
Method Article
魚にIonocytesを研究するために完全な二国間ギル脱神経と相まって時差染色技術
要約
This manuscript describes a protocol to track the re-distribution of branchial ionocytes and their innervation using a time differential staining technique coupled with full bilateral gill denervation.
要約
鰓ionocytes(IC)は、魚のイオン調節のための機能ユニットである。イオンチャネル、ポンプおよび交換の様々なビアおよびCa 2+ -成人では、それらは、フィラメント状などのNa +、Cl等の鰓どこ輸送イオンの層状上皮に見出される。硬骨魚鰓は外因フェイシャル(VI)、舌咽(IX)と迷走(X)神経によって支配されている。 IXおよびX神経はまた鰓のIC神経支配の外因性供給源である。ここでは、神経支配、増殖、およびICの分布を研究するために使用される2つの技術が説明されています。時間差染色技術と完全な二国間鰓除神経技法。簡単に言えば、金魚が不可欠ミトコンドリア固有の色素にさらされている( 例えば、のMitoTrackerレッド)ラベル(赤色蛍光)既存のIC。魚のどちらか3回復させた - 5日またはすぐにフル二国間鰓除神経を受けた。回復の5日間、G - 3の後病気を採取し、免疫組織化学のために固定されている。組織はその後、α-5一次抗体で染色された(標的細胞を含む+ / K + ATPアーゼのNa)すべての(新規および既存の)ICのグリーンラベルを付けた二次抗体と組み合わせた。共焦点イメージングを用いて、それが既存のICは(生ミトコンドリア特異的染料及びα-5の両方で標識された)が黄色に表示され、新たなICは(α-5のみで標識された)緑色に表示されていることが実証された。タンデムで使用される両方の技術は、魚が環境問題にさらされているとき、鰓フィラメント上のICの神経支配、増殖および分布を研究するために適用することができます。
概要
ICは、魚にイオン調節する機能部であり、鰓フィラメントおよびラメラ4,6-8,10の上皮表面に見られる。サブタイプの様々なユニークな特徴を有することが記載されているが、ICの多くは(したがって、それらはまた、ミトコンドリアが豊富な細胞として知られている)ミトコンドリアの高密度および/ または酵素の存在量のNa + / K +を特徴とするATPアーゼ(NKA)。典型的には、これらのICハウスイオン調節に関与する他のポンプ、イオンチャネルおよび交換の様々な( 例えば Na + / H +交換体のNa + /のCl -共輸送体、H +ポンプ)2,10,11。代償機構としてICの再分配と増殖は、特にイオン性ストレス( 例えば、イオンの乏しい水への曝露)4,8,9時のイオン恒常性を維持するための中心である。
この研究は、テクニックを染色する時間差を記述魚の鰓で新たに増殖しionocytes(IC)を識別するためのQUE 1。この技術は、鰓弓の完全な両側性除神経に結合される。金魚( キンギョ )、これらの研究に使用される種は、それらが構造的に彼らの鰓2を改造する顕著な能力を持っているので、鰓上皮細胞の増殖を研究するために適している。冷水(<15℃)または低酸素症、それぞれ3に順応している-ギルリモデリングは、(30℃、通常は15で維持)魚がラメラ細胞塊の成長または後退(ILCM)を指します。金魚に時間微分染色技術を用いた以前の研究は、4,5改造鰓の文脈における鰓上のICの再分配、神経支配および増殖に焦点を当てている。加藤と金子1はメダカで変換と鰓ICの交換を研究するために、この新規な技術を開発した(Fundulusのheteroclitus)transfe淡水(FW)に海水(SW)からRRED。本研究では、焦点は、25°Cに順化キンギョICの増殖および神経支配である。
それは鰓リモデリングの文脈では、金魚は、低酸素曝露およびその後の正常酸素の回復中にICの一定の数を維持することが示された時間差染色技術を使用して、しかし、神経支配された細胞の割合は、正常酸素回復期間5全体に減少した。これは、魚のイオン取り込み機構は神経制御12下にあることを70年以上前に提案された。硬骨魚ギルは「鰓神経」13,14と呼ばフェイシャル(VII)、舌咽(IX)と迷走(X)神経によって支配されている。ゼブラフィッシュ( ゼブラフィッシュ )鰓神経支配にJonzおよびナース(2003)による研究では、神経支配の起源は外因性(神経線維の細胞体は、鰓に外因性である)、ならびに内因性の(細胞体であることを示した神経線維)が鰓に固有のものである。同じ著者はまた鰓ICが外因7を神経支配していることを実証した。
