
Method Article
Immagini tridimensionali di cellule batteriche per rappresentazioni cellulari accurate e localizzazione precisa delle proteine
In questo articolo
Riepilogo
Questo protocollo spiega come preparare e montare campioni batterici per l'imaging tridimensionale dal vivo e come ricostruire la forma tridimensionale di E. coli da queste immagini.
Abstract
La forma di un batterio è importante per la sua fisiologia. Molti aspetti della fisiologia cellulare come la motilità cellulare, la predazione e la produzione di biofilm possono essere influenzati dalla forma delle cellule. Le cellule batteriche sono oggetti tridimensionali (3D), anche se raramente sono trattati come tali. La maggior parte delle tecniche di microscopia genera immagini bidimensionali (2D) che portano alla perdita di dati relativi alla forma effettiva delle cellule 3D e alla localizzazione delle proteine. Alcuni parametri di forma, come la curvatura gaussiana (il prodotto delle due curvature principali), possono essere misurati solo in 3D perché le immagini 2D non misurano entrambe le curvature principali. Inoltre, non tutte le celle sono piatte durante il montaggio e l'imaging 2D di celle curve potrebbe non rappresentare con precisione le forme di queste cellule. Misurare accuratamente la localizzazione delle proteine in 3D può aiutare a determinare la regolazione spaziale e la funzione delle proteine. È stata sviluppata una tecnica di convoluzione in avanti che utilizza la funzione di sfocatura del microscopio per ricostruire le forme delle cellule 3D e per localizzare con precisione le proteine. Qui, viene descritto un protocollo per la preparazione e il montaggio di campioni per l'imaging di cellule vive di batteri in 3D sia per ricostruire una forma cellulare accurata che per localizzare le proteine. Il metodo si basa su una semplice preparazione di campioni, sull'acquisizione di immagini fluorescenti e sull'elaborazione delle immagini basata su MATLAB. Molti microscopi fluorescenti di alta qualità possono essere semplicemente modificati per prendere queste misure. Queste ricostruzioni cellulari sono computazionalmente intensive e l'accesso a risorse computazionali ad alto throughput è raccomandato, anche se non è necessario. Questo metodo è stato applicato con successo a più specie batteriche e mutanti, modalità di imaging fluorescente e produttori di microscopi.
Introduzione
Le celle di tutti i tipi regolano le loro forme per funzioni specifiche. Per esempio, i neuroni sono modellati in modo diverso rispetto alle cellule del sangue e hanno funzioni diverse. Allo stesso modo, le cellule batteriche sono disponibili in una varietà di forme e dimensioni, anche se lo scopo di queste forme non è sempre noto1,2. Pertanto, è importante che la forma delle cellule batteriche sia determinata con precisione. Il metodo delineato mostra un modo facilmente implementato per raccogliere dati adatti per l'analisi 3D della maggior parte delle cellule batteriche vive o fisse.
Il metodo descritto consente di scattare immagini 3D di cellule batteriche per rappresentare con precisione la forma delle cellule 3D del campione e localizzare con precisione le proteine all'interno di queste forme. Le tecniche di microscopia tradizionali prendono immagini 2D, il che è problematico quando si studiano cellule che hanno forme anormali o non simmetriche, come mutanti di Escherichia coli, o batteri curvi come Vibrio cholerae e Helicobacter pylori. Mentre le immagini 3D ad alta risoluzione sono l'input chiave di questo metodo, il metodo non restituisce un'immagine con risoluzione avanzata. Piuttosto, questo metodo ricostruisce le coordinate della superficie 3D e la forma della cella utilizzando un algoritmo di convoluzione in avanti utilizzando contorni attivi e l'apparente funzione di sfocatura del microscopio3 (Figura 1). E 'stato utilizzato per studiare l'atto batterico omolog MreB in E. coli4,5,6,7, il nuovo elemento periskeletale CrvA in V. colerae8, e il putativo bactofilin CcmA in Helicobacter pylori9 (Figura 2).
La localizzazione delle proteine può dare una visione delle loro funzioni. Ad esempio, le proteine coinvolte nella divisione cellulare sono normalmente localizzate alla cella media10,11. Sono stati intrapresi studi ad alto contenuto di velocità sono stati intrapresi per localizzare tutte le proteine di un batterio nella speranza di ottenere informazioni sulle loro funzioni12. Sfortunatamente, questi studi sono stati eseguiti con imaging 2D e analisi 1D o 2D, rendendo impossibile misurare aspetti specifici della localizzazione delle proteine, come la localizzazione alle caratteristiche geometriche cellulari.
Ad esempio, MreB, una proteina dinamica necessaria per la forma dell'asta di molti batteri, è ipotizzata per indirizzare la localizzazione della sintesi della parete cellulare, e la sua localizzazione rispecchia la localizzazione della sintesi della parete cellulare7,13. MreB da più specie mostra arricchimento geometrico4,6,7,14,15. I polimeri di superficie dinamici, come MreB, possono accoppiare la geometria della superficie al profilo di arricchimento mediotempo 16 e possono essere in grado di orientarsi a geometrie specifiche minimizzando l'energia associata al legame alla membrana17. Mentre l'importanza della torsione, dell'aggregazione, della piegatura e della dinamica non è stata completamente risolta per MreB, è importante notare che misurazioni accurate di entrambe le curvature principali di una superficie richiedono una rappresentazione 3D completa della cella. Pertanto, per misurare in modo più accurato le curve a cui le proteine si localizzano, è preferibile utilizzare l'imaging 3D, piuttosto che 2D. L'imaging 3D elimina la necessità di stimare computazionalmente quelle curvature che non possono essere misurate in 2D, una stima che potrebbe non essere accurata nelle cellule asimmetriche18.
