
Method Article
Imaging ottico basato su fotodiodi per la registrazione di dinamiche di rete con risoluzione a singolo neurone in invertebrati non transgenici
* Questi autori hanno contribuito in egual misura
In questo articolo
Riepilogo
Questo protocollo presenta un metodo per l'imaging dell'attività della popolazione neuronale con risoluzione a singola cellula in specie di invertebrati non transgenici utilizzando coloranti sensibili alla tensione di assorbanza e un array di fotodiodi. Questo approccio consente un flusso di lavoro rapido, in cui l'imaging e l'analisi possono essere perseguiti nel corso di un solo giorno.
Abstract
Lo sviluppo di preparati di invertebrati transgenici in cui l'attività di insiemi specificabili di neuroni può essere registrata e manipolata con la luce rappresenta un progresso rivoluzionario per gli studi delle basi neurali del comportamento. Tuttavia, uno svantaggio di questo sviluppo è la sua tendenza a concentrare i ricercatori su un numero molto piccolo di organismi "progettisti" (ad esempio, C. elegans e Drosophila), potenzialmente con un impatto negativo sulla ricerca di studi comparativi su molte specie, che è necessario per identificare i principi generali della funzione di rete. Il presente articolo illustra come la registrazione ottica con coloranti sensibili alla tensione nel cervello di specie di gasteropodi non transgenici possa essere utilizzata per rivelare rapidamente (cioè nel corso temporale di singoli esperimenti) le caratteristiche dell'organizzazione funzionale delle loro reti neurali con risoluzione a singola cellula. Descriviamo in dettaglio i metodi di dissezione, colorazione e registrazione utilizzati dal nostro laboratorio per ottenere tracce di potenziale d'azione da dozzine a ~ 150 neuroni durante programmi motori comportamentalmente rilevanti nel SNC di più specie di gasteropodi, tra cui uno nuovo alle neuroscienze - il nudibranco Berghia stephanieae. L'imaging viene eseguito con coloranti sensibili alla tensione di assorbanza e un array di fotodiodi a 464 elementi che campiona a 1.600 fotogrammi / secondo, abbastanza veloce da catturare tutti i potenziali d'azione generati dai neuroni registrati. È possibile ottenere registrazioni multiple di diversi minuti per preparazione con poco o nessun segnale di sbiancamento o fototossicità. I dati ottici grezzi raccolti attraverso i metodi descritti possono essere successivamente analizzati attraverso una varietà di metodi illustrati. Il nostro approccio di registrazione ottica può essere facilmente utilizzato per sondare l'attività di rete in una varietà di specie non transgeniche, rendendolo adatto per studi comparativi su come i cervelli generano comportamento.
Introduzione
Lo sviluppo di linee transgeniche di invertebrati come Drosophila e C. elegans ha fornito potenti sistemi in cui le basi neurali del comportamento possono essere interrogate e manipolate otticamente. Tuttavia, questi preparati speciali possono avere lo svantaggio di ridurre l'entusiasmo per gli studi sui circuiti neurali di specie non transgeniche, in particolare per quanto riguarda l'introduzione di nuove specie nella ricerca neuroscientifica. Concentrarsi esclusivamente su uno o due sistemi modello è dannoso per la ricerca di principi generali della funzione di rete, perché gli studi comparativi rappresentano una via essenziale attraverso la quale tali principi vengono scoperti1,2,3,4. Il nostro obiettivo qui è quello di dimostrare un approccio di imaging su larga scala per ottenere una rapida comprensione della struttura funzionale delle reti neurali dei gasteropodi, nel tentativo di facilitare studi comparativi della funzione della rete neurale.
Molluschi gasteropodi come Aplysia, Lymnaea, Tritonia, Pleurobranchaea e altri sono stati a lungo utilizzati per studiare i principi della funzione della rete neurale, in gran parte perché i loro comportamenti sono mediati da neuroni di grandi dimensioni, spesso identificabili individualmente situati sulla superficie dei gangli, rendendoli facilmente accessibili alle tecniche di registrazione5. Nel 1970, sono stati sviluppati coloranti sensibili alla tensione (VSD) che possono integrarsi nella membrana plasmatica che presto hanno permesso le prime registrazioni prive di elettrodi dei potenziali d'azione generati da più neuroni6. Qui, dimostriamo il nostro uso di VSD per esaminare l'attività di rete in diverse specie di gasteropodi, tra cui uno nuovo alle neuroscienze, Berghia stephanieae. Il dispositivo di imaging è un array di fotodiodi a 464 elementi (PDA) disponibile in commercio che campiona a 1.600 fotogrammi / secondo (Figura 1), che, se utilizzato con VSD ad assorbimento rapido, rivela i potenziali d'azione di tutti i neuroni registrati7. I segnali registrati da tutti i diodi vengono visualizzati immediatamente dopo l'acquisizione e sovrapposti su un'immagine del ganglio nel software di acquisizione PDA, rendendo possibile lo sdodo dei neuroni di interesse con elettrodi affilati nella stessa preparazione8,9.
Nei dati PDA grezzi, molti diodi registrano in modo ridondante i neuroni più grandi e molti contengono anche segnali misti da più neuroni. Un punto di svolta è stato lo sviluppo di un metodo automatizzato di ordinamento dei picchi che utilizza l'analisi indipendente dei componenti per elaborare rapidamente ogni set di dati PDA grezzi a 464 canali in un nuovo set di tracce, in cui ogni neurone registrato appare in una traccia separata contenente solo i suoi potenziali d'azione10,11.
In questo articolo delineiamo i passaggi essenziali coinvolti nell'ottenere registrazioni del potenziale d'azione su larga scala dal sistema nervoso dei gasteropodi con un array di fotodiodi e VSD ad assorbimento rapido. Illustriamo inoltre metodi analitici che possono essere impiegati per raggruppare e mappare i neuroni registrati otticamente rispetto ai loro insiemi funzionali, e per caratterizzare caratteristiche a livello di popolazione che spesso non sono evidenti attraverso la semplice ispezione delle tracce di sparo12,13.
Protocollo
NOTA: il flusso di lavoro descritto di seguito è riepilogato nella Figura 2.
1. Ridurre al minimo le vibrazioni
- Se possibile, assicurarsi che il carro sia al piano terra e utilizzare un tavolo di isolamento a molla, che smorza una gamma più ampia di frequenze di vibrazione rispetto ai tavoli d'aria.
- Se si utilizza un tavolo a molla, assicurarsi che sia fluttuante (deve essere regolato ogni volta che si aggiunge o si toglie qualcosa dal tavolo).
- Ridurre il più possibile il rumore basato sulle vibrazioni nella sala immagini, anche fino al punto di interrompere il flusso d'aria durante l'imaging, se necessario. Ridurre al minimo le vibrazioni derivanti dalla turbolenza del fluido nei sistemi di perfusione.
NOTA: la preparazione neurale non deve muoversi durante l'acquisizione. Movimenti di qualsiasi tipo producono spostamenti di bordi di contrasto attraverso i diodi, portando a segnali artefatti. Se la procedura comporta uno stimolo durante l'acquisizione, non deve indurre il movimento di preparazione.
2. Esegui un test stenopeico per consentire il corretto allineamento delle foto del ganglio con i dati del PDA
- Posizionare un pezzo di foglio di alluminio con 3 piccoli fori infilati su un vetrino per microscopio. Scatta un'immagine dei tre fori con la fotocamera digitale montata sul suo fotoporto parfocale.
- Utilizzando il software di imaging fornito con il PDA, acquisire un breve file (ad esempio, 5 s). A metà dell'acquisizione, tocca il tavolo per indurre artefatti di vibrazione che saranno molto visibili attorno ai bordi dei fori stenopeiche, consentendo all'immagine dei fori stenopeiche di essere allineata con precisione con i dati ottici.
- Utilizzare la funzione Di sovrapposizione nel software di imaging, che si trova nella voce di menu "Visualizza | | di sovrapposizione di pagina Sovrapponi tracce con | immagine esterna Sovrapponi immagine", per sovrapporre i dati del diodo alla foto dei fori stenopeici, quindi regolare iterativamente le impostazioni x, y e di ingrandimento per la foto fino a quando i fori stenopeici si trovano direttamente in cima agli artefatti stenopeici nei dati del diodo.
- Salva questi numeri per allineare le immagini di preparazione scattate con la fotocamera con i dati del diodo negli esperimenti futuri.