本研究では、完全な両側性鰓除神経に結合された時間差染色技術は、金魚で外因性神経支配を欠いているICの増殖を調べた。繊細な外科手術を簡素化し、ionocytesは、標準の免疫組織化学的技術を用いて同定される - フルバイラテラル鰓除神経は(200gの30)は、それらの比較的大きなサイズがあるため、これらの二つのアプローチは、金魚に実現可能であるIX及びXの脳神経を切断することをいう。発達研究ハイブリドーマバンク、大学、本研究では、ICは、極めて重要なミトコンドリア特有の色素( 例えば、のMitoTracker赤)又はNa + / K + -ATPアーゼ(α-5のαサブユニットに対する一次抗体を用いて可視化したアイオワ、アイオワシティアイオワ州)の。このプロトコルは、単純なメトを提供しています魚の鰓上のICの再分配および増殖を可視化し、分析するのD。
プロトコル
両方のプロトコルは動物ケアのカナダ人評議会(CCAC)のガイドラインに準拠し、大学オタワの動物管理委員会(プロトコルBL-226)の承認を得て実施した。
1.時差染色技術:ミトコンドリアが豊富な染料バース
- ジメチルスルホキシド(100%DMSO)の94.0μlの50μgのを溶解して1 mMののMitoTrackerレッド原液を準備します。使用しないときは-20℃で暗所に原液を保管してください。凍結/解凍サイクルを避けてください。
- 600ミリリットルの最大容量で - (6箱3)暗黒の箱を準備します。システムの水400ミリリットル(水通常に保持されている魚)でボックスを記入し、O 2の供給源を提供するために、各ボックス内の空気の石を配置。金魚を入手(30〜40グラム)を400 mlの水と空気の石のボックスに配置します。 30分後、0.1μMおよび0.01%DMSOの最終濃度を得るために実行可能なミトコンドリアが豊富な染料を追加します。 4時間魚を浸すR。
- これらのコントロールの魚( すなわち、無脱神経)である場合、ボックスへの水の流れをオン染料洗い流しプロトコルに割り当てられた時間の期間、魚を回復することを可能にする。 5日 - 魚は典型的には3のために回収される。
- 時差染色技術:免疫組織化学の回復期間の後、 第3節に進んでください。これらの魚は、除神経される場合は、セクション2に進んでください: 完全な二国間除神経手順 。
2.完全な二国間除神経手順
- 学生Vannasスプリングはさみ(曲線)、標準パターン鉗子曲面の標準パターン鉗子ストレート、1組の1組、第5鉗子、組織リトラクターの1ペア、小さなコットンボール(1の2組の1組を取得 - 直径2 mm)であり、綿棒(Qヒントまたは同等)。
- 麻酔薬水浴を準備します。まず、最終容量100mlにベンゾカイン10gを溶解させるストック溶液を調製し、99%のエタノール。麻酔水浴を調製するために、必要な温度で曝気システム水30リットル中にストックベンゾカイン溶液15mlに溶解する。水槽に空気ポンプまたは中央エアラインに接続されたエアストーンを配置することによって、水を通気する。
- 麻酔薬水浴( 図1A)に魚を置きます。
- 呼吸が停止した後、手術台上の魚を配置し、 図1Bに示すように、挿管。曝気麻酔液と鰓を灌漑するために、その口腔内チューブを挿入することによってこれを行います。えらの灌漑は、魚が除神経処置中にO 2及び麻酔薬の十分な量が供給されることを保証する。頭が少し下向きに傾斜されるように魚を置きます。これは、第四鰓弓の背後にあるエリアへのより良いアクセスを可能にします。
- 優しくストレート標準パターン鉗子で蓋を持ち上げるそして蓋と頭部の内側との間の組織のリトラクターを配置。慎重に頭から蓋を維持し、露出した鰓を維持するために、組織のリトラクターを開きます。
- 麻酔液はこの手順の間鰓を灌漑していることを確認してください。リトラクターは、すべての4つの鰓のアーチへのアクセスを提供する頭の横のハンドルを置きます。
- 頭部への第四の鰓弓を装着する靭帯の張力を作成するためにそれらを開く優しく四鰓弓と後頭部の間に湾曲した標準パターン鉗子を置き、。主鰓蓋空洞に鰓弓の背側の端を接続する上皮を貫通することにより、 - (3 mm 2)と第5ピンセットで小さな開口部を作成します。主要な血管を損傷する危険性があるので、あまりにも深くに行くしないように注意してください。
- 第5鉗子で開催され、小さなコットンボールでゆっくりと慎重にIX(舌咽)とX(迷走)神経を露出するように切開を展開します。フリー再び血管を傷つけないように注意しながら、5号鉗子を使用して、すべての結合組織から神経。
注:金魚の鰓IXおよびX神経は第四の鰓弓の後ろに深い休息と鰓のアーチへの供給の主要な血管に近接している。 - 神経一度湾曲した標準形状鉗子とのオープン切開を保持した状態で湾曲した春のはさみは慎重に神経を切断するために使用することを確認されている。神経を切断した後、緩やかに湾曲鉗子を撤回。 48時間 - 上皮は非常に薄く、切開は、通常24内にそれ自身で閉じているため縫合糸で切開を閉じる必要はない。組織リトラクターを削除します。
