
Method Article
胚性ニワトリ聴覚脳幹をスライスしてトノトピック勾配とマイクロ回路を評価する
要約
ここでは、1つの脳幹スライス内のトノトピック特性と発生軌跡を調べるために、ニワトリ胚の非冠状聴覚脳幹スライスを取得するためのプロトコルを提示します。これらのスライスは、矢状、水平、および水平/横セクションを含み、従来の冠状セクションよりも個々のスライス平面内のより大きなトノトピック領域を包含する。
要約
ニワトリ胚は、聴脳幹を研究するために広く受け入れられている動物モデルであり、高度に特殊化されたマイクロ回路と、トノトピック(すなわち周波数)軸に沿って差動的に配向されたニューロントポロジーで構成されています。トノトピック軸は、吻側内側面における高周波音の分離符号化および尾側領域における低周波符号化を可能にする。伝統的に、胚組織の冠状脳幹スライスは、相対的な個々の等周波薄板の研究を可能にする。個々の等周波領域に関する解剖学的および生理学的問題を調査するのに十分であるが、より大きな聴覚脳幹領域にわたるトノトピック変異およびその発達の研究は幾分限られている。このプロトコルは、下聴脳幹の周波数領域のより大きな勾配を含むニワトリ胚からの脳幹スライス技術を報告しています。ニワトリの聴覚脳幹組織に異なるスライス法を利用することで、1つの脳幹スライス内で電気生理学的および解剖学的実験が可能になり、冠状断面よりもトノトピック特性と発生軌跡のより大きな勾配がよりよく保存されます。複数のスライス技術により、聴覚脳幹マイクロ回路の多様な解剖学的、生物物理学的、およびトノトピック特性の調査を改善することができます。
概要
ニワトリ胚は、細胞生物学、免疫学、病理学、発生神経生物学など、数多くの多様な科学分野における基本的な生物学的問題を研究するための貴重な研究モデルです。ニワトリの聴覚脳幹の微小回路は、聴覚形態と生理学の観点から理解できる高度に専門化された回路の優れた例です。例えば、Rubel and Parks (1975) は、ニワトリ核マグノセルラリス (NM) と層状核 (NL) のトノトピック配向 (すなわち周波数勾配) を、矢状面に対して~30°配向した核の軸を横切る線形関数として最初に記述した。NMとNLの個々のニューロンは、特性周波数(CF)として知られる最高の音の周波数を、吻側内側面に沿って尾外側領域にエンコードします。高周波感受性ニューロンは吻側内側領域にあり、低周波感受性ニューロンは尾側に位置する。そのため、トノトピック特性を研究するための聴覚脳幹組織の伝統的な解剖方法は、連続した冠状スライスを利用してきました。実際、発生中のニワトリ胚の聴覚マイクロ回路は、何十年にもわたって連続する尾側から吻側への冠状動脈面脳幹スライスを介したトノトピック聴覚機能の信号処理を研究するためのモデルシステムとして確立されています1,2,3,4,5,6。
しかし、NMとNLのトノトピック構成はトポロジー的・形態的に複雑である。聴神経入力は、高いCF入力が、副生NM細胞の体周囲長の少なくとも4分の1を覆う終末球様構造で終わるように分布している。逆に、低CF入力は、終末球様終末ではなく、NMニューロンの樹状突起上の複数のブートンシナプスで組織化されています。中央のCF入力は、終末球とブートン様シナプス4、7、8、9、10、11、12の両方で終了します。NLでは、非常にステレオタイプの樹枝状勾配は、樹枝状の長さだけでなく樹枝状の幅にも明らかです。このユニークな樹枝状勾配は、トノトピック軸に密接に適合しています。樹状突起は、高CFニューロンから低CFニューロンまで、それぞれ長さが11倍、幅が5倍に増加します6。コロナスライスにおけるこれらの核のこのような複雑な分布を克服するために、このプロトコルは矢状面、水平面、および水平/横面での解剖アプローチを記述します。これらのスライス技術は、個々のスライス面において最大のトノトピック特性を示す聴覚脳幹組織の例を提供する。
プロトコル
すべての手順は、ノースウェスタン大学施設動物管理および使用委員会(IACUC)によって承認され、実験動物の管理および使用に関する国立衛生研究所のガイドラインに従って実施されました。脳幹組織の解剖および調製のためのプロトコルは、以前のプロトコル5,13に準拠しています。
1.卵の取り扱い
- IACUCが承認した地元の動物サプライヤーから受精卵(Gallus gallus domestic us)を購入します。
- 卵は到着後すぐに14°Cの冷蔵庫に保管し、5日以内にインキュベートします。
注:胚の生存率は1週間後に大幅に低下します。 - 卵を70%エタノールで殺菌してから、38 ± 1°C、湿度~50%で孵化させます。
2.人工脳脊髄液(ACSF)の組成と調製
- 1 Lの18.2 MΩcm dH 2 Oに次の化学物質を混合して、10x ACSFストック溶液を作成します:NaCl(塩化ナトリウム)130 mM、NaHCO3(重炭酸ナトリウム)26 mM、KCl(塩化カリウム)2.5 mM、NaH2PO4(リン酸二水素ナトリウム)1.25 mM、デキストロース(D-(+)-グルコース)10 mM。原液を冷蔵庫に保管してください。
- MgCl 2(塩化マグネシウム)1 MおよびCaCl 2(塩化カルシウム)1 M溶液を別々に18.2 MΩcm dH2 Oで調製し、冷蔵庫に保存する。
- 使用直前に、10x ACSFを1xに希釈し、95%O 2/5%CO 2で15〜20分間連続的にバブリングし、MgCl 2およびCaCl 2を添加する。ACSFおよびdACSF(ACSFを解剖)を調製するには、それぞれMg2+ 1 mM、Ca2+ 3 mMおよびMg 2+ 3 mM、Ca 2+ 1 mMの最終濃度に調整します。