NOTA: l'allineamento stenopeico del PDA deve essere eseguito solo una volta dopo che il PDA è stato montato sul microscopio, fino a quando non viene ruotato o rimosso, momento in cui deve essere fatto di nuovo.
- Salva questi numeri per allineare le immagini di preparazione scattate con la fotocamera con i dati del diodo negli esperimenti futuri.
3. Dissezioni per tre specie di gasteropodi marini
- Per le specie che crescono fino a grandi dimensioni, come Tritonia e Aplysia, inizia con individui più piccoli, che hanno gangli più sottili e meno opachi, facilitando l'ottenimento di luce sufficiente per un segnale ottimale al rumore.
- Avere pronta acqua di mare artificiale filtrata da utilizzare come soluzione salina per le dissezioni e gli esperimenti di imaging.
NOTA: In tutte le fasi successive del protocollo, "soluzione salina" indica l'acqua di mare artificiale. - Dissezione di Tritonia diomedea
- Metti un animale in frigorifero per circa 20 minuti per anestetizzarlo.
- Per gli animali più grandi, esporre il cervello tenendo l'animale in una mano, lasciando che l'estremità della testa si drappeggi sopra l'indice per esporre il "collo". Per gli animali più piccoli, appuntarli sul lato dorsale in un piatto di dissezione rivestito di cera prima di esporre il cervello.
- Usando le forbici per dissezione, fai un'incisione della linea mediana di 3-4 cm sul lato dorsale dell'animale, sopra la massa buccale (che può essere sentita attraverso la parete del corpo).
NOTA: La porzione necessaria del SNC, costituita da gangli cerebropleurici e pedali fusi, bilaterali, è arancione e distinta nell'aspetto dal tessuto circostante; si trova immediatamente posteriore ai rinofori e in cima alla massa buccale. - Asportare il SNC recidendo quei nervi innervando il corpo dell'animale usando pinza e forbici da microdissezione, mantenendo intatti tutti quei nervi che collegano i gangli centrali. Lasciare una lunga lunghezza del nervo del pedale 3 (PdN3), o qualsiasi nervo sarà stimolato.
- Utilizzare perni minuziali per fissare il SNC sul fondo di un piatto rivestito di elastomero riempito di soluzione salina per un'ulteriore dissezione. Mantenere la temperatura di preparazione a 11 °C perfondendo il piatto con soluzione salina erogata con un sistema di raffreddamento Peltier in linea controllato a feedback utilizzando una pompa peristaltica.
- Usando pinza e forbici per microdissezione, rimuovere con cura lo strato vagamente aderente del tessuto connettivo da tutto il SNC. Lasciare la guaine fine aderente ai gangli.
- Brevemente (~10 s) immergere i gangli in una soluzione di glutaraldeide allo 0,5% in soluzione salina. Rimettere i gangli nel piatto rivestito di elastomero perfuso di soluzione salina, consentendo alla soluzione salina di lavare via la glutaraldeide prima di iniziare la colorazione VSD.
NOTA: Questa leggera correzione del tessuto connettivo e dei suoi muscoli intrinseci aiuterà a prevenire il movimento durante l'imaging.
- Dissezione di Aplysia californica
- Anestetizzare un animale di circa 40 g iniettando ~ 20 mL di MgCl2 350 mM nel corpo attraverso la superficie ventrale (piede).
- Usa i perni per posizionare il lato ventrale dell'animale verso l'alto in un piatto di dissezione rivestito di cera.
- Usando le forbici per dissezione, fai un'incisione della linea mediana di 2-3 cm lungo l'estensione più anteriore del piede. Fissare i lembi del piede su entrambi i lati dell'incisione per rivelare parte del SNC e della massa buccale.
NOTA: La porzione necessaria del SNC, costituita da gangli cerebrali fusi e gangli pleurici e pedali bilaterali strettamente apposti, è giallo-arancio e distinta nell'aspetto dal tessuto circostante; si trova dorsale e posterolaterale alla massa buccale muscolare bulbosa. - Usa pinza e forbici di dissezione per sezionare con cura la massa buccale, rivelando i gangli cerebrali.
- Asportare il SNC recidendo quei nervi innervando il corpo dell'animale usando pinza e forbici da microdissezione, mantenendo intatti tutti quei nervi che collegano i gangli centrali. Lasciare una lunga lunghezza del nervo del pedale 9 (PdN9), o qualsiasi nervo sarà stimolato.
- Utilizzare perni minuziosi per posizionare il SNC in un piatto rivestito di elastomero riempito di soluzione salina. Mantenere la temperatura di preparazione a 15-16 °C perfondendo il piatto con soluzione salina che passa attraverso un dispositivo di raffreddamento Peltier.
- Usando pinza e forbici da microdissezione, rimuovere il tessuto connettivo eccessivo dal SNC e sezionare una porzione superficiale della guaina sul ganglio o sui gangli da immaginare. Fai attenzione durante questo processo a non fare un buco nella guaiella, il che comporterebbe la fuoriuscita dei neuroni dall'interno.
- Immergere brevemente (~20 s) i gangli in una soluzione di glutaraldeide allo 0,5% in soluzione salina. Rimettere i gangli nel piatto rivestito di elastomero perfuso di soluzione salina, consentendo alla soluzione salina di lavare via la glutaraldeide prima di iniziare la colorazione VSD.
- Dissezione di Berghia stephanieae
- Metti un animale in frigorifero per circa 20 minuti per anestetizzarlo.
- Utilizzando un piatto rivestito di elastomero riempito con soluzione salina a temperatura ambiente, posizionare i perni di minuzia sia nella testa che nella coda.
- Usando le forbici da microdissezione, fai un'incisione dorsale di 5-7 mm superficiale al SNC.
NOTA: Gli occhi, macchie scure che risiedono all'interno dell'animale accanto al SNC, segnano convenientemente la posizione della porzione necessaria del SNC, che consiste in gangli cerebropleurici e pedali fusi bilateralmente e si trova in cima alla massa buccale. - Asportare il SNC recidendo quei nervi innervando il corpo dell'animale usando pinza e forbici da microdissezione. Lasciare che qualsiasi nervo venga stimolato sufficientemente a lungo per un elettrodo di aspirazione.
4. Colorare la preparazione con un colorante sensibile alla tensione
- Preparare soluzioni stock di RH155 (noto anche come NK3041) o RH482 (noto anche come NK3630 o JPW1132).
- RH155: Sciogliere 5,4 mg di colorante solido in 1 mL di EtOH al 100%, pipettando 29 μL in ciascuno dei 34 tubi di microcentrifuga. Con il contenuto di ciascun tubo esposto all'aria, lasciarli asciugare durante la notte al buio. Tappare e collocare le aliquote solide risultanti di RH155, ciascuna contenente 0,15 mg, in un congelatore a -20 °C.
- RH482: Sciogliere 2 mg di colorante solido in 100 μL di DMSO, dividere la soluzione in 20 aliquote da 5 μL, ciascuna contenente 0,1 mg di RH482, e conservare in un congelatore a -20 °C.
NOTA: Per Tritonia e Aplysia,la perfusione del bagno o l'applicazione a pressione possono essere utilizzate per caricare il VSD RH155 nelle membrane dei neuroni nella preparazione. L'applicazione a pressione ha il vantaggio di esporre solo il ganglio che viene ripreso al VSD.
- Per la perfusione in bagno, aggiungere 5 mL di soluzione salina a ciascuna delle due aliquote di cui sopra di RH155 solido e vortice in soluzione, producendo una soluzione combinata di 10 mL contenente 0,03 mg/mL RH155.
- Perfondere al buio (per evitare il fotosableaching) da 1 a 1,5 ore a 11 °C per Tritonia e a 16 °C per Aplysia. Mantenere la temperatura facendo passare la soluzione di perfusione attraverso un sistema di raffreddamento Peltier.
- Per l'applicazione in pressione, aggiungere 500 μL di soluzione salina a un'aliquota di RH155 e vortice per produrre una concentrazione di colorante di 0,3 mg/ml.
- Aspirare circa 200 μL della soluzione in tubi di polietilene utilizzando un microdispenser portatile, assicurandosi che vi sia una buona corrispondenza tra il diametro del tubo e il diametro del ganglio da colorare.
- Utilizzare un micromanipolatore per posizionare con cura l'estremità del tubo sopra il ganglio bersaglio, abbassandolo fino a formare un sigillo aderente sul ganglio. Utilizzare il tipo di sistema di raffreddamento sopra descritto per mantenere i gangli alla temperatura desiderata.
- Attenuare le luci della stanza per evitare il fotosciviazione e ruotare la manopola dell'applicatore del microdispenser ogni 5 minuti per forzare più colorante sul ganglio.