Sebbene l'imaging 2D delle cellule sia più rapido e non richieda tanto lavoro computazionale di postimaging, l'imaging 3D fornisce una rappresentazione più accurata della cella, nonché la capacità di misurare le caratteristiche superficiali, come la curvatura, che non può essere misurata in 2D. Pertanto, man mano che l'imaging 3D diventa più comune, nuove intuizioni sulla forma delle cellule e sulla localizzazione delle proteine diventeranno possibili.
Protocollo
1. Preparazione del campione
- Rendere E. coli fluorescente per l'imaging sia progettandoli per esprimere una proteina fluorescente citoplasmica6 o utilizzando un tinrito a membrana5. Altre specie batteriche possono essere utilizzate al posto di E. coli.
- Trasformare le cellule tramite elettroporazione con un plasmide che codifica la proteina citoplasmica mCherry fluorescente.
NOT: Possono essere utilizzate diverse proteine fluorescenti di altri colori di. - Coltivare i trasformatori su una piastra LB con l'antibiotico appropriato a 37 gradi centigradi. Ottenere colonie singole e identificare cloni positivi mediante microscopia. Le colonie resistenti agli antibiotici che appaiono rosse al microscopio contengono il plasmide che trasporta mCherry.
- Trasformare le cellule tramite elettroporazione con un plasmide che codifica la proteina citoplasmica mCherry fluorescente.
- Coltivare 2 mL della coltura notturna in supporti liquidi LB con gli antibiotici appropriati a 37 gradi centigradi in un'incubatrice tremante. Utilizzare una colonia da una piastra o una piccola quantità di uno stock di congelatore come inoculum.
NOT: Utilizzare i supporti necessari per le cellule batteriche selezionate per l'esperimento. - Sottocoltura la coltura notturna 1:1,000 in 5 mL di supporti LB freschi e lasciare che le cellule crescano a una fase esponenziale a 37 gradi centigradi in un incubatore tremante per 3-4 h. Misurare l'OD600 delle cellule per confermare che sono in fase esponenziale (cioè. , OD600 x 0,2-0,4).
NOT: L'imaging può essere eseguito in qualsiasi fase di crescita a seconda dell'esperimento eseguito.
2. Preparazione del vetrino
NOT: È importante utilizzare supporti con bassa autofluorescenza. È possibile utilizzare qualsiasi mezzo con bassa autofluorescenza. In questo esperimento, l'1% di agarose M63 minimal media pads e sigillamento con VaLaP19 è necessario per l'immagine delle cellule in 3D.
-
Preparare una soluzione dell'1% di agarose in 20 mL di supporti minimi.
- Microonde la soluzione fino a quando l'agarose è completamente dissolta e la soluzione appare chiara.
- Mantenere la soluzione in un bagno d'acqua a 60 gradi fino a quando non è pronto per l'uso.
NOT: Questa soluzione può essere solidificata e fusa in base alle esigenze o può essere suddivisa in quantità minori che vengono fuse in base alle esigenze. Dopo più usi il supporto può diventare scolorito e deve essere sostituito.
- Sciogliere il sigillante VaLaP su una piastra calda a 80-100 gradi centigradi.
NOT: Prepara VaLaP, per essere usato come sigillante, fondendo 50 g di gelatina di petrolio, lanolina e paraffina insieme in un bicchiere su una piastra calda a 80-100 gradi centigradi. Questa grande quantità di VaLaP può essere conservata a temperatura ambiente a tempo indeterminato e sarà sufficiente per >500 diapositive. Riscaldamento a > 100 gradi c farà sì che si degrada e il suo colore si scurirà. - Posizionare due pile ciascuna delle tre schegge di copertura da 20 mm x 20 mm sulle estremità opposte di una diapositiva (sei appendici totali).
- Pipette 200 - L di agarose pad soluzione sul vetrino.
NOT: Se non si utilizza una macchia fluorescente ingegnerizzata, un coloranti a membrana può essere aggiunto alla soluzione agarose pad prima di questo passaggio. Risospendere il tinrito in acqua e aggiungere la tintura a 1 mL della soluzione agarose pad ad una concentrazione finale di 5 g/mL. - Immediatamente e saldamente posizionare un secondo scivolo verso il basso sulla pila di scivolamenti di copertura per appiattire l'agar e lasciarlo solidificare per 1 min a temperatura ambiente.
- Scivolare con attenzione fuori dalla diapositiva superiore.
- Utilizzare l'estremità grande di una punta di pipetta da 200 litri per ritagliare le singole pastiglie dal gel (5 mm di diametro) sul vetrino e scartare il gel malformato o non necessario.
NOT: Se si immaginano più ceppi o condizioni, sarà necessario un pad separato per ciascuno di essi. Se si vuole che venga imageta una sola deformazione, fare delle pastiglie da 3/4 per supportare la copertina. - Cellule pipettrici su e giù più volte per interrompere i grumi di cellule e garantire la coltura è ben mescolata. Pipetta 1 - L di sottocultura dal gradino 1,3 su un pad.
NOT: Le ricostruzioni della forma cellulare richiedono singole celle che non toccano altre celle. Se si utilizzano colture di fase stazionaria o ad alta densità cellulare, potrebbe essere necessario diluire il campione 1:10 prima di posizionarlo sul pad. - Lasciare asciugare l'aria del campione per un valore di 5/10 min. Assicurarsi che la goccia sia completamente assorbita nel pad. Se rimane del liquido, le cellule si muoveranno nel liquido e non possono essere imageggiate.
