
Method Article
Sonde tether per tensiometri per quantificare la meccanica e l'adesione dell'integrina mediata dal fattore di crescita
In questo articolo
Riepilogo
TGT surface è una piattaforma innovativa per studiare il crosstalk fattore di crescita-integrina. Il design flessibile della sonda, la specificità del ligando di adesione e la precisa modulazione delle condizioni di stimolazione consentono solide valutazioni quantitative dell'interazione EGFR-integrina. I risultati evidenziano EGFR come un "meccano-organizzatore" che sintonizza la meccanica dell'integrina, influenzando l'assemblaggio dell'adesione focale e la diffusione cellulare.
Abstract
Gli organismi multicellulari si basano sulle interazioni tra recettori di membrana e ligandi affini nella matrice extracellulare circostante (ECM) per orchestrare molteplici funzioni, tra cui adesione, proliferazione, migrazione e differenziazione. Le forze meccaniche possono essere trasmesse dalla cellula attraverso l'integrina del recettore di adesione ai ligandi nell'ECM. La quantità e l'organizzazione spaziale di queste forze generate dalle cellule possono essere modulate dai recettori del fattore di crescita, incluso il recettore del fattore di crescita epidermico (EGFR). Gli strumenti attualmente disponibili per quantificare i cambiamenti mediati da crosstalk nella meccanica cellulare e metterli in relazione con le aderenze focali, la morfologia cellulare e la segnalazione sono limitati. I sensori di forza molecolare basati sul DNA noti come tension gauge tethers (TGT) sono stati impiegati per quantificare questi cambiamenti. Le sonde TGT sono uniche nella loro capacità di modulare la soglia di forza sottostante e di riportare le forze del recettore della scala piconewton su tutta la superficie cellulare aderente a risoluzione spaziale limitata dalla diffrazione. Le sonde TGT qui utilizzate si basano sulla dissociazione irreversibile di un duplex del DNA da parte delle forze recettore-ligando che generano un segnale fluorescente. Ciò consente la quantificazione della tensione cumulativa dell'integrina (storia della forza) della cellula. Questo articolo descrive un protocollo che impiega TGT per studiare l'impatto dell'EGFR sulla meccanica dell'integrina e sulla formazione dell'adesione. L'assemblaggio della piattaforma di rilevamento meccanico TGT è sistematicamente dettagliato e viene delineata la procedura per l'immagine delle forze, le aderenze focali e la diffusione cellulare. Nel complesso, la capacità di modulare la soglia di forza sottostante della sonda, il ligando di adesione e il tipo e la concentrazione del fattore di crescita impiegato per la stimolazione rendono questa una solida piattaforma per studiare l'interazione di diversi recettori di membrana nella regolazione delle forze mediate dall'integrina.
Introduzione
Le cellule hanno la capacità intrinseca di percepire, generare e rispondere alle forze meccaniche, portando a cambiamenti nel fenotipo cellulare e al rimodellamento del microambiente locale 1,2. Le forze svolgono un ruolo cruciale nella regolazione di molti aspetti del comportamento cellulare, tra cui adesione, migrazione, proliferazione, differenziazione e guarigione delle ferite 3,4. Le aberrazioni nello scambio meccanico bidirezionale tra una cellula e il microambiente possono portare a stati patologici, incluso il cancro5. Numerosi recettori di membrana sono coinvolti nel mantenimento dell'omeostasi della matrice cellulare; di questi, le integrine e il recettore del fattore di crescita epidermico (EGFR) hanno una robusta sinergia 6,7. Classicamente, le integrine stabiliscono il legame meccanico tra il microambiente e il citoscheletro intracellulare mentre l'EGFR regola la crescita, la proliferazione e la sopravvivenza cellulare 8,9. L'EGFR è un bersaglio terapeutico altamente studiato, focalizzato sulla regolazione esterna-in che facilita la segnalazione intracellulare. Il crosstalk EGFR-integrina è stato stabilito geneticamente e biochimicamente per regolare la progressione di più malattie, incluso il cancro10,11. Mentre gli studi indicano l'esistenza dell'interazione EGFR-integrina, i risultati sono attribuiti a vie di segnalazione lontano dalla membrana plasmatica 7,12,13,14. L'impatto dell'EGFR, o di altri fattori di crescita, sulla meccanica cellulare rimane in gran parte inesplorato in parte a causa della mancanza di strumenti per misurare le forze cellulari e i risultati di segnalazione. La sfida sta nell'identificare strumenti appropriati per studiare la comunicazione tra questi paradigmi di segnalazione parallela e per quantificare i loro contributi specifici alla meccanica cellulare.
Sono stati sviluppati diversi approcci per misurare le forze generate dai recettori di adesione cellulare e il lettore è indirizzato a revisioni approfondite di queste tecniche15,16. In breve, la microscopia della forza di trazione e il rilevamento di array di micropilastri si basano sulla deformazione di un substrato sottostante per dedurre forze nanonewton (nN), un ordine di grandezza superiore alle singole forze recettoriali17,18. Le tecniche a singola molecola, tra cui AFM e pinzette ottiche, sono sensibili alle forze piconewton a singola proteina (pN) ma misurano solo un recettore alla volta e non offrono una buona (o qualsiasi) risoluzione spaziale. Le sonde di tensione molecolare basate sul DNA e le sonde TGT (Tension Gauge Tether) offrono una risoluzione della forza pN con risoluzione spaziale limitata alla diffrazione (o migliore), conferendo loro un ruolo unico nello studio delle forze unicellulari19,20 di diversi tipi di cellule, tra cui fibroblasti, cellule tumorali, piastrine e cellule immunitarie 21,22,23,24 . Mentre le sonde di tensione molecolare hanno un elemento "a molla" estensibile, ideale per l'imaging in tempo reale, le sonde TGT si rompono irreversibilmente, lasciando dietro di sé una "storia di forza" fluorescente. I TGT modulano inoltre la soglia di tensione del substrato sottostante; una serie di sonde con composizioni chimiche simili ma diverse forze di rottura, otolleranze di tensione (T tol), possono essere utilizzate per quantificare la tensione minima richiesta per la formazione dell'adesione focale e la diffusione cellulare. Le sonde TGT sono costituite da due filamenti di DNA complementari, uno ancorato alla superficie e l'altro che presenta un ligando alla cellula. Se un recettore lega il ligando ed esercita una forza maggiore delT tol della sonda, i fili saranno separati. Ttol è definito come la forza costante necessaria per rompere il 50% delle sonde in un intervallo di 2 s in condizioni ideali. Nelle sonde TGT "turn-on", un quencher sul filamento superiore può essere separato da un fluoroforo sul filamento inferiore. Solo dove la sonda TGT è stata rotta, presumibilmente da forze maggiori o uguali a Ttol, verrà generato un segnale fluorescente. Le sonde TGT possono anche essere riparate, consentendo una facile manipolazione di sistemi biologici e test di più condizioni. Per questi motivi, le sonde TGT sono state utilizzate in questo lavoro.
Le sonde TGT sono state impiegate per studiare come l'adesione cellulare integrina-dipendente e le forze meccaniche sono modulate dall'EGFR21 attivato. Questo lavoro ha stabilito EGFR come un "mechano-organizer", ottimizzando l'organizzazione dell'adesione focale e la generazione di tensione. Inoltre, è stato riscontrato che la stimolazione EGF ha influenzato la distribuzione e la maturità delle aderenze focali e una maggiore diffusione cellulare. Questo approccio potrebbe essere utilizzato in studi futuri per studiare come i fattori di crescita influenzano le forze meccaniche nella progressione e nella dinamica del tumore. Mentre il ruolo della diafonia EGFR-integrina nella regolazione della transizione epiteliale-mesenchimale è stabilito, il ruolo delle forze meccaniche in questo processo rimane poco esplorato10.
Qui, viene presentato un protocollo dettagliato per questi esperimenti che copre la sintesi e l'assemblaggio di sonde TGT 56 pN, la generazione di superfici TGT su coperture di vetro, l'applicazione di cellule Cos-7 sulla superficie TGT e la stimolazione con EGF, fissazione e colorazione di cellule con falloidina e un anticorpo anti-paxillina, fluorescenza a riflessione interna totale ad alta risoluzione (TIRF) e microscopia a contrasto di interferenza di riflessione (RICM) imaging, e quantificazione delle immagini. Questo protocollo, sebbene scritto per studiare la stimolazione EGF delle cellule Cos-7, è facilmente adattabile per molti esperimenti basati su TGT. Diversi ligandi, Ttol, tipi di cellule, parametri di stimolazione, proteine etichettate dopo la fissazione e analisi quantitativa possono essere facilmente sostituiti, rendendo questo protocollo robusto e ampiamente utile.
