
Method Article
Caratterizzazione funzionale di RING-Type E3 Ubiquitin Ligases In Vitro e In Planta
In questo articolo
Riepilogo
L'obiettivo di questo manoscritto è quello di presentare un profilo per gli studi biochimici e funzionali completi dei legamenti dell'ubiquitina E3 di tipo RING. Questa pipeline multifase, con protocolli dettagliati, convalida un'attività enzimatica della proteina testata e dimostra come collegare l'attività alla funzione.
Abstract
L'ubiquitazione, come modifica post-traduzionale delle proteine, svolge un importante ruolo normativo nell'omeostasi delle cellule eucariotiche. L'attaccamento covalente di 76 modificatori di ubiquitina aminoacido a una proteina bersaglio, a seconda della lunghezza e della topologia della catena di poliufilia, può portare a diversi risultati che vanno dalla degradazione delle proteine ai cambiamenti nella localizzazione e/o nell'attività della proteina modificata. Tre enzimi catalizzano in sequenza il processo di ubiquitina: E1 enzima attivatota l'ubiquitina, enzima coniugante E2 ubiquitina, e E3 ubiquitina ligase. La ligasa dell'ubiquitina E3 determina la specificità del substrato e, pertanto, rappresenta un argomento di studio molto interessante. Qui presentiamo un approccio globale per studiare la relazione tra l'attività enzimatica e la funzione della ligase di ubiquitina e3 di tipo RING. Questo protocollo in quattro fasi descrive 1) come generare un mutante carente di ligase E3 attraverso la mutagenesi mirata al dominio REC. 2–3) come esaminare l'attività di ubiquitazione sia in vitro che in planta; 4) come collegare tali analisi biochimiche al significato biologico della proteina testata. Generazione di un mutante carente di ligase E3 che interagisce ancora con il suo substrato, ma non lo onniquizza più per la degradazione, facilita la sperimentazione delle interazioni enzimatica-substrato in vivo. Inoltre, la mutazione nel dominio RECà spesso conferisce un fenotipo negativo dominante che può essere utilizzato negli studi di knockout funzionali come approccio alternativo a un approccio di interferenza dell'RNA. I nostri metodi sono stati ottimizzati per studiare il ruolo biologico dell'efere di nematode parassita vegetale RHA1B, che dirotta il sistema di ubiquitinazione ospite nelle cellule vegetali per promuovere il parassitismo. Con una leggera modifica del sistema di espressione in vivo, questo protocollo può essere applicato all'analisi di qualsiasi ligase E3 di tipo RING indipendentemente dalle sue origini.
Introduzione
La stragrande maggioranza dei legamenti di ubiquitina E3 appartiene a RING(Really Interesting New Gene)-tipo) -tipo. Il dominio RING-finger è stato originariamente identificato da Freemont et al. 1 e funzionalmente descritto come un dominio che medial'interazioneproteina-proteina 2 . Il dito canonico RING è un tipo speciale di dominio di coordinamento dello zinco definito come una sequenza di consenso di otto Cis (C) e His (H) specificamente distanziati da altri residui di aminoacidi (X), C-X2-C-X9–39-C-X1–3-H-X2–3-C/H-X2-C-X4-48-C-X2-C. Due ioni di 2 o n2 sono stabilizzati dai residui di nucleo C e H attraverso una topologia "cross-brace" univoca con C1/C2 e C/H5/C6 che coordinano i primi ioni n2, mentre C3/H4 e C7/C8 legano il secondo (Figura 1A)3,4. A seconda della presenza di C o H nel quinto sito di coordinamento di N2,sono state definite due sottoclassi canoniche di proteine RING-finger: C3HC4 e C3H2C3 (rispettivamente RING-HC e RING-H2). Poiché il dominio RING della ligase elitazione e3 media l'interazione tra enzimi coniugati E2 e substrati, è stata dimostrata la mutazione di questi residui essenziali di C e H per interrompere l'attività delle ligase5. Sono state descritte altre cinque sottoclassi meno comuni di legamenti RING E3 (RING-v, RING-C2, RING-D, RING-S/T e RING-G)6. I legamenti di ubiquitina e3 di tipo RING possono essere ulteriormente suddivisi in semplici e complessi enzimi E3. La semplice singola sottounità RING E3 lega menti contengono sia il sito di riconoscimento del substrato che il dominio RING di associazione E2. Al contrario, la multisubunit RING-type E3 complesso substrato di reclutamento o media il legame del e2-ubiquitin intermedio al complesso E3. Il dominio RING Lys residue(s) che funge da sito di attacco di ubiquitina primaria per l'auto-ubiquitinazione potrebbe anche essere importante per l'attività di ligase E3.
Non tutte le proteine contenenti RING funzionano come legamenti E3. Pertanto, la previsione bioinformatica del dominio del dito DI RING e la capacità di ubiquitinazione proteica dipendente da E2 devono essere convalidate biochimicamente e collegate al ruolo biologico della proteina testata. Qui, descriviamo un protocollo passo-passo che illustra come rilevare e caratterizzare funzionalmente l'attività enzimatica dei legamenti di ubiquitina E3 di tipo RING, sia in vitro che in planta, attraverso un approccio di mutagenesi site-directed. I risultati rappresentativi di questa pipeline sono indicati per la ligase RHA1B di tipo RING. RHA1B è una proteina eflatrice prodotta dalla cisti parassitaria vegetale Globoderaa per sopprimere l'immunità delle piante e manipolare la morfologia delle cellule della radice vegetale. Per proteggersi dall'invasione patogena/parassita, le piante hanno sviluppato recettori immunitari di tipo di tipo "NB-LRR" che si prospettano recettori immunitari di tipo "gruppo" di nucleotidi e alle colonne sensibili che rilevano il sito di infezione per arrestare la colonizzazione dei patogeni. Uno di questi recettori immunitari è la proteina Gpa2 di patate che conferisce resistenza ad alcuni isolati di G. pallida (popolazioni di campo D383 e D372)7.
Utilizzando i protocolli presentati, è stato recentemente scoperto che RHA1B interferisce con la segnalazione del sistema immunitario delle piante in modo dipendente da E3 prendendo di mira l'immunorecettore Gpa2 della pianta per l'ubiquitinazione e la degradazione8.