- Posizionare un coperchio sulla parte superiore delle pastiglie.
- Sigillare lo slip di copertura con VaLaP fuso spazzolando delicatamente intorno al bordo dello scivolo di copertura con un tampone di cotone. Assicurarsi di tenerlo lontano dalla parte superiore della ricevuta di copertura dove l'obiettivo toccherà. Il VaLaP si indurisce in pochi secondi, sigillando il campione.
- Immediatamente immagini del campione.
NOT: Il campione deve essere immagine non appena la diapositiva è preparata. Le cellule possono crescere sulle pastiglie, e se dividono le ricostruzioni sarà più difficile.
3. Requisiti di imaging
- Assicurarsi che l'asse z o di messa a fuoco del microscopio possa effettuare movimenti precisi inferiori a 50 nm. Utilizzare z piezo stadi (vedi Tabella dei materiali), disponibili sui microscopi di livello di ricerca, perché i tipici dispositivi di messa a fuoco motorizzati non sono in grado di fornire questa precisione.
NOT: Supponendo che il criterio di campionamento Nyquist-Shannon20, è necessario avere la capacità di spostare 0,5 volte la dimensione del passo più piccola desiderata o 2 volte la frequenza spaziale desiderata. Per i 100 nm necessari in questo protocollo, è necessaria una fase con una precisione di 50 nm o meno. -
Assicurarsi che il microscopio contenga un obiettivo 100x con un'apertura numerica minima (NA) di 1,45.
- Raccogliere z-stack di celle diverse da 200-400 per garantire che vengano ottenute abbastanza celle per le applicazioni a valle.
NOT: Il numero di cellule necessarie dipende dalla variabilità sottostante del campione di interesse. Alcune delle cellule raccolte a questo punto non ce la faranno attraverso i passaggi di ricostruzione.
- Raccogliere z-stack di celle diverse da 200-400 per garantire che vengano ottenute abbastanza celle per le applicazioni a valle.
- Assicurarsi che lo z-stack corrisponda alle impostazioni utilizzate per il canale della forma se viene misurato un canale fluorescente secondario (proteina, etichetta metabolica, ecc.).
4. Imaging
- Inserire il vetrino sigillato sul microscopio e lasciarlo seduto per 5 min per ecloccare la temperatura con l'ambiente circostante, perché la stanza del microscopio può essere a una temperatura diversa rispetto alla stanza di preparazione del campione.
-
Prendete uno z-stack fluorescente del campione.
NOTA: lo z-stack dovrebbe coprire interamente il campione con una spaziatura z inferiore alla profondità di campo. Per un obiettivo di 1,45 NA 100x e cellule E. coli spesse 1 m, 40 passi a 100 nm per passo funzionano bene. Per le cellule più grandi o le cellule che non si trovano perfettamente piatte sulla superficie, 50 o più passi possono essere necessari. Includi passaggi sufficienti e assicurati che il campione sia completamente sfocato sopra e sotto.- Utilizzare il software associato al microscopio (vedere Tabella dei materiali) per controllare il microscopio.
- Concentrarsi al centro della cellula utilizzando le ruote di messa a fuoco al microscopio. In Acquisizione ND selezionare la casella di controllo per eseguire una z-stack. Fare clic sul pulsante Home per impostare il centro della cella come punto iniziale. Impostare la dimensione del passo su 0,1 m e impostare l'Intervallo su 4 m. Assicurarsi che il dispositivo sia impostato sullo stadio piezo.
- Impostare i canali fluorescenti sotto la finestra Lambda per le impostazioni per le molecole fluorescenti che si indagano. In questo esperimento sono stati utilizzati GFP e mCherry.
NOT: Prendere uno z-stack aggiuntivo con la stessa dimensione passo e gamma nel secondo canale di colore se si desidera la distribuzione 3D di un canale fluorescente aggiuntivo. In questo esperimento, mCherry citoplasmatico è stato utilizzato per determinare la forma delle cellule e MreB-GFP è stato utilizzato come secondo canale di colore21. - Assicurati che l'ordine dell'esperimento sia impostato su lambda (serie z) in modo che adotti uno stack z completo in ogni canale di colore prima di passare.
- Fare clic su Esegui ora per avviare l'acquisizione dell'immagine e salvare il file uno o entrambi gli stack z sono stati completati.
- Spostarsi in una nuova area del pad e ripetere i passaggi 4.2.2-4.2.5.
5. Ricostruzione delle cellule
- Ritaglia singole celle e salva le immagini come file tiff impilato in modo che vi sia una sola cella per file. Assicurarsi che questa cella sia ben isolata da qualsiasi altra cella (cioè, circa 5 volte la metà massima intera della funzione di sfocatura in xy.
NOTA: questa operazione può essere eseguita utilizzando un software di analisi delle immagini disponibile gratuitamente (vedere Tabella dei materiali).- Disegnare un riquadro intorno a una singola cella e duplicare tale cella 2x, una volta per ogni canale. Assicurarsi che la casella hyperstack duplicata sia selezionata e modificare il canale su 1 o 2, assicurandosi che le sezioni includano l'intero z-stack.
NOT: Se si immagina solo la forma delle cellule e non una proteina fluorescente aggiuntiva, sarà presente un solo canale. - Una volta che entrambe le pile sono disponibili andare a Immagini Proprietà Stacks . Proprietà Tools (Strumenti) Concatenare per combinare le immagini con il canale proteico prima e il canale di forma secondo.
- Salvare la nuova immagine come file tiff.
- Disegnare un riquadro intorno a una singola cella e duplicare tale cella 2x, una volta per ogni canale. Assicurarsi che la casella hyperstack duplicata sia selezionata e modificare il canale su 1 o 2, assicurandosi che le sezioni includano l'intero z-stack.