- 頭の反対側に同じ手順を繰り返します。
- 麻酔から魚を回収するために新鮮な曝気水に麻酔から鰓の灌漑を切り替えます。一度主鰓蓋の動きは、少なくとも24時間、静養するために実験的なタンクに魚を移動し再開した。
- は魚の別のセットの「偽」の手順を実行します。 「偽」の手順では、神経を切断することなく、第四の鰓弓の背後にある上皮を貫通することを含む。
3.時差染色技術:免疫組織化学
- まず、1×リン酸緩衝生理食塩水(; PBS 96mlの中に4グラムPFA PBS)中の4%パラホルムアルデヒド(PFA)を調製する。 PFAは室温でPBS中に簡単に溶解しない。 PFAを溶解し、水浴中で熱溶液。ヒュームフードでこれを実行します。 PFAは、溶液中になったら、使用前に冷ます。 2週間まで4℃で保管してください。
- 8シンチレーションバイアル(鰓弓あたり1バイアル)の合計のためのシンチレーションバイアルに4%PFAの4ミリリットル - 魚を安楽死と鰓組織を抽出する前に、3を配置。また、小さな量るボートに乗り、1×PBSでそれを埋める。これは、切除された後に組織を洗浄するために使用される。氷の上にすべてのソリューションを保管してください。
- 実行可能なミトコンドリアが豊富な色素露光の後実験は、ベンゾカインの過剰摂取で水浴に置くことで金魚を安楽死させる、終了した。
- 鰓鰓バスケットのそれぞれの端を切断する頭部の片側の蓋や湾曲したハサミを持ち上げるために鈍鉗子を使用してください。慎重に鈍ピンセットを用いて耙によって鰓をピックアップし、主鰓蓋空洞からそれらを持ち上げる。すぐに過剰ベンゾカインと血液を除去するためにPBS 1Xの氷冷で鰓を洗う。
- 4%PFAで満たされた別々のバイアルで切除した鰓(各鰓弓用バイアル)を配置し、4℃でO / Nを修正。
- 固定後、1×PBSで過剰PFAを洗浄し、4℃でRTまたはO / Nで6時間、シェーカー上、1%のTriton-Xの1.5ミリリットルで満たされた2ミリリットルの弾丸チューブに組織を置く。このステップでは、組織を透過性に。全体ギルは2ミリリットルチューブに置くには大きすぎる場合には、フィラメントに損傷を与えないように注意しながらチューブを収まるほど小さいのセクションに組織を切断。
- MIによる一次抗体希釈液を調製するXINGの1X PBS 992μLに各一次抗体(8μLの合計)のストック溶液4μlの。 NKAは(NKAが豊富な細胞を標識する)ことを確認し、ZN-12(ラベルニューロン)一次抗体が同じホストに提起されている。研究者は、彼らが別の宿主種で育った一次および二次抗体を使用する必要がありますいると同時に、特定のICのサブタイプを識別することにした場合、これは重要です。
- トリトンX溶液を除去し、組織を洗浄せずに1追加:NKAが豊富な細胞を検出するためにNKAモノクローナル抗体の250希釈(1×PBS中に希釈する)(α-5)とゼブラフィッシュ特定の神経抗体(ZN-12)神経線維(一次抗体)を検出し、4℃でRTまたはO / Nで6時間、シェーカー上でインキュベートする。
- 1X PBSを用いて3分間ずつ、一次抗体を3回洗浄する。これを行うには、ピペットを用いて、それを吸引することにより、チューブから一次抗体溶液を除去。
- 二次抗体を準備1:1×PBSの995μLに株式の二次抗体の5μLを混合することによって200希釈。二次抗体(アレクサフルオロ488)を適用し、シェーカー上、4℃でRTまたはO / Nで6時間インキュベート。
注:のMitoTrackerレッドは、〜594 nmの波長で励起され、赤の蛍光を発する。 〜488nmの波長で励起され、緑色の蛍光を発する二次抗体は、NKAにラベルを付け、ZN-12一次抗体を使用する必要があります。 - (ステップ3.7で説明したように)5分ごとに組織を3回洗浄することにより、過剰な二次抗体を除去します。
4.イメージング
- 洗浄後、細胞全体および神経線維の共焦点イメージングをマウントするための凹状のスライド上の組織をマウントします。組織をマウントするには、第1平坦顕微鏡スライド上に1×PBSの滴(200μL)に入れてください。これは、組織が乾燥することはありません。
- 湾曲したマイクロハサミで鰓hemibranchs区切ります。 1×PBSのドロップと凹スライドに取り付けメディアのドロップを置きます。
- 上に向けフィラメントの先端に凹面スライドに分けhemibranchsを置き、カバーガラスでカバーしています。動き回ると、組織を変位からカバースリップを防止するためにマニキュアを有するカバースリップの端を軽く。イメージングの前に15分 - 組織が10のために凹面スライドの底に沈殿することを許可する。