Protocollo
1. Preparazione di oligonucleotidi TGT
NOTA: I dettagli della sintesi della sonda oligonucleotidica sono descritti qui. Si prega di notare che alcune modifiche e passaggi di purificazione possono essere esternalizzati per la sintesi personalizzata.
- Attivare l'ammina primaria del peptide ciclo[Arg-Gly-Asp-D-Phe-Lys(PEG-PEG)] con il linker azide-NHS come descritto da Zhang et al22 mescolando in un rapporto 1:1,5 (100:150 nmoli) in un volume finale di 10 μL di dimetilformammide. Aggiungere 0,1 μL di trietilammina di base organica e incubare per 12 ore a 4° C.
- Purificare il prodotto mediante HPLC in fase inversa utilizzando 0,1 M TEAA (solvente A) e 100% acetonitrile (solvente B) con una portata di 1mL/min e la condizione iniziale del 10% solvente B impostata ad un gradiente dello 0,5%/min. Combinare i picchi eluiti (assorbanza a 203 nm) e verificare mediante spettrometria di massa MALDI-TOF. Il prodotto è cRGDfK-azide.
- Per generare il filamento superiore TGT, combinare cRGDfK-azide e oligonucleotide alchinico-21-BHQ2 (filamento superiore TGT: 5Hexynyl/GTGAAATACCGCACAGATGCG/3BHQ_2) in un rapporto 2:1 (͂200 μM: 100 μM) in 100 μL di 1x Phosphate Buffered Saline (PBS) con 5 mM di ascorbato di sodio e 0,1 μM di Cu-THPTA preformato. Lasciare che la reazione proceda per un minimo di 4 ore a temperatura ambiente (RT) o durante la notte a 4 °C.
- Elaborare la miscela attraverso il gel dissalazione P2 per rimuovere il colorante in eccesso, i sottoprodotti, il solvente organico e i reagenti non reagiti. Preparare la colonna della centrifuga con 650 μL di gel P2 pre-idratato ruotandola a 18.000 x g per 1 min. Scartare il liquido di flusso e aggiungere la miscela di reazione. Ruotare di nuovo a 18.000 x g per 1 minuto e raccogliere il flusso attraverso. Portare la miscela di reazione ad un volume finale di 300 μL con acqua ultrapura.
NOTA: Pre-idratare il gel P2 con acqua per 6 ore. - Purificare la miscela di reazione dissalata mediante HPLC in fase inversa. I solventi organici utilizzati per questa purificazione includono 0,1 M TEAA in H2O (solvente A) e 100% MeCN (solvente B, o fase mobile).
- Prima di iniettare la miscela, equilibrare la colonna con una condizione iniziale del 10% di solvente B con un gradiente dell'1%/min. Regolare la portata a 1 mL/min. Iniettare la miscela di reazione nel circuito HPLC con un ago per iniezione da 500 μL.
- Raccogliere il prodotto visualizzando il picco di assorbimento a 260 nm per il DNA e 560 nm per il quencher BHQ2. Asciugare il prodotto eluito durante la notte in un concentratore centrifugo sotto vuoto.
- Impiegare la sostituzione nucleofila per accoppiare il filamento inferiore TGT all'estere Cy3B-NHS come descritto in Ma et. al25. Miscelare 100 μM del filamento inferiore TGT da 56 pN (5Biosg/TTTTTT/iUniAmM/CGCATCTGTGCGGTATTTCACTTT) con 50 μg di estere Cy3B-NHS pre-disciolto in 10 μL di DMSO. Regolare il pH di questa miscela a 9 con bicarbonato di sodio 0,1 M e portare il volume finale a 100 μL con 1x PBS. Incubare la miscela di reazione durante la notte a RT.
- Purificare la miscela in sequenza utilizzando la filtrazione su gel P2 e l'HPLC in fase inversa per separare reagenti, sali e solventi organici non reagiti (descritti nei passaggi 1.4 e 1.5).
- Stimare la concentrazione dei coniugati oligonucleotide-colorante purificati registrando la loro assorbanza a 260 nm utilizzando uno spettrofotometro.
- Caratterizzare i prodotti purificati mediante spettrometria di massa MALDI-TOF. Sciogliere l'acido 3-idrossipicolinico in eccesso in solvente TA50 (50:50 v/v acetonitrile e 0,1% TFA in ddH2O) per preparare la matrice MALDI fresca. I pesi molecolari stimati e misurati per i prodotti etichettati sono: cRGDfK-1-BHQ2 - 8157.9 (calcolato), 8160.1 (trovato); Cy3B etichettato 56 pN TGT - 10272.7 (calcolato), 10295.8 (trovato).
- Sciogliere separatamente i fili superiore e inferiore in acqua priva di nucleasi ad una concentrazione compresa tra 30-50 μM. Utilizzare punte per pipette prive di DNasi per evitare la contaminazione delle scorte. Conservare a 4 °C per applicazioni a breve termine o -20 °C per applicazioni a lungo termine. La stabilità degli oligonucleotidi non è influenzata da ripetuti cicli di congelamento-scongelamento.
2. Preparazione della superficie
Giorno 1:
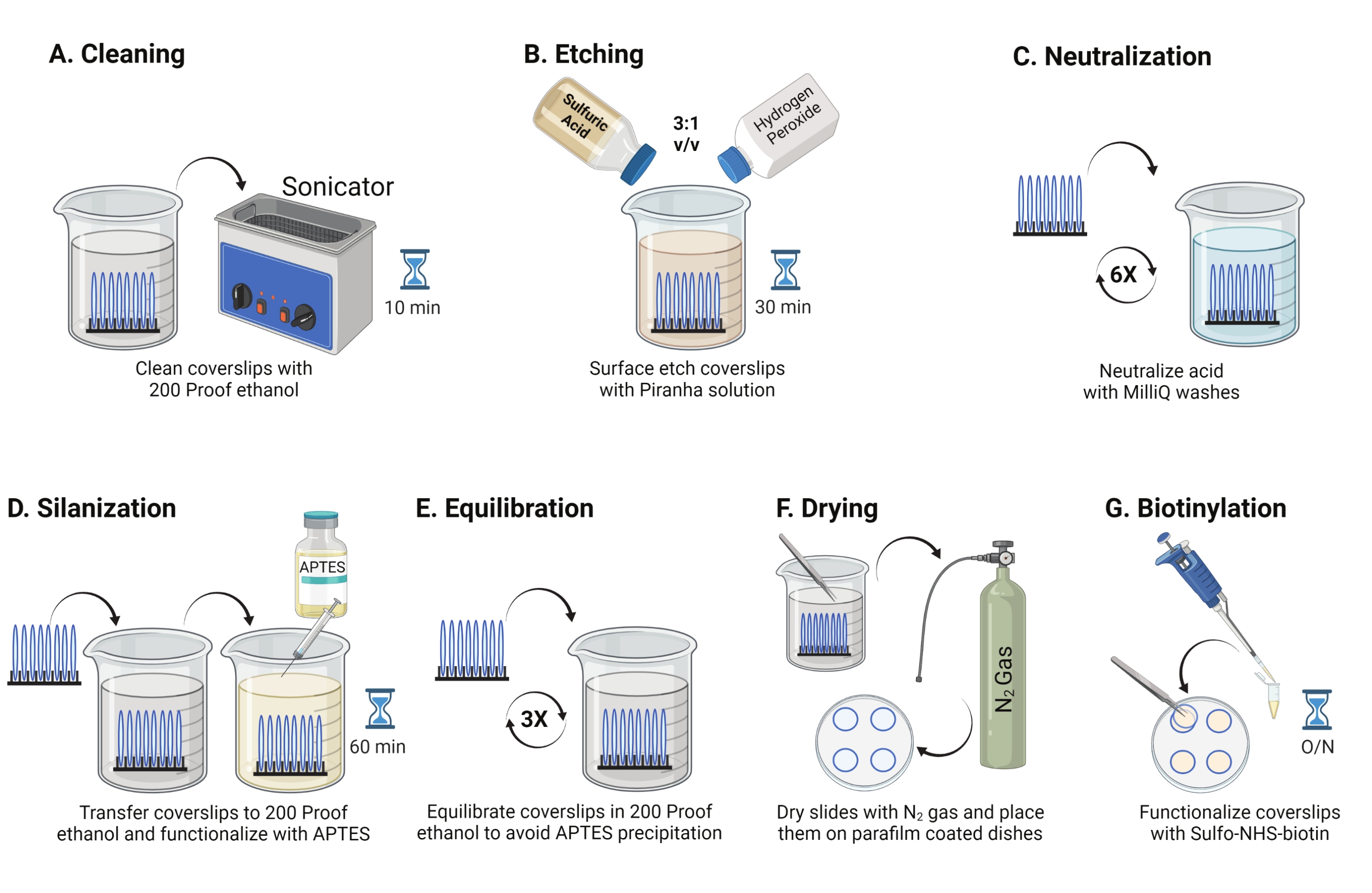
- Posizionare i coperchi di vetro da 25 mm (fino a 8) in un rack in politetrafluoroetilene. Posizionare il rack in un becher borosilicato da 50 mL contenente 40 mL di etanolo a prova di 200 ml. Coprire il becher con pellicola di paraffina per evitare l'ingresso di acqua e sonicare ad una frequenza operativa di 35 kHz per 10-15 minuti a RT (Figura 1A).
