלות
בהתאם למוסד המבצע את הניסויים, הנחיות אתיות והטיפול בעוברי עוף עשויים להיות שונים. בעוד שהנחיות המכונים הלאומיים לבריאות לטיפול ושימוש בחיות מעבדה מתירות עריפה מהירה, ישנן שיטות חלופיות להמתת חסד של עוברי עוף21. רקמת גזע המוח של עובר עוף המתפתח מוקדם היא רכה ועדינה בהשוואה לעוברים מבוגרים יותר. יש לו כמה קשרים וכלי דם על פני השטח הזקוקים לזהירות יתרה בעת הסרתם. הרקמה צריכה להישמר ב-dACSF קר כקרח ולחמצן אותה עם 95% O 2/5% CO2 כדי להגדיל את הכדאיות.
שיטת החיתוך הסגיטלית שימושית רק עבור טונוטופיה איפסילטרלית. שיטת חיתוך זו מספקת פרוסות גדולות יותר מאשר פרוסות קורונליות, שהטיפול בהן עלול להיות מסוכן. עם זאת, ניתן לחתוך את הפרוסות בשיטות מחט צולבות המתוארות בפירוט במקום אחר22. שימוש בגזע מוח משובץ של 4% LMP agarose יכול להציל מבנים עדינים בפרוסות, אך יש להיזהר שלא לשפוך אגרוז חם מדי. הגדרתו במהירות על ידי הצבת גזע המוח החסום באגרוז בסביבה מצוננת למשך ~ 1 דקות הופכת את הפרוסות לישימות יותר להקלטות אלקטרופיזיולוגיות.
יישום של superglue בכמויות עודפות יכול להיות רעיל. זה חייב להיות מיושם באופן מינימלי, כמויות עודפות יש לשטוף מיד על ידי שינוי dACSF. עבור פרוסות זוויתיות חריפות (45°), חיתוך הזווית של גוש האגרוז הוא קריטי; ניתן להשתמש במראה כדי לראות את הזווית הקדמית תוך כדי חיתוך גוש האגרוז עם להב חד. להבים מסחריים עשויים להיות בעלי ציפוי שעווה שיש לנגב עם אלכוהול ולייבש לפני השימוש. אופטימיזציה נדרשת עבור מהירות ותדירות חיתוך הוויברטום מכיוון שצינורות סיבים אקסונאליים קשים יותר מרקמת קליפת המוח או מטריקס. שמירה על משרעת גבוהה ושימוש בתמיסת דיסקציה מקוררת עשויים למנוע נזק לרקמות.
כל הפתרונות צריכים להיות מוכנים טריים, ו Ca 2 + ו Mg 2+ יש להוסיף ACSF לאחר מבעבע 95% O 2/5% CO 2 . אחרת, ייתכנו משקעים של Ca2+. יש להשתמש במכחול כדי לטפל בפרוסות בעדינות בתוך הוויברטום. שמור על זמן החיתוך הכולל מתחת ל-15 דקות במידת האפשר. ניתן להשתמש בפיפטת פסטר מזכוכית כדי לתמרן פרוסות גזע המוח.
אין להשתמש בחומרי ניקוי או חומרי כביסה קורוזיביים עבור כלי זכוכית וציוד הבאים במגע עם הפרוסות המשמשות באלקטרופיזיולוגיה. התמונות שצולמו מייצגות את המראה של רקמה בעובי 200-300 מיקרומטר תחת אופטיקה של ניגודיות הפרעה דיפרנציאלית (DIC). איכות הראייה תהיה ירודה יותר מאימונוהיסטוכימיה או ממיקרוסקופיית אלקטרונים, אך היא משקפת במדויק את מה שהנסיין יראה בעת ביצוע הקלטות אלקטרופיזיולוגיות.