- pHが7.2〜7.4、浸透圧が300〜310 mOsm / Lになるように、ACSFのバブリング速度を設定します。
注:バブリング中にACSFを氷浴に入れることは、解剖中に組織の構造的完全性をサポートする低い溶液温度を維持するのに有益です。
3.アガロース(5%)ブロック調製
- 5 gのアガロースを100 mLのdACSFに混合します。100°Cの水浴または電子レンジを2〜3分間使用し、アガロースが完全に溶解して泡立ち始めるまで塊を防ぐために30秒ごとに攪拌します。
- 溶かしたアガロースを厚さ5 mmまでの空のペトリ皿に注ぎ、室温で保って固めます。固めた後、パラフィルムを用いてシャーレを密封し、4°Cで保存する。
- アガロースを鋭利な刃で立方体のブロックに切断し、解剖時に使用します。
4.解剖プロトコルと聴覚脳幹の分離
- 70%エチルアルコール溶液スプレーを使用して解剖領域を洗浄します。
- 支持または角度の付いたアガロースブロックをビブラトームトレイに接着します。
- 希望する年齢の卵を選択してください(現在のプロトコルではE20とE21)。ステップ1のように上記のプロトコルに従って卵を扱い、孵化させます。
- 卵を明るい光の下に置き(キャンドリング)、卵の大きい側または丸い側でこのスペースを探して、空気で満たされたスペースを見つけます。
- 卵を室温に順応させ、空気で満たされた空間で殻を割って、膜嚢を露出させます。
- くちばしを露出させるために嚢を穏やかに切開します。
- メスで、首と頭を卵からそっと引き出します。
- 鋭利なハサミを使って頭を素早く斬首します。
- 断頭後、氷冷したdACSFで頭をきれいにし、解剖パッドから余分な血液を取り除きます。
- 氷冷したdACSFで頭をしっかりと保持し、吻側尾切開を行います。目の後ろと目の間の切開を開始し、収穫した首の長さに従います。
注:若い胚は、切開を行う際に必要な圧力が少なくて済む場合があります。 - 頭蓋骨を露出させるために皮膚を分離します。
- 目の後ろの頭蓋骨を正中線から横方向に切ります。両方の半球に対してこれを行います。
注:このステップは、脳組織を無傷に保ちながら、頭蓋骨の吻側部分を付着した脳から分離するのに役立ちます5。 - 頭蓋骨の吻側部分をスライスします。刃を目の後ろに置き、すばやく切ります。
注意: 付属の頭蓋骨をきれいに切るには、努力が必要な場合があります。 - 冷たいdACSFの皿に頭を浸します。
- 小さなはさみを利用して、頭蓋骨の尾部に正中線から横方向の切開を行い、組織に損傷を与えることなく頭蓋骨から脳を分離しようとします。
- 脳幹と小脳をやさしく露出させます。頭蓋骨全体の背側を引っ込め、脳幹を慎重に取り除き、穏やかなそりで細かい絵筆を使って露出させます。湾曲した鉗子を使用して、組織と血管の接続から脳幹をきれいにします。8番目の 脳神経領域に特に注意し、両側に無傷の神経線維を短く残すようにしてください。
- 花柄を切断し、血管を慎重に取り除くことにより、脳幹を小脳から分離します。追加の血管の脳幹を整えます。
注意: 手順全体が、カルボキシゲ(95%O 2/5%CO2)で連続的にバブリングされた氷冷dACSFで実行されていることを確認してください。
5.ビブラトームスライス
注意: 次の手順では、組織の背面を立方体のアガロース片で支える必要があります。
- ビブラトームブレードを水平軸に沿って置き、脳幹をスライストレイに接着します。吻側を接着し、冠状スライスのために吻側尾軸を垂直に保ちます。
- 矢状スライスのために外側内側軸を垂直に保ちます。
- 腹側を接着し、水平スライスのために背腹軸を垂直に保ちます。
- 鋭角矢状水平面を実現するには、脳幹の腹側を接着し、腹側背軸を45°の角度で切断されたアガロースブロックの斜辺表面に垂直に保ちます。アガロースブロックの反対側の表面をスライストレイに面して接着し、吻側尾側軸をブレードエッジと平行に保ちます。
6.壊れやすいまたは大きな脳幹組織の取り扱い
- ステップ5の別のアプローチでは、分離した脳幹を4%低融点(LMP)アガロースに~40°Cで35 mm x 10 mmのペトリ皿に浸します。
- 浸した脳幹にアガロースを注いだ後、ペトリ皿を氷の上に置いて固化させます。鋭いかみそりの刃を使用して、脳幹が埋め込まれた立方体のアガロースブロックを切断します。
- LMPアガロースブロックを吻側に接着し、脳幹の吻側尾軸を垂直に保ちます。
- NM領域が可視化されるまで冠状スライスを取ります。
- 鋭利な刃で接着剤からアガロースブロックを取り除きます。核を見つけるには、0.5 μLの色素(トルイジンブルーまたはオレンジG)を細い針でNMにそっと置きます。
- 矢状または水平スライスのためにこのブロックをスライストレイに再度取り付け、染色領域に対して核を特定します。
- 最高のパフォーマンスを得るには、 振動体のスライス速度を4〜5(~30 ± 4 mm / min)、 振動周波数を85〜87 Hz、 スライス振幅を4〜6(~1 ± 0.2 mm)に設定します。
- 脳幹切片作製後、200〜300μmの順次採取したスライスを市販のスライスチャンバーに入れ、ACSF中で室温で1時間平衡化し、95%O2/5%CO2(pH 7.2〜7.4、浸透圧300〜310mOsm/L)の混合物を連続的にバブリングした。これらの条件では、スライスは5〜6時間まで生存し続けます。
7.電気生理学:パッチクランプ手順
- 脳幹スライスを記録チャンバーに移し、カルボキシゲン化ACSFを連続的に灌流します~1.5 ± 0.5 mL/min。
- 先端直径1〜2μm、抵抗3〜6MΩの範囲のマイクロピペットプーラーでパッチピペットを引きます。
- ピペットにK-グルコン酸塩ベースの内部溶液を充填します(電流クランプ記録用)。