- Controllare a 30 minuti per confermare che si sta verificando una buona colorazione, quindi continuare per un tempo di colorazione totale di circa 1 ora.
- Per la colorazione in Berghia,aggiungere 1 mL di soluzione salina a un'aliquota congelata di RH482 e vortice per dissolvere.
- Trasferire 200 μL di questa soluzione in un tubo microcentrifuga contenente 800 μL di soluzione salina e vortice in soluzione, producendo una soluzione colorante finale di 0,02 mg/ml di RH482 in soluzione salina con DMSO allo 0,1%.
- Posizionare l'intero SNC nel tubo microcentrifuga, avvolgendo il tubo in un foglio di alluminio per evitare il fotosableaching e stringere la mano ogni 5-6 minuti per circa 1 ora. Conservare i restanti 800 μL della prima soluzione in frigorifero e utilizzare per un massimo di 3 giorni per macchiare i preparati successivi.
5. Appiattire la preparazione e impostare per la stimolazione nervosa
NOTA: i passaggi descritti in questa sezione devono essere eseguiti con un'illuminazione minima o con luce verde per ridurre al minimo il fotosaguamento.
- Dopo la colorazione, immergere il SNC in soluzione salina all'interno della camera di imaging e posizionare sotto un microscopio di dissezione.
- Posiziona pezzi di silicone (per Tritonia o Aplysia)o macchie di vaselina (per Berghia)a sinistra ea destra del ganglio / gangli da immaginare.
- Premere un pezzo di dimensioni appropriate di un coperchio di vetro o plastica sul preparato per appiattirlo. Premere con fermezza ma non così forte da danneggiare i neuroni.
NOTA: Appiattire la superficie convessa del preparato in questo modo renderà parfocale un numero maggiore di neuroni, aumentando così il numero di neuroni registrati, e aiuterà inoltre a immobilizzare la preparazione durante l'imaging. - Se si stimola un nervo a suscitare un programma motorio fittizio, preparare un elettrodo di aspirazione la cui punta anteriore è larga circa quanto il diametro del nervo. Eseguire questa operazione fondendo accuratamente un segmento di tubo di polietilene PE-100 su una fiamma mentre si tirano delicatamente entrambe le estremità del segmento del tubo e quindi si taglia il cono risultante nel punto desiderato.
- Disegnare un piccolo volume di soluzione salina attraverso l'estremità affusolata di un elettrodo di aspirazione in polietilene, seguito dall'estremità del nervo da stimolare, attaccando una lunghezza di tubo polimerico flessibile a pareti spesse all'estremità posteriore dell'elettrodo e utilizzando l'aspirazione della bocca per applicare una pressione negativa.
- Confermare che la soluzione salina nell'elettrodo manca di bolle che potrebbero interrompere la conduzione elettrica.
6. Preparazione e ottimizzazione per l'imaging
- Spostare la camera sul rig di imaging. Avviare la perfusione salina attraverso la camera di registrazione e posizionare la sonda di temperatura vicino alla preparazione. Impostare il termoregolatore per la temperatura desiderata per la specie oggetto dell'immagine (per Tritonia,11 °C, per Aplysia,15-16 °C, o per Berghia,26-27 °C).
- Posizionare un filo d'argento clorurato lungo l'elettrodo di aspirazione, assicurandosi che contatti la soluzione salina nell'elettrodo e mettere l'altro filo Ag-AgCl (il percorso di ritorno) nella soluzione salina del bagno, vicino all'elettrodo di aspirazione.
- Abbassare la lente ad immersione in acqua nella soluzione salina. Chiudere il diaframma di base, quindi alzare o abbassare il condensatore del sottostadio e regolare la messa a fuoco fino a quando i bordi del diaframma sono a fuoco nitido, creando l'illuminazione Köhler.
- Concentrati sulla regione della preparazione da immaginare. I pregiudizi si concentrano sui neuroni più piccoli rispetto a quelli più grandi, poiché i segnali ottici provenienti da neuroni più grandi hanno maggiori probabilità di essere registrati rispetto a quelli più piccoli anche se sono leggermente sfocati.
- Scatta una foto del ganglio da fotografare con la fotocamera digitale parfocale.
- Con l'interruttore di guadagno del pannello di controllo impostato su 1x, ispezionare l'intensità della luce a riposo (RLI) nel software di imaging facendo clic sul pulsante "RLI"e controllando il RLI medio dei diodi. Regolare il livello di tensione inviato da uno stimolatore all'alimentazione della lampada a LED e continuare a controllare il livello medio di RLI fino a quando non si trova nell'intervallo desiderato (di solito intorno a 3-4 V).
NOTA: sono auspicabili RTI elevati, corrispondenti a circa 3-4 V sul PDA. Maggiore è la luce, migliore è il rapporto segnale-rumore dei segnali ottici, tuttavia, questo deve essere bilanciato con una velocità più veloce di fotosbiancamento a RTI più elevati. Questo rischio è ridotto al minimo utilizzando lenti ad alto NA. Gli obiettivi ad immersione in acqua utilizzati sono 10x/0,6 NA, 20x/0,95 NA, 40x/0,8 NA e 40x/1,15 NA.
- Impostare l'interruttore di guadagno del pannello di controllo su 100x per la registrazione.
- Se si stimola un nervo, impostare la tensione, la frequenza e la durata desiderate su uno stimolatore separato da quello utilizzato per impostare il livello di luce. Verificare che l'attivazione TTL tra il pannello di controllo e lo stimolatore sia configurata correttamente.
NOTA: I parametri di stimolo nervoso campione in ogni specie sono i seguenti: Tritonia PdN3, 2 s, 10 Hz treno di impulsi di 5 ms, 10 V impulsi; Aplysia PdN9, 2.5 s, 20 Hz treno di impulsi di 5 ms, 8 V impulsi; un nervo a pedale Berghia, 2 s, treno di impulsi a 10 Hz di 5 ms, impulsi a 5 V. - Ricontrolla che la molla o il tavolo dell'aria siano galleggianti.
7. Registrazione ottica
- Spegnere o attenuare le luci della stanza, compresa l'illuminazione fluorescente dall'alto.
- Impostare la durata del file desiderata, il percorso e il nome del file, quindi fare clic sul pulsante "Prendi dati" nel software di imaging per acquisire i file fino alla capacità della RAM disponibile del computer. Rimanere fermi durante la registrazione ottica, poiché piccole vibrazioni possono introdurre grandi artefatti nei dati di registrazione ottica.
NOTA: per le acquisizioni che supererebbero la RAM disponibile del computer, è disponibile un programma di acquisizione C++ personalizzato tramite il Dr. Jian-young Wu della Georgetown University. - Per visualizzare i dati immediatamente dopo l'acquisizione, utilizzare la funzione Sovrapposizione nel software di imaging per sovrapporre i dati raccolti da tutti i 464 diodi sull'immagine del ganglio scattata prima della preparazione7. Fare clic su uno qualsiasi dei diodi rappresentati nel software per espandere ciò che hanno registrato in una schermata di traccia separata.
- Ottenere l'allineamento esatto dei diodi rispetto alla preparazione inserendo i fattori x, y e di ingrandimento come precedentemente determinato dal test stenopeico.
- Per massimizzare la visibilità del potenziale d'azione e migliorare la resa dei neuroni per il successivo spike-sorting14, imporre un filtro Butterworth passa-banda con tagli a 5 Hz e 100 Hz (disponibile nel software di imaging) per rimuovere sia il rumore a bassa che ad alta frequenza.
- Per salvare i dati ottici filtrati come file di testo per ulteriori analisi in una piattaforma di calcolo scientifico, selezionare prima la casella "Filtro TP" appena sotto laschermata" Pagina " nel software di imaging. Quindi, seleziona "Salva pagina come ASCII" dalla scheda "Output" e inserisci il nome file desiderato nella finestra di dialogo che appare.