Protocollo
1. Mutagenesi diretta al sito (Figura 1)
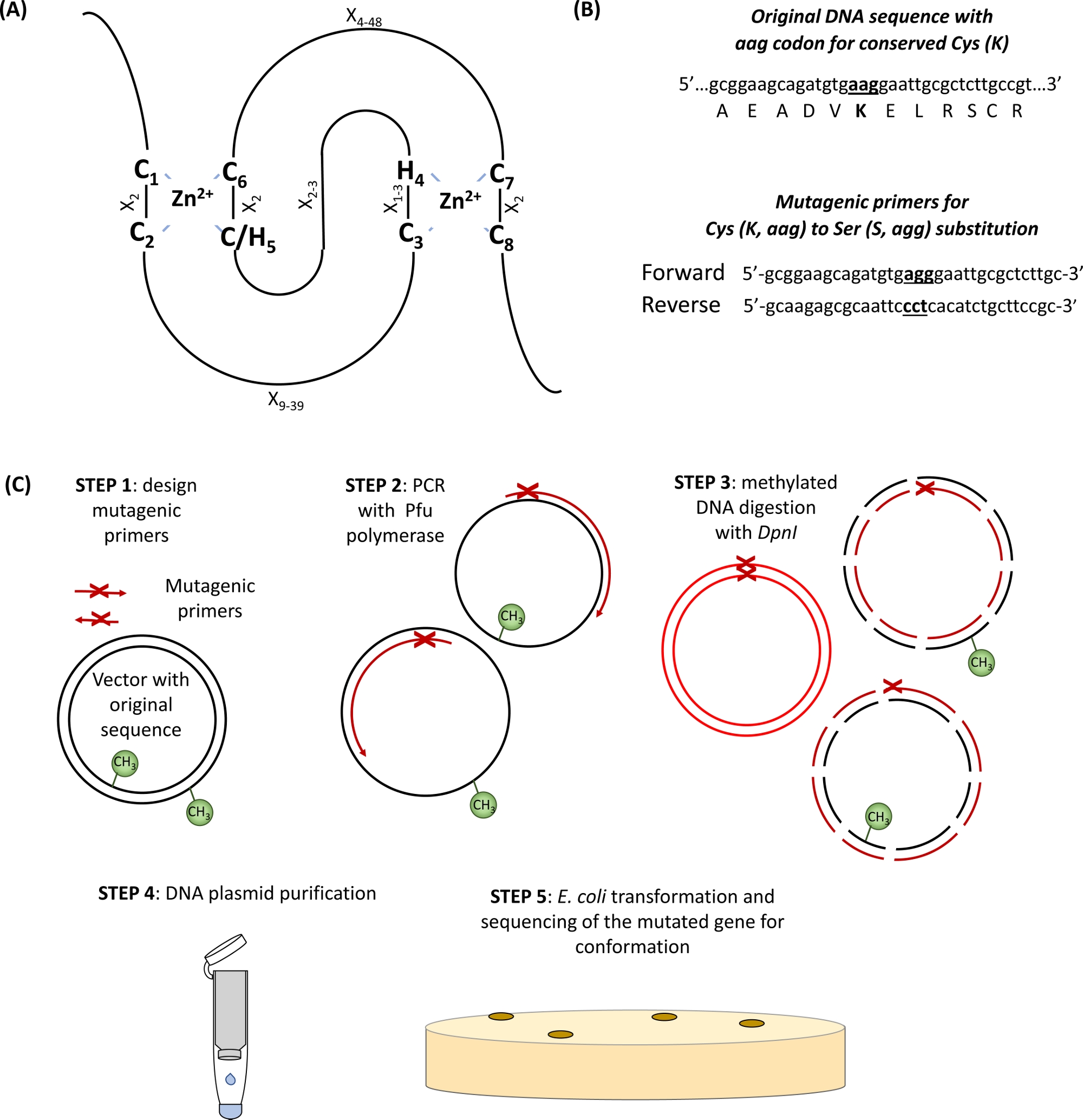
- Identificare i Cis conservati e i suoi aminoacidi nel dominio RING (Figura 1A) e i primer di progettazione che portano il codone di sostituzione degli interessi affiancato da 15 coppie di basi su entrambi i lati del sito di mutazione (Figura 1B).
- Introdurre la mutazione desiderata per amplificazione basata sulla PCR del plasmide che ospita il gene di interesse utilizzando primer mutageni e polimerasi di DNA ad alta fedeltà contenenti Pfu in 50 : L del volume totale di reazione PCR, come mostrato nella Tabella 1 e nella Tabella 2 secondo il protocollo del produttore.
- Digerire ilDNA metilato e semimetilato dei genitori con l'aggiunta di 3 -L dell'enzima di restrizione DpnI direttamente alla reazione PCR (passaggio 1.2) e l'incubazione a 37 gradi centigradi per 2 h.
NOTA: La metilazione è una modifica della proteina posttranziale che viene aggiunta al plasmide prodotto e isolato dai batteri. Nuove copie di plasmide generato da PCR mancano di metilazione, quindi, le nuove copie rimarranno intatte durante il trattamento DpnI. - Purificare i plasmidi mutagenizzati utilizzando un kit di estrazione del DNA commerciale basato sulla tecnologia della colonna di spin ed eluire il DNA con 50 gradi d'acqua.
- Trasformare le cellule chimicamente competenti del DH5, con 0,5 litri del DNA plasmico mutagenizzato recuperato secondo il protocollo del produttore. In breve, incubare celle competenti con DNA sul ghiaccio per 30 min, poi riscaldarle per 20 s a 42 gradi centigradi, e mettere di nuovo i tubi sul ghiaccio per 2 min. incubare le cellule con 500 LL di supporti LB a 37 gradi centigradi per 1 h a 250 rpm e poi distribuirli su polpi selettivi.
- Verificare la mutazione desiderata da Sanger sequenziando i plasmidi di DNA isolati da E. coli.
2. Purificazione delle proteine ricombinanti e analisi dell'ubiquitazione in vitro
- Clonare il tipo selvaggio RING e i geni RING mutati di interesse nel vettore pMAL-c2 (seguire il protocollo del produttore; Tabella n. 3) fondere questi geni con il tag dell'epitopo MBP che permette la purificazione in un solo passaggio usando la resina di amilosio. Introdurre i costrutti risultanti nel ceppo E. coli BL21 come descritto al punto 1.5.
- Coltivare il ceppo E. coli BL21 che ospita il costrutto desiderato in un mezzo liquido da 50 mL a 37 gradi centigradi per 2-3 h fino a raggiungere la fase logaritmica (OD600 di 0,4–0,6).
- Aggiungere IPTG a una concentrazione finale di 0,1-1 mM per indurre l'espressione della proteina ricombinante con etichettatura MBP di interesse e incubare la coltura di E. coli per 2-3 h a 28 gradi centigradi. Posizionare la coltura sul ghiaccio dopo l'incubazione.
NOTA: Eseguire i passi 2.4–2.13 sul ghiaccio per proteggere le proteine dalla degradazione. - Per verificare l'efficienza di induzione, raccogliere 1,5 mL di cellule indotte, ruotarle verso il basso a 13.000 x g per 2 min, rimuovere il supernatante, e risospendere nuovamente le cellule in 20 :L di 2x SDS-PAGE buffer di carico (24 mM Tris-HCl pH 6.8, 0.8% SDS, 10% (v/v) glicerol, 4 mM DTT, 0.04% (w/v) bromopheno blu).