- 各鰓弓の場合は、鰓弓あたり6枚の画像を生成する、画像化のためにランダムに6鰓フィラメントを選択します。 3μmの光学スライス - 1を取ることによって、イメージに組織を従来の共焦点顕微鏡を使用してください。
注:すべての既存の細胞がNKAのためのMitoTracker赤とポジティブで標識されるだけ黄色に表示されます。赤い表示された細胞は、唯一のNKAが含まれていない既存のICです。でも新たに増殖した細胞のみがNKAのためにポジティブになり、唯一の緑が表示されます。神経繊維も緑色に表示されます。
Ionocyte定量5.画像解析
- EACのために撮影された時間鰓フィラメントは、Zスタックのセクションをスクロールし、存在し、それらが新たに分化しているか否かを、既存の、および/または神経支配ラメラ及びフィラメント状ICの数をカウントすることによりICと関連する神経支配を定量。
- フィラメントの(mm 2)とフィラメント当たりや面積あたりのICを定量化する。 ICが定量化されたフィラメントの領域を網羅するフィラメントのラメラを概説するために画像を取得するために使用されるソフトウェアに関連した描画ツールを使用することによってこれを行います。ほとんどの共焦点イメージングソフトウェア·プログラムは、画像上で概説した領域の面積を計算するためのオプションを持っている。あなたが領域を取得するためにこれを行うことを可能にするイメージングソフトウェアのオプションを使用します。
- 単位面積( 例えば、1mm 2あたり)あたりのICの測定値を得るために、ソフトウェアによって計算領域によってカウントICを分割する。
結果
図1(図1A)を設定手術テーブル、手術中の魚の配置( 図1B)と時間差染色技術( 図1C)のための3つの最も重要なステップを示している。ステップ1では、魚が暗所で25℃で十分に通気され、水浴中で30分間維持される。 30分の期間の間、研究者は、ステップ2( 図1C)の間に水に添加し、DMSO中のミトコンドリアが豊富な染料のアリコートを調製することができる。ステップ2での潜伏期間は、ミトコンドリアが豊富な細胞( すなわち、ICS)への水からミトコンドリアが豊富な染料の取り込みが可能になります。魚はその後のどちらかが完全な二国間の除神経手続きや魚に麻酔されている偽の手順および操作operculaを受けることができるが、神経が無傷のままである。ステップ3で設定したEIを持って魚流水で提供回収室を表し、THERは完全な二国間の除神経または「偽」の手順を経て。
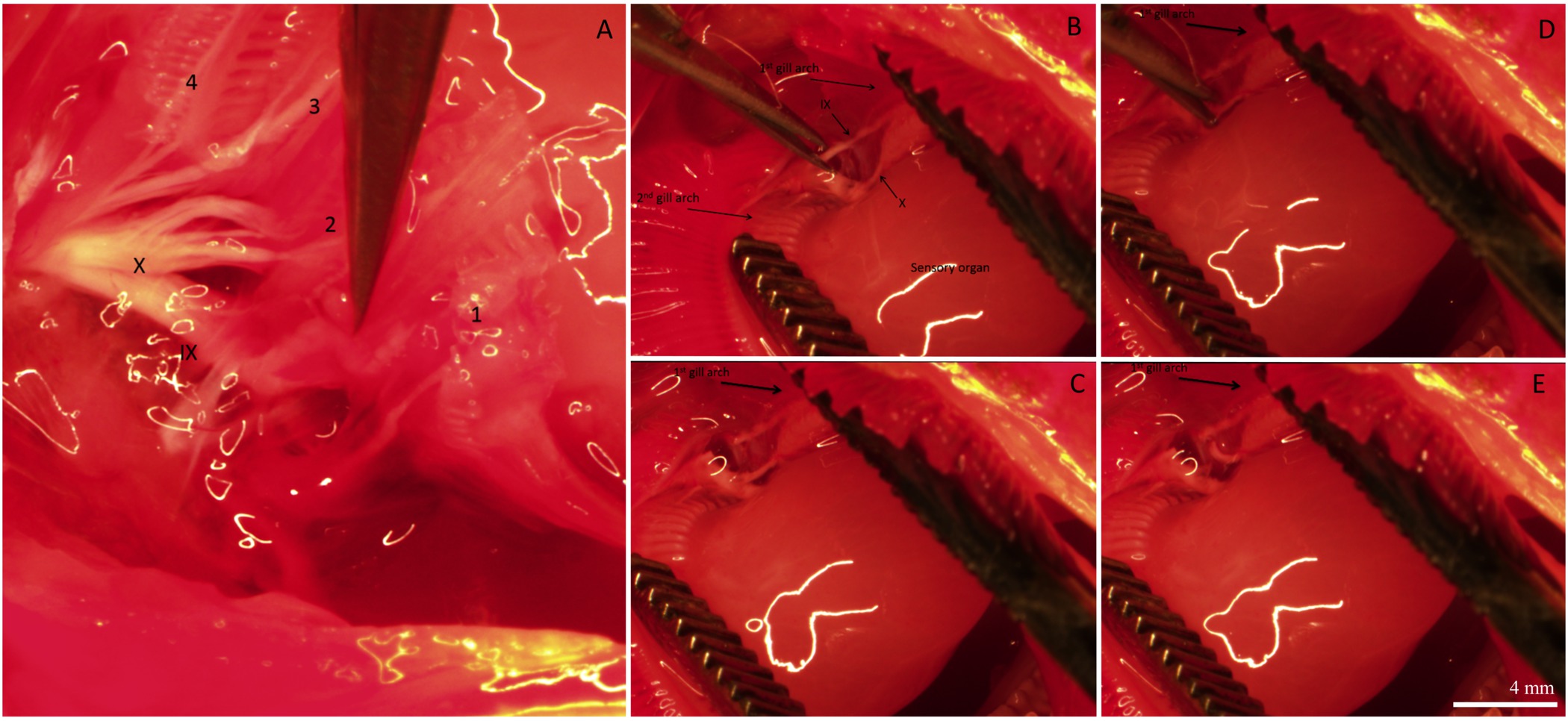
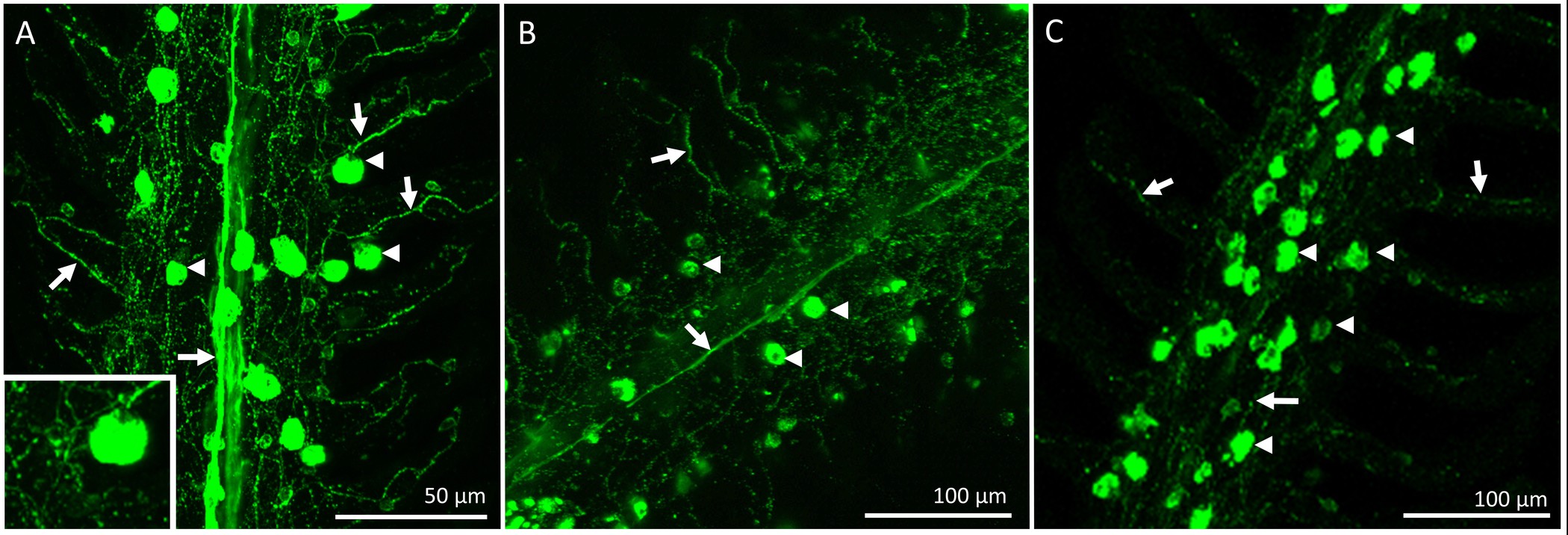
回復期間の後に、魚を安楽死させ、鰓、免疫組織化学のために摘出し、固定した。 「偽」の手順を受けた魚の鰓フィラメント上のICの全体的な分布と神経支配は、 図2に示されている。ICはフィラメント状上皮上に存在するだけでなく、ラメラ間地域の基部にある。 図2Aショーミトコンドリアが豊富な色素で標識された既存のICは(除神経/シャム手順が行われた前に、 すなわち、これらのICは存在していた)。 図2Bは、神経繊維がフィラメント状の(NKAの免疫反応性によって識別される)新たに既存のと形成されたICを支配示しており、ラメラ上皮。最後に、二つの画像のマージ( 図2C)は、明らかに既存のIC(黄色に見える)と新しいICを(緑表示されます)を明らかにする。図2Dは 、 図2A-Cに示されたフィラメント用のIC定量化の代表的なグラフである。この特定のフィラメントに、より大きい既存のICよりも1mm 2あたり新たに増殖したICの個数(N = 1)があるように思われる。外因性鰓神経支配のソースを削除するには、IXおよびX脳神経(外因性神経支配)が切断された。 図3(a)は、鰓フィラメントの後鰓蓋キャビティの背側領域を示していると神経をカバー上皮はIXおよびXを露出させ除去してすべての4つのアーチ( 図3A)を支配するために二つの主要な神経幹からスパン(ローマ数字で示す)脳神経。鰓アーチは第一鰓弓の選択的な脱神経を説明する。 図3B-Eを通じて1の番号が付けられています。最初の鰓弓が残りoを神経支配に影響を与えることなく除去することができるIX及びX脳神経( 図3B)の両方によって支配される鰓のアーチ( 図3C-E)、F。鰓フィラメントへの外因性の神経支配の段階的な損失( 図4A-C)で完全な二国間の除神経の結果。対照魚はフィラメント( 図4A)の長さに及ぶ明らか神経束を示す。完全な二国間の除神経は回復の2日間( 図4B)の後に外因性神経支配のいくつかの損失となりました。外因性神経支配のさらなる消失が除神経( 図4C)からの回復の5日後に認められた。回復の5日後のICへの残りの神経支配は、おそらく鰓フィラメント(; 図4C固有神経支配)内の細胞体と神経に由来するものであった。