Risultati
Tritonia
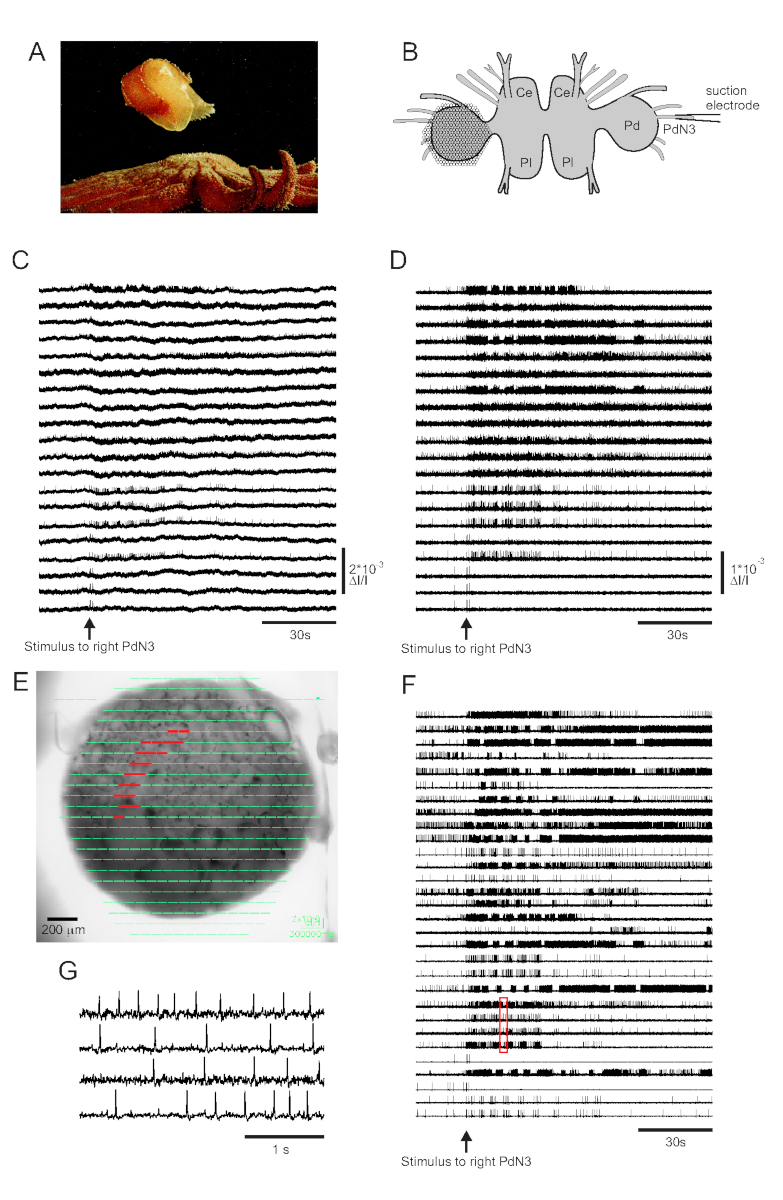
Il contatto della pelle con il suo predatore seastar innesca la nuotata di fuga di Tritonia diomedea, costituita da una serie ritmica di flessioni di tutto il corpo che la spingono verso la sicurezza (Figura 3A). Nelle preparazioni cerebrali isolate, un breve stimolo al nervo 3 del pedale (PdN3) suscita il programma motorio di nuoto ritmico (SMP) per questo comportamento, che è facilmente riconoscibile nelle registrazioni ottiche dai gangli del pedale. La Figura 3B illustra il layout di un esperimento di imaging VSD progettato per registrare l'attività di attivazione dei neuroni sulla superficie dorsale del ganglio del pedale Tritonia sinistro, su cui è stato posizionato il PDA, come stimolo al PdN3 controlaterale (a destra) che suscita l'SMP. I dati grezzi e filtrati (filtro Butterworth passabanda, cutoff a 5 e 100 Hz) dall'attività di registrazione di 20 diodi prima, durante e dopo la stimolazione di PdN3 sono mostrati rispettivamente nelle figure 3C,D. Lo stimolo nervoso è stato consegnato 20 s nel file di 2 minuti. Immediatamente dopo l'acquisizione, i segnali misurati da tutti i 464 diodi dell'array di registrazione possono essere visualizzati topograficamente su un'immagine della preparazione nel software di imaging (Figura 3E). A questo punto, molte tracce contengono picchi registrati in modo ridondante dagli stessi neuroni e alcune tracce contengono picchi da più di un neurone. Spike-sorting le tracce di diodi filtrati con ICA ha prodotto 53 tracce neuronali uniche, 30 delle quali sono mostrate nella Figura 3F. La cinetica dei singoli picchi può essere apprezzata nella Figura 3G, che espande un estratto di quattro tracce dalla Figura 3F (riquadro rosso); l'accuratezza dell'algoritmo di spike-sorting ICA è stata precedentemente verificata utilizzando registrazioni simultanee di elettrodi taglienti, che hanno dimostrato che tutti i picchi nelle tracce ordinate corrispondono a picchi registrati intracellulari da singoli neuroni11,14.
Aplysia
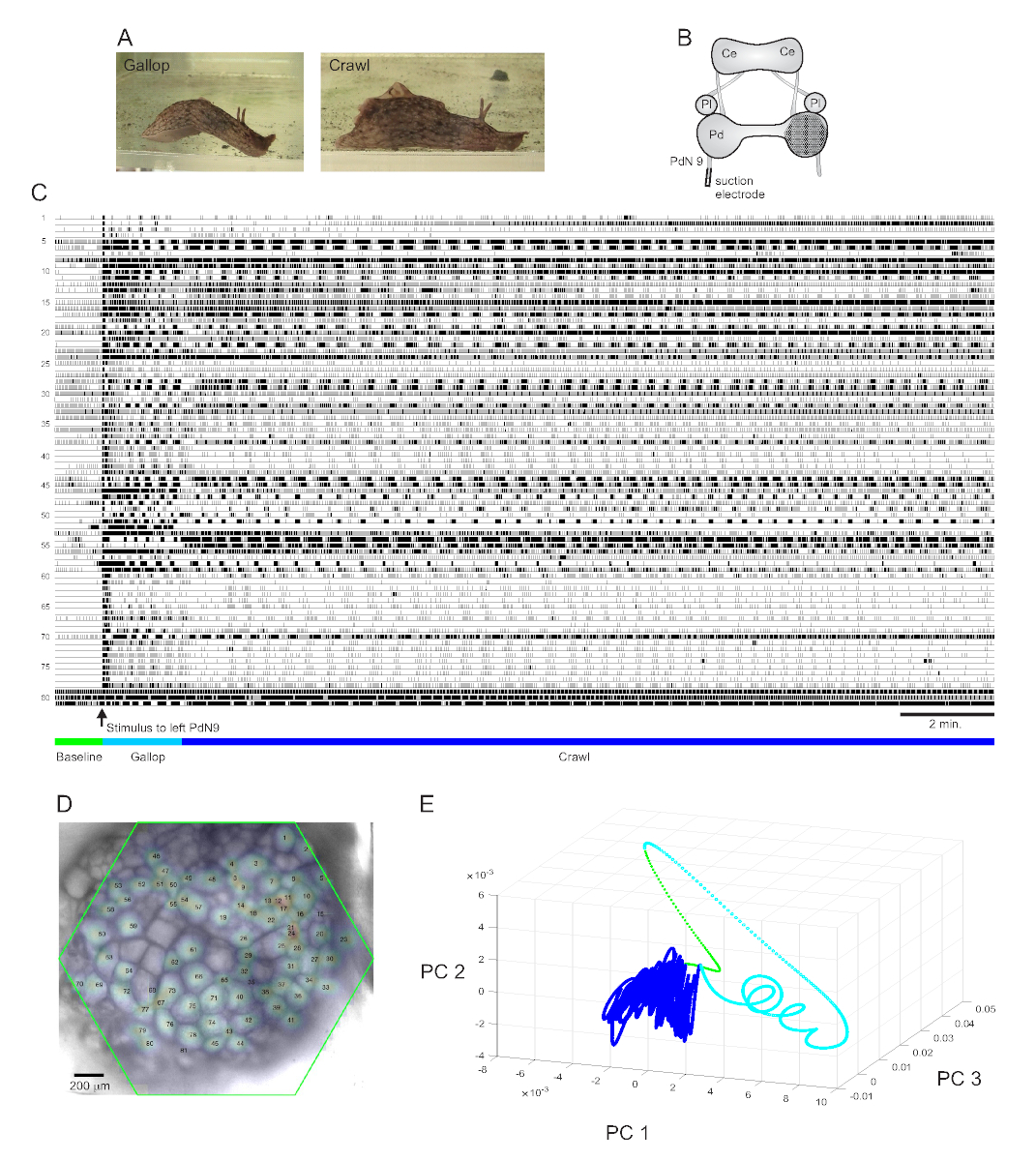
Uno stimolo di coda fortemente avversivo ad Aplysia californica suscita una risposta di fuga ritmica stereotipata in dueparti 15. La prima fase della risposta è un galoppo di diversi cicli di affondi alla testa e tiri della coda che spostano rapidamente l'animale in avanti. Questo è in genere seguito da un periodo di strisciamento, che coinvolge ripetute ondate di contrazioni muscolari testa-coda che spingono l'animale in avanti a una velocità più lenta per diversi minuti (Figura 4A). Per catturare questi programmi motori di fuga nelle registrazioni ottiche, il PDA è stato focalizzato sulla superficie dorsale del ganglio del pedale destro in una preparazione cerebrale isolata, e un elettrodo di aspirazione è stato posizionato sul nervo del pedale controlaterale (sinistro) 9 (PdN9; Figura 4B). Un minuto dopo una registrazione ottica continua di 20 minuti(Figura 4C),PdN9 è stato stimolato per suscitare la sequenza del programma motorio gallop-crawl. Le distribuzioni spaziali gaussiane probabilistiche dei segnali provenienti da tutti gli 81 neuroni registrati sono state mappate sul ganglio (Figura 4D). La riduzione della dimensionalità applicata alla registrazione completa ha rivelato che le fasi galoppo (ciano) e strisciamento (blu scuro) del programma di fuga occupavano aree distinte e formavano traiettorie diverse, simili a spirali e loop, rispettivamente, nello spazio dei componenti principali (Figura 4E).