- スライス内の異なるトノトピック領域にわたるニューロン特性をテストするには、スライス平面の両端にニューロンを配置し、記録電極でアプローチします。
- ニューロンに近づきながら、ピペットチップで正の空気圧を維持します。
- ニューロンにくぼみが視覚化されるまで、体細胞に向かって移動します。次の 2 つの手順をすばやく実行します。
- 正圧を解放してギガオーム(1GΩ)シールを作成します。
- アンプ設定を電圧クランプモードのままにし、ピペットオフセットをゼロpAに修正します。シールテスト(100Hzで10mVテストパルス)を実行します。負圧を適用して、ニューロン膜の小さなパッチを破裂させます。
- 聴覚ニューロンの能動的固有特性をテストするには、過分極および脱分極体細胞電流注射を適用します。
注:この手順の例は、補足ビデオS1、S2で視覚化できます。この手順の詳細は、ビデオの凡例に記載されています。
結果
ここに示すすべての脳幹スライスは、脳幹組織(~200-300 μm)から取得され、5倍の対物レンズおよび微分干渉コントラスト(DIC)光学系を使用して画像化されました。カメラは解剖顕微鏡に取り付けられ、画像取得ソフトウェアを備えたコンピューターに接続されました(材料表を参照)。これらの図の衛星挿入図(右パネル)は、60倍の倍率の水浸対物レンズを使用して画像化されました。デジタル画像を取得する際に、脳幹スライスのすべての領域が均等に拡大されるように注意が払われました。写真は最適な明るさと焦点で撮影されました。脳幹スライスのデジタル画像は、重なり合う領域に基づいて平面的にステッチされ、明るさ、コントラスト、およびグレースケールをさらに調整するためにデスクトップコンピューターにインポートされました。ニワトリ聴覚脳幹の基本的な微小回路は、以前の研究1,2,5,13に従って同定された。顕微鏡(5倍対物レンズ)では、聴覚核は、スライスの背側領域に沿って同側および反対側の両方で各核の周りを行き交う重有髄神経線維に隣接する領域として特定されました。
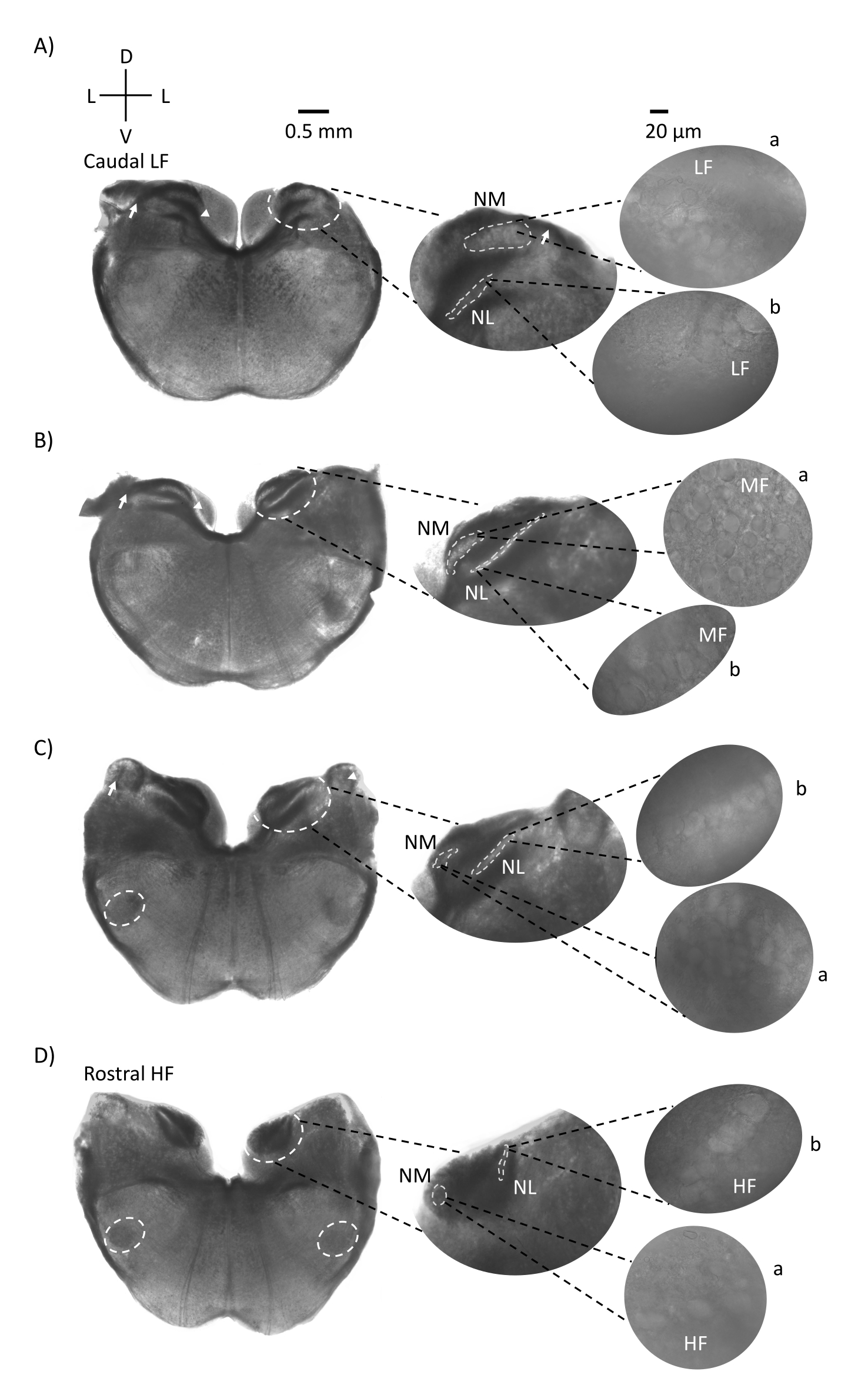
図1は、E21ニワトリ胚由来の脳幹組織(200〜300μm)の伝統的な冠状切片を示しています。ここに示す4つの冠状動脈スライスは、聴覚脳幹核の相対的な等周波領域を表しており、最も低いCF聴覚領域(図1A、尾側)から最も高いCF聴覚領域(図1D、吻側内側)まで進行しています。図1A-Dの4つの冠状スライスすべてについて、標識されたNMとNLの拡大領域が中央の列に表示され、図パネルの右側に拡大(60倍の対物レンズ)が示されています(それぞれ衛星挿入図のaとb)。図1A,Bの矢印は聴神経線維の入力を示し、矢印はスライスの左側のNM軸索の分岐を示しています。図1Cは、角核として知られる別の鳥類蝸牛核構造を示しています(NA、左矢印、右矢印)。2つの最も吻側の冠状動脈スライスは、冠状動脈スライスの腹側外側領域に沿って位置する上オリバリー核(SON)を示しています(図1C、D、白い破線の円)。
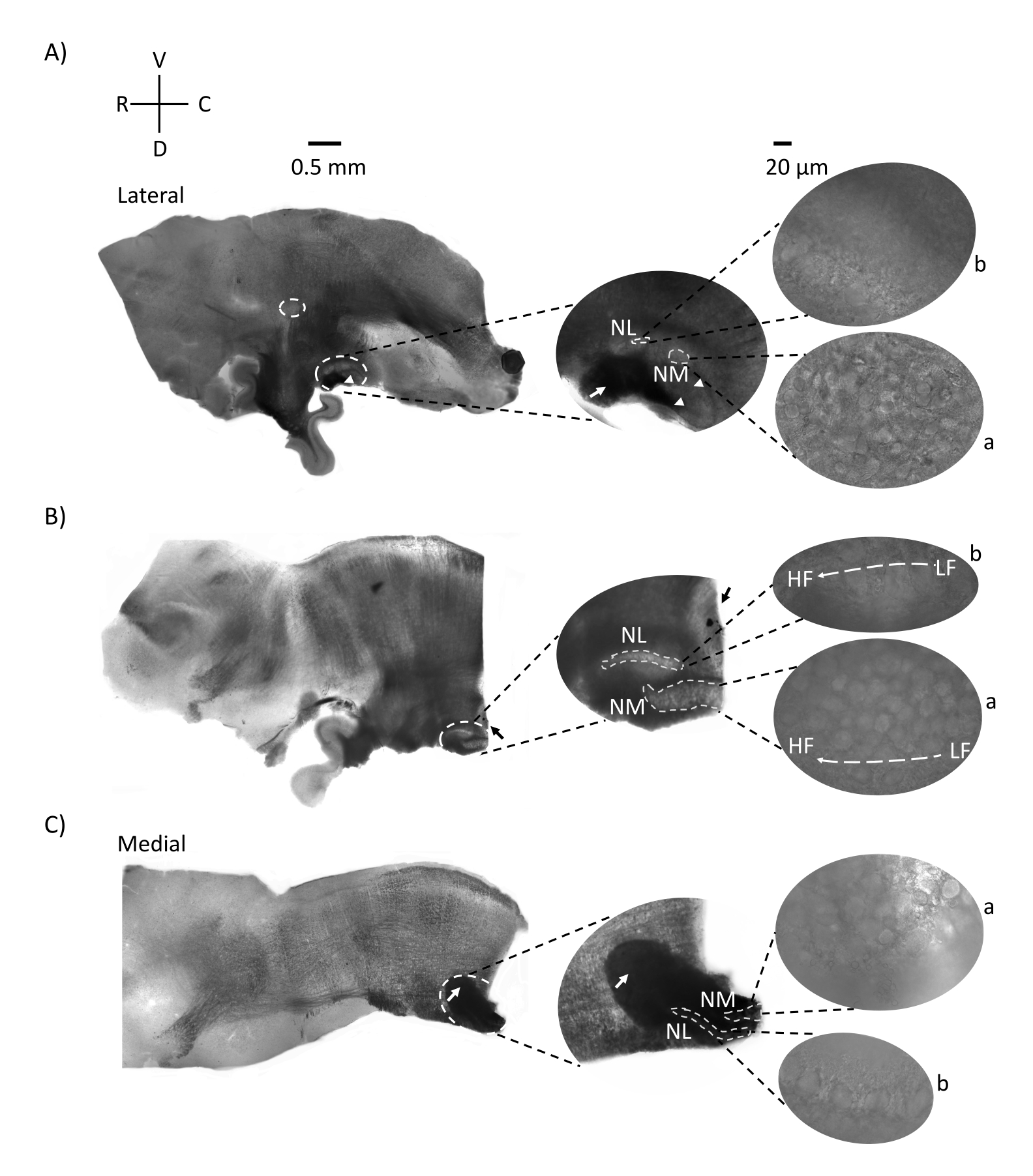
図2は、E21ニワトリ胚由来の脳幹組織(200-300μm)の矢状切片を示す。3つの矢状スライスすべて(図2A-C)について、標識されたNMとNLの拡大領域が中央の列に表示され、図パネルの右側に拡大(60倍の対物レンズ)が表示されます(衛星画像ではそれぞれaとb)。NMとNLは、聴神経線維(図2A、中央の矢印)が高倍率で観察されたニューロンのクラスターに入り(図2A、中央、小、白い破線の円と矢印)、聴覚領域の始点を強調している場所を特定しました(図2A、左、大、白い破線の円と矢印)。SONは、最も外側のスライスの吻側外側領域で同定された(図2A、小さい、白い、破線の円)。図2Bは、NMおよびNLからの比較的低CFおよび高CFの両方の聴覚領域を吻側尾軸に沿って含む拡張トノトピック領域を示す(白色の輪郭領域、衛星挿入図も参照)。図2Cは、最も内側スライスの同側および反対側の軸索房および聴覚領域の終点(左および中央矢印)を示す。ここに示すスライスの向きは、図1に見られるような従来のスライスの向き(すなわち、冠状)とは対照的です。これは、電気生理学的記録に必要なガラスピペットのアプローチに最も適した向きを表示するために行われた。
トノトピック軸の大きな領域が図2Bに表されていることを確認するために、NMニューロンからの電流クランプ電気生理学記録を行った。 図3は、冠状スライス(図3A、B)および矢状スライス(図3C、D、補足ビデオS1、S2)から記録された成熟(E21)NMニューロンの機能的類似点および相違点を示す。2つのNMニューロンを冠状スライスの内側および外側の端から選択し(図1Bに示すスライスと同様)、矢状スライスのNMの吻側端および尾側端から2つのNMニューロンを選択した(図2Bに示すスライスと同様)。図3A、Bは、体細胞電流注入と同様の電気生理学的応答特性を示しています(−100 pA〜+200 pA、+10 pA刻み、100 ms持続時間)。これら2つのNMニューロンの発火パターンは、このスライス平面において微妙な違いを示し、中周波NMニューロンの相対的な等周波ラミナを示す。図3C,Dは、発火パターンが吻側-尾側軸全体で実質的な違いを有することを示しており、低周波NMニューロン(図3C)から高周波NMニューロン(図3D)への比較的高いトノトピック勾配を示しています。両方のニューロンは、以前に報告されたように、ステレオタイプの発火パターンを示しました14,15。
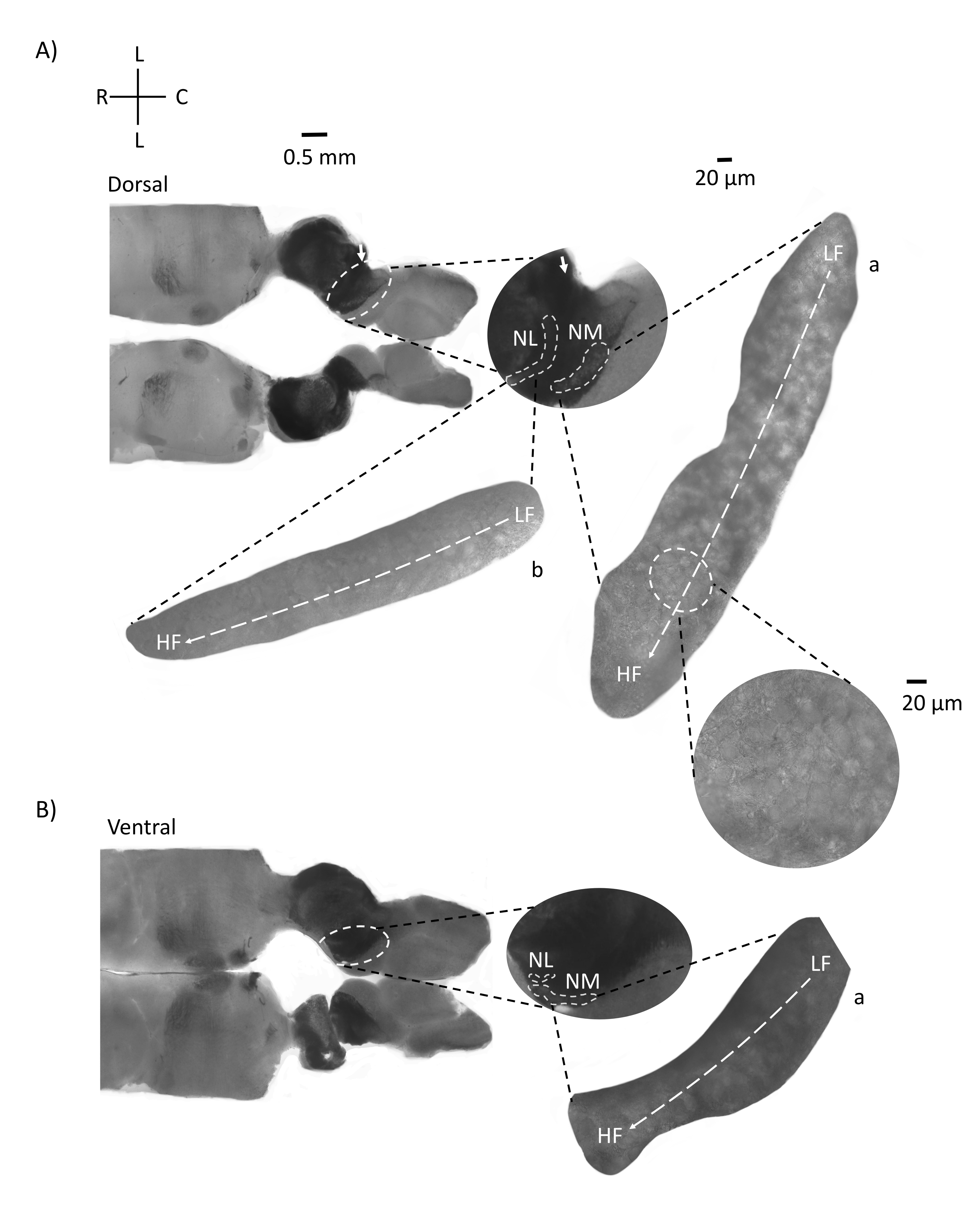