図1。実験はで使用されるステップの除神経手順およびシーケンス用に設定時差染色技術。時間微分染色技術で使用される(A)麻酔し、挿管魚の配置の手術テーブル麻酔薬と回復のタンクを示す除神経手続きのために設定。(B)例。(C)重要なステップ。ステップ1において、魚は30分間、温度制御され、静的な曝気水浴中に配置されている。ミトコンドリアが豊富な色素は0.1μMの最終濃度になるように、ステップ2で追加され、魚が4時間の最小のために溶液中で入浴することが許可されている。水の流れは魚が鰓の完全な二国間の除神経または偽手術のいずれかを受け、その時点で、ステップ3で再開される。 この図の拡大版をご覧になるにはこちらをクリックしてください。
{kind=link}
 />
/>
2日間フル二国間の除神経後に(25℃に順応)金魚における時間微分染色技術を表す図2.ライト顕微鏡写真。 (A)既存ionocytes(集積;矢印)の分布単一鰓フィラメントには、ミトコンドリアが豊富な色素染色によって明らかにされる(B)は、ICの分布(新規および既存の)および鰓神経(破線矢印)。それぞれ、α-5およびZn-12抗体で染色することにより明らかにされている。新たに増殖したICから(緑色に、矢印で示されている);矢印はICを示す(C)(A)のオーバーラップ及び(B)は、既存のICを(矢印で示す黄色に見える)を区別する。 (C)の中に挿入する神経支配既存ionocyteの倍率である。(D)パネル(AC)に示すフィラメント用ICの定量化の代表的なグラフ。パネルではN = 1のスケールバー(C)は50μmであると、すべてのパネルに適用されます。/ftp_upload/52548/52548fig2highres.jpg「ターゲット= "_空白">この図の拡大版をご覧になるにはこちらをクリックしてください。

図鰓神経支配の様々な段階を描い3.代表的な画像。すべての4鰓弓IXおよびX脳神経の神経支配を示す口腔の(A)背ビュー。ギルのアーチは、1-4の番号が付いています。神経を覆う鰓フィラメントおよび組織を第 1 の (B)。優れた神経分布を可視化するために除去し、 第 2 鰓弓を、感覚器官を明らかにするために分離され、IXおよびX脳神経の分岐が第 1鰓を支配しているアーチ。IXおよびX脳神経の枝を切断することによって1 番目の鰓弓の選択的な脱神経を示す画像の(CE)シーケンス。回路図再ために。硬骨魚の鰓神経支配のプレゼンテーション、Milsom ら図1を参照する26の画像の白い線は、顕微鏡のライトからの水の反射である;彼らは、あらゆる形態学的構造を定義していない。パネル内のスケールバー(E)は 4ミリメートルで、すべてのパネルに適用されます。 この図の拡大版をご覧になるにはこちらをクリックしてください。
{kind=link}

25℃Ionocytes(矢頭で示される) に順化金魚の1 番目の鰓弓の単一のフィラメントにionocytesの分布と神経支配を描いた図4.ライト顕微鏡写真を示した(α-5抗体と神経で染色したByの矢印)をznが-12抗体で染色した。おそらく広範囲ラメラ分岐でIX及びX神経(外因性神経支配)から生じる中枢神経束を示す制御魚の(A)ギルフィラメント。示さionocytesの一部は、(挿入)。(B)A鰓フィラメント2日間フル二国間の除神経後に神経支配されている。ラメラ神経支配は5日、外因性神経支配が大きく存在しなかったことを実証する完全二国間の除神経後に。(C)A鰓フィラメントほとんど無傷に見えた中枢神経束の減少があった。定性分析は、完全な二国間の除神経が鰓フィラメント内の細胞体(内因性神経支配)フィラメントを通って、ラメラに神経のネットワークを作成すると、神経を維持しつつ、外因性の鰓神経支配の劣化の原因となることを示唆している。 してくださいこの図の拡大版を表示するには、こちらをクリックしてください。
{kind=link}
ディスカッション
時間微分染色技術は、イオンの取り込みの動的な調節を理解し、鰓上皮にICの一時的な再配分を検討する便利なツールとなります。簡単な手順であるが、時間微分染色技術の成功に不可欠である重要なポイントがいくつかあります。金魚は、プロトコルに割り当てられた時間のためのミトコンドリアが豊富な染料にさらされなければならない。短い曝露はミトコンドリアが豊富な細胞( すなわち、ionocytes)により色素の貧しい取り込みになります。固定時には、鰓組織を迅速に切除しなければならず、フォトブリーチングを回避するために、暗所に保存。組織は、固定の2週間以内に画像化のために処理されなければならない。二国間の神経切片手続き中の魚がよく、麻酔をかけていることを確認してください。神経や血管を明瞭に識別される。それは回収タンクに移される前に、魚がいっぱい鰓蓋機能を再開します。