Tre video basati sulla registrazione Aplysia raffigurata nella Figura 4 mostrano ulteriori tipi di analisi che possono essere eseguite su tali set di dati. Il video 1 anima l'attivazione di tutti i neuroni registrati per l'intera durata della registrazione. Il periodo post-stimolante iniziale del programma motorio di fuga è stato caratterizzato da un galoppo, in cui l'attività nel ganglio è stata segnata dall'alternanza di scoppi di diversi cluster funzionali (Video 2). Il galoppo successivamente è passato a un crawl, in cui l'attività attraverso i cluster neuronali è rimasta ampiamente fasica ma ha assunto una traiettoria rotazionale in senso antiorario nel ganglio (Video 3). Gli ultimi due video incorporano anche il clustering del consenso, che rivela separatamente lo sparo e le posizioni dei diversi insiemi funzionali per le fasi di galoppo e scansione della risposta di fuga. Si noti che molti neuroni assegnati allo stesso cluster in entrambe le fasi di galoppo e strisciamento hanno mostrato vicinanza fisica l'uno all'altro nel ganglio, in linea con i risultati precedenti12.
Berghia ·
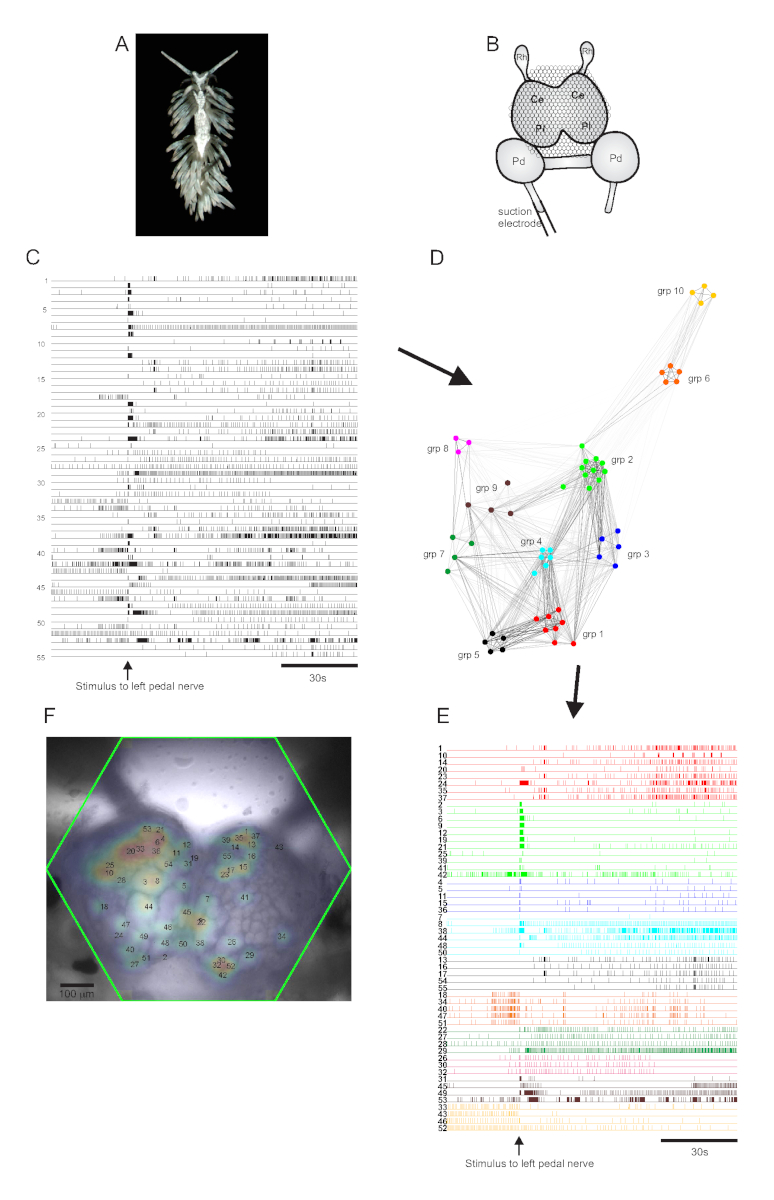
Il nudibranco eolido Berghia stephanieae (Figura 5A) rappresenta un nuovo sistema modello per le neuroscienze. La configurazione dell'imaging per un tipico esperimento berghia è mostrata nella Figura 5B. Per suscitare un'attività neuronale su larga scala, un elettrodo di aspirazione è stato posizionato sul nervo del pedale sinistro più prominente e uno stimolo nervoso è stato consegnato 30 s in una registrazione di 2 minuti. Le tracce elaborate dall'ICA hanno rivelato attività sia spontanea che evocata da stimoli in 55 neuroni (Figura 5C). Il rilevamento della comunità tramite clustering di consenso ha identificato dieci distinti insiemi funzionali, che sono rappresentati nella Figura 5D in un grafico di rete e nella Figura 5E, che riorganizza le tracce mostrate nella Figura 5C in base alle loro assegnazioni di clustering. Le distribuzioni gaussiane dei segnali provenienti da tutti i neuroni registrati sono sovrapposte a un'immagine della preparazione in Figura 5F per indicare le posizioni di tutti i 55 neuroni registrati.

Figura 1: Viste del rig di imaging ottico e del PDA (Photodiode Array). (A)Il rig di imaging ottico, con PDA, fotocamera digitale, microscopio e palcoscenico. (B) Il design interno del PDA, in cui le fibre ottiche collegano l'apertura di imaging a 464 fotodiodi. Una fila di amplificatori si trova sopra i fotodiodi. (C) La faccia esagonale dell'apertura di imaging, su cui è focalizzata l'area da immagine. Fare clic qui per visualizzare una versione più grande di questa figura.
{kind=link}

Figura 2: Diagramma di flusso che illustra il flusso di lavoro essenziale per ottenere registrazioni ottiche. I passaggi essenziali nel protocollo di imaging VSD, dalla dissezione e colorazione attraverso i dettagli dell'imaging, sono rappresentati in questo diagramma di flusso. Fare clic qui per visualizzare una versione più grande di questa figura.
{kind=link}

Figura 3: Risultati di Tritonia diomedea,che illustrano dati grezzi, filtratie ordinati con picchi. (A) Tritonia che sfugge alla stella marina predatrice Pycnopodia helianthoides attraverso la sua nuotata, che consiste nell'alternanza di flessioni dorsali e ventrali del corpo. (B) Schema della configurazione dell'immagine. Ce=lobo cerebrale del ganglio cerebropleurico; Pl = lobo pleurico del ganglio cerebropleurico; Pd = ganglio del pedale. (C) Dati grezzi da 20 fotodiodi, che mostrano l'attività nel ganglio del pedale sinistro alla stimolazione del PdN3 controlaterale (stimolo indicato dalla freccia). (D) Dati filtrati dagli stessi diodi del C (filtro Butterworth passabanda 5 e 100 Hz). (E) Uscita del software di imaging in cui le tracce compresse raccolte da tutti i 464 diodi sono sovrapposte topograficamente su un'immagine del preparato. Le posizioni dei 20 diodi le cui tracce sono mostrate in C e D sono evidenziate in rosso. (F) Trenta tracce selezionate di singolo neurone generate da spike-sorting tramite ICA. (G) Una visione espansa di quattro tracce di neuroni singoli, corrispondenti alla casella rossa in F, mostra i loro potenziali d'azione a una risoluzione temporale più elevata. Fare clic qui per visualizzare una versione più grande di questa figura.