図4は、E21ニワトリ胚の脳幹組織(200-300μm)の水平断面を示す。両方の水平スライス(図4A、B)について、標識されたNMとNLの拡大領域が中央の列に表示され、図パネルの右側に拡大(60倍の対物レンズ)が表示されます(それぞれ衛星インセットのaとb)。水平スライスでは、NMとNLが正中線に向かって識別され、ニューロンは外側内側軸に沿って広がっていました(図4A、B、中央、白、破線の輪郭領域)。拡大画像は、トノトピック勾配の大きな範囲を示しています。低周波ニューロンは尾側領域にあり、高周波ニューロンは吻側内側領域にあります(図4A、B、右、衛星)。吻側 - 尾側軸に沿って正中線を通る繊維は、聴覚核の反対側の接続を示すが、これらの繊維の構成は単純な平面内ではない。ただし、水平/横断面からの鋭い角度スライスは、これらの軸索線維を矢状面に向かってたどることができます。水平面から鋭角(45°)で厚さ200〜300μMの脳幹組織のスライスを図5に示します。聴覚脳幹核は、最も外側のスライスから始まり、最も内側のスライスで終わる大きな斜めの広がりを横切って見ることができます(図5A-C、ラベル付きの中央パネル、白い輪郭の領域)。さらに、NMおよびNL領域の角度配向は、連続した非対称スライスで視覚化することもできます(図5A-C、ラベル付き中央パネル、白、破線の輪郭領域)。拡大画像(60倍対物レンズ)は、聴覚核が吻側内側軸から尾外側軸に沿って進むときの眼圧視軸を示しています(図5A-C、右、衛星挿入図)。図 5 のスライスの向きは、図 2 の向きと似ています。それらは従来の画像の提示とは対照的ですが、電気生理学的実験により適しています。

図1:脳幹の代表的な冠状連続切片。 (A-D) 左:尾軸から吻側軸へのスライス、聴覚核、および白い破線の円でマークされた接続繊維。中央の挿入物は聴覚領域のより大きなビューであり、核は白い破線の円a:NMおよびb:NL内に示されています。矢印は聴神経求心性線維を示し、矢印はA、BのNM軸索分岐を示す。矢印はCのNAを示す。横方向の白い破線の円は、C、DのSONを示しています。右:衛星挿入物は、これらの原子核を60倍の対物レンズで示しています:a:NMおよびb:NL。略語:NM =核マグノセルラリス;NL =核ラミナリス;NA =角核;SON =優れたオリバリー核;LF =比較的低周波ニューロン。MF =中周波ニューロン;HF =高周波ニューロン;D =背側;L =横方向。V =腹側。この図の拡大版を表示するには、ここをクリックしてください。
{kind=link}

図2:脳幹の代表的な矢状連続切片。 (A-C) 左:外側軸から内側軸へのスライスで、聴覚核は白い破線の円でラベル付けされています。中央の挿入物は、同じ聴覚核領域を拡大して示しており、白い破線の円でマークされています。(A)スライスの中央にある白い破線の円は、SONを強調しています。矢印は聴神経線維を示し、矢印はNAを示す。スライスの右側の先端にある暗い黒い斑点は、イメージングアーティファクトです。小脳の領域は、左パネルのスライスA及びBの両方で聴覚領域に対して背側が見ることができる。(B)向きが冠状面に変更された矢状スライス(スライス中)。聴覚領域は青色染料(黒い矢印)で識別され、矢状面で再びスライスされました。(A-C)中央の挿入NMおよびNL領域は、白破線でマークされています。右:60倍の対物倍率で観察されたa:NMとb:NLの衛星図。聴覚核におけるLFおよびHFトノトピック勾配は、吻側尾軸に沿って示される。(C)の暗い領域を指す矢印は、内側軸を通って正中線を横切って走る重有髄NM線維を示す。繊維は聴覚核の両側を接続します。略語:NM =核マグノセルラリス;NL =核ラミナリス;NA =角核;SON =優れたオリバリー核;LF =比較的低周波ニューロン。HF =高周波ニューロン;D =背側;V =腹側;R =吻側;C =尾側。この図の拡大版を表示するには、ここをクリックしてください。
{kind=link}

図3:電流クランプモードでの体細胞電流注入(-100 pA〜+200 pA、+10 pA刻み、100 ms持続時間)に対するニューロン応答の電気生理学的記録。 ニューロンは、NMの極端な反対領域である同じスライス内の記録のために選択された。(A,B)単一の冠状動脈スライスにおける代表的なニューロン応答は、微妙な違いを伴う相対的な等周波特性を示す。応答特性は、コロナスライス内のNMの最も内側(A)および外側(B)領域から記録された2つの異なるMFニューロンを表す。(C,D)単一の矢状スライスからの代表的なニューロン記録。記録は、比較的LF NM応答(C)とHF NM応答(D)を示しており、単一の矢状断面に沿ったトノトピック勾配の実質的な違いを強調しています。略語:NM =核マグノセルラリス;LF =比較的低周波ニューロン。MF =中周波ニューロン;HF =高周波ニューロン。 この図の拡大版を表示するには、ここをクリックしてください。
{kind=link}

図4:脳幹の代表的な水平連続切片。 (A,B)左:背側から腹側軸に沿ったスライス、聴覚核は白い破線の円でマークされています。8番目の 脳神経求心性線維は、矢印でマークされた聴覚核を接続します。中央の挿入物は、耳核領域の拡大図であり、白い破線の下にマークされた聴覚核NMおよびNL領域が示されている。聴覚核の明確なトポロジカル運動は、 A、Bに見ることができます。(A,B)右: a:NMと b:NLを示す大きな衛星図。右の挿入物は、60倍の対物倍率で観察された聴覚核と、尾側外側から吻側内側軸に沿ったLFからHFまでの湾曲したトポロジカル軸を示しています。略語:NM =核マグノセルラリス;NL =核ラミナリス;LF =比較的低周波ニューロン。HF =高周波ニューロン;L =横方向。R =吻側;C =尾側。 この図の拡大版を表示するには、ここをクリックしてください。