- Misurare la funzione di sfocatura del microscopio utilizzando sottodiffrazione limitato perline fluorescenti22. Questo deve essere fatto per ogni obiettivo al microscopio e al microscopio, ma può essere eseguito prima o dopo l'imaging dei campioni di interesse.
- Media insieme più perline indipendenti con qualche intervento manuale utilizzando il software disponibile (vedi Tabella dei materiali).
NOT: Il prodotto finale dovrebbe essere un'immagine 3D della funzione di sfocatura con la stessa spaziatura xyz dei campioni di interesse.
- Media insieme più perline indipendenti con qualche intervento manuale utilizzando il software disponibile (vedi Tabella dei materiali).
- Eseguire gli script di ricostruzione della forma della cella di convoluzione in avanti utilizzando il software disponibile. L'ultima versione di questi script può essere scaricata liberamente dal https://github.com/PrincetonUniversity/shae-cellshape-public.
- Creare una cartella all'interno di una cartella sul desktop contenente le immagini ritagliate e il file cell_shape_settings_tri.txt da shae-cellshape-public/exampleData_tri.
- Modificate cell_shape_setting_tri.txt per avere le impostazioni corrette per l'esperimento di interesse. Per questo esperimento, il file di impostazioni include le seguenti righe:
nm_per_pixel 70
Scala 0,65
stack_z_dimensione 41
stack_t_dimensione 1
Fstack_z_dimensione 41
Fstack_t_size 1
stack_seperation_nm 100
Fstack_seperation_nm 100
psfScript -999 osuPSF20180726
gradiente 1
NOT: Questo file di testo è organizzato in sezioni e ogni sezione viene analizzata nella propria variabile. Anche se molte delle impostazioni non devono essere modificate da esperimento a esperimento, è necessario assicurarsi che le dimensioni dello z-stack (stack_z_size per la forma, Fstack_z_size per le proteine di interesse), la spaziatura dello z-stack (stack_seperation_ nm, Fstack_seperation_nm), spostamento focale relativo del microscopio () ,23, la dimensione in pixel in xy (nm_per_pixel) e il nome dello script che carica la funzione di sfocatura del microscopio (psfScript ) ) corrisponde all'esperimento. Il campo sfumatura deve essere impostato su 1 se il canale della forma è riempito citoplasmaticamente e 0 se si tratta di un oggetto macchiato di membrana. - Eseguire Cell_shape_detector3dConvTriFolder (shae-cellshape-public/CellShapeDetectorTri/) con la stringa nel percorso della cartella seguito dal numero della cella su cui iniziare e dal numero di celle da eseguire.
NOT: Un esempio per l'input sarà il seguente: Cell_shape_detector3dConvTriFolder ('percorso a cartella con immagini ritagliate', indice iniziale nella cartella, numero di celle). Una cellula tipica può richiedere tra 5 e 20 min perché la ricostruzione convergerà e finisca.
- Eseguire lo screening dei ricostruzioni delle celle per assicurarsi che siano corrette prima di utilizzare le celle per qualsiasi analisi statistica.
- Eseguire ScreenFits (shae-cellshape-public/shae-fitViewerGui/) per eseguire visivamente lo screening delle ricostruzioni di singole celle.
- Fare clic sul pulsante Seleziona cartella all'apertura dell'interfaccia utente grafica (GUI), quindi selezionare la cartella con i file di dati di ricostruzione (TRI.mat) creati nel passaggio 5.3.
- Selezionare la casella accanto alla ricostruzione della cella se una cella appare deforme o non converge completamente (Figura 3). Questo potrebbe sembrare una cella con un buco, un lato piatto o un ramo che ne esce. In questo modo 'FLAG' verrà aggiunto al nome del file in modo che possa essere escluso da qualsiasi analisi a valle.
NOT: La cella ricostruita può essere confrontata con le immagini originali. Questo può essere particolarmente importante se le vostre cellule provengono da una popolazione eterogina di varie forme.
- Eseguire enrichmentSmoothingSpline (shae-cellshape-public/) per creare un profilo di arricchimento della concentrazione relativa della proteina di interesse in funzione della curvatura gaussiana in superficie.
- Selezionare ogni cartella con i file TRI.mat creati nel passaggio 5.3.3.
- Selezionare il file curve.mat appena creato (5.5.1).
NOT: Verrà creato un file curve.mat per ogni cartella selezionata nella 5.5.1. Tutti i profili di arricchimento saranno presentati in un unico grafico. Se sono necessari singoli grafici rispetto all'esecuzione 5.5 per ogni cartella.
NOT: Oltre alla precisa localizzazione geometrica di una proteina fluorescente, ci sono molti altri modi per analizzare i dati dal canale secondario, incluso il conteggio del numero, delle dimensioni e dell'orientamento degli oggetti4 .
Risultati
I batteri sono disponibili in un'ampia varietà di forme e dimensioni che possono determinare le loro funzioni in natura1. Il risultato di questa procedura è una rappresentazione 3D accurata delle celle dalla convoluzione in avanti di uno z-stack di immagini (Figura 1). Questo metodo è particolarmente importante quando si tratta di celle curve (Figura 2) o con celle a forma di anormalità (Figura 4A), poiché una rappresentazione 2D non riflette accuratamente la curvatura delle celle. Per utilizzare il metodo di convoluzione in avanti (Figura 1A), le celle devono essere macchiate perifericamente o avere una macchia citoplasmatica (Figura 2B sinistra contro destra).