{kind=link}

Figura 4: Risultati di Aplysia californica, che illustrano la registrazione di lunga durata, la mappatura del segnale e la riduzione della dimensionalità. (A) Le due fasi del programma motorio di fuga sequenziale di Aplysia, il galoppo e il gattonare. (B) Schema della configurazione dell'immagine. Ce = ganglio cerebrale; Pl = ganglio pleurico; Pd = ganglio del pedale. (C) Una registrazione di 20 minuti di 81 neuroni nel ganglio del pedale destro che rispondono a uno stimolo al PdN9 controlaterale (indicato dalla freccia). Le barre verdi, ciano e blu scuro sotto le tracce indicano rispettivamente il periodo di pre-stimolo, il galoppo e le fasi di scansione del programma motorio di fuga. (D) Un'immagine del preparato con distribuzioni gaussiane probabilistiche mappate delle posizioni di tutte le 81 sorgenti di segnali neuronali identificate dall'ICA. Il contorno verde rappresenta la posizione della faccia esagonale del PDA rispetto al ganglio. I numeri su ciascun gaussiano corrispondono ai numeri traccia in C. (E) Riduzione della dimensionalità mediante l'analisi dei componenti principali che traccia i primi tre componenti principali l'uno contro l'altro nel corso del file di 20 minuti. Le epoche pre-stimolatoria di base, galoppo e strisciamento sono mostrate rispettivamente in verde, ciano e blu scuro. Vedere i video 1-3 per le animazioni di attivazione neuronale corrispondenti a questa registrazione. Fare clic qui per visualizzare una versione più grande di questa figura.
{kind=link}

Figura 5: Risultati di Berghia stephanieae,una nuova specie per le neuroscienze, che illustra la rappresentazione grafica di rete, il clustering funzionalee la mappatura bilaterale del segnale. (A) Un esemplare di Berghia. (B) Schema della configurazione dell'immagine. Ce = lobo cerebrale del ganglio cerebropleurico; Pl = lobo pleurico del ganglio cerebropleurico; Pd = ganglio del pedale; Rh = ganglio rinoforo. (C) Tracce che mostrano l'attività spontanea ed evocata dallo stimolo di 55 neuroni bilaterali nei gangli cerebropleurici (la consegna dello stimolo è indicata dalla freccia). (D) Un grafico di rete che mostra i dieci insiemi funzionali, ciascuno assegnato un colore univoco, identificato attraverso il clustering del consenso. I nodi in questo grafico rappresentano i neuroni, dove la distanza nello spazio di rete rappresenta il grado di correlazione di attivazione all'interno e tra gli insiemi. (E) Le tracce in C sono riorganizzate e codificate a colori (seguendo la combinazione di colori di D) in insiemi funzionali. (F) Un'immagine del preparato che mostra le posizioni mappate dei segnali di ogni neurone registrato e i numeri di traccia in C ed E a cui corrispondono. Fare clic qui per visualizzare una versione più grande di questa figura.
{kind=link}
Video 1: Animazione del programma locomotore di fuga Aplysia completo di 20 minuti. L'opacità delle forme bianche che sovrastanti 81 singoli neuroni nel ganglio del pedale destro (pannello di sinistra) è stata guidata dalle corrispondenti tracce neuronali (pannello di destra) e variava linearmente in funzione del tasso medio di picco (legato per ogni 0,61 s di tempo reale nella registrazione). Per ogni neurone, l'opacità completa è stata normalizzata alla sua massima velocità di attivazione per tutta la durata della registrazione. Un secondo di tempo trascorso nel video rappresenta 12,2 s di tempo reale. La barra della scala corrisponde al tempo reale, con le linee verde, ciano e blu scuro sotto le tracce che indicano rispettivamente le fasi di base, galoppo e strisciamento del programma locomotore di fuga. Le caselle gialle intorno alla fase di galoppo e una parte della fase di scansione indicano gli estratti di registrazione utilizzati per generare le animazioni nei video 2 e 3. Clicca qui per vedere questo video. (Fare clic con il pulsante destro del mouse per scaricare.)
Video 2: Animazione della fase di galoppo del programma locomotore di fuga Aplysia. Il clustering di consenso è stato eseguito su tutti gli 81 neuroni registrati nella sola fase di galoppo del programma motorio per derivare gli insiemi funzionali, utilizzando l'approccio e il software descritti e resi disponibili nel rif.12. Gli insiemi neuronali che mostrano schemi di sparo in gran parte tonici o irregolari durante questa fase del programma di fuga sono stati omessi da questo video. I potenziali d'azione dei neuroni appartenenti agli insiemi nero e verde oliva possono essere ascoltati nella traccia audio del video, con i neuroni e le tracce corrispondenti evidenziati. I tassi di picco medi sono stati normalizzati come in Video 1 e con un binning temporale equivalente; 1 s di tempo trascorso nel video corrisponde a 6,1 s di tempo reale. Clicca qui per vedere questo video. (Fare clic con il pulsante destro del mouse per scaricare.)
Video 3: Animazione della fase di scansione del programma locomotore di fuga Aplysia. Il clustering di consenso è stato eseguito su tutti gli 81 neuroni registrati nella sola fase di scansione del programma motorio per derivare gli insiemi funzionali. Gli ensemble che mostrano schemi di cottura in gran parte tonici o irregolari durante questa fase del programma motorio sono stati omessi da questo video. I tassi di picco medi sono stati normalizzati come nei video 1 e 2 e con un binning temporale equivalente; 1 s di tempo trascorso nel video corrisponde a circa 12,2 s di tempo reale. Clicca qui per vedere questo video. (Fare clic con il pulsante destro del mouse per scaricare.)
Discussione
Uno dei dettagli più importanti nell'implementazione del nostro approccio di imaging VSD su larga scala è quello di ridurre al minimo le vibrazioni, che producono movimenti di bordi di contrasto attraverso i diodi, con conseguenti grandi segnali artefatti. Poiché i VSD di assorbanza producono variazioni percentuali molto piccole nell'intensità della luce con potenziali d'azione, gli artefatti di vibrazione, se non prevenuti, possono oscurare i segnali neuronali di interesse. Impieghiamo diversi metodi per ridurre al minimo gli artefatti vibrazioni. In primo luogo, la nostra sala di imaging si trova al piano terra, che isola la preparazione dalle vibrazioni legate alla costruzione di apparecchiature di trattamento dell'aria e molte altre fonti. In secondo luogo, è stata utilizzata una tabella di isolamento a molla, che altri utenti di PDA hanno confermato fornisce un migliore smorzamento delle vibrazioni rispetto alla più comune tabella dell'aria16. In terzo luogo, sono stati utilizzati obiettivi di immersione in acqua, che eliminano le fluttuazioni dell'immagine derivanti dalle increspature superficiali. In quarto luogo, la preparazione da immagine è stata leggermente premuta tra il fondo del coverslip della camera e un frammento di coverslip premuto dall'alto che è tenuto in posizione da tappi di silicone o vaselina, stabilizzando ulteriormente la preparazione. Questo appiattisce anche la superficie convessa del ganglio o dei gangli che vengono ripresi, con conseguente aumento dei neuroni nel piano di messa a fuoco dell'obiettivo, il che aumenta il numero di neuroni registrati.