- Riempire un becher da 50 mL con 40 mL di soluzione di Piranha preparata al momento mescolando acido solforico e perossido di idrogeno in un rapporto 3:1 in un becher di pyrex. Mescolare con una pipetta di vetro. Trasferire il rack coverslip nel becher e incubare per 30 minuti a RT nella cappa aspirante per incidere la superficie del coverslip (Figura 1B).
NOTA: indossare DPI completi, tra cui un cappotto da laboratorio, guanti e occhiali, e lavorare nel cappuccio dei fumi chimici. Aggiungere lentamente il perossido di idrogeno all'acido per evitare il surriscaldamento della soluzione. - Dopo l'incisione, utilizzare una pinzetta per trasferire il rack coverslip in un becher con acqua ultrapura. Ripetere questo passaggio sei volte a intervalli di 15 s per neutralizzare completamente l'acido (Figura 1C).
NOTA: Lasciare la soluzione di Piranha nella cappa chimica durante la notte prima di scartarla nel contenitore dei rifiuti acidi. - Ispezionare visivamente le coperture per assicurarsi che le superfici appaiano pulite senza motivi o particelle di polvere sulla superficie del vetro. Ripetere i passaggi 2.1-2.4 se vengono rilevati modelli o polvere.
NOTA: Testare l'idrofilia superficiale immergendo i coperchi trattati in acqua e rimuovendoli verticalmente. L'acqua sulle coperture trattate si ritira come un foglio uniforme per formare gli anelli di Young rispetto alle coperture non trattate che formano patch. - Trasferire il rack coverslip in un becher con etanolo a prova di 200 e lavare due volte per 15 s per equilibrare le superfici al solvente organico. Trasferire il rack coverslip in una soluzione di etanolo a prova di 200 con APTES al 3% per 1 ora a RT per silanizzare i coverslip (Figura 1D). Coprire il becher con pellicola di paraffina.
NOTA: i parametri di deposizione aptes variano a seconda del metodo di pulizia della superficie, del contenuto di acqua solvente, della concentrazione di APTES, dei tempi di incubazione e della temperatura per la ricottura. - Immergere il rack in un becher pulito con una soluzione di etanolo a prova di 200. Ripetere questo lavaggio tre volte per 15 s ciascuno (Figura 1E).
- Asciugare i coperchi utilizzando gas azoto (N2) con bassa pressione di uscita. Posizionare le coperture in un piatto di polistirolo di 10 cm con un pezzo di pellicola di paraffina posato piatto al suo interno. Assicurarsi che le coperture siano asciutte e separate (Figura 1F).
- Aggiungere 100 μL di 2 mg/mL di soluzione di NHS-biotina in DMSO a quattro coverslip posti su pellicola di paraffina. Impostare un "sandwich" con gli altri quattro coverslips in cima (due coverslip rivolti l'uno verso l'altro con la soluzione di funzionalizzazione in mezzo) e incubare il piatto durante la notte a 4 °C (Figura 1G).
NOTA: a 4 °C, il reagente NHS è più stabile, il che facilita la funzionalizzazione uniforme della superficie. Inoltre, il sandwich conserva i reagenti. Evitare di aggiungere una soluzione in eccesso nel sandwich in quanto potrebbe fuoriuscire e far scivolare le coperture.
Giorno 2:
- Togliere il piatto da 4 °C e separare i coperchi a sandwich. Orientare gli slip nel rack con la superficie rivestita l'una di fronte all'altra, come mostrato nella Figura 2A. Lavali con 200 soluzioni di etanolo a prova di tre volte per 15 s ciascuno. Asciugare con gas N2 e metterli in un nuovo piatto con un film di paraffina al suo interno.
NOTA: l'orientamento delle coverslip come indicato aiuta a identificare la superficie funzionalizzata. - Lavare le coperture con 1 mL di 1x PBS tre volte per riportarle alla fase acquosa. Aggiungere 800 μL di albumina sierica bovina allo 0,1% (BSA) in 1x PBS (p/v) a ciascuno dei coverslip e incubare a RT per 30 minuti per passivare la superficie e bloccare il legame non specifico dei reagenti di funzionalizzazione successiva (Figura 2B).
- Dopo l'incubazione, lavare i coverslip tre volte con 1 mL di 1x PBS. Aggiungere 800 μL di 1 μg/mL di streptavidina in 1x PBS a RT per 45-60 min per funzionalizzare i coverslip (Figura 2C).
NOTA: Mantenere un coperchio senza streptavidina per verificare l'efficienza di passivazione (opzionale). Aggiungi10 nM molecole biotinilate e immagine utilizzando condizioni sperimentali. Questa intensità superficiale dovrebbe essere vicina al rumore scuro della fotocamera. - Contemporaneamente al passo 2.11, assemblare le sonde TGT (in alto: filo inferiore con rapporto molare 1:1) ad una concentrazione finale di 50 nM in 100 μL di 1 M NaCl in un tubo PCR utilizzando un termiciclo. Dissociare i fili a 95 °C per 5 minuti e ricottura gradualmente riducendo la temperatura a 25 °C e mantenendola per 25 minuti (Figura 2D). Evitare l'esposizione prolungata delle sonde TGT alla luce.
- Dopo l'incubazione di streptavidina, utilizzare 1x PBS per lavare i coverslip tre volte. Aggiungere 100 μL delle sonde TGT preassemblate a quattro dei coverslip e creare sandwich utilizzando i restanti 4 coverslip con il lato funzionalizzato rivolto verso le sonde (otto superfici richiedono quattro tubi di sonde TGT ibridate). Coprire con un foglio di alluminio e incubare per 1 ora a RT per consentire il legame della sonda alla superficie (Figura 2E).
- Dopo l'incubazione, separare i panini e lavare i coverslip con 1x PBS tre volte. Le superfici TGT sono ora pronte per l'imaging. Assemblare con cura i coverslip in camere di imaging prepulite e aggiungere 1x PBS per mantenere le superfici idratate (Figura 2F).
NOTA: il serraggio eccessivo delle camere spezzerà la superficie. Prevenire l'essiccazione delle superfici.