- Far bollire i campioni per 5 min ed eseguirli su un gel 10% SDS-PAGE. Per valutare visivamente l'accumulo di proteina di fusione mbP (peso molecolare della proteina di interesse - 42,5 kDa MBP), macchiare il gel per 20 min agitando con buffer di colorazione Coomassie (50% di metanolo, 10% acido acetico, 0,1% Coomassie blu) e destaining con buffer di destaining (20% metanolo, 10% acido acetico).
- Raccogliere le restanti cellule E. coli per centrifugazione a 1.350 x g per 6 min, eliminare il supernatore e risospendere nuovamente il pellet cellulare con 5 mL di buffer di colonna (20 mM Tris HCl, 200 mM NaCl, 1 mM EDTA, inibitore proteasi batterica).
NOTA: Questo è un buon posto per fermare il protocollo durante la notte. Le cellule congelate possono essere conservate fino a -20 gradi centigradi. - Abbattere le cellule di E. coli posizionando il tubo contenente i batteri in un bagno d'acqua ghiacciata e applicando 10 cicli di sonicazione: la sonicazione di 10 s al 30% dell'amplificatore seguito da 20 s si rompe.
- Centrifuga a 13.000 x g a 4 gradi centigradi per 10 min e salvare il super-natante (estratto di greggio).
- Preparare 500 l di resina di amilosia in un tubo da 15 mL. Lavare la resina aggiungendo 10 mL di buffer a colonna fredda e centrifugando a 1.800 x g, 4 gradi centigradi per 5 min. Fallo 2x.
- Aggiungere 5 mL di estratto grezzo al tubo con la resina di amilosino e incubare pernottando a 4 gradi centigradi.
- Centrifuga a 1.800 x g a 4 gradi centigradi per 5 min e scartare il supernatante.
- Aggiungere 10 mL di buffer di colonna al pellet di resina e incubare per 20 min. Quindi centrifugare a 1.800 x g a 4 gradi centigradi per 5 min. Ripetere questo passaggio 2x.
- Esolute la proteina di fusione con 0,5 mL di cuscinetto di colonna contenente 10 mM di maltosio incubando un campione per 2 h a 4 gradi centigradi. Centrifuga a 1.800 x g a 4 gradi centigradi per 5 min e raccogliere la proteina eluita. Ripetere questo passaggio 2x.
- Dialisci 1 mL della frazione proteica contro il PBS freddo. Proteina artigliante in tubi monouso (10-20 - L) per evitare lo scongelamento e conservare a -80 gradi fino a quando necessario.
- Misurare la concentrazione proteica utilizzando il prosaggio Bradford9.
- Impostare la reazione di ubiquitinazione in vitro in un volume totale di 30 gradi mescolando fino a 40 ng di E1 (ad esempio, AtUBA1), 100 ng di E2 (ad esempio, AtUBC8, SlUBC1/4/6/7/12/13/17/20/22/27/32), 1g di tipo RINGP, 1 g di proteine di tipo RINGP, e 2 g di FLAG-Ub (o HA-Ub) nel buffer di ubiquitinazione (50 mM Tris-HCl pH 7,5, 2 mM ATP, 5 mM MgCl2, 30 mM di creatina fosfato, 50 g/mL di creatina fosforosi). Incubare il composto a 30 gradi centigradi per 2 h.
NOTA: Premake 20x buffer di ubiquitinazione e conservarlo fino a -6 mesi a -20 gradi centigradi in piccoli aliquote per un singolo utilizzo. La creatina fosforokinase perde facilmente la sua attività enzimatica quando il buffer viene scongelato e congelato ripetutamente. - Terminare la reazione mescolando i 30 campioni ll con 7,5 xL di 5x buffer di carico SDS-PAGE (60 mM Tris-HCl pH 6,8, 2% SDS, 25% (v/v) glicerol, 10 mM DTT, 0,1% (w/v) bronofenolo blu) e bollente per 5 min.
- Separare le proteine con il 7,5% di elettroforesi gel SDS-polyacrilimide (SDS-PAGE), quindi trasferire sulla membrana PDVF e rilevare l'ubiquitazione mediante gonfiore occidentale utilizzando l'anti-FLAG (o anti-HA).
- Macchia rescherine la membrana PVDF con Coomassie blue per verificare l'uguale carico della proteina di tipo MBP-RING testata.
3. Agrobacterium- espressione proteica transitoria mediata nelle foglie di Nicotiana benthamiana e nell'assaggio di ubiquitinazione planta
- Streak appropriato Agrobacterium tumefaciens ceppi recanti il gene di interesse epitope (ad esempio, HA-RHA1B, HA-RHA1BC135S, HA-RHA1BK146R, HA-Ub) e vettore vuoto come un controllo sul supporto LB contenente gli antibiotici di selezione appropriati.
- Dopo 2 giorni di crescita a 28 gradi centigradi, raccogliere colonie singole e coltivarle in un mezzo liquido LB con gli antibiotici appropriati a 28 gradi centigradi/250 rpm per altri 24 h.
- Trasferire 100 l di coltura agrobatterica a 3 mL di LB fresco con gli antibiotici appropriati e incubare la coltura per ulteriori 4-6 h a 28 gradi centigradi con rotazione (250 rpm) alla fase di crescita esponenziale tardiva.
- Abbassare le cellule agrobatteriche a 1.800 x g per 6 min, eliminare il supernatante e risospendere le cellule con 3 mL di buffer di lavaggio (50 mM MES pH 5,6, 28 mM di glucosio, 2 mM NaH2PO4). Ripetere questo passaggio 2x.
- Dopo il secondo lavaggio, risospendere le cellule nel buffer di induzione (50 mME MES pH 5,6, 28 mM di glucosio, 2 mM NaH2PO4, 200 acetosyringone, 37 mM NH4Cl, 5 mM MgSO4.7H2O, 4 mM KCl, 18 .M FeSO4.7H2OM. Incubare le cellule con buffer di induzione per un ulteriore 10-12 h a 28 .
NOTA: Acetosyringone induce il trasferimento T-DNA. - Centrifugare le cellule a 1.800 x g per 6 min, scartare il supernatante e risospendere nuovamente le cellule con 2 mL di buffer di infiltrazione (10 mM MES pH 5,5, 200 acetosyringone).
NOTA: Se gli Agrobatteri si aggregano dopo l'incubazione con tampone di induzione, lasciare che le cellule aggregate affondino sul fondo del tubo lasciandolo sul banco per alcuni minuti e trasferire la chiara sospensione dell'Agrobacterium in un nuovo tubo prima di procedere con il passaggio 3.6. - Misurare la concentrazione di batteri utilizzando il valore OD600 (la densità ottica ad assorbimento di 600 nm). Regolare i valori OD600 in base a quelli desiderati.