Per massimizzare il rapporto segnale-rumore per le piccolissime variazioni del grado di assorbimento della luce VSD derivanti da un potenziale d'azione, è essenziale ottenere una luce quasi satura attraverso la preparazione al PDA, riducendo al minimo il fotosciviazione del colorante. A tal fine, in genere lavoriamo a 3-4 V di intensità luminosa a riposo, misurata con l'interruttore di guadagno del pannello di controllo PDA in posizione 1x (i 464 amplificatori del PDA saturano a 10 V di luce). Durante l'acquisizione dei dati questo fattore di guadagno viene modificato in 100x. Ottenere luce sufficiente per raggiungere i 3-4 V misurati dal PDA può essere realizzato in diversi modi. In primo luogo, utilizzare una sorgente luminosa a LED ultraluminosa che fornisca una lunghezza d'onda appropriata alle proprietà di assorbimento del colorante di assorbanza in uso. Di conseguenza, è stata utilizzata una lampada collimata a LED da 735 nm, che si sovrappone alle lunghezze d'onda di assorbimento ottimali di RH155 e RH482. In secondo luogo, se necessario, utilizzare un condensatore flip-top substage che concentra la luce dalla sorgente luminosa a LED in un'area più piccola. In terzo luogo, regolare l'altezza del condensatore per ottenere un'illuminazione Köhler, che garantisce una luminosità elevata e uniforme e la massima qualità dell'immagine. In quarto luogo, assicurarsi che non ci siano filtri di calore nel percorso ottico, che possono attenuare la lunghezza d'onda di 735 nm della lampada a LED. Quinto, rimuovere i diffusori, se è necessaria più luce, dal percorso ottico. Sesto, utilizzare obiettivi ad alto NA, che forniscono un'elevata risoluzione spaziale e consentono livelli sufficienti di luce per raggiungere il PDA a intensità della lampada inferiori. Questo ci ha permesso di ridurre al minimo il fotosciviazione nella misura in cui possiamo ottenere diversi file di acquisizione della durata di 10-20 minuti per preparazione utilizzando la stessa intensità luminosa su tutti i file e senza una significativa perdita di ampiezza del segnale o la necessità di ri-colorazione. Fondamentalmente, se lo sperimentatore desidera tracciare i neuroni attraverso questi file più lunghi, assicurarsi che il piano focale non cambi e che la preparazione non si muova. Infine, un ulteriore modo per instradare una luce sufficiente al PDA è quello di utilizzare animali più giovani, che hanno gangli più sottili e quindi meno opachi.
Di tanto in tanto scopriamo che il rapporto segnale-rumore dei segnali ottici si deteriora e/o i ritmi del programma motorio sono subottimali (ad esempio, lenti o anormali). Quando questo inizia a verificarsi in modo coerente, mescoliamo nuove soluzioni di VSD. Le aliquote di VSD rimangono tipicamente vitali per circa 6 mesi in un congelatore a -20 °C. In relazione a ciò, vale la pena notare che per Berghia, i migliori risultati sono stati finora ottenuti con l'assorbanza VSD RH482. Poiché RH482 è più lipofilo di RH155, potrebbe macchiare meglio i neuroni relativamente più piccoli di Berghiao rimanere nelle membrane neuronali in modo più efficace alla temperatura salina di registrazione più elevata utilizzata per questa specie tropicale.
Una limitazione dell'imaging dell'attività neurale basato su PDA riguarda l'accoppiamento AC dei segnali di tensione nell'hardware prima della fase di preamplificazione 100x: sebbene ciò rappresenti una caratteristica necessaria per rimuovere l'ampio offset DC prodotto dall'elevato livello di luce a riposo richiesto da questa tecnica, l'accoppiamento AC intrinseco al PDA preclude la misurazione di lenti cambiamenti nel potenziale di membrana, come quelli associati agli input sinaptici. Se si desidera registrare potenziali cambiamenti lenti o stazionario, è possibile utilizzare un sistema di imaging della telecamera CMOS accoppiato DC per acquisire l'attività sottosoglia. Byrne e colleghi hanno recentemente utilizzato una tale configurazione con RH155 per immaginare l'attività dei neuroni nel ganglio buccale di Aplysia17,18. Abbiamo utilizzato entrambi i sistemi e abbiamo scoperto che la telecamera CMOS, grazie alla sua densità molto più elevata di rilevatori (128 x 128), genera file di dati 50 volte più grandi per lo stesso tempo di imaging7. I file più piccoli del PDA facilitano l'elaborazione e l'analisi più veloci. Ciò consente anche registrazioni estese a prova singola (Figura 4) e studi di apprendimento, in cui i dati di più studi sono concatenati in un unico file di grandi dimensioni prima dell'ordinamento spike, consentendo di tenere traccia dell'organizzazione della rete man mano che l'apprendimento si sviluppa19.
In altre indagini basate su telecamere, i VSD fluorescenti sono stati utilizzati da Kristan e colleghi per esaminare la funzione di rete nei gangli segmentali della sanguisuga. In uno studio influente questo ha portato all'identificazione di un neurone coinvolto nella decisione dell'animale di nuotare o di gattonare20. In un altro studio, Kristan et al. hanno esaminato la misura in cui i comportamenti di nuoto e strisciamento della sanguisuga sono guidati da circuiti multifunzionali rispetto a quelli dedicati21. Più recentemente, Wagenaar e colleghi hanno utilizzato un microscopio a due lati per l'imaging della tensione che consente loro di registrare da quasi tutti i neuroni in un ganglio segmentale di sanguisuga22. A differenza di molti metodi di imaging basati su telecamera, un vantaggio del nostro metodo di imaging basato su PDA è l'ordinamento rapido e imparziale dei picchi da parte di ICA, una forma di separazione cieca della sorgente che non comporta decisioni sui confini neuronali per l'elaborazione dei risultati.
Per quanto riguarda la scelta dei VSD, un vantaggio dei coloranti ad assorbanza RH155 e RH482 è la fototossicità poco o nulla associata a loro23, 24,consentendo tempi di registrazione più lunghi di quelli tipici dei VSD fluorescenti. Inoltre, i VSD ad assorbimento rapido che utilizziamo sono adatti per registrare i potenziali d'azione somatici di overshooting nei preparati gasteropodi, che sono tipicamente 80 mV di ampiezza. Come mostrato nella Figura 3G,il nostro metodo ottico può registrare i potenziali d'azione (nessuna delle nostre registrazioni è tracciata): questo suggerisce che i VSD che usiamo dovrebbero essere in grado di discernere i potenziali d'azione in altri sistemi modello che si attenuano in una certa misura e quindi non sono in overshooting nel momento in cui raggiungono il soma. Tuttavia, il nostro approccio ottico potrebbe non essere l'ideale per le specie che sono note per esibire potenziali d'azione altamente attenuati quando registrate nel soma.
Gran parte della ricerca attuale sulle reti neurali si sta concentrando su un piccolo numero di specie transgeniche di design. Tuttavia, le neuroscienze traggono beneficio dallo studio di un'ampia varietà di specie filogeneticamente distinte. Lo studio di molte specie diverse fornisce approfondimenti su come i circuiti si evolvono25,26e illumina i principi della funzione di rete che possono essere comuni tra phyla1,2,3,4,27. Finora abbiamo applicato il nostro metodo di imaging a un certo numero di specie di gasteropodi, tra cui Aplysia californica8,11,12,13 ,14,28, Tritonia diomedea8,9,11,14,19,28, Tritonia festiva28, Pleurobranchaea californica (dati non pubblicati), e più recentemente Berghia stephanieae (Figura 5). Un fascino di questo approccio è che può essere facilmente applicato a molte specie, senza bisogno di animali transgenici. Desideriamo riconoscere che il nostro uso dell'imaging VSD con coloranti ad assorbimento rapido e un PDA segue le orme del lavoro pionieristico che ha raggiunto questo obiettivo in preparati Semi-intatti e comportandosi con Navanax29 e Aplysia30. La nostra enfasi sulla rapidità del nostro approccio è in parte una risposta alle preoccupazioni che molti ricercatori potrebbero essere sempre più riluttanti ad avviare studi di rete in nuove specie a causa dei timori che saranno necessari anni di studio per caratterizzare l'organizzazione di base della rete prima di essere in grado di esplorare questioni scientifiche di ampio interesse per le neuroscienze31. Di conseguenza, il nostro obiettivo qui è quello di dimostrare una tecnica che accelera notevolmente il processo, al punto che da singoli preparativi è possibile ottenere informazioni significative in giornata sull'organizzazione della rete.
Divulgazioni
Gli autori non hanno nulla da rivelare.
Riconoscimenti
Questo lavoro è stato supportato da NSF 1257923 e NIH 1U01NS10837. Gli autori desiderano riconoscere l'assistenza di Jean Wang in laboratorio.