3. Preparazione cellulare e colorazione
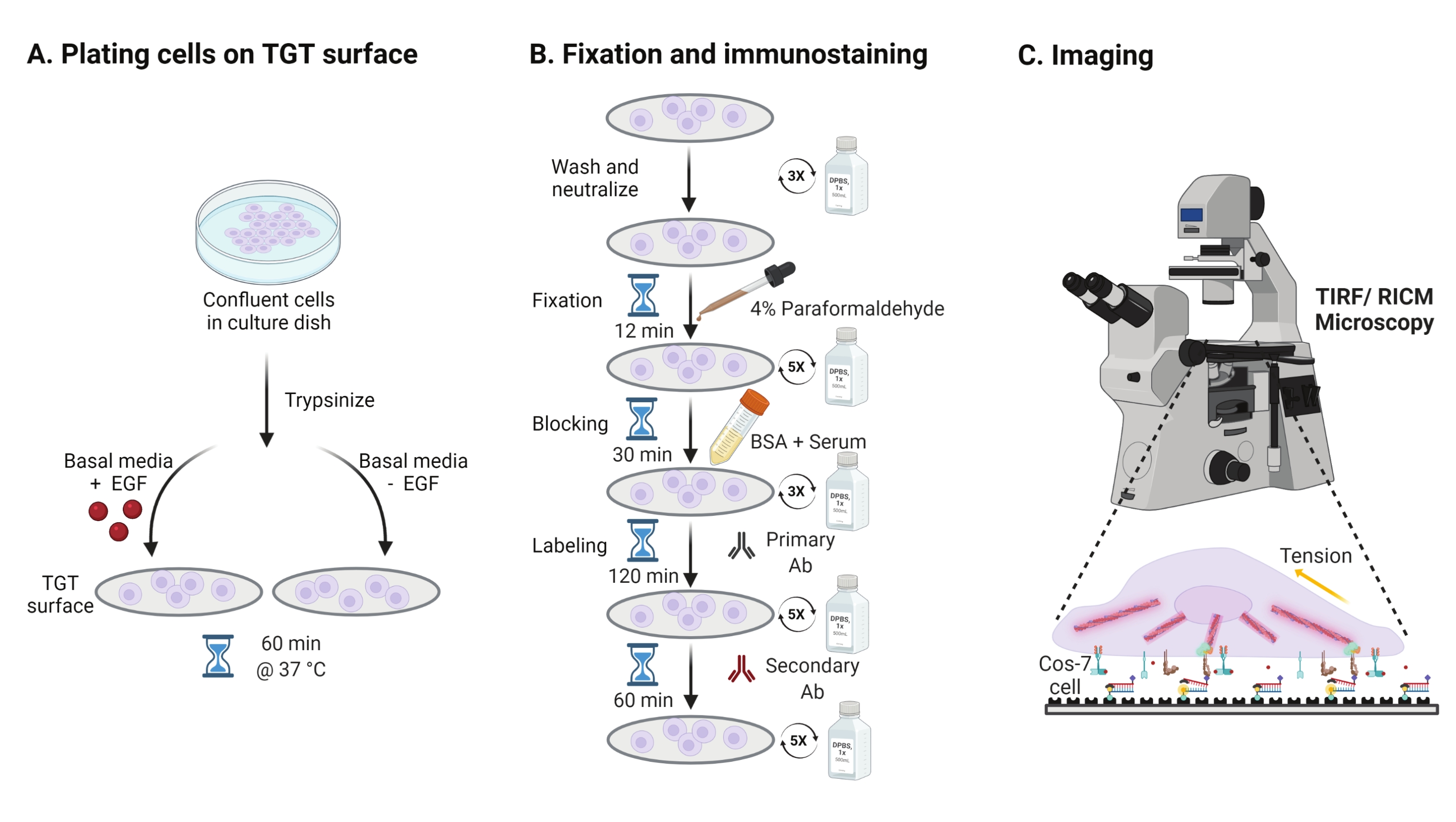
- Per studiare l'effetto della stimolazione del fattore di crescita epidermico (EGF) sulla meccanica Cos-7, l'adesione e la diffusione cellulare, tripsinizzare le cellule Cos-7 con lo 0,05% di tripsina-EDTA per 2 minuti. Neutralizzare la tripsina lavando con HBSS e centrifugando a 800 x g per 5 min. Ripetere ancora una volta il passaggio di neutralizzazione.
- Celle a piastre ad una densità di 4 x 104 celle sulle superfici TGT assemblate nel Modified Eagle Medium (DMEM) di Dulbecco integrato con 50 ng/mL EGF o DMEM senza EGF. Lasciare che le cellule si diffondano per 60 minuti a 37 °C con il 5% di CO2 in un incubatore di colture cellulari (Figura 3A).
NOTA: Le cellule vengono incubate in DMEM senza siero per evitare la stimolazione da fattori di crescita. L'EGF viene diluito in 10 mM di acido acetico per formare una scorta di 1 mg/ml. Viene utilizzato a 50 ng / mL in DMEM per esperimenti di imaging. - Dopo l'incubazione, lavare le cellule tre volte con 1x PBS e fissare con 2 ml di paraformaldeide al 4% per 12 minuti a RT (Figura 3B).
NOTA: Tutte le fasi di incubazione vengono eseguite su uno shaker rotativo a ~ 35 rpm per una diffusione uniforme delle soluzioni. Proteggere le superfici TGT dalla luce coprendole fino a quando non sono pronte per l'imaging. - Aspirare il fissativo e lavare i coverslip cinque volte con 1x PBS a intervalli di 5 minuti a RT. Opzionalmente, incubare i coverslip con 50 mM NH4Cl in 1x PBS per 30 min a 37 °C per estinguere l'autofluorescenza associata alla paraformaldeide e lavare tre volte con 1x PBS a intervalli di 5 minuti (Figura 3B).
- Aggiungere il tampone A (1x PBS, 5% siero di cavallo normale, 5% siero di capra normale, 1% BSA, 0,025% Triton X-100) e incubare per 30 minuti a 37 °C per bloccare e permeabilizzare le cellule. Lavare tre volte con 1x PBS a intervalli di 5 minuti (Figura 3B).
- Posizionare le camere di imaging con coverslips in un contenitore di umidità. Diluire l'anticorpo anti-paxillina primario (marcatore di adesione focale) a 1:250 in tampone bloccante (1x PBS, 5% siero di cavallo normale, 5% siero di capra normale, 1% BSA, 0,005% Triton x-100). Incubare con 200 μL di soluzione anticorpale primaria per coverslip per 2 ore a 37 °C (Figura 3B).
NOTA: non lasciare asciugare le superfici. - Lavare le coperture cinque volte con 1x PBS a intervalli di 5 minuti e restituirle al contenitore dell'umidità. Etichettare le cellule contemporaneamente con una miscela di anticorpo secondario anti-coniglio di capra coniugato con colorante a 1:800 diluizione e falloidina coniugata con colorante (actina) a 1:400 diluizione in 200 μL di tampone bloccante per coverslip. Incubare a 37 °C per 60 minuti (Figura 3B).
- Lavare le superfici cinque volte con 1x PBS a intervalli di 5 minuti e conservare a 4 °C fino a quando non è pronto per l'imaging (Figura 3B).
NOTA: campioni di immagini entro 3 giorni dalla preparazione della superficie per evitare la degradazione del segnale.
4. Acquisizione di immagini
- Utilizzare un obiettivo 60x ad immersione in olio con un'apertura numerica elevata (1,49) su un microscopio invertito con eccitazione TIRF 488, 561 e 647, eccitazione RICM, un sistema di messa a fuoco perfetto e una fotocamera digitale.
- Aggiungere olio all'obiettivo, pulire il fondo della camera del campione e posizionare il campione sul palco. Concentrati su una cellula e attiva la messa a fuoco perfetta.
- Metti il microscopio in modalità di imaging RICM con eccitazione di epifluorescenza e un cubo di filtro GFP con il filtro di emissione rimosso. Allineare il RICM chiudendo e centrando il diaframma dell'apertura dell'epi-illuminazione.
- Metti il microscopio in modalità TIRF con eccitazione laser e un cubo di filtro TIRF quad-pass. Focalizza il laser a 488 nm in un piccolo punto sul soffitto della stanza e aumenta l'angolo di incidenza fino a superare l'angolo critico mentre monitori la fluorescenza sulla telecamera in modalità live. Osservare una forte riduzione della fluorescenza di fondo e un singolo piano di messa a fuoco quando l'angolo critico viene superato.
NOTA: TIRF eccita una regione sottile (~ 100 nm) più vicina all'interfaccia campione-coverslip evidenziando le sonde TGT aperte e le aderenze focali, eliminando al contempo la fluorescenza fuori fuoco dall'interno della cellula. Se TIRF non è disponibile, può essere utilizzata l'epifluorescenza; tuttavia, i rapporti segnale-rumore saranno inferiori. - Identificare le cellule per l'imaging utilizzando la modalità "live" della fotocamera utilizzando RICM.
- Acquisire l'immagine RICM e le immagini TIRF di actina (640 nm ex), tensione dell'integrina (561 nm ex) e paxillina (488 nm ex). Ottenere immagini in sequenza utilizzando un tempo di esposizione di 200 ms.
NOTA: il tempo di esposizione dipende da molti fattori, tra cui l'obiettivo, la potenza del laser, i filtri di emissione e la sensibilità della fotocamera. Il segnale deve essere almeno 2 volte maggiore dello sfondo. Lo sfondo è di circa 1000 UA, quindi il segnale dovrebbe essere almeno 2000-3000 UA. - Ripetere 4,4-4,5 per almeno 30 celle. Cambia coverlips, messa a fuoco e ripeti 4.4-4.5.