NOTA: Di solito un valore OD600 compreso tra 0,2–0,4 funziona meglio per una singola espressione di macchia agrobatterica. Se viene applicata una combinazione di diversi ceppi agrobatterici, il totale dei valori di OD600 dei ceppi agrobatterici non deve superare 1. - Agroinfiltrate N. bethamiana di 4 settimane, lascia pungendole delicatamente con un ago, seguita dall'iniezione a mano di Agrobacterium con una siringa senza l'ago. Cerchiare l'area della foglia infiltrata con il marcatore (di solito 1–2 cm di diametro).
- Raccogliere i tessuti foglia infiltrati 36 h post-infiltrazione. Macinare il tessuto in polvere fine con azoto liquido.
- Polvere tissutale risospesa con 300 s l di buffer di estrazione delle proteine (50 mM Tris-HCl pH 7.5, 150 mM NaCl, 5 mM EDTA, 2 mM DTT, 10% glicerolo, 1% polivigliopolilicodolidone, 1 mM PMSF, cocktail inibitore della proteasi vegetale) e centrifugaa a 15.000 x g per 15 min a 4 gradi centigradi.
- Trasferire il supernatante in un nuovo tubo. Aggiungere 5x buffer di caricamento SDS-PAGE ad una concentrazione finale di 1x e far bollire per 5 min.
- Separare le proteine grezze su gel 10% SDS-PAGE, trasferire su membrane PVDF e sondare con anti-HA per rilevare in platani.
4. Stabilire il legame tra attività ezimatica e funzione in planta
NOTA: Ad esempio, RHA1B promuove la degradazione della proteina resistente Gpa2 per sopprimere la morte delle cellule HR. In questo passaggio viene illustrato come verificare che tali attività virulente di RHA1B siano dipendenti da E3.
- Streak appropriato Agrobacterium tumefaciens ceppi che trasportano geni taggati di interesse (in questo esempio HA-RHA1B, HA-RHA1BC135S, HA-RHA1BK146R, myc-Gpa2, RBP1) e vettore vuoto come controllo. Segui i passaggi da 3.1 a 3.8 per la preparazione e l'iniezione dell'Agrobacterium sulle foglie di N. bethamiana.
- Per la degradazione delle proteine del substrato dipendente dall'E3, seguire i passaggi da 3,9 a 3,12 ed eseguire il gonfiore occidentale utilizzando anticorpi appropriati per rilevare l'accumulo di proteine nelle cellule vegetali (ad esempio, anti-HA e anti-MYC).
- Per la risposta ipersensibile dipendente dall'E3 (HR) ha mediato l'inibizione della morte cellulare, monitorare le foglie agroinfiltrate per i sintomi di morte delle cellule HR 2-4 giorni dopo l'infiltrazione.
Risultati
In questa sezione vengono forniti risultati rappresentativi per il protocollo utilizzato per l'esame di una singola sottounità E3 ubiquitin ligase RHA1B che dispone di un dominio di tipo RING-H2 PROSITE (132–176 aminoacidi)10. Come illustrato nella Figura 1, per ottenere una proteina mutante carente di E3, almeno una delle otto C o H conservate nel dominio RING (Figura 1A) deve essere mutagenizzata (Figura 1B). Così, come primo passo, sono state generate due versioni mutanti di RHA1B, RHA1BC135S (una sostituzione di Cys by Ser nel conservato C3 del dominio RING) e RHA1BK146R (una sostituzione di Lys da Arg nell'unico Lys presente in RHA1B). Anche se una singola sottounità E3 media il trasferimento dell'ubiquitina dall'ubiquitina che ospita l'E2 al substrato piuttosto che interagire direttamente con l'ubiquitina, l'auto-oniquitazione dell'E3 a Lys potrebbe essere necessaria per la sua massima attività enzimatica.
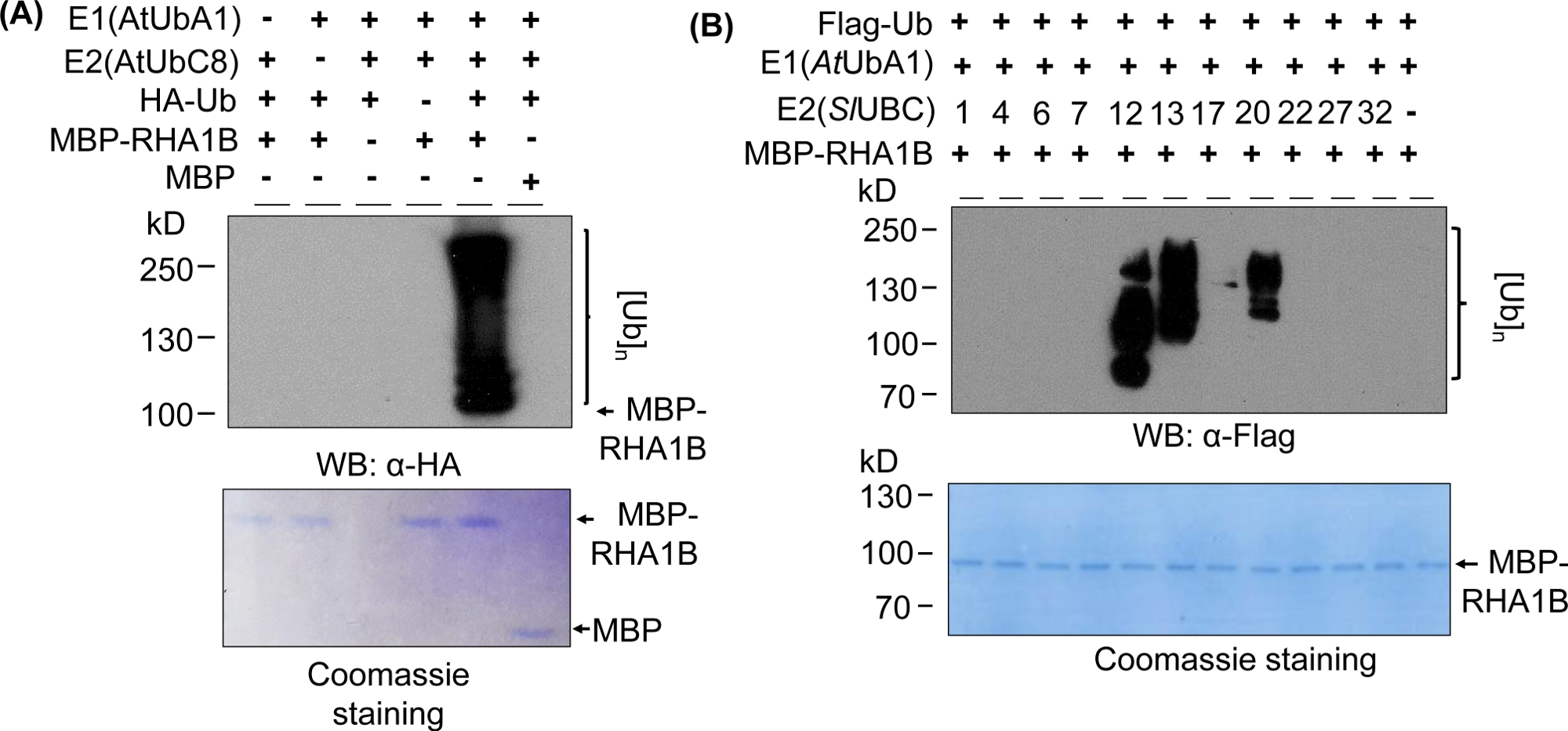
I risultati del gonfiore occidentale nella Figura 2A mostrano un tipico risultato positivo positivo del saggio in vitro, con uno striscio multibandante che inizia dal peso molecolare della proteina testata (ad esempio, RHA1B in vitro- 100 kDa) e progredisce verso l'alto. L'anticorpo anti-HA riconobbe l'Ub marcato HA incorporato nella catena poli-ubiquitinazione di diverse lunghezze, creando questo tipico striscio associato all'ubiquitina. Per convalidare i risultati positivi, Figura 2A presenta anche tutti i controlli negativi importanti mancanti singoli componenti (E1, E2, Ub, o MBP-RHA1B) o utilizzando MBP come controllo e privo del segnale di ubiquitinazione spalmato. Inoltre, la colorazione blu Coomassie della membrana PVDF ha mostrato un carico uguale di MBP-RHA1B o MBP in tutti i controlli.