"> FW環境は、受動的なイオン損失と浸透水のゲイン14のバランスを取るという二重の課題に魚を提示します。どこに彼らができる受動的なイオン損失のバランスがフィラメント状に局在しているICの間で塩の積極的な取り込みとラメラの上皮を介して起こる外部環境2,4,8,9,16と直接接触する。しかし、鰓上のICの位置は静的ではありません。直面したときに、過去30年間の研究の数は、そのいくつかのFWの魚種を示しているイオン性および/ または温度の課題に、ラメラ1,2,4,5,17-21のより遠位の領域にラメラのフィラメントまたは塩基から鰓ICを再分配する。このような再分配は、ラメラの厚さを増大させることができる鰓上皮22間でガス移動(O 2、CO 2)を危険にさらすことができます。研究者は移転emergencを追跡するために、この原稿に記述された時間微分染色技術を使用していたこれらの異なる実験条件( 図1C)1,4,5下鰓上皮上の新しいICの電子。鰓、おそらく鰓ICは、IXおよびX脳神経7,23-25 によって支配されている。これらの神経は、それぞれ、および鰓から遠心性と求心性入力の両方を運ぶ。これらは、4 番目の鰓弓の背後に口腔の背側に位置している。神経のアクセシビリティと二国間の除神経手続きを行うことができる容易さは、種特異的である。神経は組織の薄い層の下に4 番目の鰓弓の後ろに単一の平面内に位置するマスでは、例えば、頭の尖ったと平らに解剖ができます。これは神経が見えると除神経の手順を実行するには、研究者へ簡単にアクセスできるようになります。これとは対照的に、金魚は短い鼻と丸い頭を持っている。金魚のIXおよびX脳神経は、背側のSiに深くうそ異なる平面を占める第 4鰓弓の後に空洞のデ。この配向は、神経へのアクセスのしやすさを制限し、適切な神経を特定し、切断するために、より慎重なアプローチが必要です。除神経手順の目的は、それぞれ、および鰓から感覚求心性と遠心性入力を削除することです。鰓弓の除神経はまた、放射性同位体( 例えば、22のNa)を用いたイオンフラックス実験と結合することができる。これらの技術は、鰓上皮を横切るイオンの移動に神経入力の寄与を研究するために連携して使用することができる。除神経処置に対する別の制限は、神経束を切断するときにこのようにしてそれが神経支配の両方のタイプが削除されることが可能である、感覚および運動ニューロンを区別できないことである。すべての運動ニューロンを切断すると、魚の鰓と主鰓蓋運動に影響を与える可能性が。鰓除神経実験を行う際にこのように、それはまた後に換気を監視することが重要である魚は、ガスとイオン交換の両方のために十分な鰓動きがあることを確認するための手順から回復した。
暗サイクルおよび摂食市販食品ペレット:この原稿を使用し成体動物に記述プロトコルは、12:12光に保つ。これらの方法は、いくつかの方法で変更することができる。まず、ミトコンドリアに富む色素曝露後の回復期間は、研究者のプロトコル( 例えば、1,3,5、または14日間)の要件に合わせることができる。ラボでのミトコンドリアが豊富な染料曝露後の回復の最長期間は14日間4,5なっている。回復の14日後のミトコンドリアが豊富な色素の蛍光強度の有意な減少がありませんでした。第二に、α-5一次抗体を使用することが唯一のNKAの豊富な細胞を同定し、鰓ICの異なるサブタイプを区別しないに制限されています。幸いなことに、金魚にはICの大半は両方mitochondrであることが確立されたイオンが豊富な(のMitoTrackerラベル)とすべての魚種4,27の場合とできない場合があり細胞(α-5とラベル)NKAに富む。今後の実験はチャンネル、ポンプと交換、各種( 例えば、NHE、H +ポンプ )に対して特異的に向けられた抗体を使用して、特定のICサブタイプの一時的な再分配を以下に集中することができます。これまでの研究では、ionocytesの大半はミトコンドリアが豊富な色素展示NKA免疫反応4で染色したことがわかった。 ICの神経支配は、ゼブラフィッシュ由来のニューロン特異的抗原(ZN-12)に対する一次抗体を用いて検出することができる。本研究では、α-5およびZn-12を一次抗体の両方に同じ二次抗体(アレクサフルオロ488)を用いて検出した。この制限は、両方の一次抗体がマウス宿主に上昇さ及びNKA豊富細胞およびニューロンは、それらが同じ色を発するにもかかわらず、形態学的に区別することができるという事実によって克服されるためである。連続染色異なる二次抗体(アレクサフルオロ488と、例えば、最初のα-5、次いでアレクサフルオロ594-12 ZN)も、同じ色の蛍光の両方のマーカーを有するという問題を解消するために使用することができる。最後に、完全な二国間の神経切片プロトコルは、特定の鰓のアーチに神経を標的とするように変更することができます。例えば、選択的神経切片を穏やか鰓バスケット( 図2B-E)の背側末端の最初の鰓弓に通じる神経を露出させる第一および第二鰓アーチを分離することにより、第1鰓弓で行うことができる。時間微分技術はまた、開発および鰓皮膚からイオン輸送遷移として幼生ゼブラフィッシュ魚ionocytesの分布を研究するために適用することができる。