Figura 4 Mostra la localizzazione MreB nella cella. Una fusione GFP è stata fatta alla proteina actina batterica MreB21 al fine di studiare la sua precisa localizzazione sia nel tipo selvatico che nelle cellule mutanti E. coli 4. Poiché MreB è associato alla membrana, l'imaging 3D è necessario per riprodurre fedelmente la sua posizione nella cellula. Facendo queste misurazioni in 3D, siamo stati in grado di ricostruire le forme di entrambi i tipi selvatici e le cellule mutanti rod (Figura 4A). È stato dimostrato che la localizzazione di MreB è stata arricchita a piccole curve gaussiane, una caratteristica geometrica che può essere misurata solo in 3D, in modo dipendente da Rod (Figura 4B).

Figura 1: Metodo di convoluzione in avanti ricostruisce le cellule senza alcuna conoscenza preliminare della forma delle cellule. (A) L'output finale del metodo è una ricostruzione cellulare 3D derivata confrontando una forma di prova con la funzione di sfocatura calibrata del microscopio. Questo viene confrontato con lo z-stack osservato fino a quando l'immagine 3D osservata corrisponde a quella ipotetica. L'immagine qui mostrata è un rendering della ricostruzione di una cellula C. crescentus. (B) La pipeline di ricostruzione aggiorna in modo iterativo le posizioni stimate degli elementi di superficie in base alla pila di immagini osservata. Per i dettagli algoritmici completi del metodo, vedere Nguyen3. (C) La pipeline di ricostruzione inizia senza una conoscenza a priori della forma della cella e aggiorna la posizione e il numero dei vertici della superficie in modo che corrispondano allo stack z osservato. Le immagini rappresentative da ogni 30 passi durante la ricostruzione 3D sono mostrate da tre diverse angolazioni di una cellula V. colera. Le posizioni delle superfici di questa forma vengono aggiornate per ridurre al minimo la differenza tra le pile simulate e osservate. Fare clic qui per visualizzare una versione più grande di questa figura.
{kind=link}

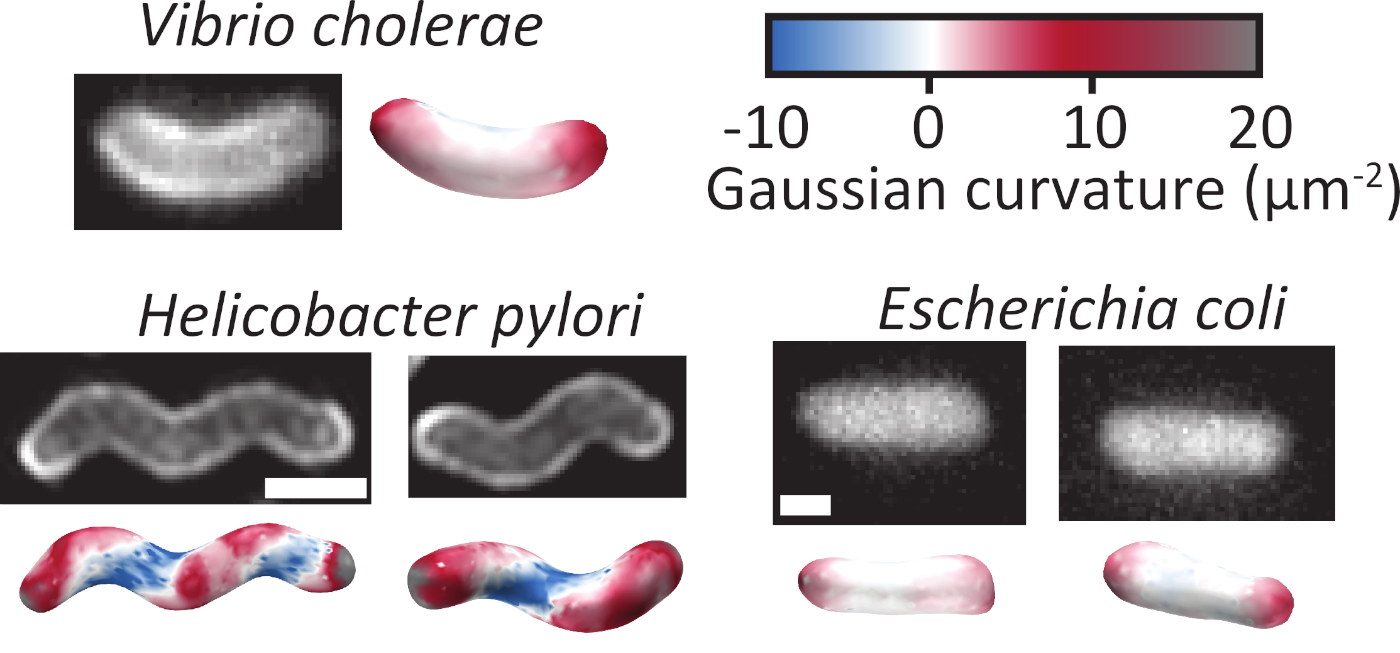
Figura 2: Rappresentazione ricostruzioni 3D di tre specie batteriche di forma diversa. Ricostruzioni tridimensionali possono essere fatte da cellule con la loro membrana macchiata (a sinistra) o riempito con un fluoroforo citoplasmico (a destra). La forma e la curvatura della cella iniziale non è importante, in quanto un'asta piegata, un'asta torsione o un'asta dritta sono tutti in grado di essere ricostruiti con precisione. Le superfici ricostruite sono colorate in base alla curvatura gaussiana locale della superficie. Barra di scala: 1 m. Fare clic qui per visualizzare una versione più grande di questa figura.