La figura 2mostra come i risultati dell'ubiquitinazione in vitro variavano a seconda della specifica combinazione E2/E3. In questo esempio sono stati testati 11 diversi E2 che rappresentano 10 diverse famiglie E2. L'attività di ubiquitinazione rilevata variava da nessun segnale (nessuna macchia) a uno striscio multibandante a partire da diversi pesi molecolari, indicando diversi modelli di ubiquitazione.
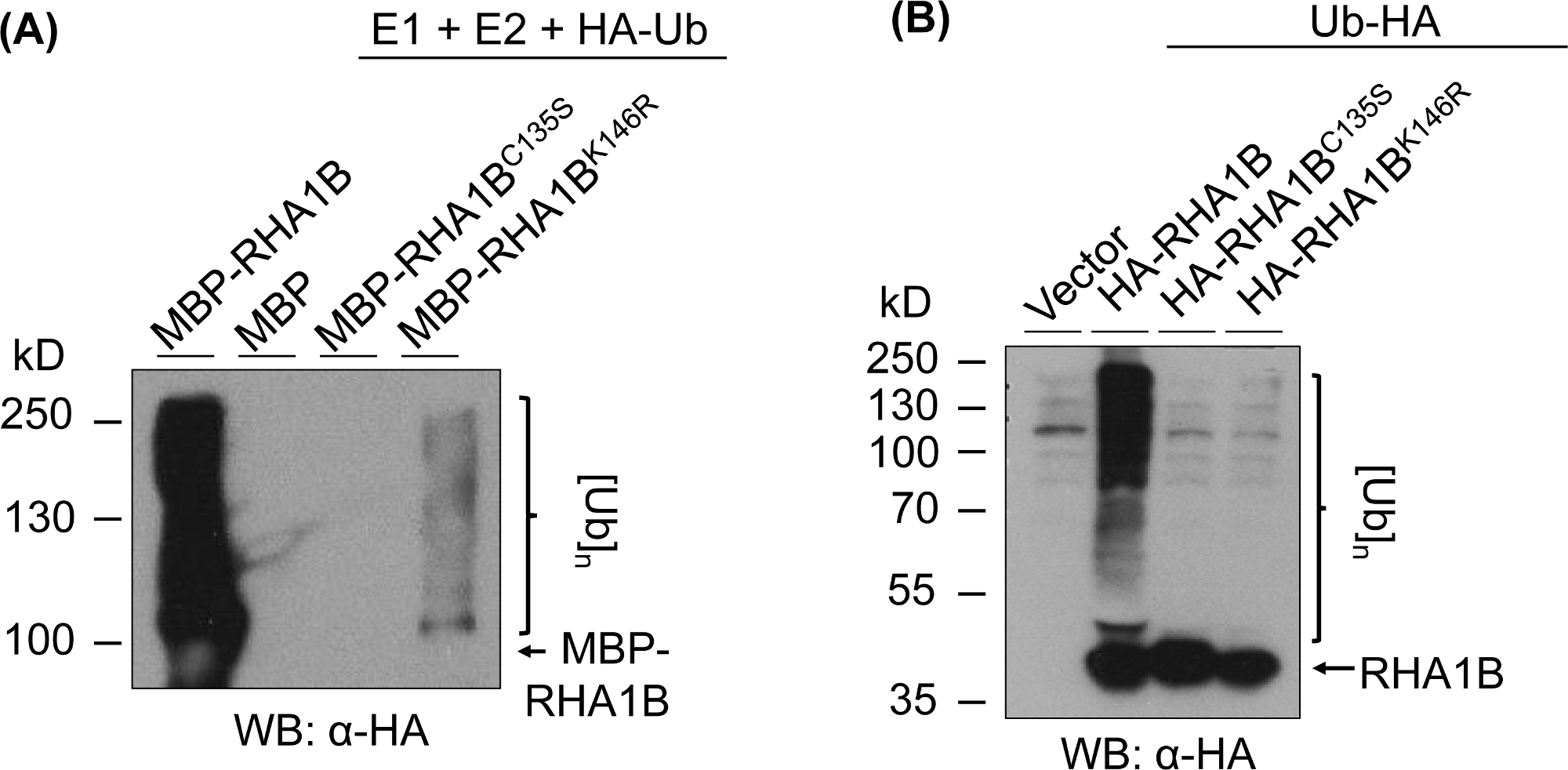
La figura 3 mostra i risultati dell'espressione di ubiquitinazione per le versioni RING- e K-mutante delle proteine testate. La mancanza di attività enzimatica per RHA1BC135S è supportata dalla sua incapacità di generare uno striscio multibandante in vitro (Figura 3A) o di promuovere il segnale poli-ubiquitination in planta (Figura 3B). È da notare che la sovraespressione di HA-tagged Ub in planta ha dato ubiquitinazione livello basale in tutti i campioni testati, compreso il controllo vettoriale, in contrasto con il forte segnale di ubiquitinazione conferito dall'attività zigmatica di tipo selvaggio RHA1B. Inoltre, l'analisi sul mutante RHA1BK146R suggerisce che il residuo K146 è anche essenziale per l'attività E3 di RHA1B. Sebbene sia stato rilevato in vitro un segnale marginale di auto-onniquitazione (Figura 3A), l'indice in planta ha determinato che il mutante è carente di E3 (Figura 3B, viene rilevato solo il segnale di ubiquitinazione di fondo).