{kind=link}

図5:代表的な水平/横の鋭角(45°)シリアルセクション。 (A-C)左:脳幹の連続切片、白い破線の円でマークされた聴覚核。中央の挿入物は、聴覚領域のより大きなビューです。(A)中央の挿入物は、これらのスライスにおけるNMおよびNLニューロンの最大広がりを示す。(B,C)中央の挿入物:白い破線でマークされた聴覚核は、(A-C)と比較すると徐々にトポロジカルな変化を示します。右:聴覚核a:NMとb:NLを60倍の対物倍率で示す衛星挿入物。NMおよびNLのLF領域からHF領域へのトノトピック軸は、外側スライスから内側スライスに向かって角度的に回転します。略語:NM =核マグノセルラリス;NL =核ラミナリス;LF =比較的低周波ニューロン。HF =高周波ニューロン;V =腹側;R =吻側;D =背側;C =尾側。この図の拡大版を表示するには、ここをクリックしてください。
{kind=link}
補足ビデオS1:過分極および脱分極体性電流注射。 低周波および高周波ニューロンから電流クランプモードでの100ミリ秒の体細胞電流注入までの応答特性。ニューロンは、同じ矢状脳幹スライスから選択された。注入の範囲は-100〜+200 pAで、+10 pA刻み、時間持続時間は100 msです。活動電位は、十分な脱分極電流ステップに応答して見られる。ビデオは、 図3Cに示す最終的なトレースに対応しています。 このファイルをダウンロードするには、ここをクリックしてください。
補足ビデオS2:過分極および脱分極体細胞電流注射。 補足ビデオ S1と同様に、このビデオは、電流クランプモードでの100ミリ秒の体細胞電流注入に対する低周波および高周波ニューロンからの応答特性を示しています。ニューロンは、同じ矢状脳幹スライスから選択された。注入の範囲は-100〜+200 pAで、+10 pA刻み、時間持続時間は100 msです。活動電位は、十分な脱分極電流ステップに応答して見られる。ビデオは、 図3Dに示す最終的なトレースに対応します。 このファイルをダウンロードするには、ここをクリックしてください。
ディスカッション
ニワトリ胚性脳幹組織の冠状切片は、何十年にもわたって相対的な個々の等周波薄板の研究を可能にしてきました1,2,5。しかし、ニワトリ聴覚脳幹のトノトピック(すなわち周波数)組織はトポロジー的に複雑であり、特定の研究課題に応じて他の解剖学的軸でよりアクセスしやすい可能性がある。個々の等周波領域に関する解剖学的および生理学的問題を調査するのに十分であるが、より大きな聴覚脳幹領域にわたるトノトピック変異およびその発達の研究は、冠状切片によっていくらか制限されている。この制限を克服するために、このプロトコルは、矢状面、水平面、および水平/横面でのアプローチを記述して、個々の脳幹セクションで最大のトノトピック特性と勾配を示す聴覚脳幹組織の追加の例を提供します。
聴覚脳幹領域の矢状切片は、冠状切片と比較して、異なるトノトピック領域がスライス内のより大きな領域に分布していることを示しています(矢状聴覚領域=~300-600μm、冠状聴覚領域=~200-350μm)。例えば、NMおよびNL領域は、矢状切片の吻側尾軸に沿ったより広い領域にわたって視覚化され(例えば、図2B)、この解剖学的軸に沿って走る機能的トノトピック勾配は、主に単一の矢状スライス内に含まれていた。これは、以前に報告されたように、吻側尾側勾配に沿って変化する固有のニューロンの違いの電流クランプ記録でさらに確認されました14,15(例えば、図3C、D)。トノトピック軸に沿った解剖学的および免疫組織化学的特性を強調する将来の実験では、単一の矢状スライス面内の聴覚特性の既知の勾配をさらに調査する可能性があります。これらには、連続した冠状断面16において以前に示されている樹状突起構造の既知の勾配ならびにNMおよびNLの固有の特性であるMAP2染色およびカリウムチャネル発現パターンが含まれるが、これらに限定されない。
聴覚脳幹領域の水平断面は、NMとNLが正中線に向かって位置していることを示しています。聴覚軸索線維の一部は、水平面に対して斜めまたは垂直に走っています(図4)。これらの繊維は、矢状面に対して45°の鋭角スライスを作ることによって続くことができる。得られた水平/横スライスは矢状または水平スライスよりも大きく、長い軸索線維は同側と反対側の両方で吻側尾軸を通り抜けました。NMとNLの両方をより大きな対角領域(~400-700 μm)で視覚化できるため、反対側の接続を外側-内側軸に沿って視覚化できます。さらに、水平/横スライス平面は、聴覚領域と結果として生じるトノトピック勾配がどのように角度回転するかも示しています(図5)。より広い領域での対側接続の角度露出により、これらのスライスは、従来の冠状スライスよりも電気生理学的刺激および微小回路研究に適しています。
その他の利点
聴覚マイクロ回路の形成には、神経細胞の生存、シナプス形成、軸索分化、樹状突起構造、および成熟を促進する手がかりの時空間的調整が必要です。したがって、ニワトリ胚聴覚マイクロ回路の代替脳幹切片は、以下の研究トピックに使用することができる:地形的に異なる次元におけるニューロンの形態学的組織;すべての聴覚および前庭核のコネクトームを整理およびマッピングする。等周波およびトノトピック平面における回路構成の活性パターンの同定と特性評価。興奮性マイクロ回路と抑制性マイクロ回路の地形的組織と特殊なニューロン集団(核)との関係。聴覚核ニューロンの空間的位置とその予測CF17;特定のトノトピーニューロンタイプの系統的標的化;前駆細胞と保存された核へのそれらの発達を追跡する。神経回路の進化に対する細胞の遺伝的系統18;種間の比較脳幹解剖学;デイター前庭複合体(DC)19のような前庭回路の調査。前庭核間の同期とクロストーク。