{kind=link}

Figura 3: I riricostruzioni possono avere esito negativo per diversi motivi. Le celle devono essere sottoposte a screening per garantire che l'algoritmo di ricostruzione conversi a un risultato ragionevole. Sinistra: tipo selvatico rappresentativo (in alto) e asta mutante (in basso) cellule E. coli che hanno superato il controllo di qualità. Destra: celle che non sono riuscite a ricostruire correttamente e non hanno superato il controllo qualità. Vengono mostrate cinque classi di convergenza non riuscita: (i) cellule che sono troppo vicine al bordo della regione di ritaglio che portano ad una marcata demarcazione, (ii) cellule che sono troppo vicine a un'altra cellula, (iii) cellule che hanno prodotto un divot, (iv) cellule che non hanno proceduto oltre il s iniziale s tatto e (v) altri errori sconosciuti. Fare clic qui per visualizzare una versione più grande di questa figura.
{kind=link}

Figura 4: Dati rappresentativi che mostrano la localizzazione delle proteine a geometrie cellulari specifiche. (A) Sono visualizzate cellule tridimensionali ricostruite di tipo selvaggio e cellule di rod-coli con curvatura gaussiana e intensità di fluorescenza MreB. Barra della scala (B) Grafici di arricchimento di MreB da cellule di tipo selvaggio e di rod-E. coli. I valori >1 mostrano l'arricchimento e i valori <1 mostrano l'esaurimento di MreB da queste regioni cellulari rispetto alla copertura uniforme della cella. Le aree ombreggiate indicano un intervallo di confidenza bootstrap medio-90% della media. La curva per ogni deformazione è una spline di levigatura cubica e viene troncata utilizzando una soglia di probabilità per curve estreme di p > 5 x 10-3. Questa cifra è stata modificata da (Bratton et al.) 4. Fare clic qui per visualizzare una versione più grande di questa figura.
{kind=link}
Discussione
Un passaggio critico in questo protocollo è l'acquisizione di immagini di alta qualità. Per ricostruire correttamente le cellule, ci deve essere abbastanza sfocatura sopra e sotto la cellula. Pertanto, è imperativo che lo z-stack preso copre una distanza sufficientemente grande. Il numero di passi effettuati durante l'acquisizione dell'immagine può essere regolato per ogni deformazione. Ad esempio, le cellule E. coli cancellate per l'asta sono più larghe e richiedono più passaggi e, di conseguenza, una distanza maggiore rispetto alle cellule di tipo selvatico. Se il campione si sposta durante l'acquisizione dell'immagine, la ricostruzione può avere errori gravi. Pertanto, è importante lasciare che la diapositiva arrivi all'equilibrio termico con il microscopio prima dell'imaging per evitare deriva durante l'acquisizione dello z-stack. Le celle devono essere visualizzate su rilievi con bassa autofluorescenza. I componenti multimediali, come quelli che si trovano nel comune supporto LB, hanno autofluorescenza che può causare problemi quando si tenta di ricostruire le cellule. La densità delle cellule sul pad di imaging è importante perché il processo di ricostruzione viene eseguito in modo indipendente su ogni cellula. Troppo poche cellule aumenteranno il tempo necessario per ottenere immagini di un numero sufficiente di cellule, mentre troppe cellule si tradurranno in campi di imaging troppo densi per ritagliare facilmente singole cellule. Poiché non tutte le cellule vengono ricostruite correttamente, le celle aggiuntive devono essere visualizzate durante la fase di acquisizione e tutte le uscite devono essere esaminate prima di procedere con l'analisi statistica (Figura 3).
Molte delle limitazioni per questo metodo sono tecniche. Sul microscopio da utilizzare, si deve avere un obiettivo che ha un'alta apertura numerica (in genere >1.4), perché questo consente la sezionamento ottico sulla scala delle dimensioni dei batteri. Inoltre, il microscopio deve essere dotato di uno stadio piezo che può fare piccoli e precisi passi nella direzione z. Inoltre, anche se non è necessario, l'accesso alle risorse di calcolo ad alta velocità effettiva per eseguire il software di analisi delle immagini è altamente raccomandato perché ridurrà il tempo di elaborazione per ricostruire le celle.
Una limitazione concettuale al metodo è che devono essere scelte le scale di energia corrette per ponderare la levigatezza della ricostruzione rispetto al rapporto segnale-rumore delle immagini. Per convalidare una scelta di parametri, le dimensioni e le forme delle cellule devono essere misurate utilizzando metodi indipendenti come la microscopia elettronica a trasmissione (TEM) o la microscopia a forza atomica (AFM). Come prova di principio, le ricostruzioni 3D delle celle sono state eseguite su un'AFM per verificare la precisione z (<50 nm) o su una griglia TEM per verificare la precisione xy (<30 nm)3. Un approccio così correlato richiede molto tempo e denaro. Un approccio più semplice può essere quello di immagini di campioni standard come celle di tipo selvaggio o 1 m di perline sferiche. Il diametro e la sferità delle ricostruzioni possono essere utilizzati per garantire che le dimensioni e le scale energetiche utilizzate nella ricostruzione siano corrette.