מחקרים הנוגעים להתפתחות מוקדמת של מיקרו-מעגלים לאורך ציר אנטומי חלופי, בין אם הם גב-גחוני, רוסטרלי-קאודלי או איפסילטרלי-קונטרלטרלי, מוגבלים בגזע המוח השמיעתי של התרנגולת. אחת הסיבות לכך היא שתפקידם של קודי שעתוק וויסות ההתפתחות הטונוטופית בגזע המוח עדיין אינו מובן במלואו. תופעות פונקציונליות כגון אפנון מלמעלה למטה ופעילות ספונטנית אובדות לעתים קרובות כאשר מתבוננים בפעילות במבחנה. עם זאת, מחקר in vivo משלים הקלטות ספציפיות וישירות של נוירונים בודדים האפשריים רק בתנאי פרוסה אלה. העידון של קבלת רקמת גזע המוח בכיוונים שונים יכול לספק מידע תובנה על ההתפתחות והמורכבות של שיפועים טונוטופיים במיקרו-מעגלים של גזע המוח השמיעתי.
Disclosures
כל המחברים מצהירים כי המחקר נערך ללא כל אינטרס מסחרי או כספי וכי אין להם כל ניגוד עניינים.
Acknowledgements
עבודה זו נתמכת על ידי מענק NIH/NIDCD R01 DC017167. אנו מודים לקריסטין מקללן על שסיפקה הערות עריכה על גרסה מוקדמת יותר של כתב היד.
Materials
| Name | Company | Catalog Number | Comments |
| Adobe photoshop 2021 | Adobe | ||
| Anti-vibration table 30"x 36" - OTMC - 63533 | TMC | ||
| Cell sens standard software | OLYMPUS | ||
| Digidata 1440A | MOLECULAR DEVICES | ||
| Digital amplifier multiclamp 700B | MOLECULAR DEVICES | ||
| DSK line-up linearslicer pro7 | TED PELLA, INC | ||
| Micromanipulator MPC-385 / OSI-MPC-385-2 | OLYMPUS AMERICA INC | ||
| Micropipette puller P-97 | SUTTER INSTRUMENTS | ||
| Microscope BX51W1 | OLYMPUS AMERICA INC | ||
| MS ICE software | Microsoft Corporation | ||
| Ohaus balance model AV212 | Ohaus Adventurer | ||
| Olympus DPSI0 /DPS80 camera | OLYMPUS | ||
| pClamp and Axoclamp data Acquisition Softwares | MOLECULAR DEVICES | ||
| pH meter lab 850 benchtop | SCHOTT INSTRUMENTS | ||
| Sharp stainless blade | Dorco/Personna | ||
| Vapor pressure osmometer model 5600 | WESCOR INC | ||
| Water purification systems Smart2pure 6UV/UF | Thermo Scientific | ||
| Chemicals- list | |||
| Agrose Low melt IB70051 | IBI SCIENTIFIC | ||
| CaCl2 (Calcium Chloride) | ACROS organics | ||
| Cynergy instant adhesive CA6001 | Resinlab | ||
| Dextrose (D-(+)-glucose) | VWR Life Science | ||
| Ethyl alcohol | IBI SCIENTIFIC | ||
| KCl (Potassium Chloride) | Amresco.Inc | ||
| MgCl2 (Magnesium Chloride) | Sigma-Aldrich | ||
| NaCl (Sodium Chloride) | Amresco.Inc | ||
| NaH2PO4 (Sodium Dihydrogen Phosphate) | Amresco.Inc | ||
| NaHCO3 (Sodium Bicarbonate) | Amresco.Inc |
References
- Rubel, E. W., Parks, T. N. Organization and development of brain stem auditory nuclei of the chicken: tonotopic organization of n. magnocellularis and n. laminaris. Journal of Comparative Neurology. 164 (4), 411-433 (1975).
- Rubel, E. W., et al. Organization and development of brain stem auditory nuclei of the chicken: ontogeny of n. magnocellularis and n. laminaris. Journal of Comparative Neurology. 166 (4), 469-489 (1976).