異なるスライス面を用いた多面的なアプローチは、脳幹微小回路の未知の解剖学的および生物物理学的特性に関する基本的な質問に答えるのに役立つ可能性があります。良い例は、主要な聴覚核(NM、NA、NL、およびSON)と、外側レムニスカス(LLDp)、半月核(SLu)20、および接線核(TN)3の背側核を含む前庭核との関係です。ただし、このプロトコルとこれらのスライスベースの研究にはいくつかの制限があります。
注意事項と制限事項
実験を行う機関によって、倫理ガイドラインや鶏胚の取り扱いが異なる場合があります。実験動物の世話と使用に関する国立衛生研究所のガイドラインは迅速な断頭を許可していますが、鶏胚安楽死の代替方法があります21。初期発達中のニワトリ胚脳幹組織は、古い胚に比べて柔らかく繊細です。表面にいくつかの接続部と血管があり、それらを取り除くときは特に注意が必要です。組織は氷冷dACSFに保持し、生存率を高めるために95%O2/5%CO2 と灌流する必要があります。
矢状スライス法は、同側トノトピーにのみ役立ちます。このスライス方法は、冠状スライスよりも大きなスライスを提供し、その取り扱いは不安定になる可能性があります。しかしながら、他の場所で詳細に説明されているクロスニードル法を用いてスライスをトリミングすることができる22。4%LMPアガロースブロック包埋脳幹を使用すると、繊細な構造をスライスして保存できますが、過度に高温のアガロースを注がないように注意する必要があります。アガロースブロックされた脳幹を冷やした環境に~1分間置くことで素早く設定すると、スライスが電気生理学的記録に実行可能になります。
過剰量の瞬間接着剤の適用は有毒である可能性があります。それは最小限に適用する必要があり、余剰量はdACSFを交換してすぐに洗浄する必要があります。鋭角(45°)スライスの場合、アガロースブロックの角度をカットすることが重要です。鏡を使って正面の角度を見ることができ、アガロースブロックを鋭利な刃で切断します。市販のブレードにはワックスコーティングが施されている場合があり、使用前にアルコールで拭き取り、乾燥させる必要があります。軸索線維の房は皮質組織やマトリックス組織よりも硬いため、ビブラトームの切断速度と頻度を最適化する必要があります。高振幅を維持し、冷やした解剖液を使用すると、組織の損傷を防ぐことができます。
全ての溶液は新鮮に調製し、95%O 2/5%CO2をバブリングした後にCa 2+およびMg2+をACSFに添加すべきである。さもなければ、Ca2+の沈殿があるかもしれません。絵筆を使用して、ビブラトーム内のスライスを優しく処理する必要があります。可能であれば、合計スライス時間を15分未満に保ちます。ガラス製のパスツールピペットを使用して、脳幹スライスを操作できます。
電気生理学で使用されるスライスに接触するガラス製品や機器には、洗剤や腐食性の洗浄剤を使用しないでください。撮影された画像は、微分干渉コントラスト(DIC)光学系の下での厚さ200〜300μMの組織の外観を表しています。視覚的品質は免疫組織化学や電子顕微鏡よりも劣りますが、電気生理学的記録を実行するときに実験者が見るものを正確に反映しています。
背側腹側、吻側尾側、同側対側のいずれであっても、代替解剖学的軸に沿ったマイクロ回路の初期発達に関する研究は、ニワトリの聴覚脳幹では限られています。その理由の一つは、脳幹における転写コードの役割とトノトピック発生の制御がまだ完全には理解されていないためです。トップダウン変調や自発的活動などの機能現象は、 in vitroで活性を観察すると失われることがよくあります。しかし、 in vivo 研究は、これらのスライス条件でのみ可能な特異的かつ直接的な単一ニューロン記録によって補完されます。さまざまな方向に沿って脳幹組織を取得することの改良は、ニワトリの聴覚脳幹微小回路におけるトノトピック勾配の発達と複雑さに関する洞察に満ちた情報を提供する可能性があります。
開示事項
すべての著者は、研究が商業的または金銭的利益なしに実施され、利益相反がないことを宣言します。
謝辞
この作業は、NIH / NIDCD R01 DC017167助成金によってサポートされています。原稿の以前のバージョンに編集上のコメントを提供してくれたクリスティン・マクレランに感謝します。
資料
| Name | Company | Catalog Number | Comments |
| Adobe photoshop 2021 | Adobe | ||
| Anti-vibration table 30"x 36" - OTMC - 63533 | TMC | ||
| Cell sens standard software | OLYMPUS | ||
| Digidata 1440A | MOLECULAR DEVICES | ||
| Digital amplifier multiclamp 700B | MOLECULAR DEVICES | ||
| DSK line-up linearslicer pro7 | TED PELLA, INC | ||
| Micromanipulator MPC-385 / OSI-MPC-385-2 | OLYMPUS AMERICA INC | ||
| Micropipette puller P-97 | SUTTER INSTRUMENTS | ||
| Microscope BX51W1 | OLYMPUS AMERICA INC | ||