5. Analisi dei dati
NOTA: eseguire l'analisi quantitativa delle immagini utilizzando il software Fiji e l'analisi utilizzando il software di statistica.
- Aprire il set di immagini per una cella.
- Create una maschera dell'area della cella (maschera RICM) tracciando il limite della cella nell'immagine RICM utilizzando lo strumento di selezione ImageJ a mano libera. Salvare la regione di interesse (ROI) nel roi manager (Analizza > strumenti > ROI manager) (Figura 4A1,2).
- Scegliete un'area rappresentativa all'esterno della cella nell'immagine di tensione dell'integrina e disegnate un ROI di almeno 200 x 200 pixel. Escludere qualsiasi altra cella o detrito fluorescente dal ROI. Misurare la fluorescenza di fondo nel ROI utilizzando lo strumento di misura (Analizza > Misura) (Figura 4A3).
- Sottrarre la fluorescenza media di fondo ottenuta nel passaggio 5.2 dall'immagine di tensione (Process > Math > Subtract) (Figura 4A4).
- Utilizzare la maschera RICM stabilita nel passaggio 5.2 per definire il segnale di tensione all'interno dell'impronta della cella (ROI manager > Seleziona > Applica maschera > Modifica > Cancella all'esterno) (Figura 4A5).
- Creare una maschera di soglia per l'immagine di tensione utilizzando il metodo di Huang per la soglia automatica (Image > Adjust > Threshold) (Figura 4A6). Assicurarsi che la maschera di soglia rappresenti al meglio l'area per la tensione di integrina generata. Come regola generale, impostare la soglia su 2 volte la fluorescenza media di fondo.
- Create una selezione della maschera di tensione con soglia (Modifica > Selezione > Crea) (Figura 4A7).
- Trasferire la maschera selezionata sull'immagine di tensione generata nel passaggio 5.4 e misurare l'intensità integrata delle sonde aperte (ROI manager > Select (Tension Mask) > Analyze > Measure > RawIntDen) (Figura 4C).
- Misurare l'area delle proprietà morfometriche della cella, la circolarità e le proporzioni dalla maschera RICM (ROI manager > Select (maschera RICM) > Applica maschera > analizza > misura) (Figura 4B).
- Misurare la densità di rottura meccanica, definita come la percentuale dell'impronta della cella con sonde rotte selezionando l'immagine della maschera di tensione e applicando la maschera RICM (ROI manager > Select (RICM Mask) > Analyze > Measure > %Area) (Figura 4C).
- Esporta le misurazioni per ulteriori analisi e visualizzazioni in un software di statistica.
- Ripetere 5.1-5.11 per ogni cella.
Risultati
Le sonde TGT attivate sono state utilizzate per studiare l'effetto del recettore del fattore di crescita epidermico attivato dal ligando (EGFR) sulla meccanica cellulare mediata dall'integrina e sulla formazione di adesione nelle cellule Cos-721. Le sonde presentano il ligando ciclico Arg-Gly-Asp-Phe-Lys (cRGDfK)21,23,25,26, che è selettivo per l'eterodimero integrina αVβ3 con solo una piccola affinità per le integrine α5β1 27,28,29,30. La sonda TGT comprende un DNA duplex funzionalizzato su una superficie di copertura in vetro attraverso il filamento inferiore utilizzando il legame biotina-streptavidina. Il filamento superiore mostra il ligando dell'integrina ed è disponibile per legarsi al recettore dell'integrina affine sulla membrana cellulare (Figura 5A). Il filo inferiore è etichettato con un fluoroforo e il filamento superiore con un quencher, portando a una fluorescenza di fondo minima quando il TGT duplex è intatto. Se un'integrina lega il ligando e applica una forza con una magnitudine maggiore delT tol della sonda, il duplex del DNA si separerà portando alla fluorescenza (Figura 5A). Qualsiasi sonda TGT che non è stata rotta da una forza meccanica rimarrà non fluorescente. Questa fluorescenza di accensione selettiva della forza consente la mappatura sistematica e quantitativa delle forze generate dall'integrina su scala pN a risoluzione limitata dalla diffrazione. Le sonde TGT modulano inoltre la soglia di tensione del substrato.
Qui è mostrato un esempio rappresentativo di una superficie TGT con unT tol di 56 pN. Le cellule Cos-7 sono state placcate su questa superficie TGT con o senza stimolazione EGF per studiare l'impatto dell'attivazione dell'EGFR con la stimolazione del ligando sull'adesione cellulare e sulla meccanica dell'integrina (Figura 5A,B). Le cellule sono state incubate con o senza EGF sulle superfici TGT per 60 minuti, fissate e immunomacchiate per mostrare la distribuzione dell'adesione focale (paxillina) e l'organizzazione del citoscheletro (F-actina) (Figura 5B). Le cellule sono state quindi fotografate utilizzando la microscopia RICM e TIRF. Come chiaramente visibile nell'immagine RICM, la diffusione delle cellule Cos-7 sulla superficie TGT a 56 pN è stata significativamente migliorata con la stimolazione EGF rispetto a quella senza stimolazione. Questo è stato quantificato per 50 cellule in ogni condizione misurando la dimensione della regione di contatto cellula-substrato dall'immagine RICM (Figura 5C). La stimolazione con EGF ha portato a una morfologia più circolare, rappresentativa delle cellule Cos-7 che si diffondono e crescono nel loro ambiente fisiologico naturale (Figura 5D). La fluorescenza delle sonde aperte è anche più alta con la stimolazione EGF come osservato nell'immagine di fluorescenza di tensione. L'intensità integrata delle sonde aperte, che è proporzionale al numero di sonde aperte, era molto più elevata con la stimolazione EGF rispetto all'esterno (Figura 5B,E). Questa è una rappresentazione di tutti gli impegni recettore-ligando in cui le integrine hanno applicato una forza maggiore di Ttol (56 pN).
La colorazione con paxillina ha mostrato che anche la distribuzione, il numero, la maturazione (dimensione) e l'organizzazione delle aderenze focali sono stati influenzati dalla stimolazione EGF. Le aderenze focali nelle cellule stimolate dall'EGF sono apparse più mature e orientate radialmente rispetto a nessun controllo EGF. Anche l'organizzazione citoscheletrica della F-actina è stata migliorata con la stimolazione EGF, come valutato dalla colorazione della falloidina (Figura 5B). Queste valutazioni qualitative sono state effettuate mediante confronto visivo delle immagini di entrambi i gruppi di trattamento. L'analisi quantitativa dell'adesione focale può essere fatta, ma esula dallo scopo di questo protocollo. In questo esperimento, la superficie TGT ha fornito una piattaforma per dettagliare sistematicamente l'effetto dell'attivazione dell'EGFR sulla diffusione cellulare, sulla meccanica dell'integrina e sulla formazione dell'adesione focale.

Figura 1: Schema per il giorno 1 della preparazione della superficie TGT. (A) Pulire i coperchi. (B) Incidere la superficie del coverslip. (C) Neutralizzare la soluzione Piranha. (D) Silanizzare la superficie per creare gruppi amminici reattivi. (E) Equilibrare i coverslips alla fase organica. F) Asciugare i coperchi con un gas inerte. (G) Biotinilare i gruppi amminici superficiali. Fare clic qui per visualizzare una versione più grande di questa figura.
{kind=link}

Figura 2: Schema per il giorno 2 della preparazione della superficie TGT. (A) Pulire e asciugare i coperchi per rimuovere qualsiasi biotina residua dal giorno precedente. (B) Passivare con BSA per evitare il legame non specifico del reagente nelle fasi successive. (C) Funzionalizzare i coverslip con streptavidina. (D) Ibridare le sonde in un termociclatore. (E) Applicare le sonde sintetizzate ai coverslip (F) Assemblare il coverslip nella camera di imaging cellulare. Fare clic qui per visualizzare una versione più grande di questa figura.
{kind=link}

Figura 3: Flusso di lavoro generale che evidenzia le fasi generali dell'intera configurazione sperimentale. (A) Processo per il distacco e la placcatura cellulare sulla superficie TGT in mezzi basali (DMEM) con o senza stimolazione EGF. (B) Diagramma di flusso delle fasi coinvolte nella fissazione e nell'immunocolorazione post-attacco e diffusione sulla superficie del TGT. (C) Dopo la colorazione, le cellule vengono fotografate su un microscopio a fluorescenza invertita con microscopia RICM e TIRF. Fare clic qui per visualizzare una versione più grande di questa figura.