Questo non è l'unico metodo che cerca di estrarre informazioni spaziali ad alta risoluzione dalle immagini di microscopia a fluorescenza. Molti articoli di revisione descrivono i recenti progressi nel campo della microscopia a super-risoluzione24,25. Tecniche di miglioramento della risoluzione come la microscopia di deconvoluzione26, la microscopia confocale del disco rotante27, la riassegnazione dei pixel28e la microscopia ad illuminazione strutturata (SIM)29 cercano di migliorare la risoluzione del immagini acquisite dal microscopio. Questi metodi non sono incompatibili con l'approccio presentato. Recentemente questo metodo è stato adattato per consentire immagini basate su SIM come ingressi9. Mentre il metodo di convoluzione in avanti condivide alcune delle sue basi con la microscopia di deconvoluzione, ha un output completamente diverso. Mentre approcci come la microscopia di deconvoluzione cercano di migliorare la risoluzione dell'immagine, questo approccio non genera un'immagine, ma piuttosto una ricostruzione della forma cellulare con una precisione di circa 50 nm. Tecniche di microscopia a controllo attivo a singola molecola basate su campioni scarsamente etichettati possono fornire livelli ancora più elevati di precisione spaziale rispetto a questo metodo. In molti casi, questi approcci a singola molecola richiedono l'ottimizzazione dei costrutti fluorescenti e possono richiedere lunghi tempi di acquisizione, rendendoli difficili da usare con campioni vivi o dinamici. Ognuno di questi metodi viene fornito con uno o più avvertimenti che questo metodo non. Ad esempio, i benefici pubblicizzati dalla microscopia confocale del disco rotante non sono applicabili ai monostrati di cellule batteriche, dove non c'è molta luce fuori dalla luce del piano. Inoltre, questo metodo fornisce una pipeline per acquisire forme cellulari 3D accurate e localizzazione delle proteine senza la necessità di fluorofori specializzati. Questo metodo ha requisiti hardware minimi (ad esempio, z piezo, obiettivo NA elevato) e richiede solo decine di immagini per punto temporale, consentendo di esaminare facilmente le strutture 3D dinamiche6 .
Ci sono stati un numero crescente di approcci per studiare l'organizzazione delle cellule batteriche in strutture 3D. Questi includono approcci concettualmente simili a questo che sfruttano le immagini di alta qualità, a fluorescenza 3D30,31,32. Questo approccio richiede cellule ben isolate e non fa supposizioni a priori sulla geometria cellulare. Tuttavia, per passare ad aggregati cellulari densi o biofilm, si presume che le cellule siano simili ad aste. Questa visione a bassa risoluzione consente ancora di studiare le disposizioni di imballaggio delle cellule, anche se l'alta densità di cellule nel biofilm impedisce l'analisi della localizzazione subcellulare di fattori specifici.
In futuro, potrebbe essere interessante sviluppare un quadro per integrare la singola molecola e gli approcci ad ampio campo con questa tecnica di ricostruzione 3D. Inoltre, potrebbe essere possibile includere questo approccio di convoluzione in avanti con gli strumenti di segmentazione della visione artificiale32 per consentire ricostruzioni di ammassi cellulari più densi.
Perché le cellule si sono evolute in forme specifiche è una questione complessa che deve riflettere l'ambiente complesso in cui vivono. Comprendere l'evoluzione e la funzione delle forme cellulari richiede di prendere misure precise e accurate di quelle forme, che è ciò che questo metodo fornisce.
Divulgazioni
Gli autori non hanno nulla da rivelare.
Riconoscimenti
Ringraziamo Jeffrey Nguyen e Joshua Shaevitz per aver contribuito a sviluppare questo metodo.
Finanziamento: RMM - NIH F32 GM103290-01A1, BPB - Glenn Centers for Aging Research and National Science Foundation PHY-1734030.
Materiali
| Name | Company | Catalog Number | Comments |
| 50 nm fluorescent beads | Invitrogen | F8795 | these are used to measure the blurring function of the microscope |
| Agarose | sigma-Aldrich | A9539 | |
| Cotton Swab | Puritan Medical Products Company LLC | S304659 | used to appy VaLaP |
| Cover Slips | VWR | 16004-302 | |
| Fiji | ImageJ | https://fiji.sc | used to cro cells |
| FM4-64 | Invitrogen | LST3166 | membrane dye used to stain cells |
| Huygens Software | Scientific Volume Imaging | Huygens essential or professional | Use to measure blurring function of microscope |
| Lanolin | Sigma-Aldrich | L7387 | combine with paraffin and petroleum jelly to make VaLaP |
| LB growth medium | BD Difco | DF0446173 | |
| M63 medium | US Biological | M1015 | |
| MATLAB | Mathworks | Needed to run forward convolution scripts | |
| Microscope Slides | Fisher | 12-550-133 | |
| NIS Elements | Nkon | ||
| Paraffin | Sigma-Aldrich | 327212 | |
| Petroleum Jelly | Equate | 49035-038-27 | |
| Piezo z stage | Nikon | 77011589 | |
| Pipet tips -p200 | USA Scientific | 1111-0730 | |
| Pipet tips- p10 | USA Scientific | 1111-3730 |
Riferimenti
- Young, K. D. The Selective Value of Bacterial Shape. Microbiology and Molecular Biology Reviews. 70 (3), 660-703 (2006).
- Persat, A., Stone, H. A., Gitai, Z. The Curved Shape of Caulobacter crescentus Enhances Surface Colonization in Flow. Nature Communications. 5, 3824(2014).
- Nguyen, J. P., Bratton, B. P., Shaevitz, J. W. Bacterial Cell Wall Homeostasis: Methods and Protocols. Hong, H. J. , Springer. New York. 227-245 (2016).
- Bratton, B. P., Shaevitz, J. W., Gitai, Z., Morgenstein, R. M. MreB Polymers and Curvature Localization are Enhanced by RodZ and Predict E. coli's Cylindrical Uniformity. Nature Communications. 9 (1), 2797(2018).
- Ouzounov, N., et al. MreB Orientation Correlates with Cell Diameter in Escherichia coli. Biophysical Journal. 111 (5), 1035-1043 (2016).