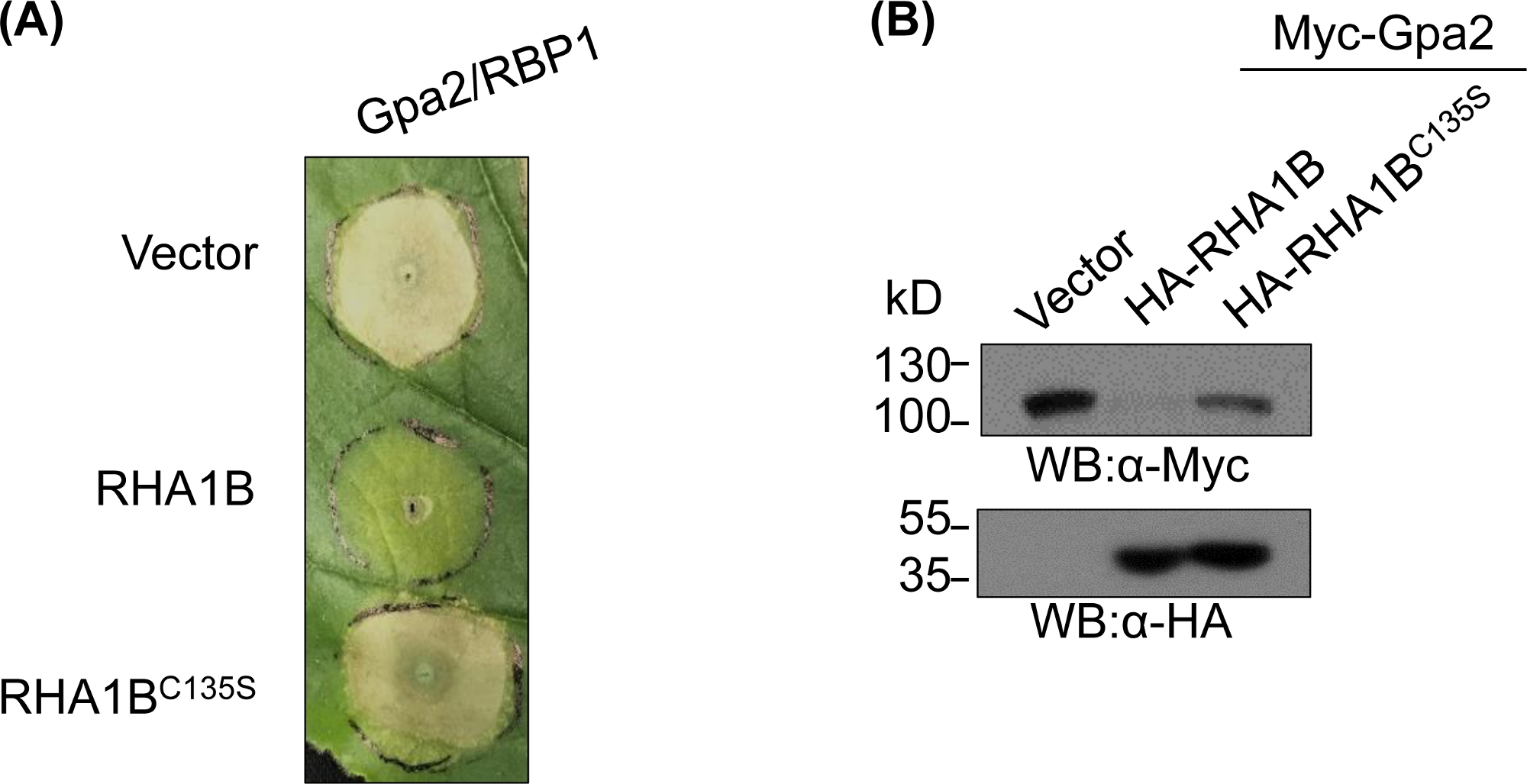
Dopo aver generato e convalidato biochimicamente il mutante carente di E3, gli studi funzionali possono essere progettati per determinare il ruolo biologico associato all'E3 della liga ubiquitina RING E3 collaudati. Nel caso di RHA1B, questo effettore nematode sopprime la segnalazione immunitaria delle piante, come si manifesta con la soppressione della morte delle cellule HR innescate da Gpa2. Come presentato nella Figura 4A, a differenza del tipo selvaggio RHA1B, il mutante RHA1BC135S privo di attività ligase E3 non ha interferito con la morte delle cellule HR. Dato che il risultato più comune dell'ubiquitinazione proteica è la sua degradazione mediata dal proteasoso, le mutazioni che risiedono nel dominio RING possono essere utilizzate anche per verificare una capacità dipendente dall'E3 di innescare la degradazione dei loro substrati diretti e/o indiretti. Così, significativamente, risultati di gonfiore occidentale in Figura 4B confermano che Gpa2 non si è accumulato in presenza di tipo selvaggio RHA1B ma RHA1BC135S non ha avuto alcun impatto sulla stabilità della proteina Gpa2.

Figura 1: Rappresentazione schematica del principio e dei passaggi coinvolti nella mutagenesi diretta al sito. (A) Evidenziato il dominio RING-CH/H2 con Cis conservati e i Suoi aminoacidi. (B) Un esempio di progettazione di primer mutageni. (C) Passi della mutagenesi diretta al sito. Fare clic qui per visualizzare una versione più grande di questa figura.
{kind=link}

Figura 2: Analisi dell'ubiquitazione rappresentativa in vitro. (A) Il gel superiore mostra il saggio di ubiquitinazione, compresi tutti i controlli negativi, e il gel inferiore mostra un carico uguale. (B) La gamma di risultati attesi a seconda degli enzimi E2. Questa cifra è stata modificata da Kud et al8. Fare clic qui per visualizzare una versione più grande di questa figura.
{kind=link}

Figura 3: Risultati del test dell'ubiquitazione per i mutanti RING e K (RHA1BC135S e RHA1BK146R). (A) Risultati dell'ubiquitazione in vitro per RHA1BC135S e RHA1BK146R. (( B) In planta risultati di prova di ubiquitinazione per RHA1BC135S e RHA1BK146R. Questa cifra è stata modificata da Kud et al8. Fare clic qui per visualizzare una versione più grande di questa figura.
{kind=link}

Figura 4: Studio funzionale rappresentativo per le funzioni biologiche dipendenti dall'E3. Un esempio di studi funzionali che mostrano la funzione biologica dipendente da E3. (A) Soppressione della morte delle cellule HR dipendenti dall'E3 e degradazione (B) di un immunorecettore vegetale Gpa2. Questa cifra è stata modificata da Kud et al8. Fare clic qui per visualizzare una versione più grande di questa figura.