{kind=link}

Figura 4: Esempio di elaborazione dei dati e analisi quantitativa. (A) Ripartizione graduale della pipeline di analisi impiegata nelle Figi (ImageJ) per RICM e quantificazione delle immagini di tensione. (B) Un esempio rappresentativo per i risultati morfometrici delle cellule analizzati utilizzando la pipeline di cui sopra. (C) Esempi rappresentativi per i risultati meccanici delle cellule analizzati utilizzando la pipeline sopra menzionata. Fare clic qui per visualizzare una versione più grande di questa figura.
{kind=link}

Figura 5: Dati di esempio da un esperimento TGT. (A) Diagramma che evidenzia la zona di contatto all'interfaccia membrana cellulare-superficie TGT. Inset proietta integrine che interagiscono con il suo ligando affine cRGDfK con (destra) o senza (sinistra) stimolazione EGF. (B) Immagini RICM e TIRF di cellule Cos-7 diffuse sulla superficie TGT a 56 pN. Le immagini sono ottenute 60 minuti dopo la placcatura con o senza stimolazione EGF. Le singole immagini RICM (come acquisite), tensione dell'integrina (scala di grigi), paxillina (arancione caldo) e F-actina (blu-verde) sono mostrate con sovrapposizioni per entrambe le condizioni di stimolazione. Scala Bar: 10 μm. L'inserto evidenzia un ROI (regione di interesse) ingrandito che descrive in dettaglio la colocalizzazione della tensione di integrina generata nei siti di formazione dell'adesione contrassegnati dalla paxillina e l'organizzazione citoscheletrica subcellulare sottostante contrassegnata dall'actina. Scala Bar: 5 μm. (C-E) Grafici a dispersione per l'area di diffusione (impronta cellulare RICM) (C), circolarità (D) e tensione integrata (E) per cellule Cos-7 con o senza stimolazione EGF. Le barre indicano la media ± s.d. Le differenze tra i gruppi sono state valutate statisticamente con il t-test dello studente; P < 0,0001. n = 50 cellule in tre esperimenti indipendenti. Fare clic qui per visualizzare una versione più grande di questa figura.
{kind=link}

Figura 6: Superfici TGT di esempio con diversi possibili problemi. (A) Immagini di tensione e RICM di una superficie TGT ideale con sonda assemblata spenta prima dell'adesione cellulare. (B) Immagini di tensione e RICM di una superficie TGT in cui la sonda TGT manca del filamento superiore (quencher). L'immagine di tensione mostra una fluorescenza uniforme dal fluoroforo aperto nel filamento inferiore. (C) Immagini di tensione e RICM per cellule diffuse su una superficie TGT ideale. (D) Immagini di tensione e RICM per cellule diffuse su una superficie TGT mal fatta con passivazione limitata o sonda degradata. (E) Le immagini di tensione, RICM e campo luminoso per cellule placcate su una superficie ideale con ligando cRGDfK che indica interazioni cRGDfK-integrina sono vitali per l'attaccamento cellulare e la generazione di tensione. (F) Immagini di tensione, RICM e campo luminoso per cellule placcate su una superficie senza ligando cRGDfK sul TGT. Mentre le cellule sono visibili nell'immagine in campo luminoso, non si osserva alcun attaccamento cellulare o tensione di integrina generata. Scala Bar: 10 μm. Fare clic qui per visualizzare una versione più grande di questa figura.
{kind=link}
Discussione
Con la dettagliata procedura passo-passo sopra descritta, è possibile preparare le superfici TGT per quantificare la morfologia cellulare e la tensione dell'integrina generata dalle cellule aderenti durante l'attaccamento cellulare e la diffusione dopo il trattamento con EGF. La semplice progettazione della sonda e la sintesi e la preparazione della superficie insieme alla semplice configurazione sperimentale hanno fornito una piattaforma stabile per studiare l'interazione di EGFR e integrine. Nel complesso, i risultati convalidano che l'attivazione ligando-dipendente di EGFR migliora la diffusione cellulare, sintonizza le proprietà di forza dei recettori dell'integrina e promuove l'organizzazione e la maturazione dell'adesione focale. I risultati ottenuti utilizzando le sonde TGT supportano l'ipotesi generale che i fattori di crescita, come l'EGFR, agiscano come "meccano-organizzatori", aumentando la quantità e l'organizzazione spaziale della tensione dell'integrina e regolando l'orientamento e la meccanica delle aderenze focali.
Dopo l'applicazione sulla superficie TGT, le cellule atterrano, si attaccano e si diffondono mentre i recettori dell'integrina (αVβ3) rilevano e si legano al ligando cRGDfK. In questo modo le sonde TGT possono essere rotte meccanicamente, generando fluorescenza nel sito di innesto del ligando. La lettura è la "storia della forza" cumulativa della cellula che interagisce con la superficie. Ci sono alcuni problemi comuni con le superfici TGT che possono essere presenti durante questi esperimenti. L'elevata fluorescenza superficiale di fondo (Figura 6A,B), l'aspetto superficiale irregolare, l'incapacità delle cellule di generare il segnale di tensione (Figura 6C,D) e la mancata diffusione delle cellule (Figura 6E,F) possono essere dovute a carenze tecniche con la sonda TGT o la sintesi superficiale. Le soluzioni a questi problemi comuni sono presentate nella Tabella 1.
Il design semplice delle sonde TGT fornisce ai biologi cellulari un potente strumento per studiare specifici risultati di segnalazione dell'integrina del fattore di crescita in isolamento senza interferenze da altri recettori della superficie cellulare fornendo solo ligandi e stimolazioni specifici. Inoltre, le sonde TGT consentono di studiare la soglia di tensione sottolineando i singoli recettori dell'integrina durante l'adesione cellulare alla sensibilità pN. Approcci alternativi non riportano le forze esercitate dai singoli recettori con alta risoluzione spaziale in campioni fissi31. La microscopia con forza di trazione è sensibile solo alle forze nN, un ordine di grandezza superiore alle forze applicate dai singoli recettori dell'integrina15, e le sonde di tensione molecolare misurano le forze pN, ma poiché sono reversibili, non resistono robustamente alla fissazione. Per questi motivi, le sonde TGT sono uno strumento interessante per studiare la meccanica delle interazioni fattore di crescita-integrina.
Ci sono diverse sfumature tecniche associate alle sonde TGT che dovrebbero essere considerate prima di progettare un esperimento. L'immagine di tensione è un'istantanea nel tempo, che rappresenta la storia della forza e non un indicatore degli impegni recettore-ligando in un dato punto temporale. Poiché la generazione del segnale dipende dalla separazione della sonda, la fluorescenza TGT deriva da sonde aperte non sotto tensione attiva dall'impegno recettore-ligando. Ciò significa che la lettura per la tensione di integrina ottenuta sulla superficie TGT è di natura storica e cumulativa che rappresenta dove c'erano forze maggiori di Ttol; le posizioni delle attuali forze recettore-ligando inferioria T tol non sono riportate 19,32. Poiché la rottura del TGT provoca la cessazione dell'impegno recettore-ligando, la diffusione cellulare è dovuta alle interazioni integrina-ligando che sperimentano forze inferioria T tol. L'utente deve quindi fare attenzione quando definisce il tempo post-placcatura per stimare i risultati meccanici associati alle aderenze a base di integrina. Infine, va considerato il significato di Ttol. Le sonde TGT qui impiegate hanno un Ttol di 56 pN, dove Ttol è la forza costante necessaria per rompere il 50% delle sonde quando applicato per 2 s. Quando si considerano sistemi biologici complicati, i TGT probabilmente sperimentano una gradazione di forza eterogenea e diversificata con dipendenze temporali variabili. Se i TGT sono rotti da forze maggiori di Ttol, la fluorescenza sarebbe una sottostima della tensione totale. In alternativa, le forze al di sottodi T tol applicate per durate più lunghe possono rompere un numero simile di sonde come forze di soglia elevate applicate per tempi più brevi. Entrambi questi scenari possono portare alla stessa lettura dell'intensità di fluorescenza, rendendo difficile risolvere l'esatta magnitudine o dinamica della tensione utilizzando le sonde TGT33,34.