| MS ICE software | Microsoft Corporation | ||
| Ohaus balance model AV212 | Ohaus Adventurer | ||
| Olympus DPSI0 /DPS80 camera | OLYMPUS | ||
| pClamp and Axoclamp data Acquisition Softwares | MOLECULAR DEVICES | ||
| pH meter lab 850 benchtop | SCHOTT INSTRUMENTS | ||
| Sharp stainless blade | Dorco/Personna | ||
| Vapor pressure osmometer model 5600 | WESCOR INC | ||
| Water purification systems Smart2pure 6UV/UF | Thermo Scientific | ||
| Chemicals- list | |||
| Agrose Low melt IB70051 | IBI SCIENTIFIC | ||
| CaCl2 (Calcium Chloride) | ACROS organics | ||
| Cynergy instant adhesive CA6001 | Resinlab | ||
| Dextrose (D-(+)-glucose) | VWR Life Science | ||
| Ethyl alcohol | IBI SCIENTIFIC | ||
| KCl (Potassium Chloride) | Amresco.Inc | ||
| MgCl2 (Magnesium Chloride) | Sigma-Aldrich | ||
| NaCl (Sodium Chloride) | Amresco.Inc | ||
| NaH2PO4 (Sodium Dihydrogen Phosphate) | Amresco.Inc | ||
| NaHCO3 (Sodium Bicarbonate) | Amresco.Inc |
参考文献
- Rubel, E. W., Parks, T. N. Organization and development of brain stem auditory nuclei of the chicken: tonotopic organization of n. magnocellularis and n. laminaris. Journal of Comparative Neurology. 164 (4), 411-433 (1975).
- Rubel, E. W., et al. Organization and development of brain stem auditory nuclei of the chicken: ontogeny of n. magnocellularis and n. laminaris. Journal of Comparative Neurology. 166 (4), 469-489 (1976).
- Shao, M., et al. Spontaneous synaptic activity in chick vestibular nucleus neurons during the perinatal period. Neuroscience. 127 (1), 81-90 (2004).
- Fukui, I., Ohmori, H. Tonotopic gradients of membrane and synaptic properties for neurons of the chicken nucleus magnocellularis. Journal of Neuroscience. 24 (34), 7514-7523 (2004).
- Sanchez, J. T., Seidl, A. H., Rubel, E. W., Barria, A. Preparation and culture of chicken auditory brainstem slices. Journal of Visualized Experiments. (49), e2527 (2011).
- Sanchez, J. T., Lu, Y., Fay, R. R., Popper, A. N., Cramer, K., Coffin, A. Glutamate signaling in the auditory brainstem. Auditory Development and Plasticity: Springer Handbook of Auditory Research. 64 (4), 75-108 (2017).
- Parks, T. N. Morphology of axosomatic endings in an avian cochlear nucleus: nucleus magnocellularis of the chicken. Journal of Comparative Neurology. 203 (3), 425-440 (1981).
- Jhaveri, S., Morest, D. K. Sequential alterations of neuronal architecture in nucleus magnocellularis of the developing chicken: a Golgi study. Neuroscience. 7 (4), 837-853 (1982).