{kind=link}
| Configurazione PCR | |
| 1 ll | plasmide (100 ng) |
| 1,5 ll | F primer mutageno (10 M) |
| 1,5 ll | R primer mutageno (10 M) |
| 1 ll | dNTP (10 mM) |
| 5 ll | buffer (10x) |
| 1 ll | Polimerasi Ultra Pfu (2,5 U/l) |
| 39 l l | ddH2O |
| 50 ll | VOLUME TOTALE |
Tabella 1: impostazione della reazione PCR
| programma termociclista | |||
| 1 | 95 gradi centigradi | 30 s | |
| 2 | 95 gradi centigradi | 30 s | |
| 3 | 60 gradi centigradi | 30 s | |
| 4 | 72 gradi centigradi | 5 min | ripetere 2-4 30 volte |
| 5 | 72 gradi centigradi | 5 min | |
Tabella 2: Programma termociclista PCR
| reazione di legatura impostata per l'esempio RHA1B | ||
| 1,5 ll | pMAL-c2::MBP vettore linearizzato per digestione con BamHI e SalI (60 ng) | |
| 7 l'uomo | Inserto RHA1B/RHA1BC135S o RHA1BK146R digerito con BamHI e SalI (25 ng) | |
| 1 ll | T4 buffer di ligase (10x) | |
| 0,5 l l | T4 ligase (400 U/ L) | |
| 10 ll | VOLUME TOTALE | |
Tabella 3: Reazione di ligazione impostata per l'esempio RHA1B.
Discussione
Chiarire la base biochimica e meccanicistica dei legamenti dell'ubiquitina di tipo E3 può contribuire notevolmente alla nostra comprensione del loro significato biologico nello sviluppo, nella segnalazione dello stress e nel mantenimento dell'omeostasi. Il protocollo qui descritto accoppia un approccio di mutagenesi con studi in vitro e in planta. Introducendo una singola sostituzione di aminoacidi nei residui conservati del dominio RING attraverso la mutagenesi site-direct, il mutante con deficit di E3 risultante può essere testato in parallelo con la proteina di tipo selvatico per collegare l'attività enzimatica con la funzionalità.
È fondamentale identificare correttamente il dominio RING, in particolare i suoi Cis conservati e i Suoi residui. Strumenti online come PROSITE possono essere utilizzati per farlo10. Per destabilizzare il dominio RING responsabile del reclutamento dell'enzima E2, Cys è normalmente sostituito con Ser, che è il suo più vicino sostituto strutturale senza la capacità di creare un legame disulfide utilizzato per il coordinamento dello zinco. Lorick ealtri ha mostrato che la mutazione in uno qualsiasi di quei residui Cis critici abolirebbe l'attività di ubiquitazione della singola sottounità RING-tipo E3 ligas5. Sebbene alcuni residui di Cys siano importanti anche per i complessi multiunità E3 ligase contenenti proteine di tipo RING, a causa della struttura tridimensionale sfaccettata e dinamica di quelle complessi di ubiquitazione e del diverso ruolo delle proteine di tipo RING, una singola sostituzione dei residui conservati nel dominio RING in multiunità E3 ligase non è riuscita a generare un fenotipo carente di ligase11.
Per la mutagenesi diretta dal sito, abbiamo scoperto che l'uso di vettori di plasmide più piccoli e di cicli di amplificazione più bassi di solito produceva una maggiore efficienza per la mutagenesi. L'enzima Pfu può essere sostituito con qualsiasi altra polimerasi del DNA ad alta fedeltà e ad alta espressività. Inoltre, se il gene di interesse contiene codoni rari, un'altra macchia di E. coli, Rosetta, può essere utilizzata per ottenere una maggiore resa della proteina ricombinante. Inoltre, sia il tempo di incubazione che la temperatura per l'induzione IPTG possono essere ulteriormente ottimizzati. Temperature più basse riducono il tasso di divisione di E. coli, che potrebbe essere favorevole per l'espressione di alcune proteine. Anche se una maggiore concentrazione di IPTG potrebbe migliorare l'espressione proteica, inibisce anche i processi di divisione di E. coli e non è raccomandato.
La singola sottounità RING tipo E3 lega non solo come scaffold molecolare che posiziona l'intermedio E2-Ub in prossimità del substrato, ma stimola anche l'attività di trasferimento dell'ubiquitina dei loro Cognati E2. Inoltre, dato che una combinazione E2/E3 è importante per la lunghezza e i collegamenti della catena di poliuzia che determina il destino di un substrato modificato, qualsiasi considerazione degli E3 di tipo RING deve includere i loro partner enzimatici, E2s12. Come illustrato nella Figura 3B, non tutti gli E2 testati sono compatibili con la ligase RHA1B. Pertanto, i saggi di ubiquitazione in vitro devono essere trasportati in parallelo con più enzimi E2 che rappresentano diverse classi E2 per evitare falsi risultati negativi.
Presentato qui è il saggio enzimatico in vitro che rileva la capacità di auto-ubiquitinazione delle proteine testate di tipo RING. Tuttavia, con piccole modifiche, questo protocollo può essere facilmente adattato per rilevare l'ubiquitinazione in vitro dei substrati. A tal fine, la miscela di ubiquitinazione in vitro dal passo 2.15 dovrebbe essere integrata con la proteina ricombinante del potenziale substrato di ligase E3 (500 ng). Dopo un'incubazione di 2 h a 30 , la proteina ubiquitinata deve essere catturata usando 15 -L di matrice di affinità anti-HA (se viene utilizzato HA-Ub, o matrice di affinità anti-FLAG se viene utilizzato FLAG-Ub) per agitazione per 2 h a 4 gradi centigradi. Dopo aver lavato le perline 4 volte con il tampone di lavaggio Ub freddo (20 mM MM Tris pH 7,5, 100 mM NaCl, 0,1 mM EDTA, 0.05% Tween 20, 1x PMSF), scartare tutto tranne 40 gradi l del buffer e passare al passo 2.16. Il segnale di ubiquitinazione, rilevato da anticorpi specifici per l'Ub e il substrato con etichettatura epitope, rispettivamente, emergendo dal peso molecolare della proteina del substrato, conferma la specificità substrato/enzima.
Inoltre, l'identificazione dei substrati e3 ligasi in vivo è di solito associata a molteplici sfide dovute all'interazione transitoria enzimatica-substrato e alla rapida degradazione della proteina bersaglio ubiquitinata. L'uso di un mutante carente di ligase E3, che interagisce ancora con il suo obiettivo ma non lo onniquizza più13,è un'alternativa molto utile all'aggiunta dell'inibitore proteasomico MG132, che non sempre interferisce sempre sufficientemente con la funzione proteasome 26S.
Una caratteristica comune dei legamenti E3 di tipo RING è la tendenza a formarsi e funzionare come omo- e/o eterodimeri. È interessante notare che la sostituzione nei residui conservati del dominio RING è di solito associata a un fenotipo negativo dominante in cui il tipo RING e3 ligase mutato blocca l'attività enzimatica di una proteina nativa di tipo selvatico13. Pertanto, la sovraespressione dei mutanti RING in planta può essere un approccio alternativo all'abbattimento del gene delle ligase E3.
Divulgazioni
Gli autori non hanno nulla da rivelare.
Riconoscimenti
Il nostro lavoro è stato reso possibile dal sostegno finanziario della sovvenzione competitiva Agriculture and Food Research Initiative (2017-67014-26197; 2017-67014-26591) dell'USDA National Institute of Food and Agriculture, USDA-NIFA Farm Bill, Northwest Potato Consorzio e ISDA Specialty Crop.