Nel complesso, le valutazioni della tensione dell'integrina con la stimolazione del fattore di crescita dovrebbero essere effettuate con attenzione progettando esperimenti con controlli interni, confrontando i profili di diffusione su altre superfici rivestite di matrice, effettuando valutazioni parallele della fluorescenza TGT nelle cellule in presenza o assenza di stimolazione del fattore di crescita e utilizzando TGT con Ttol diversi . I TGT consentono la quantificazione del ruolo della segnalazione del fattore di crescita nella regolazione della meccanica dei recettori dell'integrina, della dinamica di adesione focale e della diffusione cellulare. Questo protocollo può essere utilizzato come modello per molti esperimenti basati su TGT utilizzando sonde con diversi Ttol, diversi ligandi, diversi tipi di cellule o diverse condizioni di stimolazione. Qualsiasi proteina di interesse può essere etichettata dopo la fissazione e qualsiasi tipo di analisi quantitativa dell'immagine può essere implementata. Come tale, presentiamo un modello per numerosi esperimenti TGT.
L'uso di sonde TGT non si limita allo studio delle integrine, ma può essere esteso a una vasta gamma di recettori di membrana cellulare in diversi tipi di cellule modificando il ligando. Le sonde TGT sono state utilizzate per studiare il ruolo delle forze nella regolazione di varie cascate di segnalazione dei recettori, tra cui l'identificazione del ruolo meccanico della meccanica del recettore di Notch nello sviluppo embrionale e nella neurogenesi35, le forze che mediano l'identificazione e l'internalizzazione degli antigeni da parte dei recettori delle cellule B36 e la capacità di correzione meccanica dei recettori di superficie delle cellule T per rilevare cambiamenti nelle forze per aumentare la forza e la specificità del trasferimento del segnale37 . Insieme, questi risultati evidenziano l'immenso potenziale delle sonde TGT in una varietà di contesti sperimentali.
Divulgazioni
Gli autori non dichiarano conflitti di interesse.
Riconoscimenti
Gli autori vorrebbero riconoscere i membri del laboratorio Mattheyses per discussioni e critiche fruttuose. Riconosciamo il finanziamento ad A.L.M. da NSF CAREER 1832100 e NIH R01GM131099.
Materiali
| Name | Company | Catalog Number | Comments |
| (3-Aminopropyl)triethoxysilane | Millipore Sigma | 440140 | Surface Preparation |
| 3-hydroxypicolinic acid (3-HPA) | Millipore Sigma | 56197 | Maldi-TOF-MS matrix |
| Acetic Acid, Glacial | Fisher Scientific | A38S | Diluting EGF |
| Acetonitrile (HPLC) | Fisher Scientific | A998SK | Oligonucleotide Preparation |
| Alexa Fluor 488 Phalloidin | Cell Signaling Technology | 8878S | Immunocytochemistry |
| Ammonium Chloride | Fisher Scientific | A687 | Immunocytochemistry |
| Anti-Paxillin antibody [Y113] | Abcam | ab32084 | Immunocytochemistry |
| BD Syringes only with Luer-Lok | BD bioscience | 309657 | Surface Preparation |
| Bio-Gel P-2 | Bio-Rad | 1504118 | Oligonucleotide Preparation |
| Bovine Serum Albumin (BSA) Protease-free Powder | Fisher Scientific | BP9703100 | Surface Preparation |
| Cos-7 cells | ATCC | CRL-1651 | Cell Culture, Passage numbers 11-20 |
| Coverslip Mini-Rack, for 8 coverslips | Fisher Scientific | C14784 | Surface Preparation |
| c(RGDfK(PEG-PEG)), PEG=8-amino-3,6-dioxaoctanoic acid | Vivitide | PCI-3696-PI | Oligonucleotide Preparation |
| Cy3B NHS ester | GE Healthcare | PA63101 | Oligonucleotide Preparation |
| Dimethylformamide | Millipore Sigma | PHR1553 | Oligonucleotide Preparation |
| DMEM with L-Glutamine, 4.5g/L Glucose and Sodium Pyruvate | Fisher Scientific | MT10013C | Cell Culture |
| Epidermal Growth Factor human EGF | Millipore Sigma | E9644 | Cell Culture |
| Ethanol, 200 proof (100%) | Fisher Scientific | 22032601 | Surface Preparation |
| Falcon Standard Tissue Culture Dishes | Fisher Scientific | 08-772E | Surface Preparation |
| Fetal Bovine Serum | Fisher Scientific | 10-438-026 | Cell Culture |
| Flurobrite DMEM | Fisher Scientific | A1896701 | Cell Culture |
| Goat anti-Rabbit IgG (H+L) Cross-Adsorbed Secondary Antibody, Alexa Fluor 647 | Invitrogen | A-21244 | Immunocytochemistry |
| Goat Serum | Fisher Scientific | 16-210-064 | Immunocytochemistry |
| Hank’s balanced salts (HBSS) | Fisher Scientific | 14-170-161 | Cell Culture |
| Horse Serum | Fisher Scientific | 16050130 | Immunocytochemistry |
| Hydrogen Peroxide | Fisher Scientific | H325-500 | Surface Preparation |
| Nanosep MF centrifugal devices | Pall laboratory | ODM02C35 | Oligonucleotide Preparation |
| NHS-azide | Fisher Scientific | 88902 | Oligonucleotide Preparation |
| Nitrogen Gas Cylinder | Airgas | Surface Preparation | |
| No. 2 round glass coverslips - 25 mm | VWR | 48382-085 | Surface Preparation |
| Parafilm M Laboratory Film | Fisher Scientific | 13-374-10 | Surface Preparation |
| Paraformaldehyde 16% | Fisher Scientific | 50-980-487 | Immunocytochemistry |
| PBS, 1X | Fisher Scientific | 21-030-CV | Surface Preparation/Immunocytochemistry |
| Penicillin-Streptomycin (5,000 U/mL) | Fisher Scientific | 15-070-063 | Cell Culture |
| PYREX Low Form Griffin Beakers | Fisher Scientific | 02-540G | Surface Preparation |
| Sodium Ascorbate | Fisher Scientific | 18-606-310 | Oligonucleotide Preparation |
| Sodium Bicarbonate | Fisher Scientific | S233 | Oligonucleotide Preparation |
| Sodium Chloride | Fisher Scientific | BP358 | Surface Preparation |
| Streptavidin | Fisher Scientific | 434301 | Surface Preparation |
| Sulfo-NHS-LC-Biotin | Fisher Scientific | 21335 | Surface Preparation |
| Sulfuric Acid | Fisher Scientific | A300-500 | Surface Preparation |
| TEAA | Fisher Scientific | NC0322726 | Oligonucleotide Preparation |
| Triethylamine | Millipore Sigma | 471283 | Oligonucleotide Preparation |
| Trifluoroacetic Acid (TFA) | Fisher Scientific | PI28901 | Oligonucleotide Preparation |
| THPTA | Fisher Scientific | NC1296293 | Oligonucleotide Preparation |
| Triton X 100 Detergent Surfact Ams Solution | Fisher Scientific | 85111 | Immunocytochemistry |
| Water, DNA Grade, DNASE, Protease free | Fisher Scientific | BP24701 | Oligonucleotide Preparation |
| Equipment | |||
| Agilent AdvanceBio Oligonucleotide C18 column, 4.6 x 150 mm, 2.7 μm | Agilent | 653950-702 | Oligonucleotide Preparation |
| High-performance liquid chromatography | Agilent | 1100 | Oligonucleotide Preparation |
| Low Speed Orbital Shaker | Fisher Scientific | 10-320-813 | Immunocytochemistry |
| Matrix-assisted laser desorption/ ionization time-of-flight mass spectrometer (MALDI-TOF-MS) | Voyager STR | Oligonucleotide Preparation | |
| Molecular Probes Attofluor Cell Chamber | Fisher Scientific | A7816 | Surface Preparation |
| Nanodrop 2000 UV-Vis Spectrophotometer | Thermo Fisher | Oligonucleotide Preparation | |
| Nikon Eclipse Ti inverted microscope | pe Nikon | Microscopy | |
| Nikon Perfect Focus System | Nikon | Microscopy | |
| NIS Elements software | Nikon | Microscopy | |
| ORCA-Flash4.0 V3 Digital CMOS camera | Hamamatsu | Microscopy | |
| Quad band TIRF 405/488/561/647 cube | CHROMA | Microscopy | |
| RICM Cube | CHROMA | Microscopy | |
| SOLA v-nIR Light Engine | Lumencor | Microscopy | |
| Thermo Forma Steri Cycle 370 CO2 Incubator | Fisher Scientific | Cell Culture | |
| VWR 75D Ultrasonic Cleaner | VWR | 13710 | Surface Preparation |
| Data Analysis | Use | ||
| Fiji (Image J) | https://imagej.net/software/fiji/downloads | Quantitative Analysis | |
| Graph Pad Prism | Graph Pad | Statistical Analysis | |
| Oligo name | 5'modification/ 3' modification | Sequence (5' to 3') | Use |
| Alkyne-21-BHQ2 | 5' Hexynyl/ 3' BHQ_2 | GTGAAATACCGCACAGATGCG | Top strand TGT probe |
| 56 pN TGT | 5' Biosg/TTTTTT/iUniAmM | CGCATCTGTGCGGTATTTCACTTT | Bottom strand TGT probe |
| 12 pN TGT | 5' AmMC6/ 3' BioTEG | CGCATCTGTGCGGTATTTCACTTT | Bottom strand TGT probe |
Riferimenti
- Lim, C. -. G., Jang, J., Kim, C. Cellular machinery for sensing mechanical force. BMB Reports. 51 (12), 623-629 (2018).