Materiali
| Name | Company | Catalog Number | Comments |
| Achromat 0.9 NA swing condenser | Nikon | N/A | |
| Bipolar temperature controller | Warner Instruments | CL-100 with SC-20 | Controls perfusion saline temperature |
| Chamber thermometer | Physitemp | BAT-12 with IT-18 microprobe | |
| Digital camera | Optronics | S97808 | |
| Dissecting forceps | Dumont | #5 | |
| Dissecting scissors | American Diagnostic Corp. | ADC-3410Q | |
| Imaging microscope | Olympus | BX51WIF | |
| Imaging perfusion chamber | Siskiyou | PC-H | |
| Instant Ocean | Instant Ocean | SS6-25 | Makes 25 gallons at a time |
| Master-8 pulse stimulator | A.M.P.I. | Master-8 | |
| Microdispenser | Drummond Scientific | 3-000-752 | Dye applicator for pressure staining |
| Microdissection scissors | Moria | 15371-92 | |
| Minutien pins (0.1 mm) | Fine Science Tools | NC9677548 | For positioning and stabilizing CNS |
| Motorized microscope platform | Thorlabs | GHB-BX | Gibraltar platform |
| NeuroPlex imaging software | RedShirtImaging | NeuroPlex | Compatible with the WuTech photodiode array |
| Objective lenses | Olympus | XLPLN10XSVMP, XLUMPLFLN20XW, LUMPLFLN40XW, UAPON40XW340 | |
| PE-100 polyethylene tubing | VWR | 63018-726 | Tubing to make suction electrodes |
| Perfusion pump | Instech | P720 with DBS062SDBSU tube set | |
| Petroleum jelly | Equate | NDC 49035-038-54 | |
| Photodiode array with control panel | WuTech Instruments | 469-IV photodiode array | Contact jianwu2nd@gmail.com for ordering information |
| RH155 | Santa Cruz Biotechnology | sc-499432 | Voltage-sensitive dye |
| RH482 | Univ of Conn. Health Center | JPW-1132 | Voltage-sensitive dye; special order from Leslie Leow |
| Silicone earplugs | Mack's | Model 7 | To be use for preparation compression |
| Staining PE tubing | VWR | 63018-xxx | Different sizes depending on fit |
| Sylgard 184 silicone elastomer kit | Dow Corning | Sylgard 184 silicone elastomer kit | |
| Thorlabs LED and driver | Thorlabs | M735L2-C1, DC2100 | LED lamp and driver |
| Tygon tubing | Fisher Scientific | 14-171-xxx | |
| Vibration isolation table | Kinetic Systems | MK26 | Spring-based |
Riferimenti
- Miller, C. T., Hale, M. E., Okano, H., Okabe, S., Mitra, P. Comparative Principles for Next-Generation Neuroscience. Frontiers in Behavioral Neuroscience. 13 (12), (2019).
- Brenowitz, E. A., Zakon, H. H. Emerging from the bottleneck: benefits of the comparative approach to modern neuroscience. Trends in Neuroscience. 38 (5), 273-278 (2015).
- Bolker, J. Model organisms: There's more to life than rats and flies. Nature. 491 (7422), 31-33 (2012).
- Carlson, B. A. Diversity matters: the importance of comparative studies and the potential for synergy between neuroscience and evolutionary biology. JAMA Neurology. 69 (8), 987-993 (2012).
- Chase, R. . Behavior and its neural control in gastropod molluscs. , (2002).
- Salzberg, B. M., Grinvald, A., Cohen, L. B., Davila, H. V., Ross, W. N. Optical recording of neuronal activity in an invertebrate central nervous system: simultaneous monitoring of several neurons. Journal of Neurophysiology. 40 (6), 1281-1291 (1977).
- Frost, W. N., et al. Monitoring Spiking Activity of Many Individual Neurons in Invertebrate Ganglia. Advances in Experimental Medicine and Biology. 859, 127-145 (2015).
- Frost, W. N., Wang, J., Brandon, C. J. A stereo-compound hybrid microscope for combined intracellular and optical recording of invertebrate neural network activity. Journal of Neuroscience Methods. 162 (1-2), 148-154 (2007).
- Frost, W. N., Wu, J. -. Y., Covey, E., Carter, M. Voltage-sensitive dye imaging. Basic Electrophysiological Methods. , 169-195 (2015).
- Brown, G. D., Yamada, S., Sejnowski, T. J. Independent component analysis at the neural cocktail party. Trends in Neuroscience. 24 (1), 54-63 (2001).
- Hill, E. S., Moore-Kochlacs, C., Vasireddi, S. K., Sejnowski, T. J., Frost, W. N. Validation of independent component analysis for rapid spike sorting of optical recording data. Journal of Neurophysiology. 104 (6), 3721-3731 (2010).
- Bruno, A. M., Frost, W. N., Humphries, M. D. Modular deconstruction reveals the dynamical and physical building blocks of a locomotion motor program. Neuron. 86 (1), 304-318 (2015).
- Bruno, A. M., Frost, W. N., Humphries, M. D. A spiral attractor network drives rhythmic locomotion. ELife. 6, 27342 (2017).
- Hill, E. S., Bruno, A. M., Vasireddi, S. K., Frost, W. N., Naik, G. R. ICA applied to VSD imaging of invertebrate neuronal networks. Independent Component Analysis for Audio and Biosignal Applications. , 235-246 (2012).
- Jahan-Parwar, B., Fredman, S. M. Neural control of locomotion in Aplysia: role of the central ganglia. Behavioral and Neural Biology. 27 (1), 39-58 (1979).
- Jin, W., Zhang, R. J., Wu, J. Y. Voltage-sensitive dye imaging of population neuronal activity in cortical tissue. Journal of Neuroscience Methods. 115 (1), 13-27 (2002).
- Neveu, C. L., et al. Unique Configurations of Compression and Truncation of Neuronal Activity Underlie l-DOPA-Induced Selection of Motor Patterns in Aplysia. eNeuro. 4 (5), 17 (2017).
- Cai, Z., Neveu, C. L., Baxter, D. A., Byrne, J. H., Aazhang, B. Inferring neuronal network functional connectivity with directed information. Journal of Neurophysiology. 118 (2), 1055-1069 (2017).
- Hill, E. S., Vasireddi, S. K., Wang, J., Bruno, A. M., Frost, W. N. Memory Formation in Tritonia via Recruitment of Variably Committed Neurons. Current Biology. 25 (22), 2879-2888 (2015).
- Briggman, K. L., Abarbanel, H. D., Kristan, W. B. Optical imaging of neuronal populations during decision-making. Science. 307 (5711), 896-901 (2005).
- Briggman, K. L., Kristan, W. B. Imaging dedicated and multifunctional neural circuits generating distinct behaviors. Journal of Neuroscience. 26 (42), 10925-10933 (2006).
- Tomina, Y., Wagenaar, D. A. A double-sided microscope to realize whole-ganglion imaging of membrane potential in the medicinal leech. ELife. 6, 29839 (2017).
- Chang, P. Y., Jackson, M. B. Interpretation and optimization of absorbance and fluorescence signals from voltage-sensitive dyes. Journal of Membrane Biology. 196 (2), 105-116 (2003).
- Parsons, T. D., Salzberg, B. M., Obaid, A. L., Raccuia-Behling, F., Kleinfeld, D. Long-term optical recording of patterns of electrical activity in ensembles of cultured Aplysia neurons. Journal of Neurophysiology. 66, 316-333 (1991).
- Katz, P. S. Evolution of central pattern generators and rhythmic behaviours. Transactions of the Royal Society of London, Series B. 371 (1685), 20150057 (2016).
- Moroz, L. L. Biodiversity Meets Neuroscience: From the Sequencing Ship (Ship-Seq) to Deciphering Parallel Evolution of Neural Systems in Omic's Era. Integrative and Comparative Biology. 55 (6), 1005-1017 (2015).
- Frost, W. N., Tian, L. -. M., Hoppe, T. A., Mongeluzi, D. L., Wang, J. A cellular mechanism for prepulse inhibition. Neuron. 40, 991-1001 (2003).
- Hill, E. S., Vasireddi, S. K., Bruno, A. M., Wang, J., Frost, W. N. Variable neuronal participation in stereotypic motor programs. PLoS One. 7 (7), 40579 (2012).
- London, J. A., Zecevic, D., Cohen, L. B. Simultaneous optical recording of activity from many neurons during feeding in Navanax. Journal of Neuroscience. 7 (3), 649-661 (1987).
- Wu, J., Cohen, L. B., Falk, C. X. Neuronal activity during different behaviors in Aplysia: A distributed organization. Science. 263 (5148), 820-823 (1994).
- Marder, E., North, G., Greenspan, R. J. Searching for insight. In Invertebrate Neurobiology. , 1-18 (2007).
Ristampe e Autorizzazioni
Richiedi autorizzazione per utilizzare il testo o le figure di questo articolo JoVE
Richiedi AutorizzazioneThis article has been published
Video Coming Soon
Personale delle biblioteche
Copyright © 2025 MyJoVE Corporation. Tutti i diritti riservati