- Carr, C. E., Boudreau, R. E. Central projections of auditory nerve fibers in the barn owl. Journal of Comparative Neurology. 314 (2), 306-318 (1991).
- Köppl, C. Auditory nerve terminals in the cochlear nucleus magnocellularis: differences between low and high frequencies. Journal of Comparative Neurology. 339 (3), 438-446 (1994).
- Fukui, I., et al. Improvement of phase information at low sound frequency in nucleus magnocellularis of the chicken. Journal of Neurophysiology. 96 (2), 633-641 (2006).
- Wang, X., et al. Postsynaptic FMRP regulates synaptogenesis in vivo in the developing cochlear nucleus. Journal of Neuroscience. 38 (29), 6445-6460 (2018).
- Lu, T., Cohen, A. L., Sanchez, J. T. In ovo electroporation in the chicken auditory brainstem. Journal of Visualized Experiments. (124), e55628 (2017).
- Hong, H., Sanchez, J. T. Need for speed and precision: structural and functional specialization in the cochlear nucleus of the avian auditory system. Journal of Experimental Neuroscience. (12), 1-16 (2018).
- Hong, H., et al. Diverse intrinsic properties shape functional phenotype of low-frequency neurons in the auditory brainstem. Frontiers in Cellular Neuroscience. 12, 1-24 (2018).
- Wang, X., Hong, H., Brown, D. H., Sanchez, J. T., Wang, Y. Distinct neural properties in the low-frequency region of the chicken cochlear nucleus magnocellularis. eNeuro. 4 (2), 1-26 (2017).
- Tabor, K. M., et al. Tonotopic organization of the superior olivary nucleus in the chicken auditory brainstem. Journal of Comparative Neurology. 520 (7), 1493-1508 (2012).
- Lipovsek, M., Wingate, R. J. Conserved and divergent development of brainstem vestibular and auditory nuclei. Elife. 7, 40232 (2018).
- Passetto, M. F., et al. Morphometric analysis of the AMPA-type neurons in the Deiter's vestibular complex of the chick brain. Journal of Chemical Neuroanatomy. 35 (4), 334-345 (2008).
- Curry, R. J., Lu, Y. Intrinsic properties of avian interaural level difference sound localizing neurons. Brain Research. 1752, 147258 (2021).
- Aleksandrowicz, E., Herr, I. Ethical euthanasia and short-term anesthesia of the chick embryo. ALTEX - Alternatives to Animal Experimentation. 32 (2), 143-147 (2015).
- Palkovits, M. Isolated removal of hypothalamic or other brain nuclei of the rat. Brain Research. 14 (59), 449-450 (1973).
転載および許可
このJoVE論文のテキスト又は図を再利用するための許可を申請します
許可を申請This article has been published
Video Coming Soon
Copyright © 2023 MyJoVE Corporation. All rights reserved