- Morgenstein, R. M., et al. RodZ links MreB to Cell Wall Synthesis to Mediate MreB rotation and Robust Morphogenesis. Proceedings of the National Academy of Sciences. , 12510-12515 (2015).
- Ursell, T. S., et al. Rod-Like Bacterial Shape is Maintained by Feedback Between Cell Curvature and Cytoskeletal Localization. Proceeding of the National Academy of Science U.S.A. 111 (11), E1025-E1034 (2014).
- Bartlett, T. M., et al. A Periplasmic Polymer Curves Vibrio cholerae and Promotes Pathogenesis. Cell. 168 (1-2), 172-185 (2017).
- Taylor, J. A., et al. Distinct Cytoskeletal Proteins Define Zones of Enhanced Cell Wall Synthesis in Helicobacter pylori. bioRxiv. , 545517(2019).
- Egan, A. J. F., Vollmer, W. The Physiology of Bacterial Cell Division. Annals of the New York Academy of Science. 1277 (1), 8-28 (2013).
- Adams, D. W., Errington, J. Bacterial Cell Division: Assembly, Maintenance and Disassembly of the Z ring. Nature Reviews Microbiology. 7 (9), 642-653 (2009).
- Werner, J. N., Gitai, Z., Melvin, I. S., Brian, R. C., Alexandrine, C. High-Throughput Screening of Bacterial Protein Localization. Methods in Enzymology. 471, 185-204 (2010).
- de Pedro, M. A., Quintela, J. C., Höltje, J. V., Schwarz, H. Murein Segregation in Escherichia coli. Journal of Bacteriology. 179 (9), 2823-2834 (1997).
- Hussain, S., et al. MreB Filaments Align Along Greatest Principal Membrane Curvature to Orient Cell Wall Synthesis. eLife. 7, e32471(2018).
- Wong, F., et al. Mechanical Strain Sensing Implicated in Cell Shape Recovery in Escherichia coli. Nature Microbiology. 2, 17115(2017).
- Wong, F., Garner, E. C., Amir, A. Mechanics and Dynamics of Translocating MreB Filaments on Curved Membranes. eLife. 8, e40472(2019).
- Quint, D. A., Gopinathan, A., Grason, G. M. Shape Selection of Surface-Bound Helical Filaments: Biopolymers on Curved Membranes. Biophysical Journal. 111 (7), 1575-1585 (2016).
- Colavin, A., Shi, H., Huang, K. C. RodZ Modulates Geometric Localization of the Bacterial Actin MreB to Regulate Cell Shape. Nature Communications. 9 (1), 1280(2018).
- Valap Sealant. , Cold Spring Harbor Protocols. http://cshprotocols.cshlp.org/content/2015/2/pdb.rec082917 (2015).
- Shannon, C. E. Communication in the Presence of Noise. Proceedings of the IRE. 37 (1), 10-21 (1949).
- Billings, G., et al. De Novo Morphogenesis in L-Forms Via Geometric Control of Cell Growth. Mol Microbiol. 93 (5), 883-896 (2014).
- Sturrman, N. Measuring a Point Spread Function. , iBiology. https://www.ibiology.org/talks/measuring-a-point-spread-function (2012).
- Bratton, B. P., Shaevitz, J. W. Simple Experimental Methods for Determining the Apparent Focal Shift in a Microscope System. PLOS ONE. 10 (8), e0134616(2015).
- Moerner, W. E., Shechtman, Y., Wang, Q. Single-molecule Spectroscopy and Imaging over the decades. Faraday Discussions. 184, 9-36 (2015).
- Sahl, S. J., Hell, S. W., Jakobs, S. Fluorescence Nanoscopy in Cell Biology. Nature Reviews Molecular Cell Biology. 18, 685(2017).
- Sarder, P., Nehorai, A. Deconvolution Methods for 3-D Fluorescence Microscopy Images. IEEE Signal Processing Magazine. 23 (3), 32-45 (2006).
- Oreopoulos, J., Berman, R., Browne, M. Methods in Cell Biology. Waters, J. C., Wittman, T. 123, Academic Press. 153-175 (2014).
- Huff, J. The Airyscan detector from ZEISS: confocal imaging with improved signal-to-noise ratio and super-resolution. Nature Methods. 12, 1205(2015).
- Gustafsson, M. G. L. Surpassing the Lateral Resolution Limit by a Factor of Two Using Structured Illumination Microscopy. Journal of Microscopy. 198 (2), 82-87 (2000).
- Lee, M. K., Rai, P., Williams, J., Twieg, R. J., Moerner, W. E. Small-Molecule Labeling of Live Cell Surfaces for Three-Dimensional Super-Resolution Microscopy. Journal of the American Chemical Society. 136 (40), 14003-14006 (2014).
- Yan, J., Sharo, A. G., Stone, H. A., Wingreen, N. S., Bassler, B. L. Vibrio cholerae Biofilm Growth Program and Architecture Revealed by Single-Cell Live Imaging. Proceedings of the National Academy of Sciences. 113 (36), E5337-E5343 (2016).
- Wang, J., et al. Bact-3D: A level set segmentation approach for dense multi-layered 3D bacterial biofilms. 2017 IEEE International Conference on Image Processing (ICIP). , 330-334 (2017).
Ristampe e Autorizzazioni
Richiedi autorizzazione per utilizzare il testo o le figure di questo articolo JoVE
Richiedi AutorizzazioneThis article has been published
Video Coming Soon
Personale delle biblioteche
Copyright © 2025 MyJoVE Corporation. Tutti i diritti riservati