- Shao, M., et al. Spontaneous synaptic activity in chick vestibular nucleus neurons during the perinatal period. Neuroscience. 127 (1), 81-90 (2004).
- Fukui, I., Ohmori, H. Tonotopic gradients of membrane and synaptic properties for neurons of the chicken nucleus magnocellularis. Journal of Neuroscience. 24 (34), 7514-7523 (2004).
- Sanchez, J. T., Seidl, A. H., Rubel, E. W., Barria, A. Preparation and culture of chicken auditory brainstem slices. Journal of Visualized Experiments. (49), e2527 (2011).
- Sanchez, J. T., Lu, Y., Fay, R. R., Popper, A. N., Cramer, K., Coffin, A. Glutamate signaling in the auditory brainstem. Auditory Development and Plasticity: Springer Handbook of Auditory Research. 64 (4), 75-108 (2017).
- Parks, T. N. Morphology of axosomatic endings in an avian cochlear nucleus: nucleus magnocellularis of the chicken. Journal of Comparative Neurology. 203 (3), 425-440 (1981).
- Jhaveri, S., Morest, D. K. Sequential alterations of neuronal architecture in nucleus magnocellularis of the developing chicken: a Golgi study. Neuroscience. 7 (4), 837-853 (1982).
- Carr, C. E., Boudreau, R. E. Central projections of auditory nerve fibers in the barn owl. Journal of Comparative Neurology. 314 (2), 306-318 (1991).
- Köppl, C. Auditory nerve terminals in the cochlear nucleus magnocellularis: differences between low and high frequencies. Journal of Comparative Neurology. 339 (3), 438-446 (1994).
- Fukui, I., et al. Improvement of phase information at low sound frequency in nucleus magnocellularis of the chicken. Journal of Neurophysiology. 96 (2), 633-641 (2006).
- Wang, X., et al. Postsynaptic FMRP regulates synaptogenesis in vivo in the developing cochlear nucleus. Journal of Neuroscience. 38 (29), 6445-6460 (2018).
- Lu, T., Cohen, A. L., Sanchez, J. T. In ovo electroporation in the chicken auditory brainstem. Journal of Visualized Experiments. (124), e55628 (2017).
- Hong, H., Sanchez, J. T. Need for speed and precision: structural and functional specialization in the cochlear nucleus of the avian auditory system. Journal of Experimental Neuroscience. (12), 1-16 (2018).
- Hong, H., et al. Diverse intrinsic properties shape functional phenotype of low-frequency neurons in the auditory brainstem. Frontiers in Cellular Neuroscience. 12, 1-24 (2018).
- Wang, X., Hong, H., Brown, D. H., Sanchez, J. T., Wang, Y. Distinct neural properties in the low-frequency region of the chicken cochlear nucleus magnocellularis. eNeuro. 4 (2), 1-26 (2017).
- Tabor, K. M., et al. Tonotopic organization of the superior olivary nucleus in the chicken auditory brainstem. Journal of Comparative Neurology. 520 (7), 1493-1508 (2012).
- Lipovsek, M., Wingate, R. J. Conserved and divergent development of brainstem vestibular and auditory nuclei. Elife. 7, 40232 (2018).
- Passetto, M. F., et al. Morphometric analysis of the AMPA-type neurons in the Deiter's vestibular complex of the chick brain. Journal of Chemical Neuroanatomy. 35 (4), 334-345 (2008).
- Curry, R. J., Lu, Y. Intrinsic properties of avian interaural level difference sound localizing neurons. Brain Research. 1752, 147258 (2021).
- Aleksandrowicz, E., Herr, I. Ethical euthanasia and short-term anesthesia of the chick embryo. ALTEX - Alternatives to Animal Experimentation. 32 (2), 143-147 (2015).
- Palkovits, M. Isolated removal of hypothalamic or other brain nuclei of the rat. Brain Research. 14 (59), 449-450 (1973).
Reprints and Permissions
Request permission to reuse the text or figures of this JoVE article
Request PermissionThis article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. All rights reserved