開示事項
The authors declare that they have no competing financial interests.
謝辞
We would like to thank William Fletcher for animal care at the University of Ottawa. The authors would also like to thank Dr. William Milsom for teaching VT the full bilateral denervation technique at the University of British Columbia. A travel grant for this research was provided by the Faculty of Graduate and Postgraduate Studies at the University of Ottawa. This research is also supported by NSERC PGS-D scholarship to VT and Discovery Grants Program to SFP.
資料
| Name | Company | Catalog Number | Comments |
| MitoTracker Red | Life Technologies | M-7512 | |
| Dimethyl sulfoxide | Sigma Alrdich | D2650 | |
| α-5 primary antibody | Develompental Hybridoma Bank | a5 | |
| zn12 primary antibody | Develompental Hybridoma Bank | zn12 | |
| Alexa Fluor 488 (anti mouse) | Life Technologies | A-11001 | |
| Benzocaine | Sigma Alrdich | E1501 | 4-aminobenzoic acid ethyl ester, ethyl 4-aminobenzoate |
| Vannas spring scissors | Fine Science Tools | 15000-10 | |
| Standard pattern forceps | Fine Science Tools | 11000-12 | straight |
| Standard pattern forceps | Fine Science Tools | 11001-12 | curved |
| Dumont No. 5 forceps | Fine Science Tools | 11252-30 | |
| Tissue retractor | Fine Science Tools | 17009-07 | |
| Paraformaldehyde | Sigma Alrdich | P6148 | |
| Triton X | Sigma Alrdich | X100 | |
| Vectashield with DAPI | Vector Laboratories | H-1200 |
参考文献
- Katoh, F., Kaneko, T. Short-term transformation and long-term replacement of branchial chloride cells in killifish transferred from seawater to freshwater, revealed by morphofunctional observations and a newly established 'time-differential double fluorescent staining' technique. J. Exp. Biol. 206 (22), 4113-4123 (2003).
- Perry, S. F. Relationships between branchial chloride cells and gas transfer in freshwater fish. Comp. Biochem. Physiol. A. Mol. Integr. Physiol. 119 (1), 9-16 (1998).