- Moraes, C., Sun, Y., Simmons, C. A. (Micro)managing the mechanical microenvironment. Integrative Biology. 3 (10), 959-971 (2011).
- Vogel, V., Sheetz, M. P. Mechanical forces matter in health and disease. From Cancer to Tissue Engineering. Nanotechnology. , 233-303 (2010).
- Wang, J. H. C., Li, B. Mechanics rules cell biology. BMC Sports Science, Medicine and Rehabilitation. 2 (1), 16 (2010).
- Moeendarbary, E., Harris, A. R. Cell mechanics: principles, practices, and prospects. Wiley Interdisciplinary Reviews: Systems Biology and Medicine. 6 (5), 371-388 (2014).
- Streuli, C. H., Akhtar, N. Signal co-operation between integrins and other receptor systems. Biochemical Journal. 418 (3), 491-506 (2009).
- Chiasson-MacKenzie, C., McClatchey, A. I. EGFR-induced cytoskeletal changes drive complex cell behaviors: The tip of the iceberg. Science Signaling. 11 (515), (2018).
- Kechagia, J. Z., Ivaska, J., Roca-Cusachs, P. Integrins as biomechanical sensors of the microenvironment. Nature Reviews Molecular Cell Biology. 20 (8), 457-473 (2019).
- De Luca, A., et al. The role of the EGFR signaling in tumor microenvironment. Journal of Cellular Physiology. 214 (3), 559-567 (2008).
- Javadi, S., Zhiani, M., Mousavi, M. A., Fathi, M. Crosstalk between Epidermal Growth Factor Receptors (EGFR) and integrins in resistance to EGFR tyrosine kinase inhibitors (TKIs) in solid tumors. European Journal of Cell Biology. 99 (4), 151083 (2020).
- Eliceiri, B. P. Integrin and growth factor receptor crosstalk. Circulation Research. 89 (12), 1104-1110 (2001).
- Dan, L., Jian, D., Na, L., Xiaozhong, W. Crosstalk between EGFR and integrin affects invasion and proliferation of gastric cancer cell line, SGC7901. OncoTargets and Therapy. 5, 271-277 (2012).
- Giancotti, F. G., Tarone, G. Positional control of cell fate through joint integrin/receptor protein kinase signaling. Annual Reviews: Cell and Developmental Biology. 19, 173-206 (2003).
- Ricono, J. M., et al. Specific cross-talk between epidermal growth factor receptor and integrin alphavbeta5 promotes carcinoma cell invasion and metastasis. Cancer Research. 69 (4), 1383-1391 (2009).
- Polacheck, W. J., Chen, C. S. Measuring cell-generated forces: a guide to the available tools. Nature Methods. 13 (5), 415-423 (2016).
- Hang, X., et al. Nanosensors for single cell mechanical interrogation. Biosensors and Bioelectronics. 179, 113086 (2021).
- Style, R. W., et al. Traction force microscopy in physics and biology. Soft Matter. 10 (23), 4047-4055 (2014).
- Schoen, I., Hu, W., Klotzsch, E., Vogel, V. Probing cellular traction forces by micropillar arrays: contribution of substrate warping to pillar deflection. Nano Letters. 10 (5), 1823-1830 (2010).
- Ma, V. P. -. Y., Salaita, K. DNA Nanotechnology as an Emerging Tool to Study Mechanotransduction in Living Systems. Small. 15 (26), 1900961 (2019).
- Kim, Y., Kim, K. A., Kim, B. C. Double-stranded DNA force sensors to study the molecular level forces required to activate signaling pathways. Journal of the Korean Physical Society. 78 (5), 386-392 (2021).
- Rao, T. C., et al. EGFR activation attenuates the mechanical threshold for integrin tension and focal adhesion formation. Journal of Cell Sciences. 133 (13), (2020).
- Zhang, Y., Ge, C., Zhu, C., Salaita, K. DNA-based digital tension probes reveal integrin forces during early cell adhesion. Nature Communications. 5, 5167 (2014).
- Liu, Y., et al. DNA-based nanoparticle tension sensors reveal that T-cell receptors transmit defined pN forces to their antigens for enhanced fidelity. Proceedings of the National Academy of Sciences of the United States of America. 113 (20), 5610-5615 (2016).
- Zhang, Y., et al. Platelet integrins exhibit anisotropic mechanosensing and harness piconewton forces to mediate platelet aggregation. Proceedings of the National Academy of Sciences of the United States of America. 115 (2), 325-330 (2018).
- Ma, V. P. -. Y., et al. Mechanically induced catalytic amplification reaction for readout of receptor-mediated cellular forces. Angewandte Chemie International Edition. 55 (18), 5488-5492 (2016).
- Wang, X., Ha, T. Defining single molecular forces required to activate integrin and notch signaling. Science. 340 (6135), 991-994 (2013).
- Chen, Y., Lee, H., Tong, H., Schwartz, M., Zhu, C. Force regulated conformational change of integrin αVβ3. Matrix Biology. 60-61, 70-85 (2017).
- Kantlehner, M., et al. Surface coating with cyclic RGD peptides stimulates osteoblast adhesion and proliferation as well as bone formation. ChemBioChem. 1 (2), 107-114 (2000).
- Kapp, T. G., et al. A comprehensive evaluation of the activity and selectivity profile of ligands for RGD-binding integrins. Scientific Reports. 7, 39805 (2017).
- Kok, R. J., et al. Preparation and functional evaluation of RGD-modified proteins as alpha(v)beta(3) integrin directed therapeutics. Bioconjugate Chemistry. 13 (3), 128-135 (2002).
- Li, I. T. S., Ha, T., Chemla, Y. R. Mapping cell surface adhesion by rotation tracking and adhesion footprinting. Scientific Reports. 7 (1), 44502 (2017).
- Wang, Y., et al. Force-activatable biosensor enables single platelet force mapping directly by fluorescence imaging. Biosensors and Bioelectronics. 100, 192-200 (2018).
- Murad, Y., Li, I. T. S. Quantifying molecular forces with serially connected force sensors. Biophysical Journal. 116 (7), 1282-1291 (2019).
- Yasunaga, A., Murad, Y., Li, I. T. S. Quantifying molecular tension-classifications, interpretations and limitations of force sensors. Physical Biology. 17 (1), 011001 (2019).
- Luca, V. C., et al. Notch-Jagged complex structure implicates a catch bond in tuning ligand sensitivity. Science. 355 (6331), 1320-1324 (2017).
- Spillane, K. M., Tolar, P. B cell antigen extraction is regulated by physical properties of antigen-presenting cells. The Journal of Cell Biology. 216 (1), 217-230 (2017).
- Brockman, J. M., Salaita, K. Mechanical proofreading: a general mechanism to enhance the fidelity of information transfer between cells. Frontiers in Physics. 7, 14 (2019).
Ristampe e Autorizzazioni
Richiedi autorizzazione per utilizzare il testo o le figure di questo articolo JoVE
Richiedi AutorizzazioneThis article has been published
Video Coming Soon
Personale delle biblioteche
Copyright © 2025 MyJoVE Corporation. Tutti i diritti riservati