Materiali
| Name | Company | Catalog Number | Comments |
| Acetic acid | Sigma-Aldrich | A6283 | |
| Acetosyringone | Sigma-Aldrich | D134406 | |
| Amylose resin | NEB | E8021S | |
| ATP | Sigma-Aldrich | A1852 | |
| Bacterial protease inhibitor | Sigma-Aldrich | P8465 | |
| Bromphenol Blue | VWR | 97061-690 | |
| CaCl2 | Sigma-Aldrich | C1016 | |
| Centrifuge | Beckman Coulter | model: Avanti J-25 | |
| Commassie Blue | VWR | 97061-738 | |
| Creatine phosphate | Sigma-Aldrich | P7936 | |
| Creatine phosphokinase | Sigma-Aldrich | C3755 | |
| DNA clean & concentrator Kit | ZYMO RESEARCH | D4029 | |
| DpnI | NEB | R0176S | |
| DTT | Sigma-Aldrich | D0632 | |
| E. coli BL21 | Thermo Fisher Scientific | C600003 | |
| E. coli DH5α competent cells | Thermo Fisher Scientific | 18265017 | |
| EDTA | Sigma-Aldrich | 324504 | |
| FeSO4 7H2O | Sigma-Aldrich | F7002 | |
| FLAG-Ub | BostonBiochem | U-120 | |
| Glucose | VWR | 188 | |
| Glycerol | Sigma-Aldrich | G5516 | |
| HA-Ub | BostonBiochem | U-110 | |
| Heat block | VWR | model: 10153-318 | |
| Incubator | VWR | model: 1525 Digital Incubator | |
| Incubator shaker | Thermo Fisher Scientific | model: MaxQ 4000 | |
| IPTG | Roche | 10724815001 | |
| KCl | Sigma-Aldrich | P9333 | |
| LB Broth | Sigma-Aldrich | L3022 | |
| Liquide nitrogen | university chemistore | ||
| Maltose | Sigma-Aldrich | 63418 | |
| MES | Sigma-Aldrich | M3671 | |
| Methanol | Sigma-Aldrich | 34860 | |
| MgCl2 | Sigma-Aldrich | 63138 | |
| MgSO4 7H2O | Sigma-Aldrich | 63138 | |
| Microcentrifuge | Eppendorf | model: 5424 | |
| Miniprep plasmid purification kit | ZYMO RESEARCH | D4015 | |
| monoclonal anti-FLAG antibody | Sigma-Aldrich | F3165 | |
| monoclonal anti-HA antibody | Sigma-Aldrich | H9658 | |
| monoclonal anti-MYC antibody | Sigma-Aldrich | WH0004609M2 | |
| Mortar | VWR | 89038-144 | |
| NaCl | Sigma-Aldrich | S7653 | |
| NaH2PO4 | Sigma-Aldrich | S8282 | |
| NanoDrop | Thermo Fisher Scientific | model: 2000 Spectrophotometer | |
| Needle | Thermo Fisher Scientific | 14-826-5C | |
| NH4Cl | Sigma-Aldrich | A9434 | |
| PCR machine | Bio-Rad | model: C1000 | |
| Pestle | VWR | 89038-160 | |
| Pfu Ultra | Agilent Technologies | 600380 | |
| Plant protease inhibitor coctail | Sigma-Aldrich | P9599 | |
| pMAL-c2 | NEB | N8076S | |
| PMSF | Sigma-Aldrich | P7626 | |
| Polyvinylpolypyrrolidone | Sigma-Aldrich | P6755 | |
| SDS | Sigma-Aldrich | 1614363 | |
| Sonicator | Qsonica Sonicators | model: Q125 | |
| Syringe | Thermo Fisher Scientific | 22-253-260 | |
| Tris | Sigma-Aldrich | T1503 | |
| T4 ligase | NEB | M0202S |
Riferimenti
- Freemont, P. S., Hanson, I. M., Trowsdale, J. A novel gysteine-rich sequence motif. Cell. 64, 483-484 (1991).
- Borden, K. L. B. RING fingers and B-boxes: Zinc-binding protein-protein interaction domains. Biochemistry and Cell Biology. 76, 351-358 (1998).
- Barlow, P. N., Luisi, B., Milner, A., Elliott, M., Everett, R. Structure of the C3HC4 Domain by 1H-nuclear Magnetic Resonance Spectroscopy: A New Structural Class of Zinc-finger. Journal of Molecular Biology. 237, 201-211 (1994).
- Borden, K. L. B., et al. The solution structure of the RING finger domain from the acute promyelocytic leukaemia proto-oncoprotein PML. The EMBO Journal. 14, 1532-1541 (1995).
- Lorick, K. L., et al. RING fingers mediate ubiquitin-conjugating enzyme (E2)-dependent ubiquitination. Proceedings of the National Academy of Sciences. 96, 11364-11369 (1999).
- Jiménez-López, D., Muñóz-Belman, F., González-Prieto, J. M., Aguilar-Hernández, V., Guzmán, P. Repertoire of plant RING E3 ubiquitin ligases revisited: New groups counting gene families and single genes. PLoS ONE. 13, 1-28 (2018).
- Sacco, M. A., et al. The Cyst Nematode SPRYSEC Protein RBP-1 Elicits Gpa2- and RanGAP2-Dependent Plant Cell Death. PLoS Pathogens. 5, 1-14 (2009).
- Kud, J., et al. The potato cyst nematode effector RHA1B is a ubiquitin ligase and uses two distinct mechanisms to suppress plant immune signaling. PLoS Pathogens. 15, 1007720(2019).
- Bradford, M. M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Analytical Biochemistry. 72, 248-254 (1976).
- Sigrist, C. J. A., et al. New and continuing developments at PROSITE. Nucleic Acids Research. 41, 344-347 (2013).
- Dove, K. K., Stieglitz, B., Duncan, E. D., Rittinger, K., Klevit, R. E. Molecular insights into RBR E3 ligase ubiquitin transfer mechanisms. EMBO Reports. 17, 1221-1235 (2016).
- Metzger, M. B., Pruneda, J. N., Klevit, R. E., Weissman, A. M. RING-type E3 ligases: Master manipulators of E2 ubiquitin-conjugating enzymes and ubiquitination. Biochimica et Biophysica Acta - Molecular Cell Research. 1843, 47-60 (2014).
- Xie, Q., et al. SINAT5 promotes ubiquitin-related degradation of NAC1 to attenuate auxin signals. Nature. 419, 167-170 (2002).
Ristampe e Autorizzazioni
Richiedi autorizzazione per utilizzare il testo o le figure di questo articolo JoVE
Richiedi AutorizzazioneThis article has been published
Video Coming Soon
Personale delle biblioteche
Copyright © 2025 MyJoVE Corporation. Tutti i diritti riservati