- Sollid, J., Weber, R. E., Nilsson, G. E. Temperature alters the respiratory surface area of crucian carp Carassius carassius and goldfish Carassius auratus. J. Exp. Biol. 208 (6), 1109-1116 (2005).
- Mitrovic, D., Perry, S. F. The effects of thermally induced gill remodeling on ionocyte distribution and branchial chloride fluxes in goldfish (Carassius auratus). J. Exp. Biol. 212 (6), 843-852 (2009).
- Tzaneva, V., Vadeboncoeur, C., Ting, J., Perry, S. F. Effects of hypoxia-induced gill remodelling on the innervation and distribution of ionocytes in the gill of goldfish, Carassius auratus. J. Comp. Neurol. 522 (1), 118-130 (2014).
- Greco, A. M., Fenwick, J. C., Perry, S. F. The effects of soft-water acclimation on gill structure in the rainbow trout Oncorhynchus mykiss. Cell Tissue Res. 285 (1), 75-82 (1996).
- Jonz, M. G., Nurse, C. A. Epithelial mitochondria-rich cells and associated innervation in adult and developing zebrafish. J. Comp. Neurol. 497 (5), 817-832 (2006).
- Laurent, P., Hebibi, N. Gill morphometry and fish osmoregulation. Can. J. Zool. 67 (12), 3055-3063 (1989).
- Perry, S. F., Laurent, P. Adaptational responses of rainbow trout to lowered external NaCL concentration: contribution of the branchial chloride cell. J. Exp. Biol. 147, 147-168 (1989).
- Evans, D. H., Piermarini, P. M., Choe, K. P. The multifunctional fish gill: dominant site of gas exchange, osmoregulation, acid-base regulation, and excretion of nitrogenous waste. Physiol. Rev. 85 (1), 97-177 (2005).
- Hwang, P. P., Lee, T. H., Lin, L. Y. Ion regulation in fish gills: recent progress in the cellular and molecular mechanisms. Am. J. Physiol. Regul. Integr. Comp. Physiol. 301 (1), R28-R47 (2011).
- Krogh, A. The Active Uptake of Ions into Cells and Organisms. Proc. Natl. Acad. Sci. U. S. A. 25 (6), 275-277 (1939).
- Nilsson, G. E., Randall, D. J., Hoar, W. S. Innervation and pharmacology of the gills. Fish Physiology. , 185-272 (1984).
- Sundin, L., Nilsson, S. Branchial innervation. J. Exp. Zool. 293 (3), 232-248 (2002).
- Dymowska, A. K., Hwang, P. P., Goss, G. G. Structure and function of ionocytes in the freshwater fish gill). Respir. Physiol. Neurobiol. 184 (3), 282-292 (2012).
- Witters, H., Berckmans, P., Vangenechten, C. Immunolocalization of Na+/K+-ATPase in the gill epithelium of rainbow trout, Oncorhynchus mykiss. Cell Tissue Res. 283 (3), 461-468 (1996).
- Bindon, S., Fenwick, J. C., Perry, S. F. Branchial chloride cell proliferation in the rainbow trout, Onchorhynchus mykiss: implications for gas transfer. Can. J. Zool. 72 (8), 1395-1402 (1994).
- Bradshaw, J. C., Kumai, Y., Perry, S. F. The effects of gill remodeling on transepithelial sodium fluxes and the distribution of presumptive sodium-transporting ionocytes in goldfish (Carassius auratus). J. Comp. Physiol. B. 182 (3), 351-366 (2012).
- Chou, M. Y., et al. Effects of hypothermia on gene expression in zebrafish gills: upregulation in differentiation and function of ionocytes as compensatory responses. J. Exp. Biol. 211 (19), 3077-3084 (2008).
- Kaneko, T., Katoh, F. Functional morphology of chloride cells in killifish Fundulus heteroclitus, a euryhaline teleost with seawater preference. Fisheries Science. 70 (5), 723-733 (2004).
- Ouattara, N., et al. Changes in gill ionocyte morphology and function following transfer from fresh to hypersaline waters in the tilapia Sarotherodon melanotheron. Aquaculture. 290 (1-2), 155-164 (2009).
- Bindon, S., Gilmour, K., Fenwick, J., Perry, S. The effects of branchial chloride cell proliferation on respiratory function in the rainbow trout, Oncorhynchusmykiss. J. Exp. Biol. 197 (1), 47-63 (1994).
- Dunel-Erb, S., Bailly, Y., Laurent, P. Pattern of gill innervation in two teleosts, the perch and the trout. Can. J. Zool. 71, 18-25 (1993).
- Jonz, M. G., Nurse, C. A. New developments on gill innervation: insights from a model vertebrate. J. Exp. Biol. 211 (15), 2371-2378 (2008).
- Jonz, M. G., Zaccone, G. Nervous control of the gills. Acta Histochem. 111 (3), 207-216 (2009).
- Milsom, W. K., Reid, S. G., Rantin, F. T., Sundin, L. Extrabranchial chemoreceptors involved in respiratory reflexes in the neotropical fish, Colossoma macropomum (the tambaqui). J. Exp. Biol. 205 (12), 1765-1774 (2002).
- Hwang, P., Lee, T. New insights into fish ion regulation and mitochondrion-rich cells. Comp Biochem Physiol A Mol Integr Physiol. 148 (3), 479-497 (2007).
転載および許可
このJoVE論文のテキスト又は図を再利用するための許可を申請します
許可を申請さらに記事を探す
This article has been published
Video Coming Soon
Copyright © 2023 MyJoVE Corporation. All rights reserved